
Abstract
In the present study, the diversity of 11 Thymus species was assessed using molecular and morphological markers. Essential oil content and morphological traits were also investigated during two agronomic years. The result of the analysis of variance showed considerable differences among morphological traits. In the first and second years, the essential oil content of the species varied from 0.63 to 1.94% and 0.86 to 2.34%, respectively. T. vulgaris and T. migricus showed the highest essential oil content in two agronomic years. In this research, nine ISSR primers were also used to amplify 151 polymorphic bands in 77 accessions belonging to 11 Thymus species. Cluster and principal component (PCA) analyses classified the species in three major groups. Among the species, T. vulgaris and T. fedtschenkoi presented relatively higher genetic distance in comparison with other species. Analysis of molecular variance (AMOVA) revealed that 72.34% of the total variation was belonged to within-species variation, while 27.66% was associated among the species. High gene flow (Nm = 1.11) and genetic differentiation (Gst = 0.31) were also observed among the species. T. transcaspicus exhibited the highest genetic variation (0.19), polymorphism % (57.69%), and Shannon index (0.29). The STRUCTURE analysis also showed a high admixture of Thymus species that might be originated from a high rate of natural hybridization. Finally, based on molecular and morphological information, T. vulgaris and T. carmanicus can be suggested as good candidate species for further breeding programs in Thymus species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Thyme (Thymus spp.) is one of the most important medicinal plants of the Lamiaceae family with more than 200 species (Tohidi et al. 2019). Mediterranean regions have been assumed as the center of origin of this genus (Ghasemi Pirbalouti et al. 2011). In Iranian flora, 14 Thymus species have been found in which four of them including, T. daenensis, T. carmanicus, and T. trautvetteri, T. persicus are endemic (Rahimmalek et al. 2009; Talebi et al. 2015). Thyme, a perennial herb, and aromatic plant has always been widely used in food and pharmaceutical industries due to its valuable components (Trindade et al. 2019). Essential oils are important aromatic constituents which are derived from different part of plants such as flowers, buds, seeds, leaves, fruits, and roots (Eirini et al. 2017). From an industrial point of view, thyme has been considered as an antibacterial and antifungal plant with different applications in food industries (Rasooli et al. 2006; Ghasemi Pirbalouti et al. 2011; Mohammadi et al. 2014). In folk medicine, Thymus species are widely used due to their carminative, anti-inflammatory, antispasmodic, and digestive properties. So, the species of the genus Thymus are characterized as a biological source by having an excellent richness in essential oil (Rustaiee et al. 2013).
The current geographical distribution of plant taxa can be attributed to the present, as well as historical factors leading in the genetic structure and phylogenetic patterns (Tohidi et al. 2020). Genetic variation is a prerequisite for further breeding programs. The amount and separation of genetic variation among and within populations are also related to life history characteristics and environmental factors (Rahimmalek et al. 2009; Fadaei Heidari et al. 2016). Knowledge of genetic diversity among and within species can provide new information regarding the evolutionary process as well as new insights for conservation management of the plant species. Interspecies genetic variations lead to some phenotypical changes. In this respect, knowledge of interspecies genetic variations may also be beneficial to manage the extinction risks brought about by different intrinsic or extrinsic factors such as inbreeding and environmental factors (Lu et al. 2018). So, evaluating of inter- and intraspecies genetic variation can also be useful for selecting further breeding programs.
Nowadays, molecular markers can provide reliable information in comparison to morphological characteristics. Among the molecular markers, ISSR (inter simple sequence repeat) markers have widely been applied because of high reproducibility, high polymorphism, and simple use. ISSR marker amplifies the sequences between simple sequence repeats (Fadaei Heidari et al. 2016; Verma et al. 2017). ISSR is a PCR-based marker with high polymorphism and affordability that made it a strong tool for genetic diversity studies in plant species (Gupta et al. 1996). ISSR have been efficiently applied for genetic diversity analysis of T. caramanicus (Hadian et al. 2014) and T. daenensis (Rahimmalek et al. 2009) natural populations.
Thymus is one of the most important medicinal plant genera worldwide, and most of the research has been focused on its secondary metabolites (Tohidi et al. 2017). However, there are some reports regarding the genetic diversity of some Thymus species in different countries including T. deanaesis (Rahimmalek et al. 2009; Talebi et al. 2015), T. caespititius (Trindade et al. 2008, 2009), T. kotschianus (Yousefi et al. 2015), T. caramanicus (Hadian et al. 2014) and T. sibthorpii (Abraham et al. 2018). Most of the previous researches used one molecular marker, one or limited studied species. However, comprehensive research has not yet been performed to assess the inter- and intraspecific variation of Thymus species. In Iran, 14 Thymus species have been reported in which in the present research for the first time, inter and intraspecies variation of 11 species viz. Thymus migricus Klokov & Des.-Shost., T. fallax Fisch. & C. A. Mey., T. serpyllum L., T. trautvetteri Klokov & Desj.-Shost., T. transcaspicus Klokov, T. carmanicus Jalas, T. fedtschenkoi Ronneger, T. daenensis Celak subsp. daenensis, T. pubescens Boiss. & Kotschy ex Celak, T. kotschyanus Boiss. & Hohen., and T. vulgaris L. in two years was evaluated using molecular, morphological, and essential oil features.
The aims of the present research were (1) to assess the genetic variation among and within 11 Thymus species using ISSR markers; (2) to evaluate the morphological characteristics and essential oil content of studied species in two agronomic years.
Materials and methods
Plant materials and field experiment
The seeds of 11 Thymus species comprising T. daenensis, T. migricus, T. vulgaris, T. fallax, T. trautvetteri, T. serpyllum, T. transcaspicus, T. pubescens, T. fedtschenkoi, T. kotschyanus, T. carmanicus, were collected from gene bank of Research Institute of Forests and Rangeland, Tehran. The species were identified by Dr. Mozaffarian (Research Institute of Forests and Rangeland, Tehran), according to Flora Iranica (Rechinger 1963). The species codes and the detailed information of the species are exhibited in Table 1. A randomized block design (RCBD) with three replicates was used as a statistical design.
The study was performed at Isfahan University of Technology experimental farm. The area had a clay loam soil (pH 7.5) with an average bulk density of 1.48 g/cm3 in the top 60-cm layer of the soil surface. The mean annual precipitation and temperature were 140 mm and 15 °C, respectively. Seeds were planted with 35 cm distance on the rows and 50 cm space between the rows at 0.5 cm depth in a 2 × 1 m2plots. The aerial parts of studied Thymus species were collected at the flowering period in the two agronomic years of 2018 and 2019.
DNA extraction
Young leaves in full flowering stage were used to extract for DNA using the CTAB method with minor modifications (Murry and Thompson 1980). NanoDrop evaluated the quantity and quality of DNA at the ratio of OD260/OD280. Suitable DNA concentration was observed between the ratios of 1.6–1.8. The quality of DNA was also investigated by agarose gel in which the sharped bands without any smearing was useful to select DNAs with higher quality.
ISSR analysis
Among 13 ISSR stock primers, nine produced high numbers of repeatable bands. The obtained information of the ISSR primers is illustrated in Table 2. Amplification reactions were conducted using a thermocycler (Bio-Rad) in a total volume of 15 µL, as described previously (Fadaei Heidari et al. 2016). The optimum annealing temperature was determined according to the melting temperature for each primer. The PCR was performed under the following operation program: 2 min of initial denaturation (94 °C) followed by 40 cycles of 1 min at 94 °C; 1 min at the specific annealing temperature; extension at 72 °C for 2 min; and final extension step of 10 min at 72 °C. DNA amplified fragments were separated in a 1.2% agarose gel in 1 × TBE buffer and visualized on a gel image system.
Essential oil isolation
Aerial parts were collected at the full flowering stage. The plants were dried in shade conditions at 25 °C. Clevenger- apparatus was applied for essential oil extraction from thyme plants. For this purpose, 500 ml of distilled water was added to 40 g powder of each leaf sample and boiled for 5 h. The obtained essential oil was transferred to glass containers and stored at 4 °C. Essential oil content was measured from 77 accessions belonging to 11 Thymus species, and the average of essential oil content in each species was calculated. The essential oil content was evaluated according to dry matter, and measurements were performed in triplicates (Tohidi et al. 2017).
Morphological trait analysis
The morphological traits were measured by randomly selecting individuals of each replicate. The means of each studied species were considered for further analysis. The studied morphological traits were plant height (cm), days to flowering, days to 50% flowering, days to 100% flowering, dry weight (g/plant), fresh weight (g/plant), canopy extension (cm), leaf length (mm), leaf width (mm), stem diameter (mm), inflorescence diameter (mm), inflorescence length (mm), number of floret in inflorescence, and essential oil content (% v/w) (Table 3).
Data analysis
The polymorphic ISSR bands in each gel were recorded as absent (0) or present (1). The principal coordinate (PCoA) and cluster analyses were carried out through NTSYSpc, ver. 2. 02 (Rohlf 1998). The polymorphic information content (PIC) was computed based on previous research (Anderson et al. 1993). Genetic similarity among samples was assessed based on the Jaccard's similarity index (Jaccard 1908). The genetic structure parameters were assessed using Pop Gene software, ver. 32 (McDermott and McDonald 1993). The Arlequin ver. 3 software was also used to estimate the variation among and within the Thymus species. Also, STRUCTURE software was used to assess the admixture of genotypes based on Evanno’s Method (Evanno et al. 2005). The K value (2–10) was evaluated based on those explained by Pritchard et al. (2000). Morphological and essential oil data were analyzed using SAS ver. 9.4 and Statgraphics ver. 18.1.08.
Results and discussion
Molecular results
In a pre-screening of primers, 13 ISSR primers were tested in which nine showed high polymorphism and sharpness in the tested genotypes. The products showed the range of 150–2000 bp in size. The average polymorphism (%) was 94.43%. The number of polymorphic bands in primer varied from 7 to 35, with an average of 16.77 (Table 2). The average value of the PIC was 0.29 (Table 2). P13 and P15 showed the lowest PIC (0.24), while P17 revealed the highest value (0.39). Thymus species have shown different polymorphism content, including T. daenensis (89%) (Rahimmalek et al. 2009), T. daenensis (81.5%) (Heydari et al. 2019). T. sibthorpii (71%) (Abraham et al. 2018) and higher polymorphism observed in this study might be attributed to inter and intrageneric variation of 11 Thymus species. Similar inter- and intravariation was also obtained in similar studies on T. caramanicus (Hadian et al. 2014) and T. serpyllum (Sostaric et al. 2012).
ISSR genetic structure
Based on AMOVA (Analysis of the molecular variance), 72.34% of the variation was attributed to “within species”, while the “among species” variation assigned for 27.66% of the variation (Table 4). These results were in line with the results of Kumar et al. (2019). The results also showed a relatively moderate differentiation in allele frequencies (Fst = 0.27) and relatively high gene flow (Nm = 0.92) among the studied species (Table 4). Besides, most of the genetic variation was obtained within species (Table 4). The genetic structure criteria are shown in Table 5. The highest values for genetic variation (0.19), the number of effective alleles (1.29), and the Shannon index (I) (0.29) were obtained for T. transcaspicus from Khorasan Razavi. In contrast the lowest values were observed in T. serpyllum and T. trautvetteri from Shahedieh (Table 5).
Cluster analysis was performed to investigate the genetic relationships among the species. According to the Mantel test, the highest correlation was obtained between the Jaccard similarity coefficient and cophenetic matrices (r = 0.81). Cluster analysis was done using the UPGMA algorithm. The clustering classified Thymus species into four main clusters. The first main cluster (Group I) included five species, including T. carmanicus, T. transcaspicus, T. pubescens, T. fedtschenkoi, and T. trautvetteri (Fig. 1). The second main cluster consisted of T. daenensis, T. fallax, T. migricus, and T. kotschyanus (Group II). The third cluster (Group III) included T. vulgaris (Fig. 1), and the rest of the species were classified in group IV (Fig. 1). However, the classification did not distinctly separate the species. For instance, some of the accessions of one species were classified in different groups, while T. vulgaris was classified in one group. This kind of discrepancy in the cluster might be interpreted by ploidy level, cross-pollination, high gene flow, and natural hybridization (Fadaei Heidari et al. 2016; Ghafouri and Rahimmalek 2018).

Dendrogram of 77 studied Thymus genotypes based on ISSR markers according to the Unweighted Pair Group Mean Algorithm (UPGMA) with the Jaccard similarity index (r = 0.81). TM: T. migricus; TF: T. fallax; TC: T. carmanicus; TK: T. kotschyanus; TP: T. pubescens; TD: T. daenensis; Ttv: T. trautvetteri; Tts: T. transcaspicus; Tfed: T. fedtschenkoi; TV: T. vulgaris; TS: T. serpyllum
Principal coordinate analysis (PCoA) also confirmed the dendrogram data (Fig. 2). The first three principal coordinates explained 37.2% of the total variation implying high distribution of ISSR markers throughout the genome.

Plot of Thymus samples by principal coordinate analysis using the Jaccard’s similarity coefficients. T. migricus; T. fallax; T. carmanicus; T. kotschyanus; T. pubescens; T. daenensis, T. trautvetteri; T. transcaspicus; T. fedtschenkoi; T. vulgaris; T. serpyllum
High variation among the species might be due to processes like a recombination of genes and chromosomes, gene mutation, the genetic structure of species, and the number of effective individuals (Ghafouri and Rahimmalek 2018). Hadian et al. (2014) and Talebi et al. (2015) also observed high genetic diversity among the Thymus species. High genetic variation within the species can be due to the different fertility of seeds, dispersal, and reproduction method (Gupta et al. 1999). Furthermore, AMOVA revealed significant variation among the species (P < 0.001). In this research, the Fst value of 0.27 was obtained, which represented the mean difference between species. T. serpyllum and T. pubescens showed the highest similarity (0.945), while the least was gained between T. trautvetteri and T. fallax (0.830). The highest and lowest genetic distance was also observed between T. fedtschenkoi and T. vulgaris (0.191), and T. pubescens and T. serpyllum (0.056), respectively (Table 6). The low gene diversity within T. pubescens and T. serpyllum species might be interpreted as the low gene flow, as compared with other species (Fadaei Heidari et al. 2016).
In the present study, high genetic differentiation was observed among the species (Gst = 0.35). Three classification types have been reported for the coefficient of genetic differentiation including strong (Gst > 0.15), moderate (0.15 > Gst > 0.05), and low (Gst < 0.05) (Ghafouri and Rahimmalek 2018). Based on this classification, the value of Gst showed a high genetic differentiation in the Thymus species. The results were in agreement with those obtained by Ray et al. (2018) in Hedychium coronarium. STRUCTURE analysis was also performed for the better expounding of admixture in some species obtained via cluster and principal component analysis (PCA). Accordingly, six autonomous runs produced consistent outcomes. For the given data collection, the K value was appraised by the posterior probability of the data for a given K, Pr (X|K) (Pritchard et al. 2000). The highest value was afforded at K = 3. Thus, the analysis was accomplished according to three colors viz. red (A), blue (B), and green (C) for the studied Thymus species, as displayed in Fig. 3. The admixture model with correlated allele frequency was applied to inter genetic structure commentary. One of these clusters (Cluster A in Fig. 3) included the species containing T. carmanicus, T. fedtschenkoi, and T. transcaspicus. Cluster B contained species such as T. daenensis, T. fallax, T. migricus, and T. kotschyanus. The remaining species including, T. pubescens, T. vulgaris, T. serpyllum, and T. trautvetteri were classified in cluster C. The high admixture was perceived in this analysis. For instance, T. fallax, T. pubescens, and T. serpyllum showed high admixture in this study. One eventual reason for this admixture could be due to natural hybridization in some Thymus species out of lack of self-pollination strategies such as incompatibility mechanism.

Genetic structure of 77 Thymus genotypes as inferred by STRUCTURE software with nine ISSR markers data set. Single vertical line represents an individual accessions and different colors represent genetic stocks/gene pools. Segments of each vertical line show extent of admixture in an individual (For interpretation of the references to colour in the text, the reader is referred to the web version of this article). Tm: T. migricus; Tf: T. fallax; Tc: T. carmanicus; Tk: T. kotschyanus; Tp: T. pubescens; Td: T. daenensis; Ttrau: T. trautvetteri; Tt: T. transcaspicus; Tfed: T. fedtschenkoi; Tv: T. vulgaris; Ts: T. serpyllum
Essential oil content
In the first year (2018), essential oil content ranged from 0.63 to 1.94%, with a mean value of 1.26% (Fig. 4). The highest essential oil content and the lowest was obtained in T. migricus (1.94%), and T. fallax, respectively. In 2019, essential oil content varied from 0.86 to 2.34% to exhibit a significant variation with an average value of 1.52% (Fig. 4). T. vulgaris and T. carmanicus revealed the highest (2.34%) and lowest (0.86%) essential oil content.

Essential oil content of studied Thymus species
The following descending arrangement was gained for essential oil content in both years:
Environmental factors and plant species might highly effect on essential oil content (Llorens et al. 2014; Tohidi et al. 2017). Previous researches reported different ranges for the essential oil content in various Thymus species, including T. pannonicus (1.29%) (Arsenijević et al. 2019) and T. capitatus (2.37%) (Tammar et al. 2018).
In the present study, high variation was observed in terms of essential oil content in two studied years. Similarly, Arabsalehi et al. (2018) revealed a high variation in Stachys lavandulifolia essential oil content in two agronomic years.
Morphological trait analysis
Analysis of variance was performed to specify the effects of replication, species, year, and their interactions on morphological traits of Thymus species (Table 7). In both agronomic years, the coefficient of phenotypic variation for plant height was higher than other traits (CV = 13.80%). However, the lowest value of the coefficient of variation belonged to days to 50% flowering (CV = 0.39%) (Table 7).
The results of the two-year analysis of variance showed considerable differences among the studied species for all the traits. Also, the effect of species was significant for all the traits, and interactions of S × R (Species × Replication) was meaningless for all the characters (Table 7).
During the first agronomic year, T. fallax displayed the highest value for days to flowering (183.1 days), while T. pubescens (165 days) showed the lowest one (Table 3). The highest value of days to 100% flowering belonged to T.carmanicus and T. vulgaris, while the lowest was related to T. kotschyanus (Table 3). Leaf length varied from 5.3 in T. vulgaris to 16.2 mm in T. trautvetteri (Table 3). T. pubescens revealed the highest canopy extension, while T. fallax had the latest canopy extension (Table 3). Plant height ranged considerably from 11.9 (T. transcaspicus) to 21.3 cm (T. vulgaris) (Table 3). T. vulgaris and T. carmanicus possessed the highest and lowest fresh weight, respectively. Dry weight varied from 9.2 (T. carmanicus) to 28.9 (T. vulgaris) (g/plant) (Table 3). The highest and the lowest essential oil content were recorded for T. migricus (1.93%) and T. carmanicus (0.67%), respectively, with an average value of 1.25% (Table 3).
In the second agronomic year, inflorescence diameter ranged from 12.5 to 23.6 mm, with a means of 19.59 mm (Table 3). The highest number of floret in inflorescence was obtained in T. serpyllum, while the lowest one was measured in T. migricus (Table 3). Stem diameter varied from 0.65 to 1.32 mm, with an average value of 0.91 mm (Table 3). T. serpyllum was the shortest (13.4 cm), while T. vulgaris (21.1 cm) was the tallest species (Table 3). Fresh weight and dry weight ranged from 43.4 to 123.8 g/plant, and 18–92 g/plant, with average values of 74.50 and 40.36 g/plant (Table 3) T. pubescens had the earliest flowering time, while the latest time was recorded for T. fallax (181.3 days). Inflorescence length varied from 16.6 mm (T. migricus) to 31.1 mm (T. carmanicus) (Table 3). T. pubescens and T. fallax exhibited the highest and the lowest canopy extension, respectively. Leaf length ranged from 5.8 (T. vulgaris) to 12.4 mm (T. serpyllum) with an average value of 8.93 (Table 3). T. vulgaris had the highest essential oil content (2.33%), while T. fallax recorded the least (0.93%) essential oil content. Finally, the highest days to 100% flowering was observed in T. carmanicus (204.1 days), while the lowest belonged to T. kotschyanus (195.7 days).
PCA and cluster analysis for morphological traits
The clustering patterns of the Thymus species according to their morphological characters gained from Ward’s method for both agronomic years are presented in Fig. 5. Results of the two-year data classified the species into three clusters. The first cluster (Cluster I) was comprised of T. trautvetteri, T. kotschyanus, T. transcaspicus, and T. fedtschenkoi. Cluster II included T. fallax, T. carmanicus, T. daenensis, T. pubescens, T. serpyllum. T. migricus and T. vulgaris with the highest essential oil content in comparison with other species in both agronomic years were grouped in cluster II, and overall, they had relatively low essential oil content than other species.

Cluster of 77 studied Thymus genotypes based on morphological traits. Td: T. daenensis, Tm: T. migricus, Tv: T. vulgaris, Tf: T. fallax, Ttv: T. trautvetteri, Ts: T. serpyllum, Tts: T. transcaspicus, Tp: T. pubescens, T. fed: T. fedtschenkoi, Tk: T. kotschyanus, Tc: T. carmanicus
PCA confirmed the results of clustering in most cases (Fig. 6). The first three PCs showed the highest variation. PC1 explained 30.82% of the total variation and showed positive correlations, with the number of floret in inflorescence, and leaf width, inflorescence diameter, inflorescence length though with negative ones with plant height, fresh weight, dry weight, and essential oil content. PC2 explained 23.49% of the total variance and had positive correlations with days to flowering, days to 50% flowering, days to 100% flowering, and leaf width but negative correlations with canopy extension, number of floret in inflorescence, dry weight, and essential oil content. PC3 revealed leaf width, stem diameter, and essential oil content as positive components. Finally, canopy extension and plant height as negative compounds that account for 10.41% of the total variance.

PCA of morphological traits in 11 studied Thymus species. Td: T. daenensis, Tm: T. migricus, Tv: T. vulgaris, Tf: T. fallax, Ttv: T. trautvetteri, Ts: T. serpyllum, Tts: T. transcaspicus, Tp: T. pubescens, T. fed: T. fedtschenkoi, Tk: T. kotschyanus, Tc: T. carmanicus
The introduction of new species with high essential oil content and elite morphological traits is one of the major goals in the breeding of medicinal plants. Selection of high essential oil content, high genetic distance as well as similar ploidy level is a prerequisite for interspecific hybridization in Thymus species. The studied T. vulgaris and T. carmanicus were tetraploid (data are not shown). Moreover, these species were extreme in some critical breeding traits such as essential oil content and plant height, fresh weight, and dry weight, and consequently, they can be suggested for further breeding strategies in thyme. High variation among the studied species could be due to processes such as gene mutation, number of effective individuals, recombination of genes and chromosomes, and the genetic structure of species. High admixture in the species can be interpreted by different factors such as pollination efficiency. In this study, significant morphological variation was observed among the Thymus species during two agronomic years. Thus, the results can help the researchers to begin new breeding programs in the studied species. T. migricus and T. pubescens were diploid (data are not shown). These species are important for future breeding programs in terms of essential oil content and genetic distance. Furthermore, thymol has been considered as the major component of Thymus species. In this regard, based on a previous research on chemical components in most of the studied species (Tohidi et al. 2017); it can be suggested that these two selected species viz. T. migricus (high thymol content) and T. pubescens (low thymol content) are suitable for further breeding programs in this genus.
Conclusion
In this research, the genetic structure and variation of 11 Thymus species were analyzed using ISSR and morphological markers. The results of the study revealed the relatively high efficiency of ISSR markers for distinguishing among the different Thymus species from various samples. Furthermore, relatively high gene flow and differentiation were observed among the species. The highest genetic distance was obtained between T. vulgaris and T. fedtschenkoi, and the highest genetic similarity was between T. serpyllum and T. pubescens. T. transcaspicus and T. trautvetteri showed the highest and lowest gene diversity, respectively. Based on the results, the high admixture of samples was observed among the species. The results of the morphological analysis indicated significant variations in morphological characteristics, as well as the essential oil content of the studied species. T. vulgaris and T. migricus were selected as the best species in terms of essential oil content in the two agronomic years. Finally, based on the genetic distances, essential oil content, and morphological traits among the studied species, T. vulgaris, and T. carmanicus may be suggested for further breeding programs after complimentary assessments.
References
Abraham EM, Aftzalanidou A, Ganopoulos I, Osathanunkul M, Xanthopoulou A, Avramidou E, Sarrou E, Aravanopoulos F, Madesis P (2018) Genetic diversity of Thymus sibthorpii Bentham in mountainous natural grasslands of Northern Greece as related to local factors and plant community structure. Ind Crops Prod 111:651–659
Anderson JA, Churchill GA, Autrique JE, Tanksley SD, Sorrells ME (1993) Optimizing parental selection for genetic linkage maps. Genome 36:181–186
Arabsalehi F, Rahimmalek M, Ehtemam MH (2018) Phytochemical and morphological variation of Stachys lavandulifolia Vahl. populations as affected by genotype× year interaction. Ind Crops prod 112:342–352
Arsenijević J, Drobac M, Šoštarić I, Jevđović R, Živković J, Ražić S, Moravčević Đ, Maksimović Z (2019) Comparison of essential oils and hydromethanol extracts of cultivated and wild growing Thymus pannonicus All. Ind Crops Prod 130:162–169
Evanno G, Regnaut S, Goudet J (2005) Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol 14:2611–2620
Fadaei Heidari E, Rahimmalek M, Mohammadi S, Ehtemam MH (2016) Genetic structure and diversity of ajowan (Trachyspermum ammi) populations based on molecular, morphological markers, and volatile oil content. Ind Crops Prod 92:186–196
Ghafouri F, Rahimmalek M (2018) Genetic structure and variation in different Iranian myrtle (Myrtus communis L.) populations based on morphological, phytochemical and molecular markers. Ind Crops Prod 123:489–499
Ghasemi Pirbalouti A, Rahimmalek M, Malekpoor F, Karimi A (2011) Variation in Antibacterial Activity, thymol and carvacrol Contents of Wild Populations of 'Thymus daenensis subsp. daenensis' Celak. Plant Omics 4:209
Gupta PK, Balyan HS, Sharma PC, Ramesh B (1996) Microsatellites in plants: a new class of molecular markers. Curr Sci 70:45–54
Gupta PK, Varshney RK, Sharma PC, Ramesh B (1999) Molecular markers and their applications in wheat breeding. Plant Breed 118:369–390
Hadian J, Bigdeloo M, Nazeri V, Khadivi-Khub A (2014) Assessment of genetic and chemical variability in Thymus caramanicus. Mol Biol Rep 41:3201–3210
Heydari A, Hadian J, Esmaeili H, Kanani MR, Mirjalili MH, Sarkhosh A (2019) Introduction of Thymus daenensis into cultivation: Analysis of agro-morphological, phytochemical and genetic diversity of cultivated clones. Ind Crops Prod 131:14–24
Jaccard P (1908) Nouvelles researches sur la distribution florale. Bull Soc Vaud Sci Nat 44:223–270
Kumar J, Heikrujam M, Sharma K, Agrawal V (2019) SRAP and SSR marker-assisted genetic diversity, population structure analysis and sex identification in Jojoba (Simmondsia chinensis). Ind Crops Prod 133:118–132
Llorens L, Llorens-Molina JA, Agnello S, Boira H (2014) Geographical and environment-related variations of essential oils in isolated populations of Thymus richardii Pers. in the Mediterranean basin. Biochem Syst Ecol 56:246–254
Lu Y, Boswell M, Boswell W, Kneitz S, Klotz B, Savage M, Salinas R, Marks R, Regneri J, Postlethwait J, Warren WC (2018) Gene expression variation and parental allele inheritance in a Xiphophorus interspecies hybridization model. PLoS Genet 14:e1007875
McDermott JM, McDonald BA (1993) Gene flow in plant pathosystems. Annu Rev Phytopathol 31:353–373
Mohammadi A, Nazari H, Imani S, Amrollahi H (2014) Antifungal activities and chemical composition of some medicinal plants. J Mycol Med 24:e1–e8
Murray MG, Thompson WF (1980) Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res 8:4321–4326
Pirbalouti AG, Hashemi M, Ghahfarokhi FT (2013) Essential oil and chemical compositions of wild and cultivated Thymus daenensis Celak and Thymus vulgaris L. Ind Crops Prod 48:43–48
Pritchard JK, Stephens M, Donnelly P (2000) Inference of population structure using multilocus genotype data. Genetics 155:945–959
Rahimmalek M, Bahreininejad B, Khorrami M, Tabatabaei BES (2009) Genetic variability and geographic differentiation in Thymus daenensis subsp. daenensis, an endangered medicinal plant, as revealed by inter simple sequence repeat (ISSR) markers. Biochem Genet 47:831
Rasooli I, Rezaei MB, Allameh A (2006) Ultrastructural studies on antimicrobial efficacy of thyme essential oils on Listeria monocytogenes. Int J Infect Dis 10:236–241
Ray A, Jena S, Haldar T, Sahoo A, Kar B, Patnaik J, Ghosh B, Panda PC, Mahapatra N, Nayak S (2019) Population genetic structure and diversity analysis in Hedychium coronarium populations using morphological, phytochemical and molecular markers. Ind Crops Prod 132:118–133
Rechinger KH (1963) Flora Iranica No. 158. Akademische Druke-U, Verlagsanstalt, Wien Austria, pp 49–71
Rohlf F (1998) NTSYS-pc Version 2.0. Numerical taxonomy and multivariate analysis system. Exeter Software Setauket, New York
Rustaiee AR, Yavari A, Nazeri V, Shokrpour M, Sefidkon F, Rasouli M (2013) Genetic diversity and chemical polymorphism of some Thymus species. Chem Biodivers 10:1088–1098
Sarrou E, Tsivelika N, Chatzopoulou P, Tsakalidis G, Menexes G, Mavromatis A (2017) Conventional breeding of Greek oregano (Origanum vulgare ssp. hirtum) and development of improved cultivars for yield potential and essential oil quality. Euphytica 213:104
Sostaric I, Liber Z, Grdisa M, Marin PD, Stevanovic ZD, Satovic Z (2012) Genetic diversity and relationships among species of the genus Thymus L. (section Serpyllum). Flora 207:654–661
Talebi M, Rahimmalek M, Norouzi M (2015) Genetic diversity of Thymus daenensis subsp. daenensis using SRAP markers. Biologia 70:453–459
Tammar S, Salem N, Bettaieb Rebey I, Sriti J, Hammami M, Khammassi S, Marzouk B, Ksouri R, Msaada K (2019) Regional effect on essential oil composition and antimicrobial activity of Thymus capitatus L. J Essent Oil Res 31:129–137
Tohidi B, Rahimmalek M, Arzani A, Trindade H (2020) Sequencing and variation of terpene synthase gene (TPS2) as the major gene in biosynthesis of thymol in different Thymus species. Phytochem 169:112126
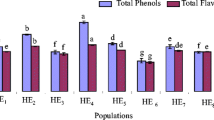
Tohidi B, Rahimmalek M, Arzani A (2017) Essential oil composition, total phenolic, flavonoid contents, and antioxidant activity of Thymus species collected from different regions of Iran. Food Chem 220:153–161
Tohidi B, Rahimmalek M, Trindade H (2019) Review on essential oil, extracts composition, molecular and phytochemical properties of Thymus species in Iran. Ind Crops Prod 134:89–99
Trindade H, Costa MM, Sofia BLA, Pedro LG, Figueiredo AC, Barroso JG (2008) Genetic diversity and chemical polymorphism of Thymus caespititius from Pico, Sao Jorge and Terceira islands (Azores). Biochem Syst Ecol 36:790–797
Trindade H, Costa MM, Lima SB, Pedro LG, Figueiredo AC, Barroso JG (2009) A combined approach using RAPD, ISSR and volatile analysis for the characterization of Thymus caespititius from Flores, Corvo and Graciosa islands (Azores, Portugal). Biochem Syst Ecol 37:670–677
Trindade H, Pedro LG, Figueiredo AC, Barroso JG (2018) Chemotypes and terpene synthase genes in Thymus genus: State of the art. Ind crops prod 124:530–547
Verma KS, ul Haq S, Kachhwaha S, Kothari SL (2017) RAPD and ISSR marker assessment of genetic diversity in Citrullus colocynthis (L.) Schrad: a unique source of germplasm highly adapted to drought and high-temperature stress. 3 Biotech 7:288
Yousefi V, Najaphy A, Zebarjadi A, Safari H (2015) Molecular characterization of Thymus species using ISSR markers. J Anim Plant Sci 8:55
Acknowledgements
The authors appreciate their gratitude to Isfahan University of Technology, Iran for their financial support. We also thank Dr. Behnaz Tohidi for editing of the article.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical statement
The manuscript has not been previously published, is not currently submitted for review to any other journal, and will not be submitted elsewhere before one decision is made.
Rights and permissions
About this article

Cite this article
Sarfaraz, D., Rahimmalek, M., Saeidi, G. et al. Genetic relations among and within wild and cultivated Thymus species based on morphological and molecular markers. 3 Biotech 10, 289 (2020). https://doi.org/10.1007/s13205-020-02274-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13205-020-02274-6