
Abstract
Pancreatic cancer is a highly lethal disease due to its rapid dissemination and resistance to conventional chemotherapy. MicroRNAs (miRNAs) are emerging as novel regulators of chemoresistance, which modulate the expression of drug resistance-related genes. MiRNA-221 has been reported to be associated with chemoresistance in various types of cancer. But the detailed molecular mechanism about miR-221-3p regulating 5-fluorouracil (5-FU) resistance in human pancreatic cancer remains to be clarified. In this study, we investigated the association between miR-221-3p expression and 5-FU sensitivity. Studies on pancreatic cancer cell lines suggested an increased 5-FU resistance with miR-221-3p over-expression. In addition, the results indicated that miR-221-3p down-regulated RB1 expression by directly binding to its 3′-UTR and therefore caused increased several aspects of pancreatic cancer pathogenesis, including proliferation, migration, invasion, and epithelial-mesenchymal transition (EMT). Collectively, our findings revealed the important role of miR-221-3p in promoting 5-FU resistance of pancreatic cancer cells and provided a potential therapeutic target for pancreatic cancer.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Pancreatic cancer has the poorest survival among all human cancers, mostly due to its difficulty for early diagnosis, local aggression, and rapid progression [1]. To date, surgery is the primary treatment for long-term survival of pancreatic cancer patients. However, only approximately 10–20 % of patients with pancreatic cancer are suitable for surgery at the time of diagnosis because of tumor invasion and metastasis [2]. Therefore, chemotherapy becomes one of the important treatments for pancreatic cancer patients. A frequently used chemotherapeutic drug is 5-FU, which can effectively improve quality of life and increase the long-term survival of pancreatic cancer patients. Nevertheless, widespread 5-FU drug resistance leads to the poor prognosis of pancreatic cancer [3, 4]. Therefore, there is a dire need for understanding the drug resistance mechanisms and discovering potential new targets for improving therapeutic efficacy of pancreatic cancer patients.
It has been found that multiple regulators are involved in cancer cell chemoresistance, including aberrant gene expression, mutations, deregulation of key signaling pathways, epithelial-mesenchymal transition (EMT), and the presence of highly resistant cells and stem cells [5]. Each of those mechanisms contributes to drug resistance of pancreatic cancer from different aspects and suggests different therapeutic targets.
Studies show that miRNAs play critical roles in regulating gene expression by affecting mRNA stability, translation, and the post-transcriptionally regulation of target genes [6–9]. The mature miRNAs imperfectly bind to the 3′ untranslated region (3′UTR) of target mRNA, causing either degradation of mRNA or inhibition of their translation to functional proteins [10, 11]. A series of recent studies have shown that miRNAs are associated with a wide range of biological processes, such as embryonic development, organ formation, and cell proliferation and apoptosis [12]. Besides, miRNAs play important roles in tumorigenesis and chemoresistance [13–15]. MiR-221 are reported to be involved in many epithelial cancers, such as glioma, prostate carcinoma, hepatocellular cancer, lung cancer, and breast cancer [1, 16–22]. Study also shown that miR-221 is significantly up-regulated in pancreatic cancer cell lines [23]. However, the detailed function, the mechanism, and the correlation between miR-221-3p and 5-FU resistance in pancreatic cancer are still poorly understood.
Moreover, emerging evidence indicated that the malignant behavior of pancreatic cancer is partly due to its intrinsic and extrinsic drug resistance characteristics, which are associated with the acquisition of the EMT phenotype [24, 25]. The EMT is a process that epithelial cells with a cobblestone phenotype acquire mesenchymal cell characteristics with a spindle-shaped fibroblast-like morphology. This process involves the disassembly of cell-cell junctions, the down-regulation of epithelial cell phenotype markers (E-cadherin, zonula occludens-1), as well as the translocation of β-catenin from the cellular membrane to the nucleus, reorganization of the actin cytoskeleton, and up-regulation of mesenchymal cell phenotype markers (vimentin, fibronectin, and N-cadherin) [26]. And the adhesive capacity of mesenchymal phenotypic cells is reduced in EMT, which leads to increased cell migration and invasion, resulting in tumor aggressiveness [27]. Moreover, accumulating evidence indicates the correlation between EMT and drug resistance [28–30]. However, whether the change of EMT markers contributes to pancreatic cancer 5-FU resistance was less studied.
In the present study, we identified miR-221-3p as a key regulator of 5-FU resistance in human pancreatic cancer cells by targeting RB1 and facilitating several aspects of pancreatic cancer pathogenesis, including proliferation, invasion, metastasis, drug resistance, and EMT. Furthermore, the expression of miRNAs and the specific targets can predict the effectiveness of the chemotherapy drugs for particular patients, which also implies miR-221-3p can be developed as a new prognostic marker for chemotherapy of pancreatic cancer.
Materials and methods
Cell lines and cell culture
The stable human pancreatic cancer PANC-1 cell line and PATU8988 cell line were obtained from the Department of General Surgery, Peking Union Medical College (Beijing, China), and 293TN cells were obtained from American Type Culture Collection. The 293TN cell line and PANC-1 cell line were cultured in Dulbecco’s modified Eagle medium (Invitrogen, USA), and PATU8988 cell line was cultured in RPMI-1640 medium (PAA). All media were supplemented with 10 % fetal bovine serum (FBS; from PAA Laboratories, USA), 100 U/ml penicillin, and 100 mg/ml streptomycin. All of the cells were maintained at 37 °C in a humidified atmosphere consisting of 5 % CO2.
Construction and transduction of recombinant lentivirus
Human miR-221-3p expression construct was generated by insertion of the coding sequence (CDS) of miR-221-3p into the downstream of CMV promoter in pMIRNA1 to generate pMIRNA1-miR-221-3p. The pMIRNA1 plasmid and pPACKH1 lenti-vector packaging kit were purchased from SBI (System Biosciences, USA). Recombinant lentiviruses were produced by transient transfection of HEK293TN cells, along with package vectors, using Lipofectamine 2000 (Invitrogen). Viral packaging was performed according to the manufacturer’s instructions. After transfection for 48 h, virus particles (lenti-miR221-3p and lenti-GFP) were harvested and concentrated using PEG-it Virus Precipitation Solution (SBI), and virus titer was determined in 293TN cells using global ultrarapid lentiviral titer kit (SBI). For gene transduction into PATU8988 and PANC-1 cells, the recombinant virus were added to the culture medium of the cells as MOI = 3~5.
Construction and transfection of expression plasmids
The coding sequence of RB1 were obtained and inserted into pmiR reporter plasmid. A 500-bp DNA fragment flanking pre-miR-221-3p was inserted into PcDNA3.1 (+) plasmid. The plasmid inserted with complementary sequences of miR-221-3p was used as positive control. The recombinant expression plasmids were transfected into PATU8988 and PANC-1 cells using Lipofectamine 2000 (Invitrogen) transfection reagent according to the protocols. The transfection efficiency was confirmed by RT-PCR.
RNA isolation and quantitative real-time PCR
Total RNA was isolated from pancreatic cancer PANC-1 cell line and PATU8988 cell line using Trizol reagent (Invitrogen, CA, USA) according to the manufacturer’s instructions. RNA was quantified by absorbance at 260 nm, and cDNA was synthesized by M-MLV reverse transcriptase (Invitrogen) from 2 μg of total RNA. Oligo (dT) 18 was used as the RT primers for reverse transcription of mRNA. A stem-loop RT primer was used for the reverse transcription of miRNA. For mRNAs, quantitative real-time PCR was carried out in a Bio-Rad CFX96 real-time PCR System (Bio-Rad, CA, USA) using SYBR Premix Ex Taq kit (Takara, Dalian, China) according to the manufacturer’ s instructions. The PCR conditions were as follows: 95 °C for 1 min, followed by 40 cycles of 95 °C for 30 s, 60 °C for 30 s, 70 °C for 1 min. For mRNAs, the data were normalized using the endogenous GAPDH control. For miRNAs, U6 snRNA was used as the endogenous control.
In vitro cytotoxicity tests
Cell viability (PATU8988 and PANC-1 cell) was identified by 5-FU and gemcitabine assay. 5-Fluorouracil and gemcitabine were purchased from Eli Lilly and Company (USA). Briefly, PATU8988 cells were plated (8 × 103 cells/well), and PANC-1 cells were plated (1 × 104 cells/well) in 96-well plates and incubated overnight to allow cell attachment. Then, 5-FU (fourfold serial dilution, from 1 × 103 to 9.54 × 10−4 μg/ml)/gemcitabine (twofold serial dilution, from 1.024 × 104 to 1.25 ng/ml) was added and incubated for 72 h.
Cell proliferation assay
2.5 × 103 cells of both PATU8988 and PANC-1 cells were split into 96-well plates. The cells were treated with or without 10 μl CCK-8 (Dojindo, Japan) for 2 h, followed by a cell proliferation assay. Proliferation rates were determined at 0, 24, 48, 72, and 96 h after seeding. All experiments were performed at least three times.
Wound healing assay
A wound healing assay was performed to examine the capacity of cell migration. After the PATU8988 and PANC-1 cells grew to 90–95 % confluence in six-well plates, a single scratch wound was generated with a 200-μl disposable pipette tip. The scratch wounds were photographed at 0, 12, 24, and 36 h, with a Nikon inverted microscope with an attached digital camera, and their widths were quantitated with the ImageJ software. The data were plotted as the percentage of wound opening, setting the initial scratch width as 100 %. The results are presented as the means ± SD of triplicate measurements per condition in three independent experiments.
Cell invasion assay
The invasive behaviors of the cells were tested using a Matrigel transmembrane invasion assay. Transwell chambers (Millipore, 8 mm pore size) were coated with Matrigel (15 mg/filter). Cells (1 × 104 cells/well) were suspended in serum-free medium and seeded into the upper chamber of each insert, while the lower chamber contained medium with 10 % FBS. Cells were allowed to invade across the Matrigel-coated membrane for 24 h at 37 °C in 5 % CO2. After 24 h of incubation, cells were removed from the upper surface of the filter by scraping with a cotton swab. The invaded cells that adhered to the bottom of the membrane were fixed with methanol and stained with the 0.1 % crystal violet and counted. All experiments were performed at least three times. The number of cells that penetrated the membrane was determined by counting the mean cell number of five randomly selected high-power fields.
Dual luciferase reporter assay
For miRNA target analysis, the 293TN cells were co-transfected with 0.4 μg of the reporter construct, 0.02 μg of pRL-TK-Renilla luciferase plasmid (Promega, Madison, WI, USA), and miR-221-3p over-expression construct or control. Cells were harvested 48 h post-transfection and assayed with Dual Luciferase Assay (Promega) according to manufacturer’s instructions. All transfection assays were carried out in triplicates.
Western blotting
For protein isolation from PATU8988 and PANC-1 cells, lysis buffer supplemented with proteinase inhibitor cocktail was used. The protein concentration was determined using the BCA protein assay kit (Bio-Rad, Italy), and cell lysates were subjected to SDS/PAGE (10 % separation gel) and transferred onto a PVDF membrane (Millipore, USA). The following antibodies were used for Western blot: GAPDH Rabbit mAb antibody (Santa Cruz Biotechnology) and RB1 Rabbit mAb antibody (Cell Signal Technology, Danvers, MA, USA). We evaluated the relative expression of protein by Gel-pro Analyzer.
Statistical analysis
Statistical analyses were performed using IBM SPSS Statistics (version 22, IBM SPSS Inc., Chicago, IL) or the Graph Pad Prism 5.0 software package (GraphPad Software, Inc., USA). Survival curves were generated by Kaplan-Meier analysis and tested for significance using the Mantel-Cox log rank test using TCGA pancreatic adenocarcinoma (PAAD) miRNA expression by RNA seq. The Spearman correlation analysis was examined by Spearman p test using GEO database (GSE41368 and GSE41369). Student’s t test (two-tailed) was performed to analyze data from the experiments in three triplicates. All data are presented as the means ± SD. p values <0.05 were considered significant, as indicated by the asterisks (*p < 0.05, **p < 0.01, ***p < 0.001).
Results
Establishment of 5-FU-resistant cell lines and identification of differentially expressed miRNAs
In order to systematically screen the miRNAs involved in the induction of 5-FU drug resistance, we created 5-FU-resistant pancreatic cancer cell lines and examined its response to 5-FU using MTT assay. IC50 were 2.456 μg/ml for PATU8988 cells and 51.83 μg/ml for PATU8988/5-FU cells (Fig. 1a). The resistance of PATU8988/5-FU cells to 5-FU was estimated to be approximately 25 times higher than that of the parental cells (Fig. 1b). Further, we investigated the miRNA expression profile of PATU8988/5-FU and PATU8988 cells through microarray to find the differentially expressed miRNAs. The results showed that there were many miRNAs differentially expressed in PATU8988/5-FU cells compared with PATU8988 cells. MiR-221-3p was significantly up-regulated in the PATU8988/5-FU cells compared with that in PATU8988 cells with a fold change of 2.8 by RT-PCR analysis (***p < 0.0001, Fig. 1c). These results led us to speculate that up-regulation of miR-221-3p may be associated with 5-FU resistance of pancreatic cancer cells. The resistant cells were continuously maintained in culture medium containing 5-FU for the following study.

Up-regulation of miR-221-3p in PATU8988/5-FU cells and prognostic value of miR-221-3p in pancreatic cancer patients. a Representative curves of growth-inhibitory effects in 5-FU-resistant PATU8988/5-FU and PATU8988 cells. IC50, half-maximal inhibitory concentration. b Resistant index of PATU8988 and 5-FU-resistant PATU8988. c Real-time PCR analysis of miR-221-3p expression between 5-FU-resistant PATU8988/5-FU cells and its parental PATU8988 cells. d The relative expression of miR-221-3p in pancreatic cancer tissues compared to their paired normal tissues (n = 9) with the data from GEO database (GSE41369) (**p < 0.01). e Kaplan-Meier overall survival analysis of miR-221-3p expression on the pancreatic cancer patients (n = 179) from TCGA database. f Kaplan-Meier relapse-free survival analysis of miR-221-3p expression on the pancreatic cancer patients (n = 179) from TCGA database. p value was calculated by log-rank test. All data are presented as the means ± SD. Student’s t test (two-tailed) was performed to analyze data from the experiments in triplicate. p values <0.05 were considered significant, as indicated by the asterisks (**p < 0.01, ***p < 0.001)
Association of miR-221-3p expression level with the clinic pathological significance in pancreatic cancer
To investigate the biological role of miR-221-3p in human pancreatic cancer, we assessed the prognostic value of miR-221-3p with the data from GEO database (GSE41369). We found that miR-221-3p was significantly up-regulated in pancreatic cancer tissues compared with their paired normal tissues (p = 0.0039, Fig. 1d). We also evaluated the clinical value of miR-221-3p in 179 pancreatic cancer patients in TCGA database. According to the TCGA pancreatic adenocarcinoma (PAAD) miRNA expression by RNA seq, we found that lower expression of miR-221-3p was correlated with longer overall survival, while shorter overall survival was associated with higher expression of miR-221-3p (Fig. 1e, p = 0.028). The similar result was found in the correlation analysis of expression level of miR-221-3p with the relapse-free survival (Fig. 1f, p = 0.035). These results suggested that higher expression of miR-221-3p might increase 5-FU resistance and then reduce therapeutic effect.
Over-expression of miR-221-3p induces resistance to 5-FU in pancreatic cancer cells
To determine the contribution of miR-221-3p to 5-FU resistance in pancreatic cells, PATU8988 and PANC-1 cells were infected with lenti_miR-221-3p and lenti_GFP (as a control), respectively, and treated with 5-FU. MiR-221-3p was over-expressed successfully in PATU8988 and PANC-1 cells as determined by quantitative RT-PCR (Fig. 2a, d). As a result, miR-221-3p decreased the sensitivity to 5-FU in both PANC-1 and PATU8988 cells compared with the control. IC50 was evaluated as 4.727 μg/ml for miR-221-3p over-expression group, 1.804 μg/ml for the control in PANC-1 cells, and 0.6982 vs. 0.1784 μg/ml in PATU8988 cells (Fig. 2b, e). Thus, the IC50 values of miR-221-3p over-expressed cells were approximately three times higher than that of the control cells. Gemcitabine was also one of the most popular chemotherapy drugs used in pancreatic cancer treatment, so we also detected the effect of miR-221-3p on regulating gemcitabine resistance. We found that IC50 of gemcitabine was evaluated as 143.4 ng/ml for miR-221-3p over-expression group, 22.09 ng/ml for the control in PANC-1 cells, and 149.8 vs. 32.33 ng/ml in PATU8988 cells (Fig. 2c, f). Thus, the IC50 of gemcitabine in miR-221-3p over-expressed cells were both higher than that of the control cells. According to these data, the expansion of miR-221-3p might have an important role for the acquisition of 5-FU and gemcitabine resistance.

Over-expression of miR-221-3p induces drug resistance in pancreatic cancer cells. a Real-time PCR analysis of miR-221-3p expression in PANC-1 cells after lentivirus infection with lenti_GFP control and lenti_miR-221-3p. b Representative curves of growth-inhibitory effects of 5-FU on PANC-1 cells which were infected with lenti_miR-221-3p and lenti_GFP. c Representative curves of growth-inhibitory effects of gemcitabine on PANC-1 cells which were infected with lenti_miR-221-3p and lenti_GFP. d Real-time PCR analysis of the expression of miR-221-3p in PATU8988 cells after lentivirus infection with lenti_GFP control and lenti_miR-221-3p. e Representative curves of growth-inhibitory effects of 5-FU on PATU8988 cells after infecting with lenti_miR-221-3p and lenti_GFP. f Representative curves of growth-inhibitory effects of gemcitabine on PATU8988cells which were infected with lenti_miR-221-3p and lenti_GFP. All data are presented as the means ± SD. Student’s t test (two-tailed) was performed to analyze data from the experiments in triplicate. p values <0.05 were considered significant, as indicated by the asterisks (*p < 0.05, **p < 0.01, ***p < 0.001)
Over-expression of miR-221-3p promotes proliferation and migration of pancreatic cancer cells
To explore whether miR-221-3p can regulate the aggressive characteristics of pancreatic cancer cells, we measured the proliferation and migration in PANC-l cells and PATU8988 cells with miR-221-3p over-expression. The proliferation rate of pancreatic cells was measured at 0, 24, 48, 72 h and 96 h, which showed that miR-221-3p promoted the growth of pancreatic cancer cell lines PANC-1 and PATU8988(Fig. 3a, b). To further validate whether pancreatic cancer cells with miR-221-3p over-expression have enhanced motility, we detected cell migration using wound-healing assay. Our migration results showed that over-expression of miR-221-3p increased cell migration compared with their control cells in PANC-l cells and PATU8988 cells. We found that miR-221-3p significantly promoted the migration rate during the closure of a wound created in a confluent monolayer in PANC-1 cell and PATU8988 cell (Fig. 3c, d). The relative ratio of PATU8988 cell wound opening per field at 0, 12, 24, and 36 h is shown (Fig. 3e, f). Taken together, we demonstrated that miR-221-3p promotes proliferation and migration of pancreatic cancer cells.

Over-expression of miR-221-3p promotes pancreatic cancer cell proliferation and migration. a Proliferation assays by cck-8 at 0, 24, 48, 72 h and 96 h after infecting with lenti_miR-221-3p or lenti_GFP in PANC-1 cells. b Proliferation assays by cck-8 at 0, 24, 48, 72 h and 96 h after infecting with lenti_miR-221-3p or lenti_GFP in PATU8988 cell. c PANC-1 cells were infected with lenti_miR-221-3p or lenti_GFP for 36 h and wounds were made. d PATU8988 cells were infected with lenti_miR-221-3p or lenti_GFP for 36 h and wounds were made. Bar, 100 μM. e The relative ratio of PANC-1 cell wound opening per field at 0, 12, 24, and 36 h. f The relative ratio of PATU8988 cell wound opening per field at 0, 12, 24, and 36 h. All data are presented as the means ± SD. Student’s t test (two-tailed) was performed to analyze data from the experiments in triplicate. p values <0.05 were considered significant, as indicated by the asterisks (*p < 0.05, **p < 0.01, ***p < 0.001)
Over-expression of miR-221-3p promotes invasion of pancreatic cancer cells
To further confirm the function of miR-221-3p in pancreatic cancer cells, we investigated the effects of miR-221-3p on cell invasion, which is an important function for malignant progression and metastases, using a Matrigel invasion assay. We used PANC-1 and PATU8988 cells with transwell assays for these experiments. PANC-1 cells exhibited significantly increased cell invasion (3.5-fold) compared with the control cells at 24 h after seeding the cells transfected with the miR-221-3p expression (***p < 0.0001, Fig. 4a). PATU8988 cells with miR-221-3p over-expression also exhibited markedly increased cell invasion (4.6-fold) at 24 h after transfection compared with the control cells (***p < 0.0001, Fig. 4b). These findings revealed that miR-221-3p was critically involved in invasion characteristics in pancreatic cancer cells. Altogether, our results demonstrated that miR-221-3p plays an important role in modulating biological functions of 5-FU resistance in human pancreatic cancer cells. Over-expression of miR-221-3p can promote cell proliferation, migration, and invasion of pancreatic cancer cells.

Over-expression of miR-221-3p promotes pancreatic cancer cell invasion and EMT. a PANC-1 cells were infected with lenti_miR-221-3p or lenti_GFP for 24 h, and transwell invasion assay was performed. The representative migrated cells per field are shown. b PATU8988 cells were infected with lenti_miR-221-3p or lenti_GFP for 24 h, and transwell invasion assay was performed. The representative migrated cells per field are shown. We selected three visual fields for calculating. c, d Real-time PCR assay was conducted to detect the expression of “EMT” markers in PANC-1 cells and PATU8988 cells. Left panel: the expression of β-cadherin, E-cadherin in PANC-1 and PATU8988 cells. Right panel: the expression of Fibronectin, N-cadherin, vimentin, ZEB1, ZEB2, and Snail in PANC-1 and PATU8988 cells. All data are presented as the means ± SD. Student’s t test (two-tailed) was performed to analyze data from the experiments in triplicate. p values <0.05 were considered significant, as indicated by the asterisks (*p < 0.05, **p < 0.01, ***p < 0.001)
Over-expression of miR-221-3p promotes EMT molecular marker changes in pancreatic cancer cells
Recent studies have shown that epithelial-mesenchymal transition (EMT) is associated with drug resistance and cancer cell metastasis [25, 31, 24]. Thus, an understanding of the molecular biology of EMT in pancreatic cancer may provide insights into the mechanisms of tumor proliferation, invasion, and metastatic progression and facilitates the development of alternative therapeutic approaches to improve the treatment outcomes for patients suffering from pancreatic cancer [32]. To investigate whether miR-221-3p promotes EMT to facilitate drug resistance of pancreatic cancer cells, we measured the mRNA levels of EMT markers using RT-PCR in PANC-1 and PATU8988 cells. We found that epithelial molecule β-cadherin and E-cadherin mRNAs were down-regulated, while the mRNA levels of mesenchymal markers including Fibronectin, ZEB1, and ZEB2 were up-regulated in PANC-1 cells with over-expression of miR-221-3p (Fig. 4c). And the similar results were found in PATU8988 cells (Fig. 4d). These results further suggested that miR-221-3p promoted pancreatic cancer cells to acquire a mesenchymal phenotype to facilitate drug resistance.
MiR-221-3p targets RB1 in pancreatic cancer cells
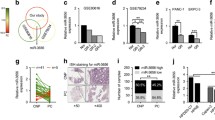
Emerging evidence suggests that the expression of genes is fundamental to the acquisition of the EMT phenotype, and the aggressiveness of tumor cell is regulated by miRNAs which lead to either translational repression or the degradation of target mRNAs [33, 34]. The putative targets of miR-221-3p were predicted using target prediction programs, TargetScan, PicTar, and miRanda. Our analysis revealed that RB1 was a potential target of miR-221-3p. The 3′-UTR of RB1 mRNA contains complementary sites for the seed region of miR-221-3p (Fig. 5a). To determine whether RB1 is a direct target of miR-221-3p, wild-type 3′-UTR binding site was cloned into the downstream of firefly luciferase-coding region in pmiR luciferase reporter vector. The constructs were co-transfected with miR-221-3p over-expression construct or control into 293TN cells. The relative luciferase activity was reduced to 40 % in miR-221-3p over-expression group (Fig. 5b). To further determine whether RB1 was a direct target of miR-221-3p, we over-expressed miR-221-3p in the two pancreatic cancer cell lines and detected RB1 protein levels. The protein levels of RB1 were substantially decreased after ectopic over-expression of miR-221-3p in pancreatic cancer cell lines PANC-1 and PATU8988 (Fig. 5c) as evidenced by western blot assays (Fig. 5d). Taken together, these findings indicate that RB1 can be negatively regulated by miR-221-3p, and miR-221-3p modulates 5-FU resistance in human pancreatic cancer cells by targeting RB1.

RB1 is verified as a direct target of miR-221-3p in pancreatic cancer cells lines. a The prediction of the binding between miR-221-3p and RB1 by Pictar. b Relative luciferase activity of the indicated RB1 reporter constructs in 293TN cells. Error bars presented standard deviation obtained from three independent experiments. c Real-time PCR analysis of the expression of miR-221-3p in PANC-1 and PATU8988 cells after lentivirus infection with lenti_GFP control and lenti_miR-221-3p. d Western blot analysis of RB1 protein in PANC-1 and PATU8988 cells after miR-221-3p over-expression. e The inverse correlation of RB1 and miR-221 expression levels was examined by Spearman correlation analysis using GEO database (GSE41368 and GSE41369) (R 2 = 0.0955, p = 0.032). All data are presented as the means ± SD. Student’s t test (two-tailed) was performed to analyze data from the experiments in triplicate. p values <0.05 were considered significant, as indicated by the asterisks (*p < 0.05, **p < 0.01, ***p < 0.001)
Then, the Spearman correlation analysis using GEO database (GSE41368 and GSE41369) was applied to compare the relative expression levels of RB1 and miR-221-3p in the pancreatic cancer specimens. We obtained a statistically significant inverse correlation (R 2 = 0.0955, p = 0.032) in pancreatic cancer tissues (Fig. 5e). These data suggested that miR-221-3p expression was inversely correlated with RB1, and RB1 was a direct target of miR-221-3p in pancreatic cancer.
Discussion
Pancreatic cancer is the fourth leading cause of cancer death in the USA. In addition, 80–85 % of patients diagnosed with pancreatic cancer are in the terminal period. Therefore, chemotherapy becomes one of the important treatments for patients with pancreatic cancer. Recently, a lot of researches focused on the gemcitabine drug resistance in pancreatic cancer; however, 5-FU as the earliest pancreatic cancer chemotherapy drug which is frequently used in the treatment of pancreatic cancer was less studied. As 5-FU primary and secondary resistance phenomenon is widespread and becomes one of the important factors affecting the effect of chemotherapy, so whether or not we can predict the effectiveness of the chemotherapy drugs for particular patients, make individualized chemotherapy come true, and reverse chemotherapy drug resistance is the core issue of the current research on pancreatic cancer chemotherapy.
Many studies have found that various factors are involved in the chemosensitivity of cancer cell, and miRNAs have been extensively studied in recent years. Presently, about 28,645 human miRNAs have been annotated in the miRBase database [35], and many of them are correlated with cancer chemoresistance [36, 37]. MiRNAs are a group of regulators which modulated target gene expression and involved in a wide range of biological processes [38–41]. In addition, numerous studies on miRNAs have indicated that miRNAs can function as oncogenes or tumor suppressor genes and participate in the development and progression of different tumors [42]. In our study, we created 5-FU-resistant pancreatic cancer cell lines and analyzed the expression profile of miRNAs using miRNA microarray, and we found that miR-221-3p was significantly up-regulated in the PATU8988/5-FU cells compared with that in the PATU8988 cells. Studies show that miR-221-3p plays critical roles in varieties of cell types, such as hematopoietic cells, endothelial cells, and tumor cells [43–46]. A series of reports pointed out that miR-221-3p is involved in the malignant behavior of the cancer, including proliferation, apoptosis, invasion, metastasis, and chemoresistance [47–49] and plays as an oncogenic microRNA in prostate cancer [50–52]. Similar with the precious study, we demonstrated that miR-221-3p played an important role in regulating 5-FU resistance in pancreatic cancer.
Recently, emerging evidence show that miR-221-3p has been found to play important roles in the acquisition of the EMT phenotype in cancer cells, like miR-221/221 targets ADIPOR1 to promote the EMT in breast cancer [53]. Moreover, it has become increasingly clear that the EMT plays an important role in the progression of cancer and is also responsible for the resistance of cancer cells to conventional chemotherapeutics. Such as, EMT may account for the movement of cells from a primary tumor into the circulation, suggesting that EMT is important in forming initial metastases [54]. Our current study demonstrated that miR-221-3p promoted cell proliferation, migration, and invasion and resulted in the acquisition of the EMT phenotype by up-regulation of mesenchymal cell markers (ZEB1, ZEB2, and fibronectin) and down-regulation of an epithelial cell marker (β-cadherin, E-cadherin) in both PANC-1 cells and PATU8988 cells. Thus, miR-221-3p may promote the acquisition of the EMT phenotype, which will result in drug resistance in the treatment of pancreatic cancer disease.
Further investigation into the mechanism revealed that miR-221-3p modulates 5-FU resistance in human pancreatic cancer cells by targeting RB1. As we know, RB1 is a tumor suppressor protein that is dysfunctional in several major cancers [55]. Precious studies show that RB1 can prevent excessive cell growth by inhibiting cell cycle progression [56], also participates in the regulation of apoptosis, which partly accounts for cancer development and chemotherapy resistance [57]. In our present study, we demonstrate that miR-221-3p directly targeted the 3′-UTR of RB1 mRNA and repressed its expression, lead to drug resistance in pancreatic cancer.
Taken together, miR-221-3p has been considered to be important in human pancreatic cancer. In the current study, a growing body of literature strongly suggests that over-expression of miR-221-3p can promote the processes of cell proliferation, migration, invasion, EMT, and promote resistance to 5-FU in pancreatic cancer cells. We have further demonstrated that miR-221-3p may represent a novel therapeutic by targeting RB1, and which is likely to have a significant therapeutic impact on human pancreatic cancer.
References
Siegel R, Ma J, Zou Z, Jemal A. Cancer statistics, 2014. CA Cancer J Clin. 2014;64(1):9–29. doi:10.3322/caac.21208.
Cartwright T, Richards DA, Boehm KA. Cancer of the pancreas: are we making progress? A review of studies in the US Oncology Research Network. Cancer Control. 2008;15(4):308–13.
Li W, Ma Q, Liu J, Han L, Ma G, Liu H, et al. Hyperglycemia as a mechanism of pancreatic cancer metastasis. Front Biosci (Landmark Ed). 2012;17:1761–74.
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–74. doi:10.1016/j.cell.2011.02.013.
Long J, Zhang Y, Yu X, Yang J, LeBrun DG, Chen C, et al. Overcoming drug resistance in pancreatic cancer. Expert Opin Ther Targets. 2011;15(7):817–28. doi:10.1517/14728222.2011.566216.
Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–97.
Chen K, Rajewsky N. The evolution of gene regulation by transcription factors and microRNAs. Nat Rev Genet. 2007;8(2):93–103.
Lu H, Buchan RJ, Cook SA. MicroRNA-223 regulates Glut4 expression and cardiomyocyte glucose metabolism. Cardiovasc Res. 2010;86(3):410–20. doi:10.1093/cvr/cvq010.
Ma F, Liu X, Li D, Wang P, Li N, Lu L, et al. MicroRNA-466l upregulates IL-10 expression in TLR-triggered macrophages by antagonizing RNA-binding protein tristetraprolin-mediated IL-10 mRNA degradation. J Immunol. 2010;184(11):6053–9. doi:10.4049/jimmunol.0902308.
Liu J, Valencia-Sanchez MA, Hannon GJ, Parker R. MicroRNA-dependent localization of targeted mRNAs to mammalian P-bodies. Nat Cell Biol. 2005;7(7):719–23.
Saxena S, Jonsson ZO, Dutta A. Small RNAs with imperfect match to endogenous mRNA repress translation. Implications for off-target activity of small inhibitory RNA in mammalian cells. J Biol Chem. 2003;278(45):44312–9. doi:10.1074/jbc.M307089200.
Ambros V. MicroRNA pathways in flies and worms: growth, death, fat, stress, and timing. Cell. 2003;113(6):673–6.
Hummel R, Hussey DJ, Haier J. MicroRNAs: predictors and modifiers of chemo- and radiotherapy in different tumour types. Eur J Cancer. 2010;46(2):298–311. doi:10.1016/j.ejca.2009.10.027.
Lima RT, Busacca S, Almeida GM, Gaudino G, Fennell DA, Vasconcelos MH. MicroRNA regulation of core apoptosis pathways in cancer. Eur J Cancer. 2011;47(2):163–74. doi:10.1016/j.ejca.2010.11.005.
Schoof CR, Botelho EL, Izzotti A, Vasques Ldos R. MicroRNAs in cancer treatment and prognosis. Am J Cancer Res. 2012;2(4):414–33.
Garofalo M, Quintavalle C, Di Leva G, Zanca C, Romano G, Taccioli C, et al. MicroRNA signatures of TRAIL resistance in human non-small cell lung cancer. Oncogene. 2008;27(27):3845–55. doi:10.1038/onc.2008.6.
Zhao JJ, Lin J, Yang H, Kong W, He L, Ma X, et al. MicroRNA-221/222 negatively regulates estrogen receptor alpha and is associated with tamoxifen resistance in breast cancer. J Biol Chem. 2008;283(45):31079–86. doi:10.1074/jbc.
Gramantieri L, Fornari F, Ferracin M, Veronese A, Sabbioni S, Calin GA, et al. MicroRNA-221 targets Bmf in hepatocellular carcinoma and correlates with tumor multifocality. Clin Cancer Res. 2009;15(16):5073–81. doi:10.1158/1078-0432.CCR-09-0092.
Lu X, Zhao P, Zhang C, Fu Z, Chen Y, Lu A, et al. Analysis of miR-221 and p27 expression in human gliomas. Mol Med Rep. 2009;2(4):651–6. doi:10.3892/mmr_00000152.
Zhang C, Kang C, You Y, Pu P, Yang W, Zhao P, et al. Co-suppression of miR-221/222 cluster suppresses human glioma cell growth by targeting p27kip1 in vitro and in vivo. Int J Oncol. 2009;34(6):1653–60.
Zhang C, Zhang J, Hao J, Shi Z, Wang Y, Han L, et al. High level of miR-221/222 confers increased cell invasion and poor prognosis in glioma. J Transl Med. 2012;10:119. doi:10.1186/1479-5876-10-119.
Zhang J, Han L, Ge Y, Zhou X, Zhang A, Zhang C, et al. miR-221/222 promote malignant progression of glioma through activation of the Akt pathway. Int J Oncol. 2010;36(4):913–20.
Papaconstantinou IG, Manta A, Gazouli M, Lyberopoulou A, Lykoudis PM, Polymeneas G, et al. Expression of microRNAs in patients with pancreatic cancer and its prognostic significance. Pancreas. 2013;42(1):67–71. doi:10.1097/MPA.0b013e3182592ba7.
Sabbah M, Emami S, Redeuilh G, Julien S, Prevost G, Zimber A, et al. Molecular signature and therapeutic perspective of the epithelial-to-mesenchymal transitions in epithelial cancers. Drug Resist Updat. 2008;11(4–5):123–51. doi:10.1016/j.drup.2008.07.001.
Fuchs BC, Fujii T, Dorfman JD, Goodwin JM, Zhu AX, Lanuti M, et al. Epithelial-to-mesenchymal transition and integrin-linked kinase mediate sensitivity to epidermal growth factor receptor inhibition in human hepatoma cells. Cancer Res. 2008;68(7):2391–9. doi:10.1158/0008-5472.
Christiansen JJ, Rajasekaran AK. Reassessing epithelial to mesenchymal transition as a prerequisite for carcinoma invasion and metastasis. Cancer Res. 2006;66(17):8319–26.
Kong D, Li Y, Wang Z, Sarkar FH. Cancer stem cells and epithelial-to-mesenchymal transition (EMT)-phenotypic cells: are they cousins or twins? Cancers (Basel). 2011;3(1):716–29. doi:10.3390/cancers30100716.
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan A, Zhou AY, et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell. 2008;133(4):704–15. doi:10.1016/j.cell.2008.03.027.
Wang Z, Li Y, Ahmad A, Banerjee S, Azmi AS, Kong D, et al. Pancreatic cancer: understanding and overcoming chemoresistance. Nat Rev Gastroenterol Hepatol. 2011;8(1):27–33. doi:10.1038/nrgastro.2010.188.
Castellanos JA, Merchant NB, Nagathihalli NS. Emerging targets in pancreatic cancer: epithelial-mesenchymal transition and cancer stem cells. Onco Targets Ther. 2013;6:1261–7. doi:10.2147/OTT.
Cheng GZ, Chan J, Wang Q, Zhang W, Sun CD, Wang LH. Twist transcriptionally up-regulates AKT2 in breast cancer cells leading to increased migration, invasion, and resistance to paclitaxel. Cancer Res. 2007;67(5):1979–87 doi:67/5/1979.
Jiang JH, Liu C, Cheng H, Lu Y, Qin Y, YF X, et al. Epithelial-mesenchymal transition in pancreatic cancer: is it a clinically significant factor? Biochim Biophys Acta. 2015;1855(1):43–9. doi:10.1016/j.bbcan.2014.11.004.
Cano A, Nieto MA. Non-coding RNAs take centre stage in epithelial-to-mesenchymal transition. Trends Cell Biol. 2008;18(8):357–9. doi:10.1016/j.tcb.2008.05.005.
Garzon R, Fabbri M, Cimmino A, Calin GA, Croce CM. MicroRNA expression and function in cancer. Trends Mol Med. 2006;12(12):580–7.
Kozomara A, Griffiths-Jones S. miRBase: annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014;42(Database issue):D68–73. doi:10.1093/nar/gkt1181.
Li JH, Luo N, Zhong MZ, Xiao ZQ, Wang JX, Yao XY, et al. Inhibition of microRNA-196a might reverse cisplatin resistance of A549/DDP non-small-cell lung cancer cell line. Tumour Biol. 2015. doi:10.1007/s13277-015-4017-7.
Liu RL, Dong Y, Deng YZ, Wang WJ, Li WD. Tumor suppressor miR-145 reverses drug resistance by directly targeting DNA damage-related gene RAD18 in colorectal cancer. Tumour Biol. 2015;36(7):5011–9. doi:10.1007/s13277-015-3152-5.
Li Z, Yu X, Shen J, Jiang Y. MicroRNA dysregulation in uveal melanoma: a new player enters the game. Oncotarget. 2015;6(7):4562–8.
Bader AG, Brown D, Winkler M. The promise of microRNA replacement therapy. Cancer Res. 2010;70(18):7027–30. doi:10.1158/0008-5472.
Pencheva N, Tavazoie SF. Control of metastatic progression by microRNA regulatory networks. Nat Cell Biol. 2013;15(6):546–54. doi:10.1038/ncb2769.
Sivadas VP, Kannan S. The microRNA networks of TGFbeta signaling in cancer. Tumour Biol. 2014;35(4):2857–69. doi:10.1007/s13277-013-1481-9.
Garzon R, Calin GA, Croce CM. MicroRNAs in cancer. Annu Rev Med. 2009;60:167–79. doi:10.1146/annurev.med.59.053006.104707.
Davis BN, Hilyard AC, Nguyen PH, Lagna G, Hata A. Induction of microRNA-221 by platelet-derived growth factor signaling is critical for modulation of vascular smooth muscle phenotype. J Biol Chem. 2009;284(6):3728–38. doi:10.1074/jbc.M808788200.
Kawaguchi T, Komatsu S, Ichikawa D, Morimura R, Tsujiura M, Konishi H, et al. Clinical impact of circulating miR-221 in plasma of patients with pancreatic cancer. Br J Cancer. 2013;108(2):361–9. doi:10.1038/bjc.2012.546.
le Sage C, Nagel R, Egan DA, Schrier M, Mesman E, Mangiola A, et al. Regulation of the p27(Kip1) tumor suppressor by miR-221 and miR-222 promotes cancer cell proliferation. EMBO J. 2007;26(15):3699–708.
Stinson S, Lackner MR, Adai AT, Yu N, Kim HJ, O'Brien C, et al. TRPS1 targeting by miR-221/222 promotes the epithelial-to-mesenchymal transition in breast cancer. Sci Signal. 2011;4(177):ra41. doi:10.1126/scisignal.
Ciafre SA, Galardi S, Mangiola A, Ferracin M, Liu CG, Sabatino G, et al. Extensive modulation of a set of microRNAs in primary glioblastoma. Biochem Biophys Res Commun. 2005;334(4):1351–8.
Besson A, Gurian-West M, Schmidt A, Hall A, Roberts JM. p27Kip1 modulates cell migration through the regulation of RhoA activation. Genes Dev. 2004;18(8):862–76. doi:10.1101/gad.1185504.
Nikiforova MN, Tseng GC, Steward D, Diorio D, Nikiforov YE. MicroRNA expression profiling of thyroid tumors: biological significance and diagnostic utility. J Clin Endocrinol Metab. 2008;93(5):1600–8. doi:10.1210/jc.2007-2696.
Pang Y, Young CY, Yuan H. MicroRNAs and prostate cancer. Acta Biochim Biophys Sin Shanghai. 2010;42(6):363–9.
Urbich C, Kuehbacher A, Dimmeler S. Role of microRNAs in vascular diseases, inflammation, and angiogenesis. Cardiovasc Res. 2008;79(4):581–8. doi:10.1093/cvr/cvn156.
Vasilatou D, Papageorgiou S, Pappa V, Papageorgiou E, Dervenoulas J. The role of microRNAs in normal and malignant hematopoiesis. Eur J Haematol. 2010;84(1):1–16. doi:10.1111/j.1600-0609.2009.01348.x.
Hwang MS, Yu N, Stinson SY, Yue P, Newman RJ, Allan BB, et al. miR-221/222 targets adiponectin receptor 1 to promote the epithelial-to-mesenchymal transition in breast cancer. PLoS One. 2013;8(6):e66502. doi:10.1371/journal.pone.0066502.
Huber MA, Kraut N, Beug H. Molecular requirements for epithelial-mesenchymal transition during tumor progression. Curr Opin Cell Biol. 2005;17(5):548–58.
Murphree AL, Benedict WF. Retinoblastoma: clues to human oncogenesis. Science. 1984;223(4640):1028–33.
Shao Z, Robbins PD. Differential regulation of E2F and Sp1-mediated transcription by G1 cyclins. Oncogene. 1995;10(2):221–8.
Indovina P, Pentimalli F, Casini N, Vocca I, A. G. .RB1 dual role in proliferation and apoptosis: cell fate control and implications for cancer therapy. Oncotarget. 2015;6(20):17873–90.
Acknowledgments
The authors thank Ming tai Chen and Zheng yi Zhang from IBMS of PUMC for technical assistance. This work was supported by grants from the National Natural Science Foundation of China (31371322, to J.Y.; 91440111, to J.Y.; 31571523, to Y.N.M.)
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflicts of interest
None
Additional information
Lijun Zhao and Dongling Zou contributed equally to this work.
Rights and permissions
About this article

Cite this article
Zhao, L., Zou, D., Wei, X. et al. MiRNA-221-3p desensitizes pancreatic cancer cells to 5-fluorouracil by targeting RB1. Tumor Biol. 37, 16053–16063 (2016). https://doi.org/10.1007/s13277-016-5445-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13277-016-5445-8