
Abstract
BTB (broad-complex, tramtrack, and bric-a-brac) family proteins are characterized by the presence of a protein–protein interaction BTB domain. BTB proteins have diverse functions, including transcriptional regulation, protein degradation, chromatin remodeling, and cytoskeletal regulation. However, little is known about this gene family in tomato (Solanum lycopersicum), the most important model plant for crop species. In this study, 38 BTB genes were identified based on tomato whole-genome sequence. Phylogenetic analysis of BTB proteins in tomato revealed that SlBTB proteins could be divided into at least 4 subfamilies. The SlBTB proteins contains 1–3 BTB domains, and several other types of functional domains, including KCTD (Potassium channel tetramerization domain-containing), the MATH (meprin and TRAF homology), ANK (Ankyrin repeats), NPR1 (nonexpressor of pathogenesis-related proteins1), NPH3 (Nonphototropic Hypocotyl 3), TAZ zinc finger, C-terminal Kelch, Skp1 and Arm (Armadillo/beta-catenin-like repeat) domains are also found in some tomato BTB proteins. Moreover, their expression patterns in tissues/stages, in response to different abiotic stress treatments and hormones were also investigated. This study provides the first comprehensive analysis of BTB gene family in the tomato genome. The data will undoubtedly be useful for better understanding the potential functions of BTB genes, and their possible roles in mediating hormone cross-talk and abiotic stress in tomato as well as in some other relative species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The BTB (broad-complex, tramtrack, and bric-a-brac) protein found primarily in Drosophila contains a highly conserved domain of approximately 115 amino acids (Zollman et al. 1994). Also known as the POZ (Pox virus and zinc finger) domain, the BTB domain is an evolutionarily conserved protein–protein interaction motif (Bardwell and Treisman 1994; Cheng et al. 2014). The BTB domain core structurally consists of five α-helices, with A1/2 and A4/5 forming α-helical hairpins and three β-strands forming a β-sheet (Stogios et al. 2005). In addition the BTB core domain, the different BTB subclasses include amino (N)- and carboxy (C)-terminal BTB extension regions that contribute to protein-specific functions (Chaharbakhshi and Jemc 2016).
BTB proteins have been widely studied in animals given its vital role in multiple developmental and disease contexts, including transcription repression (Ahmad et al. 2003; Melnick et al. 2000), protein ubiquitination/degradation (Furukawa et al. 2003; Geyer et al. 2003; Pintard et al. 2003; Wilkins et al. 2004; Xu et al. 2003), lymphocyte development, axon guidance, gonad morphogenesis, and leukemia (Ito et al. 2013; Ko et al. 2006; Lim 2014; Robinson and Cooley 1997; Siggs et al. 2012; Weyers et al. 2011). Recently, the BTB protein has been discovered to be crucial in plant development (Chevrier et al. 2014; Juranic et al. 2012; Robert et al. 2009; Xu et al. 2016), nitrogen use efficiency (Araus et al. 2016), germination (Kim et al. 2016), and biotic (Boyle et al. 2009) and abiotic stresses (Mandadi et al. 2009).
The BTB family proteins with DNA-binding domains suggestively regulate gene expression; however, the BTB domain also combines with other domains, thereby extending the functional range of the BTB protein family (Chaharbakhshi and Jemc 2016). Subclasses of zinc finger (ZF), Kelch, BTB and C-terminal Kelch (BACK), meprin and TRAF homology (MATH), ANK (Ankyrin repeats), PHR, and Ras homology (Rho) domains are common in some BTB proteins. With the presence of these domains, the BTB protein gene family can be divided into numerous subfamilies, including BTB-only proteins, BTB-ZF proteins, BTB-Kelch proteins, BTB-BACK proteins, BTB-BACK-Kelch proteins, MATH-BTB proteins, BTB-ANK proteins, BTB-BACK, BTB-PHR proteins, and Rho-BTB proteins (Cheng et al. 2014; Stogios et al. 2005).
Family members of the BTB protein gene are common in eukaryotes, endosymbiotic bacteria, and viruses, but are absent in other prokaryotic systems (Chaharbakhshi and Jemc 2016; Perez-Torrado et al. 2006). Genome-wide investigation of the BTB protein gene family has been performed in several species, including humans (Homo sapiens), nematodes (Caenorhabditis elegans), mice (Mus musculus), Arabidopsis thaliana, fruit flies (Drosophila melanogaster), malaria mosquitos (Anopheles gambiae), and silkworms (Bombyx mori) (Cheng et al. 2014; Stogios et al. 2005). Tomato (Solanum lycopersicum) is a nutritive vegetable and a model plant for research. However, abiotic stresses, such as drought, salinity, and extreme temperature, inhibit the growth, development, and productivity of crops, and have eventually become overwhelming with global climate changes (Hou et al. 2009; Mittler 2006). In our previous expression profiling of two drought-tolerant introgression lines (IL) derived from Solanum pennellii (wild drought-tolerant species, LA0716) and their recurrent parent S. lycopersicum (M82) (Gong et al. 2010), several BTB genes were differentially expressed in response to drought stress. Therefore, we identified all BTB members in the tomato genome to improve our understanding of the BTB genes in tomato. This present work identified 38 putative BTB genes and analyzed their gene classification, chromosome distribution, phylogenetic comparison, and exon–intron organization. Moreover, the expression patterns under various hormones and stress treatments were investigated. This study details the BTB gene family and provides a solid basis for the functional characterization of BTB genes in tomato.
Materials and methods
Plant materials
Tomato (S. pennellii LA0716 and S. lycopersicum cv. M82, referred to as “LA0716” and “M82”, respectively) plants were grown in a glasshouse with a 16 h light/8 h dark cycle. For the gene expression profiling analysis, identical five-leaf stage tomato plants, including LA0716 and M82, were subjected to various stresses or plant growth regulator treatments simultaneously. Tomato leaves were directly sprayed with 100 μM abscisic acid (ABA), 100 μM gibberellic acid (GA), 100 μM salicylic acid (SA), 1 mM ethephon (Eth), and 100 μM methylviologen (MV) until run-off. Salt stress was simulated by watering plants with 200 mM NaCl solution, and drought dehydration (DH) stress was simulated by placing detached leaves on filter paper under 70% relative humidity at 25 °C. Cold stress was imposed by transferring plants to a growth chamber and holding the plants at 4 °C. Control plants were mock-treated with water or no treatment. Untreated plants were used as corresponding controls to avoid the effects of the circadian clock on the differences in gene expression. Then, the leaves were harvested at 0, 1, 4, 6, 12, and 24 h, immediately frozen in liquid nitrogen, and stored at −80 °C. For all experiments, three biological samples were collected for further analysis.
Identification of putative tomato BTB proteins
The first complete cDNA sequence of the tomato BTB gene was isolated from different expression profiles between drought-tolerant ILs and their recurrent parent, M82 (Gong et al. 2010). To identify new BTB homologs in tomato, the amino acid sequences of BTB genes were subjected to BLASTP searches in the SOL Genomics Network (SGN: http://solgenomics.net/) annotation group release 2.4 tomato proteins (2.40) (E value ≤1 × 10−5). Moreover, homolog searching was performed using “BTB” or “POZ” as a keyword in the SGN tomato database (Search Genes and Loci: https://solgenomics.net/search/locus). Afterward, they were confirmed using BLASTN search against the SGN and National Center for Biotechnology Information database. Redundant sequences were removed after similarity comparison. The open reading frames were determined by Softberry FGENESH (http://linux1.softberry.com) and homologous alignment.
Phylogenetic analysis and sequence alignment
Multiple sequence alignments of SlBTB proteins were performed using ClustalW (http://www.ebi.ac.uk/Tools/msa/clustalo/), and a phylogenetic tree was constructed using the neighbor-joining (NJ) method with MEGA (version 5.05) software (Tamura et al. 2011). Domains were identified with SMART (http://smart.embl-heidelberg.de/), pfam (http://pfam.xfam.org/), and InterProscan (http://www.ebi.ac.uk/interpro/search/sequence-search) programs.
Chromosomal location, gene structure, and duplication analysis
The BTB genes were mapped to tomato chromosomes by identifying their chromosomal positions according to the SGN database (Consortium TG 2012). Accordingly, their cDNA sequences and the corresponding genomic DNA sequences of the BTB members were obtained; afterward, exons and introns were identified by comparing the genomic DNA and cDNA sequences using Gene Structure Display Server (GSDS; http://gsds.cbi.pku.edu.cn) (Guo et al. 2007). MCScanX software (http://chibba.pgml.uga.edu/mcscan2/) was used to find potential tomato BTB gene duplicates. All tomato BTB genes were compared with themselves and others using the BLASTp program, with an E value <1 × 10−5. The BLAST search outputs were imported into MCScanX software using the default criterion.
Cis-element prediction for BTB gene promoter
The BTB sequences were used as BLASTN search queries against the tomato whole genome scaffolds (version 2.40) and S. pennellii WGS Chromosome data at the SGN website. Promoter sequences (1.5 kb upstream of 5′UTR) of all the annotated BTB genes were submitted to the PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) for cis-element prediction.
Tissue expression analysis of SlBTBs of tomato
To analyze the expression patterns of SlBTBs, we used RNA-seq data, as generated previously in S. lycopersicum cv Heinz and the tomato expression platform (Bostan and Chiusano 2015; Tomato Genome 2012). The data include reads from 20 collections of tomato representing two biological replicates from ten different tissues/stages, including leaf, root, flower, flower bud, 1, 2, 3 cm fruit, mature green fruit, breaker fruit, and fruit at 10 days. Gene expression level was defined based on the iTAG mRNA loci and was normalized by reads per kilobases per million (RPKM) for each tissue/stage, and the log2 transformation was selected from the platform (http://www.cab.unina.it/NexGenEx-Tom).
RNA isolation and real-time RT-PCR
Gene expression patterns were examined by isolating the total RNA using the TRIzol reagent (Sangon, Shanghai, China). Approximately 1 μg of total RNA was reverse transcribed using a PrimeScript RT Reagent Kit with a gDNA Eraser (TaKaRa Dalian, China). Quantitative RT-PCR was performed on a CFX96 Real-Time PCR (Bio-Rad, USA) with Taq-HS PCR SYBR Green I Master Mix Kit (NOVA, China) using gene-specific primers (Supplementary Table S1), with β-actin (Solyc11g005330.1.1) transcripts as internal control. PCR amplification consisted of an initial incubation at 94 °C for 5 min, followed by 40 cycles of 94 °C for 10 s, 58 °C for 15 s, and 72 °C for 20 s. Data were gathered during the extension step. Melting-curve acquisition and analyses were performed on the cycler.
Results
Identification of BTB genes in tomato
In this study, 38 BTB genes in the tomato genome cDNA release 2.40 were identified (Table 1). For consistency, we named these genes SlBTB1 to SlBTB38 based on their distributions and relative linear orders among their respective chromosomes. Detailed information of each SlBTB, including gene name, Soly ID, chromosome location, genomic position, length of coding sequence and protein, theoretical isoelectric point, and molecular weight are presented in Table 1. The 38 deduced proteins had divergent lengths, resulting in diverse isoelectric points and molecular weights. The sequence length of the deduced proteins ranged from 93 (SlBTB16) to 963 (SlBTB31). The tomato BTB genes encode proteins with predicted molecular weights of 10.5–107.6 kDa and theoretical isoelectric points from 4.67 (SlBTB15) to 9.41 (SlBTB6). Most tomato BTB proteins were predicted to be located in the nuclei and cytoplasm, but a few might be located in other subcellular compartments, such as the chloroplast and mitochondria .
Protein sequence, phylogenetic, and duplication analyses of the tomato BTB family
All identified tomato BTB proteins contained at least one BTB domain. Most BTB proteins consisted of 1 BTB domain, except for the SlBTB38, SlBTB31, and SlBTB21, which had 2 BTB domains, whereas the BTB10 contained 3 BTB domains. These domains were irregularly scattered on the amino acid, whereas some BTB domains were located in the N-terminal and some were in the C-terminal (Fig. 1). Among the 38 BTB proteins shown in the Fig. 1, 18 BTB domains were located in the N-terminal and 8 BTB domains were located in C-terminal. Moreover, the tomato BTB proteins contained other domains, such as the MATH domain in SlBTB1 and SlBTB12. Five members (SlBTB4, SlBTB8, SlBTB21, SlBTB29, and SlBTB30) contained two Ank domains. Four SlBTBs (SlBTB6, SlBTB7, SlBTB18, and SlBTB19) consisted of one TAZ (Transcription Adaptor putative Zinc finger) domain, two (SlBTB14 and SlBTB15) had one Skp1 domain, and the SlBTB14 contained four tandem Arm (Armadillo/beta-catenin-like repeat) domains (Fig. 1). The BTB proteins in tomato consisted of 1–3 BTB domains, with most containing a combination of one BTB domain and other domains, such as MATH, Ank, TAZ, and Skp1.

Structure of the tomato BTB proteins. Numbers indicate amino acid position of the corresponding conserved domains. Scale bar represents 100 amino acids. Domains were identified with SMART (http://smart.embl-heidelberg.de/), pfam (http://pfam.xfam.org/), and InterProscan (http://www.ebi.ac.uk/interpro/search/sequence-search) programs
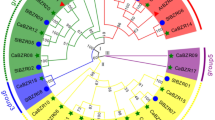
To explore the phylogenetic relationship and divergence of the BTB family in tomato, a phylogenetic tree was constructed using MEGA5 according to the aligned 38 BTB protein sequences. As shown in Fig. 2, the tomato BTB family was divided into four subfamilies based on the phylogenetic analysis. Group I comprised 24 BTB proteins, including potassium channel tetramerization domain-containing (KCTD) proteins (SlBTB23, SlBTB28, SlBTB36, and SlBTB37), TAZ (SlBTB6, SlBTB7, SlBTB18, and SlBTB19), IPR013089 Kelch-related (SlBTB11, SlBTB17, SlBTB32, and SlBTB33), MATH (SlBTB1, SlBTB2, SlBTB12, SlBTB26, and SlBTB34), NPR1-like protein (SlBTB4, SlBTB8, SlBTB29, and SlBTB30), and Arm (SlBTB20), according to the classification results of BTB domain proteins in 17 eukaryotes (Stogios et al. 2005). Group II comprised BTB-NPH3 proteins, SlBTB5, SlBTB9, and SlBTB13. Group III contained five BTB proteins, and Group IV consisted of the SKP1 (SlBTB14, SlBTB15, and SlBTB16) clade.

Phylogenetic analysis of the tomato BTB family. Trees were based on the protein sequence alignments with the full-length BTB proteins sequence. Different sharpness values of the circles indicate the different subfamily members according to sequence similarity annotation analysis, and red stars denote the BTB genes selected for expression. The phylogenetic tree was constructed using MEGA5 program using the neighbor-joining method at 1000 bootstrap replicates. (Color figure online)
Chromosomal localization, gene structure, and duplication
To determine the genomic distribution of tomato BTB genes, the genes were mapped onto the chromosomes of the published tomato genome (Fig. 3). BTB genes were distributed on all 12 chromosomes. Among them, only one BTB gene was distributed on chromosome 3 and 9; two on chromosomes 5 and 7; three on chromosomes 1, 2, 4, 8, and 12; four on chromosome 11; five on chromosome 10; and eight on chromosome 6. Among the tomato BTB genes, only eight were segmentally duplicated and one pair was arranged in tandem (SlBTB29 and SlBTB30) (Fig. 3).

Chromosome distribution and duplication events of tomato BTB genes. Chromosomal mapping was based on the physical position (Mb) in 12 tomato chromosomes. Chromosome number is indicated at the top of each bar. Positions of the tomato BTB genes in the chromosomes were obtained from Sol Genomics Network database (SGN, https://solgenomics.net/). Four paralogous gene pairs connected with solid dotted lines represent segmental duplication. Solid blocks indicate the centromere positions in chromosomes. Scale represents 10 Mb chromosomal distance
As shown in the Fig. 4, the tomato BTB gene exon–intron organizations were diverse. Most of the BTB genes contained 1–5 introns, except for SlBTB14, SlBTB15, and SlBTB16, which had no introns; and SlBTB3, SlBTB9, SlBTB20, SlBTB27, SlBTB28, and SlBTB31, which contained 11, 8, 18, 10, 11, and 18 introns, respectively. However, the members of a subfamily showed structural similarities. All BTB genes in the MATH group (SlBTB1, SlBTB2, SlBTB12, SlBTB26, and SlBTB34) showed a 4-exon/3-intron structure, and the SKP1-type BTB protein (SlBTB14, SlBTB15, and SlBTB16) had no intron.

Gene structure of the tomato BTB family generated from GSDS. The yellow block indicates the coding sequence (CDS), the blue block refers to upstream or downstream of the genes, the solid line represents the intron, and the red underline separate the gene family into four groups according to the phylogenetic analysis. Scale bar indicates DNA sequence length. (Color figure online)
Cis-elements in the promoters of tomato BTB genes
Cis-elements participate in gene regulation by interacting with their corresponding trans-regulatory factors; therefore, studying the putative cis-elements would provide valuable information regarding the tomato BTB genes expression. Hence, the promoter regions of all the BTB genes were retrieved and submitted to the PlantCARE database for cis-element identification (Supplementary Table S2). A total of 27 cis-elements were identified in more than 15 BTB genes (Tables 2 and S3). As expected, conventional promoter elements (TATA-box and CAAT-box) were detected in all SlBTBs promoters. The remaining 25 cis-acting elements can be divided into four groups. A total of 13 cis-elements were light-responsive, namely, Box 4, G-box, circadian, G-Box, Box I, TCT-motif, Sp1, I-box, ATCT-motif, GAG-motif, AE-box, ACE and GT1-motif. Four cis-elements were hormone-responsive, namely, CGTCA-motif, TGACG-motif, TCA-element, and ABRE. Six cis-elements function as stress-responsive elements: TC-rich repeats, HSE, ARE, MBS, W box and Box-W1. The fourth group had two cis-elements (Skn-1_motif and CAT-box), which are required for endosperm and meristem expression and were identified in 34 and 16 BTB gene promoters.
Expression profiles of BTB genes in tomato
To investigate the tissue-specific expression profiles of SlBTBs, transcriptome data were studied in ten tissues. As indicated in Fig. 5, most SlBTBs are constitutive and highly expressed in the tissues/stages, including SlBTB1, SlBTB2, SlBTB3, SlBTB4, SlBTB7, SlBTB9, SlBTB10, SlBTB12, SlBTB18, SlBTB28, SlBTB33 and SlBTB34. Conversely, the mRNAs of SlBTB5, SlBTB6, SlBTB8, SlBTB17, and SlBTB19 accumulated in the root and flower, but decreased during fruit development. Moreover, the SlBTB13, SlBTB20 and SlBTB24 were specifically expressed during fruit development. The raw read count in SlBTB14, SlBTB15, SlBTB16, SlBTB25, SlBTB36, SlBTB37, and SlBTB38 was 0, indicating low expression levels for the corresponding tissues/stages (Fig. 5).

Sources of samples were as follows: leaf, root, flower (Fl), flower bud, 1, 2, 3 cm fruit, mature green (MG) fruit, breaker (Br) fruit, and fruit at 10 days. Blank indicates no raw reads detected in the RNA-seq data
Heat map representation of BTB genes in various tissues/stages. The RNA-seq expression data of ten tissues were used to re-construct expression patterns of SlBTB genes.
The expression patterns of BTBs under multi-abiotic stresses between LA0716 and M82
LA0716 is tolerant to drought and salt stress, whereas the cultivated tomato M82 is sensitive to stress (Bolger et al. 2014). A previous study indicated the possible involvement of BTB genes in the drought response of tomatoes (Gong et al. 2010). To determine the expression differences of BTB genes between the wild LA0716 and cultivated tomato M82, cis-elements in the promoter region of each BTB gene were analyzed, as well as the expression patterns of the representative BTB genes under drought treatment (Supplementary Table S2, Fig. 6). Every BTB gene of LA0716 and M82 carried different cis-elements in their promoter regions (Supplementary Table S2). The major different cis-elements of the whole family were related to light, hormone, and stress responses. For instance, LTR and TATC-box elements were identified in the SpBTB3 promoter but were deficient in the SlBTB3 promoter of M82, where these elements are involved in low-temperature and GA responses, respectively.

Expression profiles of tomato BTB genes under different abiotic stresses and exogenous hormones in Solanum pennellii LA0716 (P) and S. lycopersicum cv. M82 (M). Numbers 1, 4, 6, 12, and 24 indicate the time (h) after treatments. Plants without stress at the same time served as controls. All samples were collected at the indicated time intervals from three biological replicates of each treatment. Error bars indicate ±SE of the means (n = 3)
Gene expression patterns can provide important clues for gene function; therefore, we performed quantitative RT-PCR to characterize the gene transcription profiles of the tomato BTB genes under different abiotic stresses, including cold, high salinity, dehydration drought, and oxidative stress. Describing the expression profiles of all tomato BTB genes is exhaustively difficult; therefore, only five members (SlBTB3, SlBTB5, SlBTB12, SlBTB18 and SlBTB31) covering four superfamilies of tomato BTB genes were assessed (Figs. 2, 6). As shown in Fig. 6, the SlBTB3 was induced by cold, high salinity, and dehydration drought stresses in M82 and LA0716, but displayed different expression patterns between LA0716 and M82. The increase of SlBTB3 was dramatic and slight in LA0716 and M82, respectively, under cold, high salinity, and dehydration drought stresses. However, under MV oxidative stress, the increase of SlBTB3 was slight and dramatic in LA0716 and M82, respectively. The expression levels of SlBTB5 and SlBTB31 were similar under abiotic stresses between LA0716 and M82. Moreover, SlBTB5 and SlBTB31 levels decreased in M82 but dramatically increased in LA0716 under cold, high salinity, and dehydration drought stresses. Under MV stress, the SlBTB5 and SlBTB31 levels showed no significant changes after 1, 4, 12 and 24 h in both LA0716 and M82. However, SlBTB5 and SlBTB31 levels dramatically increased in both genotypes after 6 h of treatment. The expression of SlBTB12 was inhibited by cold, high salinity, and dehydration drought stresses in M82 and LA0716, but displayed different expression patterns between LA0716 and M82. The SlBTB12 level was slightly decreased in LA0716 but was dramatically increased in M82 under cold, high salinity, and dehydration drought stresses. However, under MV oxidative stress, the SlBTB12 remained constant at 1, 4, 12 and 24 h in both LA0716 and M82, and dramatically increased in LA0716 and M82 after 6 h of treatment. The increase in SlBTB18 levels were dramatic and slight in LA0716 and M82 under cold and high salinity stresses, but the degrees of increase in SlBTB18 levels reversed in LA0716 and M82 under dehydration drought stress. Under MV oxidative stress, the SlBTB18 levels were constant at 1 and 12 h in both LA0716 and M82, but dramatically increased in M82 upon 4 and 6 h treatments and significantly increased in LA0716 after 24 h of treatment. All results indicated that BTB gene expressions are responsive to multi-abiotic stresses, but displayed different expression patterns between the cultivar species M82 and the wild species LA0716.
Expression of BTB genes in response to exogenous hormones
Hormones play vital roles in plant growth and development. To examine the effects of various hormones on the expression of the tomato BTB gene family, qRT-PCR was used to analyze the transcriptional levels of all BTB genes under GA, ABA, SA, and Eth treatments (Fig. 6).
As shown in the Fig. 5, the expression of SlBTB3 was induced by GA, ABA, SA and Eth in M82 and LA0716. The increase in SlBTB3 levels was dramatic and slight in LA0716 and M82, respectively, under ABA and GA treatment, but their degrees of increase reversed in the two genotypes under SA and Eth treatments. Moreover, the expression levels of SlBTB5 and SlBTB31 were similar in response to exogenous hormones between LA0716 and M82. The expression levels of SlBTB5 and SlBTB31 decreased in M82 but dramatically increased in LA0716 under GA, ABA, and Eth treatments. Under SA treatment, both SlBTB5 and SlBTB31 showed no significant changes except for the dramatic up-regulation of SlBTB5 after 6 h in the M82, whereas SlBTB31 was down-regulated after the 12 h in LA0716. The SlBTB12 was inhibited by GA and ABA in all time intervals and was induced by SA and Eth after 6 h of treatment in LA0716; however, it displayed no significant difference after GA, SA, and Eth treatments and was up-regulated to approximately threefold in M82. The SlBTB18 was induced by GA, ABA, and SA but was inhibited by Eth in both M82 and LA0716, with different response times between LA0716 and M82. For instance, after 1 h treatment of ABA, SlBTB18 rapidly increased by approximately 42-fold in M82 but did not show any response in LA0716. However, the SlBTB18 levels dramatically increased by 3.8-fold in LA0716 and decreased by 0.3-fold in M82 after 1 h of SA treatment. These results indicated that BTB genes respond to hormones, but display different expression patterns between M82 and LA0716.
Discussions
BTB proteins are widespread in various species and not conserved proteins
The present study characterized the structure, phylogenetic relationship, duplication events, and expression profiles of the BTB gene family in tomatoes. We found 38 and 34 BTB genes in cultivated tomato M82 and in wild tomato species (S. pennellii), respectively, and the differences in the number of BTB genes is caused by long-term evolution. The propensity of gene loss during evolution seems to be a direct reflection of its biological importance (Krylov et al. 2003); therefore, the genes lost in LA0716 (SlBTB17, SlBTB18, SlBTB19 and SlBTB20) may have biological functions in M82.
As reported, the numbers of genes belonging to the BTB family vary greatly among species. A total of 183 BTB proteins were found in H. sapiens, 178 in the C. elegans, 195 in the M. musculus, 77 in the A. thaliana, and 5 in Saccharomyces cerevisiae (Stogios et al. 2005). The difference in the number of BTB genes was large and the BTB gene composition varied among species. In our study, all BTB genes isolated from the tomato contained at least one BTB/POZ domain in the protein structure (Fig. 1), which is consistent with previous reports (Bardwell and Treisman 1994; Zollman et al. 1994). With the exception of the BTB/POZ domain, the BTB protein consisted of several other domains, including the MATH domain, Ankyrin repeat, NPR1/NIM1-like defense protein C-terminal, the NPH3 family, TAZ zinc finger, BACK, Skp1, and Arm. However, the enrichment of BTB-ZF (Zinc finger) proteins in vertebrates cannot be found in tomatoes and is consistent with the absence of BTB-ZF in Arabidopsis (Stogios et al. 2005). Through searching the SGN genome annotation database, we found 28 and 17 BTB/POZ genes present in the genomes of pepper and potato, respectively (data not shown), which are similar with the low number (38) of BTB genes in the tomato, as opposed to 77 in A. thaliana. Moreover, the total number of BTB protein genes in the tomato genome and the other four Solanaceae species surveyed were not similar, indicating that expansion has occurred in the BTB protein gene family during Solanaceae evolution. Similar findings showed that BTB protein genes have undergone a lineage-specific expansion in vertebrates compared with the fruit fly and malaria mosquito (Stogios et al. 2005).
BTB genes may play various functions in plants
BTB proteins are present in poxviruses and many eukaryotes and have diverse functions, ranging from transcriptional regulation and chromatin remodeling to protein degradation and cytoskeletal regulation. Functionally, the BTB domain acts as a protein–protein interaction module capable of both self-association and interaction with non-BTB proteins (Stogios et al. 2005). Two prominent mechanisms mainly govern BTB protein functions (Perez-Torrado et al. 2006): BTB domain-based protein–protein interactions (Bonchuk et al. 2011; Weber et al. 2005) and transcriptional regulation by DNA binding domains, such as zinc finger domains (Cho et al. 2012; Korutla et al. 2005; Liu et al. 2011; Qi et al. 2006). In this study, several other types of functional domains, including KCTD, MATH, Kelch, ANK, NPR1, NPH3, TAZ, BACK, Skp1, and Arm, were present in some tomato BTB proteins (Fig. 1). These additional domains in BTB proteins confer diverse functions, suggesting the multiple roles of BTB protein genes in plants.
The KCTD contain proteins that are involved in fundamental biological processes, including protein ubiquitination and degradation, suppression of proliferation or transcription, and human genetic disease risk (Smaldone et al. 2016). The BTB-containing KCTD proteins represent an emerging class of proteins involved in fundamental physiopathological processes (Liu et al. 2013; Skoblov et al. 2013). The SlBTB23, SlBTB28, SlBTB36, and SlBTB37 from the KCTD family have conserved N-terminal regions (Supplementary Fig. S1), indicating the involvement of the tomato KCTD BTB proteins in physiopathological ubiquitination.
MATH domains share a conserved region of approximately 180 residues (Sunnerhagen et al. 2002) and are involved in plant responses to abiotic stress (Kushwaha et al. 2016). The BTB conjunction with the MATH domain protein, MATH-BTB, is important in ABA signaling and other developmental processes (Lechner et al. 2011). The AtCUL3a and AtCUL3b in Arabidopsis can assemble with BTB/POZ-MATH proteins to form functional E3 ligases (Weber et al. 2005). The MATH-BTB gene MBA1 in maize forms homodimers, interacts with the E3 ubiquitin ligase component, Cullin 3a (CUL3a), and regulates spindle length and nuclei identity (Juranic et al. 2012). The BTB/POZ-MATH proteins can interact with members of the ERF/AP2 transcription factor family and respond to multi-abiotic stresses (Weber and Hellmann 2009).
NPR1 contains two distinct protein–protein interaction motifs: ankyrin repeats and a BTB/POZ domain (Boyle et al. 2009). The ankyrin repeat domain consists of 33 residues that are repeated in tandem that mediates protein–protein interactions (Michaely and Bennett 1992; Mosavi et al. 2002). The genes of the ANK protein family in tomato respond to biotic and abiotic stresses and are crucial for plant growth and development (Yuan et al. 2013). NPR1 is a key transcriptional co-regulator of immunity and physiology in many plant species (Caldwell and Michelmore 2009; Mukhtar et al. 2009; Withers and Dong 2016). The tomato NPR1 SlBOP genes (SlBTB8, SlBTB29 and SlBTB30) interacted with the TERMINATING FLOWER (TMF) protein to control the inflorescent architecture (Xu et al. 2016). The SlBTB4, SlBTB8, SlBTB29, and SlBTB30 are classified under the NPR1 subfamily and are highly similar to each other (Supplementary Fig. S1), implying the possible importance of the classified SlBTB4 in plant development.
The NPH3 is apparently specific to higher plants and acts as a key component in blue light-mediated phototropic responses (Motchoulski and Liscum 1999). Recently, NPH3 has been revealed to interact with the blue light receptor phot1 and functions as a substrate adapter in a CULLIN3-based E3 ubiquitin ligase (Roberts et al. 2011). In our study, SlBTB5, SlBTB9, and SlBTB13 belong to the NPH3 subfamily and, thus, may function during blue light-mediated phototropic responses.
The BTB and TAZ domain (BT) proteins in Arabidopsis have divergent functions. In Arabidopsis, five members of the BT protein subfamily (BT1–BT5) have a typical domain structure only observed in land plants. The BT family members perform a non-essential function; or, alternatively, that functional redundancy exists among BT family members during Arabidopsis development (Robert et al. 2009). BT2 has a negative role in ABA response and acts as a central component of an interconnected signaling network that detects and responds to nutrients, stresses, and hormones (Mandadi et al. 2009). The BT gene family members (BT1 and BT2) in rice act as conserved negative regulators of nitrate uptake genes (Araus et al. 2016). As shown in Supplementary Fig. S1, the amino acid sequences of SlBTB6, SlBTB7, SlBTB18, and SlBTB19 contain a conserved TAZ domain in the C-terminus, indicating that these genes probably have similar functions in the plant.
The BACK domain in the majority of proteins contains both the BTB domain and Kelch repeats. The Kelch repeats in the BACK proteins form a beta propeller that interacts with actin filaments (Stogios and Prive 2004). The BACK constitutes a novel conserved domain, which is an adaptor and/or a chaperone in the ubiquitin–proteasome protein-degradation pathway and functions during the assembly of Cullin3-based E3 ligase complexes (Canning et al. 2013; Friedman et al. 2009). The SlBTB11, SlBTB17, SlBTB22, and SlBTB33 have a C-terminal Kelch-conserved domain and share high similarity with each other (Supplementary Fig. S1); therefore, they may be involved in plant ubiquitin degradation.
As a critical component of the Cul1-based SCF complex, Skp1 forms the structural link between Cul1 and substrate recognition proteins (Stebbins et al. 1999), which serve as adaptors that simultaneously binds sequences near the N-terminus of Cul1 (cullin-RING ligase 1) and the F-box motif of an F-box protein during ubiquitin degradation (Zhuang et al. 2009). The tomato SlBTB14, SlBTB15, and SlBTB16 contain the Skp1 structure (Fig. 1) and consequently has possible functions in the ubiquitination pathway.
The SlBTB20 contains four tandem Armadillo (Arm) repeats in its N-terminus and a BTB domain located in its C-terminus. The Arm is a 42-amino acid protein–protein interaction motif that was first identified in the Drosophila segment polarity gene armadillo (Peifer et al. 1994; Riggleman et al. 1989). Animal Arm-repeat proteins function in various processes, including intracellular signaling and cytoskeletal regulation (Hatzfeld 1999). A subset of these proteins is conserved across eukaryotic kingdoms. Some Arm-repeat proteins in higher plants function in intracellular signaling, similar to their mammalian counterparts, and have other novel functions (Coates 2003). The Arabidopsis ARIA (armadillo repeat protein interacting with ABF2) protein contains armadillo repeats and a BTB/POZ domain and affects ABA-regulated gene expression, seedling growth, ABA sensitivity, and stress tolerance of plants (Kim and Choi 2006), whereas the ABAP1 (Armadillo BTB Arabidopsis protein) participates in a negative feedback loop that regulates mitotic DNA replication during plant leaf development by repressing gene transcription (Masuda et al. 2008). These functions help in the exploration of Arm-BTB protein SlBTB20 function in tomato.
BTB domain-containing proteins are present in the proteins of poxvirus and other eukaryotes, including zebrafish, mouse, and human (Chaharbakhshi and Jemc 2016); however, plant BTBs, especially in model plants, need further studies. In this study, five representative BTB genes responded to abiotic stresses (cold, salt, drought, and oxidative stress) and hormones (GA, ABA, SA, and Eth) and show different expression patterns in LA0716 and M82 (Fig. 6). LA0716 is a stress-tolerant wild tomato accession (Bolger et al. 2014), whereas the cultivated tomato M82 is stress-sensitive. These five genes possessed at least one of the stress response cis-elements (HSE, TC-rich, MBS, and ARE) in their promoter regions, thereby implying their potential roles in stress response. These BTB genes might be involved in hormone signaling as transcriptional regulators that modulate plant tolerance responses to abiotic stress.
Conclusions
In this study, 38 SlBTB genes in the tomato genome were comprehensively analyzed, covering the gene structure, subcellular localization predication, phylogenetic and duplication analyses, and cis-regulatory elements prediction. Moreover, the BTB subclass domain were classified, and the transcription of some SlBTBs were revealed to be induced by hormones, abiotic stress, or differential expressions between LA0716 and M82, which possibly indicate the vital functions of these BTB genes in physiological activities. Data from this present study should be considered in future studies for genetically improving agronomic traits and/or stress tolerance in tomatoes and other Solanaceae plants.
References
Ahmad KF, Melnick A, Lax S, Bouchard D, Liu J, Kiang CL, Mayer S, Takahashi S, Licht JD, Prive GG (2003) Mechanism of SMRT corepressor recruitment by the BCL6 BTB domain. Mol Cell 12:1551–1564
Araus V, Vidal EA, Puelma T, Alamos S, Mieulet D, Guiderdoni E, Gutierrez RA (2016) Members of BTB gene family of scaffold proteins suppress nitrate uptake and nitrogen use efficiency. Plant Physiol 171:1523–1532
Bardwell VJ, Treisman R (1994) The POZ domain: a conserved protein–protein interaction motif. Genes Dev 8:1664–1677
Bolger A, Scossa F, Bolger ME, Lanz C, Maumus F, Tohge T, Quesneville H, Alseekh S, Sørensen I, Lichtenstein G (2014) The genome of the stress-tolerant wild tomato species Solanum pennellii. Nat Genet 46:1034–1038
Bonchuk A, Denisov S, Georgiev P, Maksimenko O (2011) Drosophila BTB/POZ domains of “ttk group” can form multimers and selectively interact with each other. J Mol Biol 412:423–436
Bostan H, Chiusano ML (2015) NexGenEx-Tom: a gene expression platform to investigate the functionalities of the tomato genome. BMC Plant Biol 15:48
Boyle P, Le Su E, Rochon A, Shearer HL, Murmu J, Chu JY, Fobert PR, Despres C (2009) The BTB/POZ domain of the Arabidopsis disease resistance protein NPR1 interacts with the repression domain of TGA2 to negate its function. Plant Cell 21:3700–3713
Caldwell KS, Michelmore RW (2009) Arabidopsis thaliana genes encoding defense signaling and recognition proteins exhibit contrasting evolutionary dynamics. Genetics 181:671–684
Canning P, Cooper CD, Krojer T, Murray JW, Pike AC, Chaikuad A, Keates T, Thangaratnarajah C, Hojzan V, Marsden BD (2013) Structural basis for Cul3 protein assembly with the BTB-Kelch family of E3 ubiquitin ligases. J Biol Chem 288:7803–7814
Chaharbakhshi E, Jemc JC (2016) Broad-complex, tramtrack, and bric-a-brac (BTB) proteins: critical regulators of development. Genesis 10:505–518
Cheng D, Qian W, Meng M, Wang Y, Peng J, Xia Q (2014) Identification and expression profiling of the BTB domain-containing protein gene family in the silkworm, bombyx mori. Int J Genom 2014:865065
Chevrier S, Emslie D, Shi W, Kratina T, Wellard C, Karnowski A, Erikci E, Smyth GK, Chowdhury K, Tarlinton D, Corcoran LM (2014) The BTB-ZF transcription factor Zbtb20 is driven by Irf4 to promote plasma cell differentiation and longevity. J Exp Med 211:827–840
Cho JH, Kim MJ, Kim KJ, Kim JR (2012) POZ/BTB and AT-hook-containing zinc finger protein 1 (PATZ1) inhibits endothelial cell senescence through a p53 dependent pathway. Cell Death Differ 19:703–712
Coates JC (2003) Armadillo repeat proteins: beyond the animal kingdom. Trends Cell Biol 13:463–471
Consortium TG (2012) The tomato genome sequence provides insights into fleshy fruit evolution. Nature 485:635–641
Friedman JS, Ray JW, Waseem N, Johnson K, Brooks MJ, Hugosson T, Breuer D, Branham KE, Krauth DS, Bowne SJ (2009) Mutations in a BTB-Kelch protein, KLHL7, cause autosomal-dominant retinitis pigmentosa. Am J Hum Genet 84:792–800
Furukawa M, He YJ, Borchers C, Xiong Y (2003) Targeting of protein ubiquitination by BTB-Cullin 3-Roc1 ubiquitin ligases. Nat Cell Biol 5:1001–1007
Geyer R, Wee S, Anderson S, Yates J, Wolf DA (2003) BTB/POZ domain proteins are putative substrate adaptors for cullin 3 ubiquitin ligases. Mol Cell 12:783–790
Gong P, Zhang J, Li H, Yang C, Zhang C, Zhang X, Khurram Z, Zhang Y, Wang T, Fei Z, Ye Z (2010) Transcriptional profiles of drought-responsive genes in modulating transcription signal transduction, and biochemical pathways in tomato. J Exp Bot 61:3563–3575
Guo A, Zhu Q, Chen X, Luo J (2007) GSDS: a gene structure display server. Yi Chuan 29:1023–1026
Hatzfeld M (1999) The armadillo family of structural proteins. Int Rev Cytol 186:179–224
Hou X, Xie K, Yao J, Qi Z, Xiong L (2009) A homolog of human ski-interacting protein in rice positively regulates cell viability and stress tolerance. Proc Natl Acad Sci USA 106:6410–6415
Ito H, Sato K, Yamamoto D (2013) Sex-switching of the Drosophila brain by two antagonistic chromatin factors. Fly (Austin) 7:87–91
Juranic M, Srilunchang KO, Krohn NG, Leljak-Levanic D, Sprunck S, Dresselhaus T (2012) Germline-specific MATH-BTB substrate adaptor MAB1 regulates spindle length and nuclei identity in maize. Plant Cell 24:4974–4991
Kim SY, Choi H-i (2006) Nucleic acid molecule encoding an armadillo repeat protein, aria and a method utilizing aria to generate salt tolerant plants. US Patent No. 7049482
Kim H, Kim SH, Seo DH, Chung S, Kim SW, Lee JS, Kim WT, Lee JH (2016) Aba-hypersensitive BTB/POZ protein 1 functions as a negative regulator in ABA-mediated inhibition of germination in Arabidopsis. Plant Mol Biol 90:303–315
Ko JH, Son W, Bae GY, Kang JH, Oh W, Yoo OJ (2006) A new hepatocytic isoform of PLZF lacking the BTB domain interacts with ATP7B, the Wilson disease protein, and positively regulates ERK signal transduction. J Cell Biochem 99:719–734
Korutla L, Wang PJ, Mackler SA (2005) The POZ/BTB protein NAC1 interacts with two different histone deacetylases in neuronal-like cultures. J Neurochem 94:786–793
Krylov DM, Wolf YI, Rogozin IB, Koonin EV (2003) Gene loss, protein sequence divergence, gene dispensability, expression level, and interactivity are correlated in eukaryotic evolution. Genome Res 13:2229–2235
Kushwaha HR, Joshi R, Pareek A, Singla-Pareek SL (2016) MATH-domain family shows response toward abiotic stress in Arabidopsis and rice. Front Plant Sci 7:923
Lechner E, Leonhardt N, Eisler H, Parmentier Y, Alioua M, Jacquet H, Leung J, Genschik P (2011) MATH/BTB CRL3 receptors target the homeodomain-leucine zipper ATHB6 to modulate abscisic acid signaling. Dev Cell 21:1116–1128
Lim JH (2014) Zinc finger and BTB domain-containing protein 3 is essential for the growth of cancer cells. BMB Rep 47:405–410
Liu Q, Yao F, Wang M, Zhou B, Cheng H, Wang W, Jin L, Lin Q, Wang JC (2011) Novel human BTB/POZ domain-containing zinc finger protein ZBTB1 inhibits transcriptional activities of CRE. Mol Cell Biochem 357:405–414
Liu Z, Xiang Y, Sun G (2013) The KCTD family of proteins: structure, function, disease relevance. Cell Biosci 3:45
Mandadi KK, Misra A, Ren S, McKnight TD (2009) BT2, a BTB protein, mediates multiple responses to nutrients, stresses, and hormones in Arabidopsis. Plant Physiol 150:1930–1939
Masuda HP, Cabral LM, De Veylder L, Tanurdzic M, de Almeida Engler J, Geelen D, Inzé D, Martienssen RA, Ferreira PC, Hemerly AS (2008) ABAP1 is a novel plant Armadillo BTB protein involved in DNA replication and transcription. EMBO J 27:2746–2756
Melnick A, Ahmad KF, Arai S, Polinger A, Ball H, Borden KL, Carlile GW, Prive GG, Licht JD (2000) In-depth mutational analysis of the promyelocytic leukemia zinc finger BTB/POZ domain reveals motifs and residues required for biological and transcriptional functions. Mol Cell Biol 20:6550–6567
Michaely P, Bennett V (1992) The ANK repeat: a ubiquitous motif involved in macromolecular recognition. Trends Cell Biol 2:127–129
Mittler R (2006) Abiotic stress, the field environment and stress combination. Trends Plant Sci 11:15–19
Mosavi LK, Minor DL Jr, Peng ZY (2002) Consensus-derived structural determinants of the ankyrin repeat motif. Proc Natl Acad Sci USA 99:16029–16034
Motchoulski A, Liscum E (1999) Arabidopsis NPH3: a NPH1 photoreceptor-interacting protein essential for phototropism. Science 286:961–964
Mukhtar MS, Nishimura MT, Dangl J (2009) NPR1 in plant defense: it’s not over’til it’s turned over. Cell 137:804–806
Peifer M, Berg S, Reynolds AB (1994) A repeating amino acid motif shared by proteins with diverse cellular roles. Cell 76:789–791
Perez-Torrado R, Yamada D, Defossez PA (2006) Born to bind: the BTB protein–protein interaction domain. Bioessays 28:1194–1202
Pintard L, Willis JH, Willems A, Johnson JL, Srayko M, Kurz T, Glaser S, Mains PE, Tyers M, Bowerman B, Peter M (2003) The BTB protein MEL-26 is a substrate-specific adaptor of the CUL-3 ubiquitin-ligase. Nature 425:311–316
Qi J, Zhang X, Zhang HK, Yang HM, Zhou YB, Han ZG (2006) ZBTB34, a novel human BTB/POZ zinc finger protein, is a potential transcriptional repressor. Mol Cell Biochem 290:159–167
Riggleman B, Wieschaus E, Schedl P (1989) Molecular analysis of the armadillo locus: uniformly distributed transcripts and a protein with novel internal repeats are associated with a Drosophila segment polarity gene. Genes Dev 3:96–113
Robert HS, Quint A, Brand D, Vivian-Smith A, Offringa R (2009) BTB and TAZ domain scaffold proteins perform a crucial function in Arabidopsis development. Plant J 58:109–121
Roberts D, Pedmale UV, Morrow J, Sachdev S, Lechner E, Tang X, Zheng N, Hannink M, Genschik P, Liscum E (2011) Modulation of phototropic responsiveness in Arabidopsis through ubiquitination of phototropin 1 by the CUL3-ring E3 ubiquitin ligase CRL3NPH3. The Plant Cell 23:3627–3640
Robinson DN, Cooley L (1997) Drosophila kelch is an oligomeric ring canal actin organizer. J Cell Biol 138:799–810
Siggs OM, Li X, Xia Y, Beutler B (2012) ZBTB1 is a determinant of lymphoid development. J Exp Med 209:19–27
Skoblov M, Marakhonov A, Marakasova E, Guskova A, Chandhoke V, Birerdinc A, Baranova A (2013) Protein partners of KCTD proteins provide insights about their functional roles in cell differentiation and vertebrate development. Bioessays 35:586–596
Smaldone G, Pirone L, Pedone E, Marlovits T, Vitagliano L, Ciccarelli L (2016) The BTB domains of the potassium channel tetramerization domain proteins prevalently assume pentameric states. FEBS Lett 590:1663–1671
Stebbins CE, Kaelin WG Jr, Pavletich NP (1999) Structure of the VHL-ElonginC-ElonginB complex: implications for VHL tumor suppressor function. Science 284:455–461
Stogios PJ, Prive GG (2004) The back domain in BTB-kelch proteins. Trends Biochem Sci 29:634–637
Stogios PJ, Downs GS, Jauhal JJ, Nandra SK, Prive GG (2005) Sequence and structural analysis of BTB domain proteins. Genome Biol 6:R82
Sunnerhagen M, Pursglove S, Fladvad M (2002) The new MATH: homology suggests shared binding surfaces in meprin tetramers and TRAF trimers. FEBS Lett 530:1–3
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28:2731–2739
Tomato Genome C (2012) The tomato genome sequence provides insights into fleshy fruit evolution. Nature 485:635–641
Weber H, Hellmann H (2009) Arabidopsis thaliana BTB/POZ-MATH proteins interact with members of the ERF/AP2 transcription factor family. FEBS J 276:6624–6635
Weber H, Bernhardt A, Dieterle M, Hano P, Mutlu A, Estelle M, Genschik P, Hellmann H (2005) Arabidopsis AtCUL3a and AtCUL3b form complexes with members of the BTB/POZ-MATH protein family. Plant Physiol 137:83–93
Weyers JJ, Milutinovich AB, Takeda Y, Jemc JC, Van Doren M (2011) A genetic screen for mutations affecting gonad formation in Drosophila reveals a role for the slit/robo pathway. Dev Biol 353:217–228
Wilkins A, Ping Q, Carpenter CL (2004) RhoBTB2 is a substrate of the mammalian Cul3 ubiquitin ligase complex. Genes Dev 18:856–861
Withers J, Dong X (2016) Posttranslational modifications of NPR1: A single protein playing multiple roles in plant immunity and physiology. PLoS Pathog 12:e1005707
Xu L, Wei Y, Reboul J, Vaglio P, Shin TH, Vidal M, Elledge SJ, Harper JW (2003) BTB proteins are substrate-specific adaptors in an SCF-like modular ubiquitin ligase containing CUL-3. Nature 425:316–321
Xu C, Park SJ, Van Eck J, Lippman ZB (2016) Control of inflorescence architecture in tomato by BTB/POZ transcriptional regulators. Genes Dev 30:2048–2061
Yuan X, Zhang S, Qing X, Sun M, Liu S, Su H, Shu H, Li X (2013) Superfamily of ankyrin repeat proteins in tomato. Gene 523:126–136
Zhuang M, Calabrese MF, Liu J, Waddell MB, Nourse A, Hammel M, Miller DJ, Walden H, Duda DM, Seyedin SN (2009) Structures of SPOP-substrate complexes: insights into molecular architectures of BTB-Cul3 ubiquitin ligases. Mol Cell 36:39–50
Zollman S, Godt D, Prive GG, Couderc JL, Laski FA (1994) The BTB domain, found primarily in zinc finger proteins, defines an evolutionarily conserved family that includes several developmentally regulated genes in Drosophila. Proc Natl Acad Sci USA 91:10717–10721
Acknowledgements
This work was supported by grants from the Basic and Frontier Research Project of Chongqing (No. cstc2015jcyjA80030), The National Natural Science Foundation of China (Nos. 31301779 and 31401884), Natural Science Foundation of Youth in Jiangsu Province (No. BK20140739).
Author information
Authors and Affiliations
Contributions
The work presented here was carried out in collaboration between all authors. JL and XZ conceived the study; XS and WY performed the experiments, JL collected and analyzed the data; CS, YP and YW provided reagents and tools; JL written the manuscript; all authors read and approved of the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
All authors declare that they have no conflict of interest. Jinhua Li declares that he has no conflict of interest. Xiaoxing Su declares that she has no conflict of interest. Yinlei Wang declares that he has no conflict of interest. Wei Yang declares that he has no conflict of interest. Yu Pan declares that she has no conflict of interest. Chenggang Su declares that he has no conflict of interest. Xingguo Zhang declares that he has no conflict of interest.
Research involving human and animal participants
This article does not contain any studies with human participants or animals performed by any of the authors.
Electronic supplementary material
Below is the link to the electronic supplementary material.
13258_2017_604_MOESM3_ESM.xlsx
Table S2 The differences about the cis-elements in the promoter sequence of BTB genes between S. pennellii and M82 (XLSX 194 KB)
Rights and permissions
About this article

Cite this article
Li, J., Su, X., Wang, Y. et al. Genome-wide identification and expression analysis of the BTB domain-containing protein gene family in tomato. Genes Genom 40, 1–15 (2018). https://doi.org/10.1007/s13258-017-0604-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13258-017-0604-x