
Abstract
Wnt/β-catenin signaling pathway plays a major role in the cancer metastasis. Several microRNAs (miRNAs) are contributed to the inhibition of breast cancer metastasis. Here, we attempted to find novel targets and mechanisms of microRNA-100 (miR-100) in regulating the migration and invasion of breast cancer cells. In this study, we found that miR-100 expression was downregulated in human breast cancer tissues and cell lines. The overexpression of miR-100 inhibited the migration and invasion of MDA-MB-231 breast cancer cells. Inversely, the downregulation of miR-100 increased the migration and invasion of MCF-7 breast cancer cells. Furthermore, FZD-8, a receptor of Wnt/β-catenin signaling pathway, was demonstrated a direct target of miR-100. The overexpression of miR-100 decreased the expression levels not only FZD-8 but also the key components of Wnt/β-catenin pathway, including β-catenin, metalloproteniase-7 (MMP-7), T-cell factor-4 (TCF-4), and lymphoid enhancing factor-1 (LEF-1), and increased the protein expression levels of GSK-3β and p-GSK-3β in MDA-MB-231 cells, and the transfection of miR-100 inhibitor in MCF-7 cells showed the opposite effects. In addition, the expression of miR-100 was negatively correlated with the FZD-8 expression in human breast cancer tissues. Overall, these findings suggest that miR-100 suppresses the migration and invasion of breast cancer cells by targeting FZD-8 and inhibiting Wnt/β-catenin signaling pathway and manipulation of miR-100 may provide a promoting therapeutic strategy for cancer breast treatment.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Breast cancer is the most common cancer in women worldwide, and the vast majority of breast cancer-related deaths are due to metastatic diseases [1]. It is well established that cancer metastasis, a complex and multistep process, is modulated via aberrant cell signaling pathways by their dynamic interactions. Wnt [2], TGF-β [3], and Notch pathways [4] among numerous pathways have been identified to play key roles in breast cancer metastasis.
Wnt signaling pathway is a highly conserved pathway which modulates cell proliferation, differentiation, and cell fate during embryonic development [5]. Wnt/β-catenin pathway is a critical and widely studied Wnt pathway. When Wnt ligands bind to a seven-pass transmembrane Frizzled (Fz) receptor and its co-receptor, low-density lipoprotein receptor-related protein 6 (LRP6) or its close relative LRP5, the Wnt/β-catenin pathway is activated. The LRP6 phosphorylation and activation and the recruitment of the scaffolding protein Axin-the tumor suppressor adenomatous polyposis coli gene product (APC)-casein kinase 1 (CK1)-glycogen synthase kinase 3 (GSK3) complex to the receptor inhibit β-catenin phosphorylation and increase the stabilization of β-catenin. Subsequently, the β-catenin accumulated localizes to the nucleus and forms complexes with T-cell factor/lymphoid enhancing factor (TCF/LEF) transcription factors and consequently activates target genes, such as matrix metalloproteniase-7 (MMP-7), C-myc, Cyclin D1, and others [6]. It has been found that Wnt/β-catenin pathway was abnormally activated in a variety of cancers including breast cancer [7–9]. Wnt/β-catenin pathway regulates various biological processes that are important for cancer progression, including tumor initiation, tumor growth, and metastasis [2].
MicroRNAs (miRNAs) are small noncoding RNAs that regulate gene expression at a post-transcriptional level and monitor several biological processes [10]. Several human miRNAs have been shown in regulating the metastasis of breast cancer cells [11, 12]. MicroRNA-100 (miR-100) is a member of the miR-100 family of miRNAs and widely expressed in vertebrates [13]. The role of miR-100 in cancers seems controversial, since it can serve either as an oncogene or as a tumor suppressor in different tumor types [14, 15]. In recent years, miR-100 was reported to suppress proliferation- and survival-promoting oncogene insulin-like growth factor (IGF) 2 and inhibit breast tumorigenesis [16], and target HOXA1 to suppress breast cancer cells movement and invasion [17]. However, the role of miR-100 in breast cancer cells metastasis requires more extensive investigation.
In the present study, after finding the downregulated expression of miR-100 in breast cancer tissues and cell lines, we observed that miR-100 could inhibit the migration and invasion of breast cancer cells by transfecting miR-100 mimic in MDA-MB-231 cells and transfecting miR-100 inhibitor in MCF-7 cells. Furthermore, our results showed that miR-100 directly inhibited the expression of FZD-8 and inactivated the Wnt/β-catenin pathway in breast cancer cells. These findings suggested that miR-100 functions as a tumor suppressor in breast cancer cells and manipulation of miR-100 may provide a promoting therapeutic strategy for breast cancer treatment.
Materials and methods
Cell lines and clinical samples
The MCF-7, MDA-MB-231 cell lines, and 293T cells were all purchased from the American Type Culture Collection (ATCC). MCF-7 and 293T cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM) high glucose medium (Gibco), containing 10 % fetal bovine serum (HyClone), 100 U/ml penicillin, and 100 mg/ml streptomycin in a humidified atmosphere with 5 % CO2 at 37 °C. MDA-MB-231 cells were maintained in L15 culture medium (Gibco) in similar conditions.
Human breast cancer tissues and adjacent normal tissues for qRT-PCR analysis were obtained from 69 breast cancer patients and for in situ hybridization and immunohistochemistry were obtained from 55 breast cancer patients, who underwent surgery at First Affiliated Hospital of China Medical University between 2011 and 2012. The study has been approved by the Institutional Review Board of China Medical University, and all subjects gave their informed consent prior to their inclusion in the study.
Cell transfection
The miR-100 mimic, negative control mimic (miR-NC), miR-100 inhibitor, and negative control inhibitor (NC inhibitor) were all purchased from RiboBio (Guangzhou, China). MCF-7 and MDA-MB-231 cells (3 × 105) were seeded on 6-well plates and cultured overnight. Cells were then transfected with 15 nM miR-100 mimic, miR-NC, miR-100 inhibitor, or NC inhibitors using Lipofectamine 2000 according to the manufacturer’s instruction (Life Technologies, USA). After 48 h, the cells were used for Western blot and qRT-PCR analysis.
RNA isolation and qRT-PCR analysis
Total RNA and miRNA from breast cancer cells were isolated using a miRNA isolation Kit (Bioteke, China). The quality and quantity of the RNA and miRNA samples were assessed by nanro 2000 (thermo, USA). The reaction conditions of PCR were described as our previous reports [18]. For miR-100 expression analysis, U6 small nuclear RNA (U6 snRNA) was used as an internal control. For FZD-8, TCF-4, and LEF-1 expression analysis, GAPDH gene was used as an endogenous control. The PCR amplification primer sequences are shown in Table 1. The fold change for each miRNA and mRNA relative to the control was calculated using the 2−∆∆Ct method. The data were calculated from three independent experiments.
Western blot analysis
Cells were harvested and total proteins were extracted as previously described [18]. Proteins were resolved by SDS–PAGE and transferred onto polyvinylidene fluoride membranes by electroblotting. Membranes were blocked with 5 % milk in Tris-buffered saline with 0.1 % Tween 20 and then incubated with primary antibodies against human FZD-8 (1:1000, Abcam), β-catenin (1:1000, perprotech), MMP-7 (1:500, Boster), GSK-3β (1:1000, Abcam), and p-GSK-3β (1:1000, Abcam), and then with peroxidase-conjugated secondary antibody (Santa Cruz Biotechnology, USA), and visualized using chemiluminescence (ECL, Amersham, Freiburg, Germany), according to the manufacturer’s protocol. The results of protein expression were quantitatively analyzed with FluorChem V2.0 software (Alpha Innotech Corp, USA). The expression of GAPDH was used as a normalization control for protein loading.
Transwell migration and invasion assays
Transwell migration and invasion assays were carried out using 24-well transwell migration chambers (Corning, USA) with 8-μm pore size polyethylene membranes. For the migration assay, cells were placed in the upper chamber of each insert without Martigel coating. For the invasion assay, cells were placed in the upper chamber of each insert, which was precoated with Matrigel (BD Biosciences, USA). For both assays, MCF-7 cells and MDA-MB-231 cells were starved overnight and trypsinized and suspended in serum-free medium. The 1 × 104 cells (for the migration assay) or 1 × 105 cells (for the invasion assay) were seeded in starvation medium on the top chamber. The bottom chamber was filled with 10 % FBS in DMEM or L15 medium which acted as chemoattractant. After 24-h incubation, the cells migrating or invading to the lower chamber were fixed with 4 % paraformaldehyde for 20 min, stained with crystal violet solution for 30 min, and counted using a microscope. Three independent experiments were performed, and the data are presented as the average ± SD.
Dual luciferase reporter assay
The 293T cells (1.2 × 104) were seeded in triplicate in 24-well plates and allowed to settle over 24 h. The luciferase reporter plasmids (10 ng) containing either wild type or mutant FZD-8-3′-UTR and miR-100 mimic or miR-NC (15 nM) were co-transfected into 293T cells. Luciferase and renilla signals were measured 48 h after transfection using the Dual Luciferase Reporter Assay Kit (Promega, USA), according to the manufacturer’s protocol. Three independent experiments were performed, and the data are presented as the average ± SD.
Immunohistochemistry
Immunohistochemistry was performed as previously described [19]. Briefly, tissue sections (4 μm thick) obtained from paraffin-embedded tumor tissues from breast cancer patients were incubated with primary antibody against FZD-8 (1:200, Abcam). Images from each section were captured by a Digital Sight digital camera under a Nikon Eclipse 80i microscope (Japan, at ×200 magnification). The immunoreactivity was evaluated by two independent investigators blinded to the patients’ clinicopathological characteristics according to the percentage of stained cells and the intensity of the immunoreactivity. The intensity of immunoreactivity was scored as follows: 0 for no staining, 1 for weak staining, 2 for moderate staining, and 3 for strong staining. The percentage of stained cells was scored as follows: 0 for <5 % staining, 1 for 5–25 % staining, 2 for 26–50 % staining, 3 for 51–75 % staining, and 4 for >75 % staining. Five random fields without overlaps from each section were counted. The final immunoreactive score was determined multiplying the intensity score with the score for the percentage of positively stained cells.
In situ hybridization
A synthetic oligonucleotide probe (Shanghai Samgon Biological Engineering Technology &Serbices Co, Ltd, China) with the sequences of 5′-aacccguagauccgaacuugug-3′ was used. It was labeled by 3′ and 5′-tailing with digoxin. In situ hybridization was performed using Enhanced Sensitive ISH Detection Kit II as specified by the manufacturer (MK1030, boster, China). Sections were washed in xylene and rehydrated with serial dilutions of alcohol. The sections were then treated with 3 % H2O2 for 10 min and protease K (20 μg/ml) for 15 min at 37 °C, pre-hybridized in pre-hybridization solution for 2 h at 37 °C, and hybridized in hybridization solution at 37 °C overnight. Probe concentration was 8 μg/ml. Sections were incubated in 10 % normal goat serum for 30 min at 37 °C, then in biotinylated mouse anti-digoxigenin for 60 min at 37 °C, ultimately in SABC-AP for 30 min at 37 °C. DAB (3,3-diaminobenzidine) substrate was applied to the sections. Sections were counterstained with hematoxylin. Quantification of the staining intensity of miR-100 was performed through image analysis the same as immunohistochemistry.
Statistical analysis
Analyses were performed using SPSS17.0. A two-tailed Student’s t test was used to evaluate the statistical significance of the differences between two groups. One-way analysis of variance (ANOVA) was used to compare the differences among three or more groups. The correlation of the expression levels of miR-100 and FZD-8 with the lymph node metastasis in human breast cancer tissues was analyzed by Mann-Whitney U test. The Pearson rank correlation analysis was applied to assess the association between the expression of miR-100 and FZD-8. Probability values <0.05 were considered statistically significant.
Results
MiR-100 expression is downregulated in breast cancer tissues and cell lines
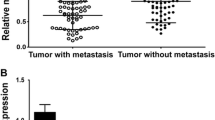
To assess the clinical relevance of miR-100 in human breast cancer patients, we first measured the expression of miR-100 in 69 pairs of human breast cancer tissues and adjacent normal breast tissues by qRT-PCR. We observed that miR-100 expression in human breast cancer tissues was significantly lower than in adjacent normal breast tissues (Fig. 1a, P < 0.01). In addition, the expression of miR-100 was decreased at least 2-fold compared with adjacent normal breast tissues in 47.8 % (33/69) of human breast cancer cases (Fig. 1b). Furthermore, the low expression of miR-100 was found to be closely correlated with lymph node metastasis by Mann-Whitney U test (Fig. 1c, P < 0.05). We also measured miR-100 expression in human mammary epithelial MCF-10A cells, breast cancer SKBR3, MCF-7, T47D, BT549, MDA-MB-231, and MDA-MB-435S cells by qRT-PCR. MiR-100 expression was significantly downregulated in all these breast cancer cell lines compared with MCF-10A cells (Fig. 1d, P < 0.0001). Taken together, the data suggested that the expression of miR-100 was downregulated in breast cancer tissues and established cell lines and the low expression of miR-100 might be relevant to metastasis of breast cancer.

The expression of miR-100 is downregulated in breast cancer tissues and cell lines. a MiR-100 expression in 69 pairs of human breast cancer tissues (cancer) and adjacent normal tissue (normal) by qRT-PCR. The expression was normalized to the level of U6 in each sample. b Fold changes of miR-100 of each individual paired sample. The data were represented as log2-fold change (cancer/normal). c The low expression of miR-100 was correlated with lymph node metastasis in 69 cases of human breast cancer tissues by Mann-Whitney U test. d MiR-100 expression in human mammary epithelial MCF-10A cell, breast cancer SKBR3, MCF-7, T47D, BT549, MDA-MB-231, and MDA-MB-435S cells was measured by qRT-PCR. Data are presented as average ± SD from three independent experiments. ***P < 0.0001 versus MCF-10A cells
MiR-100 suppresses the migration and invasion of breast cancer cells
To observe the effects of miR-100 on the migration and invasion of breast cancer cells, we first detected the expression levels of miR-100 after transfecting miR-100 mimic in invasive MDA-MB-231 cells using qRT-PCR analysis. It is noteworthy that the expression of miR100 was increased approximately 290-fold at 48 h after transfection with the miR-100 mimic in comparison to transfection with miR-negative control (miR-NC) (P < 0.01, Fig. 2a). Next, we measured the effects of transfecting miR-100 mimic on the migration and invasive abilities of MDA-MB-231 cells by transwell migration and invasion assays. The results showed that the overexpression of miR-100 suppressed MDA-MB-231 cells migration to 55 % of the control and invasion to 50 % of the control (P < 0.05, Fig. 2b).

MiR-100 suppresses the migration and invasion of breast cancer cells. a The relative expression of miR-100 was detected by qRT-PCR at 48 h after transfection with miR-100 mimic or miR-NC in MDA-MB-231 breast cancer cells. b The cells migration and invasion abilities were measured by transwell migration and invasion assays after transfection with miR-100 mimic or miR-NC in MDA-MB-231 cells. Cells that migrated and invaded into the lower transwell chambers were counted. The cell number migrating and invading into the lower chambers in miR-NC group were set as 1. c The relative expression of miR-100 was detected by qRT-PCR at 48 h after transfection with miR-100 inhibitor or NC inhibitor in MCF-7 breast cancer cells. d The cell migration and invasion abilities were measured by transwell migration and invasion assays after transfection with miR-100 inhibitor or NC inhibitor in MCF-7 cells. Cells that migrated and invaded into the lower transwell chambers were counted (100×). The cell number migrating and invading into the lower chambers in NC-inhibitor group was set as 1. Data are presented as average ± SD from three independent experiments. *P < 0.05; **P < 0.01 versus miR-NC group or NC inhibitor group
Inversely, the expression of miR-100 in MCF-7 cells at 48 h after transfection with the miR-100 inhibitor was inhibited by approximately 55 % compared to transfection with the NC inhibitor (P < 0.05, Fig. 2c). Furthermore, the inhibition of miR-100 expression increased MCF-7 cells migration to 160 % of the control and invasion to 185 % of the control (P < 0.01, Fig. 2d). Taken together, the data suggested that miR-100 suppressed breast cancer cell migration and invasion.
FZD-8 is a direct target of miR-100
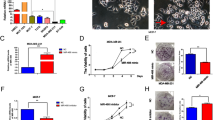
To further explore the mechanisms underlying the inhibitory role of miR-100 in the migration and invasion of breast cancer cells, we predicted the putative targets of miR-100 by TargetScan and RNAhybrid. We focused on the genes related to Wnt signaling pathway involved in the tumor metastasis. We found that FZD-8, a receptor of Wnt signaling pathway, was one of the targets of miR-100 (Fig. 3a). To test whether FZD-8 was a direct target of miR-100, we constructed a luciferase reporter plasmid containing FZD-8-3′-untranslated region (3′-UTR) harboring a conserved miR-100 binding site (pGL3-FZD-8-3′UTR) and a plasmid containing FZD-8-3′-UTR with miR-100 target sequences mutated (pGL3-FZD-8-3′UTR mu). The pGL3-FZD-8-3′UTR or pGL3-FZD-8-3′UTR mu was cotransfected with the miR-100 mimic or miR-NC into 293T cells. The reporter assay showed that miR-100 mimic significantly decreased the luciferase activity by approximately 55 % in 293T cells co-transfected with the pGL3-FZD-8-3′UTR (P < 0.05, Fig. 3b). However, the luciferase activity in the cells co-transfected with the pGL3-FZD-8-3′UTR mu was not significantly reduced. Our findings suggested that FZD-8 was a direct target of miR-100.

FZD-8 is a direct target of miR-100 in breast cancer cells. a Schematics of highly conserved miR-100 binding site in human FZD-8-3′UTR. b Luciferase reporter assay in 293T cells co-transfected with miR-100 mimic or miR-NC and PGL3-FZD-8-3′UTR or PGL3-FZD-8-3′UTR mu. Relative luciferase activity was measured at 48 h after transfection. c The relative expression of FZD-8 was detected by qRT-PCR at 48 h after transfection with miR-100 mimic or miR-NC in MDA-MB-231 breast cancer cells. d Representative western blot images and the relative FZD-8 protein expression were analyzed with GAPDH as an internal control at 48 h after transfection with miR-100 mimic or miR-NC in MDA-MB-231 breast cancer cells. e The relative expression of FZD-8 was detected by qRT-PCR at 48 h after transfection with miR-100 inhibitor or NC inhibitor in MCF-7 breast cancer cells. f Representative western blot images and the relative FZD-8 protein expression were analyzed with GAPDH as an internal control at 48 h after transfection with miR-100 inhibitor or NC inhibitor in MCF-7 breast cancer cells. Data are presented as average ± SD from three independent experiments. *P < 0.05; **P < 0.01 versus miR-NC group or NC inhibitor group
Next, we found that the transfection of miR-100 mimic could decrease FZD-8 mRNA and protein expression levels to approximately 40 and 70 % of the miR-NC in MDA-MB-231 cells (P < 0.05, Fig. 3c, d). In addition, the inhibition of endogenous of miR-100 by miR-100 inhibitor increased the FZD-8 mRNA and protein expression levels to approximately 6.3-fold and 144 % of the NC inhibitor in MCF-7 cells (P < 0.05, Fig. 3e, f). These results demonstrated that miR-100 could inhibit the expression of FZD-8 in breast cancer cells.
Ectopic miR-100 expression inhibits the activation of Wnt/β-catenin signaling pathway
FZD-8 is an important receptor of Wnt signaling pathway. To further investigate whether miR-100 inhibits the activation of Wnt pathway by targeting FZD-8 in breast cancer cells, we detected the protein expression levels of β-catenin, GSK-3β, and p-GSK-3β, the important components of Wnt/β-catenin pathway, and MMP-7, a major target gene of Wnt/β-catenin pathway related to metastasis, in MDA-MB-231 cells transfected with miR-100 mimic. The results showed that the transfection of miR-100 mimic significantly suppressed the protein expression levels of β-catenin and MMP-7 (Fig. 4a, P < 0.05, P < 0.01) and increased the expression of GSK-3β and p-GSK-3β in MDA-MB-231 cells, compared with miR-NC (Fig. 4b, P < 0.01). In addition, we also found that the ectopic miR-100 expression obviously decreased the mRNA expression levels of T cell factor-4 (TCF-4) and lymphoid enhancer factor-1 (LEF-1), the important transcription factors of Wnt/β-catenin pathway, in MDA-MB-231 cells (Fig. 4c, d, P < 0.05).

Ectopic miR-100 expression inhibits the activation of Wnt/β-catenin signaling pathway. a Representative western blot images and the relative protein expression levels of β-catetin and MMP-7, and b GSK-3β and p-GSK-3β were analyzed with GAPDH as an internal control at 48 h after transfection with miR-100 mimic or miR-NC in MDA-MB-231 breast cancer cells. The relative mRNA expression levels of TCF-4 c and LEF-1 d were detected by qRT-PCR at 48 h after transfection with miR-100 mimic or miR-NC in MDA-MB-231 breast cancer cells. Data are presented as average ± SD from three independent experiments. *P < 0.05; **P < 0.01 versus miR-NC group
Furthermore, the inhibition of endogenous of miR-100 by miR-100 inhibitor significantly increased the protein expression levels of β-catetin and MMP-7 (Fig 5a, P < 0.05) and reduced the protein expression levels of GSK-3β and p-GSK-3β (Fig 5b, P < 0.05), and increased mRNA expression levels of TCF-4 and LEF-1 in MCF-7 breast cancer cells (Fig. 5c, d, P < 0.01, P < 0.05). Taken together, the data suggested that miR-100 could suppress the migration and invasion of breast cancer cells by targeting FZD-8 and inhibiting the activation of Wnt/β-catenin signaling pathway.

MiR-100 inhibitor increases the activation of Wnt/β-catenin signaling pathway. a Representative western blot images and the relative protein expression levels of β-catetin and MMP-7, and b GSK-3β and p-GSK-3β were analyzed with GAPDH as an internal control at 48 h after transfection with miR-100 inhibitor or NC inhibitor in MCF-7 breast cancer cells. The relative mRNA expression levels of TCF-4 (c) and LEF-1 (d) were detected by qRT-PCR at 48 h after transfection with miR-100 inhibitor or NC inhibitor in MCF-7 breast cancer cells. Data are presented as average ± SD from three independent experiments. *P < 0.05; **P < 0.01 versus NC inhibitor group
The expression of miR-100 is negatively correlated with the expression of FZD-8 protein in human breast cancer tissues
To further evaluate the relevance of the endogenous expression of miR-100 and FZD-8, we measured the expression of miR-100 using in situ hybridization and the expression of FZD-8 protein by immunohistochemistry in 55 pairs of human breast cancer tissues and adjacent normal tissues with tissue microarray (TMA). As shown in Fig. 6a, b, the expression of FZD-8 was significantly higher in human breast cancer tissues compared with the adjacent normal tissues (P < 0.01). We also analyzed the correlation of the expression levels of miR-100 and FZD-8 with the lymph node metastasis in 55 cases of human breast cancer tissues by Mann-Whitney U test and found that the high expression of FZD-8 was closely correlated with lymph node metastasis and the low expression of miR-100 was closely correlated with lymph node metastasis (Fig. 6c, d, P < 0.01). Pearson rank correlation analysis showed that the expression of miR-100 was inversely related to the expression of FZD-8 protein in breast cancer tissues (Fig. 6e, P < 0.01).

The expression of miR-100 is negatively correlated with the expression of FZD-8 in human breast cancer tissues. a Representative images of miR-100 expression measured by in situ hybridization and FZD-8 expression measured by immunohistochemistry in 55 pairs of human breast cancer tissues (cancers) and adjacent normal tissues (normal) with tissue microarray (TMA). b The expression of FZD-8 in 55 pairs of human breast cancer tissues and adjacent normal tissues. c The correlation of expression of FZD-8 with lymph node metastasis was analyzed in 55 cases of human breast cancer tissues by Mann-Whitney U test. d The correlation of expression of miR-100 with lymph node metastasis was analyzed in 55 cases of human breast cancer tissues by Mann-Whitney U test. e Correlation analysis showed that the miR-100 expression was negatively related to the expression of FZD-8 in breast cancer tissues
Discussion
Tumor metastasis is the leading cause of death for cancer patients. It is very important to find potent factors to inhibit the metastasis of cancer cells. Several miRNAs have been found to function as tumor suppressors in tumor metastasis. Han et al. found that miR-29a suppressed the growth, migration, and invasion of lung adenocarcinoma cells by targeting carcinoembryonic antigen-related cell adhesion molecule 6 (CEACAM6) [20]. Nishikawa et al. reported that miR-29s acted as tumor suppressors, contributing to cancer cell migration and invasion and directly targeting laminin signaling in prostate cancer [21]. In our studies, we found that miR-100 inhibited the migration and invasion of breast cancer cells by targeting FZD-8 and inhibiting the Wnt/β-catenin signaling pathway.
Recently, other groups have reported the functions of miR-100 in different types of cancers. Chen et al. reported that downregulation of miR-100 correlated with tumor progression and poor prognosis in hepatocellular carcinoma [22]. Peng et al. found that miR-100 could affect the growth of epithelial ovarian cancer cells by post-transcriptionally regulating polo-like kinase 1 (PLK1) expression [23]. Petrelli et al. demonstrated that miR-100 inhibited maintenance and expansion of breast cancer stem cells in basal-like cancer through PLK1 downregulation [24]. In the present study, we observed that the expression of miR-100 was significantly downregulated in human breast cancer tissues than the adjacent normal breast tissues and also lower in breast cancer cell lines compared with MCF-10A cells. In addition, we found that the low expression of miR-100 was correlated with lymph node metastasis of breast cancer patients. Furthermore, we demonstrated that the overexpression of miR-100 suppressed the migration and invasion of MDA-MB-231 breast cancer cells, and the inhibition of miR-100 expression increased the migration and invasion of breast cancer MCF-7 cells, suggesting that miR-100 was an anti-metastasis-miR in breast cancer cells. Our findings were consistent with the reports by Chen et al. [17]. Differently, they showed that miR-100 targeted and inhibited the expression of HOXA1 to inhibit tumor proliferation, invasion, and migration, while we first demonstrated that the expression of miR-100 was negatively correlated with the FZD-8 expression in human breast cancer tissues and miR-100 directly targeted and inhibited the expression of FZD-8 and the activation of WNT/β-catenin signaling pathway, resulting in the decreased migration and invasion abilities of breast cancer cells.
Wnt/β-catenin signaling pathway has been demonstrated to play an important role in the development and tumor metastasis. Wnt/β-catenin pathway transduced by LRP 5/6 and Frizzled receptor complexes leads to nuclear translocation of β-catenin and its interaction with TCF/LEF factors to regulate transcription [6]. Several miRNAs have been shown to target the key components of WNT/β-catenin pathway and regulate the biological function of cancer cells. MiR-612 was found to suppress the stemness, migration, and invasion of liver cancer cells via Wnt/β-catenin signaling pathway [25]. Subramanian et al. reported that miR-29b antagonized transactivation of β-catenin target genes by downregulating coactivators of β-catenin in human colorectal cancer cells [26]. Zhao et al. demonstrated that miR-30c inhibited multiple myeloma cell proliferation, invasion, and migration regulated by the Wnt/BCL9/β-catenin transcriptional complex [27]. Previous studies have shown that FZD-8, one of the Frizzled family of receptors for the Wnt ligands, played a major role in human lung cancer [28]. Recently, it was reported that FZD-8-mediated Wnt/β-catenin pathway played an important role in the drug resistance of triple-negative breast cancer cells [29]. In this study, we showed that FZD-8 was a direct target of miR-100. The transfection of miR-100 mimic could decrease FZD-8 mRNA and protein expression levels, suppress the protein expression levels of β-catenin and MMP-7, and the mRNA expression levels of TCF-4 and LEF-1, and increase the protein expression levels of GSK-3β and p-GSK-3β in MDA-MB-231 cells, and the transfection of miR-100 inhibitor in MCF-7 cells showed the opposite effects. Moreover, we also found that miR-100 expression was negatively correlated with FZD-8 expression, and the high expression of FZD-8 and the low expression of miR-100 were closely related to the lymph node metastasis in breast cancer tissues. Our results suggested that miR-100 inhibited the activation of Wnt/β-catenin pathway by targeting FZD-8 in breast cancer cells.
In summary, our findings suggest that miR-100 can suppress the migration and invasion of breast cancer cells by targeting FZD-8 and inhibiting the Wnt/β-catenin signaling pathway. Our studies provide new insights into the molecular mechanisms underlying the metastasis of breast cancer. However, the complex regulatory network of miR-100 in regulating the migration and invasion of breast cancer should be explored further.
References
Gangadhara S, Barrett-Lee P, Nicholson RI, Hiscox S. Pro-metastatic tumor-stroma interactions in breast cancer. Future Oncol. 2012;8(11):1427–42. doi:10.2217/fon.12.134.
Klemm F, Bleckmann A, Siam L, Chuang HN, Rietkotter E, Behme D, et al. beta-catenin-independent WNT signaling in basal-like breast cancer and brain metastasis. Carcinogenesis. 2011;32(3):434–42. doi:10.1093/carcin/bgq269.
Tang B, Vu M, Booker T, Santner SJ, Miller FR, Anver MR, et al. TGF-beta switches from tumor suppressor to prometastatic factor in a model of breast cancer progression. J Clin Invest. 2003;112(7):1116–24. doi:10.1172/JCI18899.
Hu YY, Zheng MH, Zhang R, Liang YM, Han H. Notch signaling pathway and cancer metastasis. Adv Exp Med Biol. 2012;727:186–98. doi:10.1007/978-1-4614-0899-4_14.
Clevers H. Wnt/beta-catenin signaling in development and disease. Cell. 2006;127(3):469–80. doi:10.1016/j.cell.2006.10.018.
MacDonald BT, Tamai K, He X. Wnt/beta-catenin signaling: components, mechanisms, and diseases. Dev Cell. 2009;17(1):9–26. doi:10.1016/j.devcel.2009.06.016.
Khramtsov AI, Khramtsova GF, Tretiakova M, Huo DZ, Olopade OI, Goss KH. Wnt/beta-Catenin Pathway Activation Is Enriched in Basal-Like Breast Cancers and Predicts Poor Outcome. Am J Pathol. 2010;176(6):2911–20. doi:10.2353/ajpath.2010.091125.
Arend RC, Londono-Joshi AI, Straughn Jr JM, Buchsbaum DJ. The Wnt/beta-catenin pathway in ovarian cancer: a review. Gynecol Oncol. 2013;131(3):772–9. doi:10.1016/j.ygyno.2013.09.034.
Serafino A, Moroni N, Zonfrillo M, Andreola F, Mercuri L, Nicotera G, et al. WNT-pathway components as predictive markers useful for diagnosis, prevention and therapy in inflammatory bowel disease and sporadic colorectal cancer. Oncotarget. 2014;5(4):978–92.
Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–97.
Liu P, Tang H, Chen B, He Z, Deng M, Wu M, et al. miR-26a suppresses tumour proliferation and metastasis by targeting metadherin in triple negative breast cancer. Cancer Lett. 2015;357(1):384–92. doi:10.1016/j.canlet.2014.11.050.
Chan SH, Huang WC, Chang JW, Chang KJ, Kuo WH, Wang MY, et al. MicroRNA-149 targets GIT1 to suppress integrin signaling and breast cancer metastasis. Oncogene. 2014;33(36):4496–507. doi:10.1038/onc.2014.10.
Christodoulou F, Raible F, Tomer R, Simakov O, Trachana K, Klaus S, et al. Ancient animal microRNAs and the evolution of tissue identity. Nature. 2010;463(7284):1084–8. doi:10.1038/nature08744.
Zheng YS, Zhang H, Zhang XJ, Feng DD, Luo XQ, Zeng CW, et al. MiR-100 regulates cell differentiation and survival by targeting RBSP3, a phosphatase-like tumor suppressor in acute myeloid leukemia. Oncogene. 2012;31(1):80–92. doi:10.1038/onc.2011.208.
Li ZP, Li X, Yu C, Wang M, Peng F, Xiao J, et al. MicroRNA-100 regulates pancreatic cancer cells growth and sensitivity to chemotherapy through targeting FGFR3. Tumor Biol. 2014;35(12):11751–9. doi:10.1007/s13277-014-2271-8.
Gebeshuber CA, Martinez J. miR-100 suppresses IGF2 and inhibits breast tumorigenesis by interfering with proliferation and survival signaling. Oncogene. 2013;32(27):3306–10. doi:10.1038/onc.2012.372.
Chen D, Sun Y, Yuan Y, Han Z, Zhang P, Zhang J, et al. miR-100 induces epithelial-mesenchymal transition but suppresses tumorigenesis, migration and invasion. PLoS Genet. 2014;10(2):e1004177. doi:10.1371/journal.pgen.1004177.
Ma MT, He M, Wang Y, Jiao XY, Zhao L, Bai XF, et al. MiR-487a resensitizes mitoxantrone (MX)-resistant breast cancer cells (MCF-7/MX) to MX by targeting breast cancer resistance protein (BCRP/ABCG2). Cancer Lett. 2013;339(1):107–15. doi:10.1016/j.canlet.2013.07.016.
Bai X, Song Z, Fu Y, Yu Z, Zhao L, Zhao H, et al. Clinicopathological significance and prognostic value of DNA methyltransferase 1, 3a, and 3b expressions in sporadic epithelial ovarian cancer. PLoS One. 2012;7(6):e40024. doi:10.1371/journal.pone.0040024.
Han HS, Son SM, Yun J, Jo YN, Lee OJ. MicroRNA-29a suppresses the growth, migration, and invasion of lung adenocarcinoma cells by targeting carcinoembryonic antigen-related cell adhesion molecule 6. FEBS Lett. 2014;588(20):3744–50. doi:10.1016/j.febslet.2014.08.023.
Nishikawa R, Goto Y, Kojima S, Enokida H, Chiyomaru T, Kinoshita T, et al. Tumor-suppressive microRNA-29s inhibit cancer cell migration and invasion via targeting LAMC1 in prostate cancer. Int J Oncol. 2014;45(1):401–10. doi:10.3892/ijo.2014.2437.
Chen P, Zhao X, Ma L. Downregulation of microRNA-100 correlates with tumor progression and poor prognosis in hepatocellular carcinoma. Mol Cell Biochem. 2013;383(1-2):49–58. doi:10.1007/s11010-013-1753-0.
Peng DX, Luo M, Qiu LW, He YL, Wang XF. Prognostic implications of microRNA-100 and its functional roles in human epithelial ovarian cancer. Oncol Rep. 2012;27(4):1238–44. doi:10.3892/or.2012.1625.
Petrelli A, Carollo R, Cargnelutti M, Iovino F, Callari M, Cimino D, et al. By promoting cell differentiation, miR-100 sensitizes basal-like breast cancer stem cells to hormonal therapy. Oncotarget. 2015;6(4):2315–30.
Tang J, Tao ZH, Wen D, Wan JL, Liu DL, Zhang S, et al. MiR-612 suppresses the stemness of liver cancer via Wnt/beta-catenin signaling. Biochem Biophys Res Commun. 2014;447(1):210–5. doi:10.1016/j.bbrc.2014.03.135.
Subramanian M, Rao SR, Thacker P, Chatterjee S, Karunagaran D. MiR-29b downregulates canonical Wnt signaling by suppressing coactivators of beta-catenin in human colorectal cancer cells. J Cell Biochem. 2014;115(11):1974–84. doi:10.1002/jcb.24869.
Zhao JJ, Lin JH, Zhu D, Wang XJ, Brooks D, Chen M, et al. miR-30-5p functions as a tumor suppressor and novel therapeutic tool by targeting the oncogenic wnt/beta-catenin/bcl9 pathway. Cancer Res. 2014;74(6):1801–13. doi:10.1158/0008-5472.CAN-13-3311-T.
Wang HQ, Xu ML, Ma J, Zhang Y, Xie CH. Frizzled-8 as a putative therapeutic target in human lung cancer. Biochem Biophys Res Commun. 2012;417(1):62–6. doi:10.1016/j.bbrc.2011.11.055.
Yin S, Xu L, Bonfil RD, Banerjee S, Sarkar FH, Sethi S, et al. Tumor-initiating cells and FZD8 play a major role in drug resistance in triple-negative breast cancer. Mol Cancer Ther. 2013;12(4):491–8. doi:10.1158/1535-7163.MCT-12-1090.
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (Grant No. 81373427), Program for Liaoning Innovative Research Team in University, LNIRT, China (Grant No. LT2014016), the Liaoning Provincial Science and Technology Program, China (Grant No. 2013225079), Program for Liaoning Excellent Talents in University, China (Grant No. LJQ2014084), and the S&T Projects in Shenyang, China (Grant No. F14-232-6-05).
Authors’ contributions
Minjie Wei and Miao He designed the experiments. Qian Jiang, Huizhe Wu, Zhaojin Yu, Longyang Jiang, Yan Wang, and Xingyue Zong performed the experiments. Qian Jiang, Mengtao Ma, and Miao He analyzed the data. Shu Guan and Feng Jin gave technical and material support. Miao He, Qian Jiang, and Minjie Wei wrote and reviewed the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
None.
Rights and permissions
About this article

Cite this article
Jiang, Q., He, M., Guan, S. et al. MicroRNA-100 suppresses the migration and invasion of breast cancer cells by targeting FZD-8 and inhibiting Wnt/β-catenin signaling pathway. Tumor Biol. 37, 5001–5011 (2016). https://doi.org/10.1007/s13277-015-4342-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13277-015-4342-x