
Abstract
Areolae of the crustose lichen Rhizocarpon geographicum (L.) DC., are present on the peripheral prothallus (marginal areolae) and also aggregate to form confluent masses in the centre of the thallus (central areolae). To determine the relationships between these areolae and whether growth of the peripheral prothallus is dependent on the marginal areolae, the density, morphology, and size frequency distributions of marginal areolae were measured in 23 thalli of R. geographicum in north Wales, UK using image analysis (Image J). Size and morphology of central areolae were also studied across the thallus. Marginal areolae were small, punctate, and occurred in clusters scattered over the peripheral prothallus while central areolae were larger and had a lobed structure. The size-class frequency distributions of the marginal and central areolae were fitted by power-law and log-normal models respectively. In 16 out of 23 thalli, central areolae close to the outer edge were larger and had a more complex lobed morphology than those towards the thallus centre. Neither mean width nor radial growth rate (RaGR) of the peripheral prothallus were correlated with density, diameter, or area fraction of marginal areolae. The data suggest central areolae may develop from marginal areolae as follows: (1) marginal areolae develop in clusters at the periphery and fuse to form central areolae, (2) central areolae grow exponentially, and (3) crowding of central areolae results in constriction and fragmentation. In addition, growth of the peripheral prothallus may be unrelated to the marginal areolae.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
The yellow-green species of the crustose lichen genus Rhizocarpon comprise discrete areolae containing the alga Trebouxia which develop on a black prothallus composed of fungal hyphae, a growth form which may represent one of the earliest types of lichen symbiosis (Galløe 1930; Armstrong and Bradwell 2010a; Armstrong 2011). The prothallus extends beyond the region of areolae to form a peripheral concentric ring up to 2 mm in width (Armstrong and Bradwell 2001, 2010a). Galløe (1930) preferred the term ‘inter-areolar thallus’ to prothallus as areolae do not appear to form on the surface of the prothallus but to develop by local transformation of the prothallus as a result of thickening and changes in colour of superficial hyphae. However, prothallus has become the generally accepted term for the basal hyphae. The growing edge of the lichen, i.e., the peripheral prothallus, advances very slowly, recorded annual radial growth rates (RaGR) being in the range 0.02 to 1 mm yr-1 (Hale 1967; Armstrong 1983; Innes 1985; Armstrong 2005; Trenbirth and Matthews 2010; Armstrong and Bradwell 2010a; Armstrong 2011). The deepest basal hyphae of the prothallus appear colourless and hence, the dark pigment in the upper layers may act as a protection against high light intensities (Galløe 1930).
The development of Rhizocarpon follows a different course from that of foliose species, squamulose species such as Cladonia (Galløe 1954), and many crustose species such as Lecidea (Galløe 1927). Initial development of the areolae and the formation of the prothallus have been studied in Rhizocarpon lecanorinum (Flörke ex Körb) Anders (Clayden 1998). When ascospores of R. lecanorinum germinate, the hyphae remain short and do not initially form a prothallus. A compact soredium-like granule develops which associates with a compatible species of the green alga Trebouxia. Free-living Trebouxia cells are often the first to colonize a bare substratum and such cells are often present on the surface before any lichens become established (Mukhtar et al. 1994). Thallus differentiation occurs, accompanied by the deposition of rhizocarpic acid within the developing cortical layer, resulting in the formation of a typical areole in which the algae occur in well developed clusters (Galløe 1930). The radiating prothallus is initiated by growth from the basal margin of the areolae. The pattern of development of R. lacanorinum may represent an evolutionary reduction of an ancestral, more prolonged development as in R. geographicum (Galløe 1930; Clayden 1998), the latter species having a more divergent mycelial growth in the early stages and a slower rate of development.
Asta and Letrouit-Galinou (1995) identified four further processes in the development and growth of a mature Rhizocarpon thallus: (1) the formation of new areolae on the peripheral prothallus, (2) growth and division of existing areolae, (3) the confluence of areolae, and (4) the fusion of smaller thalli to form larger individuals. Galløe (1930) observed the formation of areoles in various species of Rhizocarpon resulting from presumed deposition of algal cell on the prothallus. Different species varied in the density and dispersion of areolae over the marginal prothallus and in the degree of prominence of the marginal prothallus. Hence, species growing in more extreme environments often had more scattered areolae (Galløe 1930; Runemark 1956). Galløe (1930) also observed that newly formed areolae were small and spatially separated and described them as ‘cushion-shaped’ but in time they increased in height and width by intercalary growth.
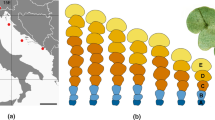
Ideally, developmental processes should be studied by direct observation from sequential photographs but the slow growth and longevity of Rhizocarpon thalli make this approach impractical other than over long periods of time. It is possible, however, that because of the radially growing thallus developmental processes could be inferred indirectly by studying areolae at different locations across the thallus. Hence, areolae of Rhizocarpon geographicum (L.) DC. are highly variable in size and shape (Runemark 1956; Armstrong and Bradwell 2010a) (Fig. 1). Areolae in the peripheral zone (marginal areolae), are frequently small and punctate while those in the central confluent region (central areolae) are significantly larger and exhibit a more complex angular or lobed shape (Galløe 1930; Runemark 1956). Whether the central areolae develop from the marginal areolae is unknown. In addition, it is unclear whether radial growth of the peripheral prothallus depends on algae in marginal as well as central areolae (Armstrong and Smith 1987, 2009). Hence, this study had two main objectives: (1) using image analysis, to study the possible relationship between marginal and central areolae and (2) to determine whether RaGR of the peripheral prothallus was related to density, size, or area fraction of the marginal areolae.

A thallus of Rhizocarpon geographicum (L.) DC. showing marginal areolae (MA) and central areolae (CA). Clearly visible are the punctate MA on the marginal prothallus (MP), the larger multi-lobed areolae at the outer edge of the central region (CA1), and the smaller areolae towards the thallus centre (CA2). Note the boundary between the peripheral and central region and that in the central confluent region the prothallus is only visible as a thin black line around the areolae (SF = 1 mm2 sample field on marginal prothallus, A-B = transect drawn across central region
2 Materials and methods
2.1 Site
The study site was a south-facing maritime rock surface located in the Dovey estuary at ‘Picnic island,’ Aberdovey, north Wales (Grid Ref. SN 6196) in an area of Ordovician slate rock described previously (Armstrong 1974). The surface possesses a rich lichen flora characteristic of maritime siliceous rock in the west of the UK (James et al. 1977), has a high proportion of crustose species (Armstrong 1974), including a large population of R. geographicum (Armstrong 1974, 2002).
2.2 Taxonomy
Molecular phylogenetic studies (Ihlen and Ekman 2002) have suggested that the frequently used subgenus and section classification of the yellow green Rhizocarpon species (Poelt 1988) is no longer tenable. Hence, the thalli used in the present study were identified as R. geographicum (L.) DC. using the following criteria: (1) thallus was yellow-green and areolae possessed flat black apothecia, (2) spores were dark brown and two-four celled (Galløe 1930), (3) epihymenium was not black, and (4) medulla was I+ intense blue/violet. None of the thalli encountered exhibited a more continuously lichenised and finely fissured thallus which would indicate the presence of R. geographicum subsp. prospectans (Räsänen) D. Hawksw. & Sowter, a common subspecies in coastal regions (Purvis et al. 1992; Smith et al. 2009).
2.3 Selection of thalli and photography
A random sample of 30, R. geographicum thalli, 0.5–2.4 cm in diameter, was selected on the rock surface for study. However, seven thalli were excluded because they exhibited a degree of discolouration or fragmentation of the centre, did not possess a distinct marginal prothallus at least 1 mm in width, or had potentially competing lichen thalli touching or overgrowing the marginal prothallus. Each of the remaining 23 thalli was photographed it its entirety using a Canon IXUS-70 digital camera, which incorporates a x12 zoom lens, and which provides a particularly clear image of the peripheral prothallus and the marginal areolae (Armstrong and Bradwell 2010b) (Fig. 1). A scale measure was placed next to each thallus.
2.4 Image analysis
There may not be an abrupt boundary between the peripheral and central regions at all points around the thallus (Fig. 1) Hence, for analysis, an approximate boundary was established between the peripheral prothallus and the central confluent region and was located where the central areolae become confluent and the prothallus was visible only as a black line surrounding the areolae. First, density, sizes, and area fraction of the marginal areolae were measured using ‘Image J’ software developed by the National Institute of Health (NIH), Bethesda, USA (Syed et al. 2000; Girish and Vijayalakshmi 2004; Armstrong and Bradwell 2010b). Each lichen image was opened using the software and magnified to clearly reveal the marginal areolae. Three to eight, 1 mm square sample fields, the number depending on thallus size, were placed at random locations on the peripheral prothallus of each thallus, with one side of the field aligned along the edge of the growing margin. The image was ‘cropped’ to include only the selected sample field. The image was then ‘thresholded’ to the maximum limit, a procedure which results in only the white outlines of the marginal areolae remaining visible. Each marginal areola ≥0.1 mm in diameter, which was not touching the edge of the sample field, was included in the sample. The following data were then obtained from each sample field: (1) total number of marginal areolae, (2) diameter of each areola estimated as the greatest diameter of the ellipse of best shape fitted to the areola, and (3) area of each areola. At this stage, each of the marginal areolae identified were examined and ambiguous structures, e.g., those not in clear focus, excluded from the sample. The area fraction of the sample field occupied by the remaining marginal areolae was then calculated. In addition, areolae were divided into three morphological subtypes: (1) simple ‘punctate’ (unlobed) areolae, (2) lobed areolae with two distinct segments, and (3) multi-lobed areolae with more than two segments. The frequency of each morphological type of areola was then determined in each sample field.
Second, to study central areolae, a transect was drawn across the greatest diameter of the central region (Fig. 1). The maximum diameter of each areola that touched the transect line was then measured and its morphology assigned to one of the three groups defined above. Each transect line was also divided into three segments to compare the morphology of central areolae located at the outer edges with those towards the thallus centre. Each outer edge comprised 20 % of the transect length, and the remaining central portion comprising 60 % of the transect length.
2.5 Radial growth rate (RaGR)
RaGR of the marginal prothallus (Armstrong and Smith 1987; Armstrong 2006) was measured at each location used to sample the marginal areolae, at 3-month intervals over 1 year from July 2010 to July 2011 using the method described by Armstrong (1973, 1975, 2006). The advance of the prothallus, using a micrometer scale, was measured in relation to fixed markers on the substratum. Radial growth increments were summed over 3-month periods and averaged for each thallus.
2.6 Statistical analysis
Statistical analysis was carried out using STATISTICA Software (Statsoft Inc., 2300 East 14th St, Tulsa, OK 74104, USA). First, the correlation between the mean width of the prothallus and RaGR, averaged over all locations, and the mean density, diameter, and area fraction of the marginal areolae were tested using Pearson’s correlation coefficient ‘r’.
Second, a Poisson distribution was fitted to the marginal areolae data from all thalli (Armstrong 2007). If the marginal areolae were distributed at random over the peripheral prothallus then the probability (P) that the fields contain various numbers of areolae is given by the Poisson distribution. Hence, the Poisson distribution can be used to calculate the expected number of sample fields containing 0, 1, 2, 3, …, n, individuals and deviations of the observed from the expected frequencies tested using either a Kolmogorov-Smirnov (KS) or chi-square (χ 2) ‘goodness of fit’ test. In addition, the variance (V) of a Poisson distribution is equal to its mean (M) and hence, the V/M ratio is unity for a random distribution. The V/M ratio can therefore be used as is an index of dispersion, uniform distributions having a V/M ratio less than unity and clustered distributions greater than unity (Armstrong 2007).
Third, areolae diameters, combined from all thalli, were used to construct size class-frequency distributions of the marginal and central areolae. Three statistical models were fitted to these distributions: (1) a negative exponential decay model, by transforming the frequencies (Y variable) to logarithms and fitting a linear regression to the data, the goodness of fit to a linear model being tested using Pearson’s ‘r’, (2) a ‘power-law’ model by transforming both the frequencies (Y variable) and diameters (X variable) to logarithms, and fitting a linear regression, and (3) a log-normal distribution (Pollard 1979) which is often used to describe the size distributions of plant populations (Limpert et al. 2001). Goodness-of-fit to a log-normal model was tested using the KS test.
Fourth, polynomial curves were fitted to the relationship between maximum diameter of a central areola and its location along the transect. For each thallus, polynomials up to the fourth order were fitted successively to the data. At each stage, the correlation coefficients (Pearson’s ‘r’), regression coefficients, standard errors (SE), values of t, and the residual mean square were obtained. The reduction in the sums of squares was tested for significance as each term was included and the analysis continued until either a non-significant value of the variance ratio (F) was obtained or there was little gain in the explained variance (Snedecor and Cochran 1980).
Fifth, variations in the frequency of areolae of different morphology were compared using χ 2 contingency table tests: (1) between marginal and all central areolae and (2) within the central region, between areolae at the outer edges of the central zone compared with those towards the thallus centre.
3 Results
The density (mm−2), mean diameter (mm), and mean area fraction (mm2 mm−2) of the marginal areolae in the peripheral zone together with the mean RaGR (mm yr−1) of the marginal prothallus are shown for each thallus in Table 1. The density of the marginal areolae was in the range 1.5–10.9 per mm2 of prothallus, and their diameters in the range 0.10–0.26 mm. There were no significant correlations between either RaGR or mean prothallus width of a thallus and the mean density, diameter, or area fraction of marginal areolae.
The fit of the frequency distribution of number of areolae per sample field to the Poisson distribution is shown in Fig. 2. Marginal areolae exhibited a significant deviation from a Poisson distribution towards clustering (χ 2 = 31.91, 3DF, P < 0.001, V/M = 2.33) with a greater number of fields than expected with zero and greater than eight areolae, and fewer than expected with between four and eight areolae.

Frequency distribution of the number of marginal areolae per sample field of a sample of 23 thalli of Rhizocarpon geographicum (L.) DC. Fit to Poisson distribution (χ 2 = 31.91, 3DF, P < 0.001, V/M = 2.33)
The size-class frequency distributions of the areolae are shown in Figs. 3 and 4. Frequency of the marginal areolae declined rapidly with increasing size (Fig. 3). A power-law model provided the best fit to this distribution (goodness-of-fit: r = −0.93, P < 0.001). By contrast, the size-class distribution of the central areolae (Fig. 4) was positively skewed with a modal class at an upper size limit of 0.6 mm and a maximum diameter of 2.8 mm. This distribution was best fitted by a log-normal model (deviation from model: KS = 0.05, P > 0.05).

Size-class frequency distribution of marginal areolae of Rhizocarpon geographicum (L.) DC. Fit to a power-law model (r = −0.93, P < 0.001)

Size-class frequency distribution of central areolae of Rhizocarpon geographicum (L.) DC. Deviation from log-normal model (KS = 0.05, P > 0.05)
Variation in diameter of central areolae across a single thallus is shown in Fig. 5. This distribution was fitted by a fourth-order (quartic) polynomial (r = 0.70, P < 0.001), central areolae having greater diameters at the outer edges of the central region compared with the thallus centre, 16/23 thalli exhibiting a similar pattern of distribution (Table 2). Areolae diameter increased linearly across the central region of the thallus in one individual, areolae were larger at one edge only than in the centre in two thalli, areolae were larger in the centre than at both edges in one thallus, and in two thalli, there were no significant differences in diameter across the thallus.

Variation in the diameter of areolae across the central region of a single thallus (Thallus 7) of Rhizocarpon geographicum (L.) DC. Distribution was fitted by a fourth-order (quartic) polynomial (r = 0.70, P < 0.001)
The frequency of areolae of different morphology is shown in Table 3. The marginal and central areolae exhibited a different range of morphologies (χ 2 = 160.75, P < 0.001), marginal areolae being more frequently punctate and central areolae having a lobed structure. In addition, in the central confluent region, areolae in the outer zone were more frequently multi-lobed compared with those in the zone towards the thallus centre (χ 2 = 31.32, P < 0.001).
4 Discussion
Two problems were encountered using Image J to count and measure marginal areolae in R. geographicum. First, not all structures identified by the software, especially those smaller than 0.1 mm, could be confirmed as areolae and therefore, no areola smaller than 0.1 mm was included in a sample. In addition, after analysis of each sample field, the marginal areolae were examined and those not clearly in focus eliminated. Second, on a few occasions the software could not identify the individual areolae within a cluster and these areolae were also eliminated from the measured sample.
Peripheral and central areolae exhibited a number of differences. Hence, marginal areolae were small, punctate in shape, and occurred in clusters widely scattered over the peripheral prothallus (Galløe 1930). By contrast, central areolae were significantly larger and had a more complex, lobed morphology. Marginal areolae could originate from: (1) overgrowth by the peripheral prothallus of free-living algal cells located on the substratum (Galløe 1930; Hill 1984; Armstrong and Smith 1987; Clayden 1998; Sanders and Rico 1992), (2) algal cells which are deposited on the surface of the prothallus (Galløe 1930), (3) algal cells originating from the central areolae ‘pushed’ into the growing area by specialised hyphae (Nienburg 1926), or (4) zoospores from central areolae that swim and colonise the marginal prothallus (Slocum et al. 1980). The smallest and largest marginal areolae, however, were frequently located adjacent to either the growing margin of the prothallus margin or the edge of the central region respectively, suggesting that at least some marginal areolae could have originated from algae on the substratum (Mukhtar et al. 1994). If this hypothesis is correct then the number of the marginal areolae may depend on the density of available algal cells (Galløe 1930). It was also observed by Galløe (1930) that algal cells either trapped by the advancing prothallus or which fall onto the prothallus were enveloped by a protective layer of black hyphae under which they divided and propagated, subsequently differentiating into areolae.
The size-class frequency distributions of the marginal and central areolae were also markedly different. Size frequency distribution of the marginal areolae was fitted by a power-law model consistent with a rapid decline in frequency with increasing size. This distribution could reflect the recent appearance of large numbers of areolae on the peripheral prothallus, a high rate of mortality of areolae, or that small marginal areolae, especially those developing in clusters, rapidly coalesce at the margin. By contrast, the size-class distribution of the central areolae was fitted by a log-normal model. Both the normal and log-normal distributions result from a variety of factors acting independently, additive effects resulting in a normal distribution and multiplicative effects in a log-normal distribution. As a consequence, many physical, chemical, biological, and statistical processes create random variables that follow a log-normal distribution (Hattis and Burmaster 1994). The size and age structure of many plant populations, for example, frequently follow a log-normal distribution which is the result of exponential growth, but with growth of individuals exhibiting random variation (Limpert et al. 2001). The data suggest a similar model for R. geographicum, viz., the organism comprising a slow-growing fungal prothallus associated with a colony of independently and exponentially growing areolae.
The diameter and morphology of the areolae also varied across the central region, the largest multi-lobed areolae being located at its edges. The large areolae at the edge of the central region could be the result of two processes, viz., relatively unrestricted exponential growth and fusion of marginal areolae. Within the central confluent region, areolae located towards the centre of the thallus are usually smaller and possess fewer lobes than those at the edges. Rapid growth of central areolae at the edge together with a slow rate of advance of the prothallus is likely to result in crowding of areolae in the centre. Overcrowding in the centre could then result in constriction and/or splitting of areolae which could explain the frequency of smaller, less extensively lobed areolae in the centre.
Whether marginal areolae contribute to growth of the marginal prothallus is unclear (Armstrong 2011). No correlations were observed between either prothallus width or RaGR and the density, diameter, or area fraction of adjacent marginal areolae. Two features were notable about the growth data. First, RaGR, averaged over thalli, was approximately half that recorded in previous studies in north Wales (Armstrong 1983) and second, there was considerable variation in RaGR between thalli. The lower growth rates may be attributable either to the maritime site studied or to the exceptionally cold and dry winter of 2010 in north Wales. Variability in growth rates does not appear to be associated with thallus diameter, a result which was also apparent in R. geographicum thalli measured in situ in the Pacific Northwest (Armstrong 2005). Lichen growth on a rock surface is likely to be controlled by many factors including microscale differences in aspect, slope, surface permeability, roughness, and differential flow of nutrients together with variations in height on the surface. Armstrong (2005) found that growth of R. geographicum in the Pacific Northwest was unrelated to slope but was correlated with aspect. In addition, Haworth et al. (1986) found variations in the growth of Rhizocarpon thalli both between and within sites in the central Brooks Range, Alaska. Thalli at sites subjected to high to moderate light intensities grew approximately twice as fast as those at shaded sites (Haworth et al. 1986). However, in the present study there was no evidence that growth rates were related to height on the face and there was little significant variation in aspect and slope over the substratum. Hence, microscale variations in surface permeability, nutrient availability, or roughness could have been responsible for growth rate variation.
The growth data suggest that RaGR of the peripheral prothallus is not related to the density, sizes, or area fraction of the marginal areolae. Hence, complete removal of the central areolae resulted in no measurable growth of the prothallus over 18 months while removal of central areolae to within 1 mm and 2 mm of the prothallus significantly reduced growth in proportion to the width of central areolae present (Armstrong and Smith 1987, 2009). These results suggest that the large central areolae at the margin of the central areolae are a more likely source of carbohydrate for marginal growth (Armstrong 2009).
The growth form of R. geographicum, which may represent one of the most primitive types of lichen symbiosis, occurs consistently in several other genera including Buellia (Galløe 1930), Lecanactis and Bacidia (Galløe 1930), Lecidea (Galløe 1927), and Lecanora, Verrucaria, Polyblastia, Staurothele, Arthonia, Arthropyrenia, Opegrapha, and Enterographa (Galløe 1972). Within many of these genera, a possible evolutionary sequence can be postulated towards a more integrated thallus by suppression of the basal prothallus and fusion of areolae to form a more continuous structure (Galløe 1927, 1930, 1972). Within the genus Rhizocarpon itself, this result may be seen in R. umbilicatum (Ram.) Flag., in which the thallus is cracked-areolate but essentially continuous without an obvious prothallus. This hypothetical evolutionary sequence can also be observed in Lecidea (Galløe 1927) in which there are areolate species with a black or white prothallus, e.g., Lecidea aglaea Sommerf., species in which the thallus is granular on a dark prothallus, such as L. limitata (Scop.) Gray, and species with a more continuous thallus but with a less obvious prothallus, e.g., L. tumida Massal. It is also possible that such a growth form may provide the ancestral type for the primary thallus of many species of Cladonia (Galløe 1954), in which the ancestral prothallus disappears and the independent areolae become more leaf-like, developing ultimately into basal squamules.
In conclusion, the data suggest a possible hypothesis to explain the development of areolae in R. geographicum: (1) marginal areolae develop on the peripheral prothallus largely in clusters, (2) these clusters of areolae fuse to form the large multi-lobed areolae often observed at the outer edge of the confluent central region, (3) central areolae increase in size exponentially as a consequence of fusion and growth, and (4) overcrowding in the centre results in confluence and splitting of central areolae. In addition, the data also suggest the marginal areolae contribute little to the growth of the peripheral prothallus.
References
Armstrong RA (1973) Seasonal growth and growth rate colony size relationships in six species of saxicolous lichens. New Phytol 72:1023–1030
Armstrong RA (1974) The descriptive ecology of saxicolous lichens in an area of South Merionethshire, Wales. J Ecol 62:33–45
Armstrong RA (1975) The influence of aspect on the pattern of seasonal growth in the lichen Parmelia glabratula ssp. fuliginosa. New Phytol 75:245–251
Armstrong RA (1983) Growth curve of the lichen Rhizocarpon geographicum. New Phytol 94:619–622
Armstrong RA (2002) The effect of rock surface aspect on growth, size structure, and competition in the lichen Rhizocarpon geographicum. Environ Exp Bot 48:187–194
Armstrong RA (2005) Radial growth of Rhizocarpon section Rhizocarpon lichen thalli over six years at Snoqualmie Pass in the Cascade Range, Washington State. Arct Antarct Alp Res 37:411–415
Armstrong RA (2006) Seasonal growth of the crustose lichen Rhizocarpon geographicum (L.) DC. in south Gwynedd, Wales. Symbiosis 41:97–102
Armstrong RA (2007) Measuring the spatial arrangement patterns of pathological lesions in histological sections of brain tissue. Folia Neuropathol 44:229–237
Armstrong RA (2011) The biology of the crustose lichen Rhizocarpon geographicum. Symbiosis 55:53–68
Armstrong RA, Bradwell T (2001) Variation in prothallus width and the growth of the lichen Rhizocarpon geographicum (L.) DC. Symbiosis 30:317–328
Armstrong RA, Bradwell T (2010a) Growth of crustose lichens: a review. Geogr Ann 92A:3–17
Armstrong RA, Bradwell T (2010b) The use of lichen growth rings in lichenometry: some preliminary findings. Geogr Ann 92A:141–147
Armstrong RA, Smith SN (1987) Development and growth of the lichen Rhizocarpon geographicum. Symbiosis 3:287–300
Armstrong RA, Smith SN (2009) Carbohydrates in the prothallus and areolae of the crustose lichen Rhizocarpon geographicum (L.) DC. Symbiosis 49:95–100
Asta J, Letrouit-Galinou MA (1995) Observations on the early growth of Rhizocarpon geographicum thalli. Herzogia 11:239–252
Clayden SR (1998) Thallus initiation and development in the lichen Rhizocarpon lecanorinum. New Phytol 139:685–695
Galløe O (1927) Natural history of Danish lichens: Original investigations based on new principles. Part 1. H. Aschehoug & Co, Copenhagen
Galløe O (1930) Natural history of Danish lichens: Original investigations based on new principles. Part 3. H. Aschehoug & Co, Copenhagen
Galløe O (1954) Natural history of Danish lichens: Original investigations based on new principles. Part IX. H. Ejnar Munksgaard, Copenhagen
Galløe O (1972) Natural history of Danish lichens: Original investigations based on new principles. Part X. (Ed. MS Christiansen), Ellen Galløe, Copenhagen
Girish V, Vijayalakshmi A (2004) Affordable image analysis using NIH Image/Image J. Indian J Cancer 41:47
Hale ME (1967) The Biology of Lichens, 1st edn. Arnold, London
Hattis DB, Burmaster DE (1994) Assessment of variability and uncertainty distributions for practical risk assessments. Risk Anal 14:713–730
Haworth LA, Calkin PE, Ellis JM (1986) Direct measurement of lichen growth in the central Brooks Range, Alaska USA, and its application to lichenometric dating. Arct Alp Res 18:289–296
Hill DJ (1984) Studies on the growth of lichens 1. Lobe formation and the maintenance of circularity in crustose lichens. Lichenologist 16:273–278
Ihlen PG, Ekman S (2002) outline of phylogeny and character evolution in Rhizocarpon (Rhizocarpaceae, lichenized Ascomycota) based on nuclear ITS and mitochondrial SSU ribosomal DNA sequences. Biol J Linn Soc 77:535–546
Innes JL (1985) Lichenometry. Prog Phys Geogr 9:187–254
James PW, Hawksworth DL, Rose F (1977) In: Seaward MRD (ed) Lichen communities in the British Isles: A preliminary conspectus. Academic, London and New York, pp 295–419
Limpert E, Stahel WA, Abbt M (2001) Log-normal distributions across the sciences: keys and clues. BioScience 51:341–352
Mukhtar A, Garty J, Galun M (1994) Does the lichen alga Trebouxia occur free-living in nature: further immunological evidence? Symbiosis 17:247–253
Nienburg W (1926) Anatomie der Flechten. In: Linstauer K (ed) Anatomie der Flechten, Handbuch der Pflanzenanatomie, 6. Borntraeger, Berlin, pp 1–137
Poelt J (1988) Rhizocarpon Ram. em. Th. Fr. subgenus Rhizocarpon in Europe. Arct Alp Res 20:292–298
Pollard JH (1979) Numerical and Statistical Techniques. Cambridge University Press, Cambridge
Purvis OW, Coppins BJ, Hawksworth DL, James PW, Moore DM (1992) The Lichen Flora of Great Britain and Ireland. Natural History Museum Publication, London
Runemark H (1956) Studies in Rhizocarpon. I. Taxonomy of the yellow species in Europe. Opera Botanica 2:1–152
Sanders WB, Rico VJ (1992) Lichenizing rhizomorphs and thallus development in the squamulose lichen Aspicilea crespiana Rico. (Lecanorales, Ascomycetales). Botanica Acta 105:449–456
Slocum RD, Admadjian V, Hildreth KC (1980) Zoosporogenesis in Trebouxia gelatinosa: untrastructure, potential for zoospore release and implications for the lichen association. Lichenologist 12:173–187
Smith CW, Aptroot A, Coppins BJ, Fletcher A, Gilbert OL, James PW, Wolseley PA (eds) (2009) The Lichen Flora of Great Britain and Ireland. Natural History Museum Publication, London
Snedecor GW, Cochran WG (1980) Statistical methods, 7th edn. Iowa State University Press, Ames
Syed A, Armstrong RA, Smith CUM (2000) Quantification of axonal loss in Alzheimer’s disease: an image analysis study. Alzheimer’s Reports 3: 19–24
Trenbirth HE, Matthews JA (2010) Lichen growth rates on glacier forelands in southern Norway: preliminary results from a 25-year monitoring programme. Geogr Ann Phys Geogr 92A:19–40
Acknowledgments
The assistance of Dr K. M. Wade who carried out the digital photography is gratefully acknowledged. Grateful thanks are also due to the reviewers whose critical comments substantially improved this article.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Armstrong, R.A. Development of areolae and growth of the peripheral prothallus in the crustose lichen Rhizocarpon geographicum: an image analysis study. Symbiosis 60, 7–15 (2013). https://doi.org/10.1007/s13199-013-0234-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13199-013-0234-2