
Abstract
Background
Prevention and improvement of disease symptoms are important issues, and probiotics are suggested as a good treatment for controlling the obesity. Human gut microbiota has different community structures. Because gut microbial composition is assumed to be linked to probiotic function, this study evaluated the efficacy of probiotics on obesity-related clinical markers according to gut microbial enterotype.
Methods
Fifty subjects with body mass index over 25 kg/m2 were randomly assigned to either the probiotic or placebo group. Each group received either unlabeled placebo or probiotic capsules for 12 weeks. Body weight, waist circumference, and body composition were measured every 3 weeks. Using computed tomography, total abdominal fat area and visceral fat area were measured. Blood and fecal samples were collected before and after the intervention for biochemical parameters and gut microbial compositions analysis.
Results
Gut microbial compositions of all the subjects were classified into two enterotypes according to Prevotella/Bacteroides ratio. The fat percentage, blood glucose, and insulin significantly increased in the Prevotella-rich enterotype of the placebo group. The obesity-related markers, such as waist circumference, total fat area, visceral fat, and ratio of visceral to subcutaneous fat area, were significantly reduced in the probiotic group. The decrease of obesity-related markers was greater in the Prevotella-rich enterotype than in the Bacteroides-rich enterotype.
Conclusion
Administration of probiotics improved obesity-related markers in obese people, and the efficacy of probiotics differed per gut microbial enterotype and greater responses were observed in the Prevotella-dominant enterotype.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The prevalence of obesity has risen over the last few decades and has become a public health concern [1]. Prevention and improvement of obesity are important issues because obesity is not only a problem of its own, but it can be a risk factor for metabolic syndromes such as diabetes and cardiovascular disease [2, 3]. Recently, evidence that support obesity itself can increase mortality risk is increasing. And now it is more obvious that obesity itself can increase mortality risk. Study result of more than 10.1 million subjects from 239 large studies have shown that being overweight and obese is associated with higher risk of dying prematurely than normal weight [4]. Obesity also has a significant impact on the development of musculoskeletal diseases, such as osteoarthritis, rheumatoid arthritis, and fibromyalgia, by casing degenerative and inflammatory changes [5]. Fat accumulation in the liver, so-called hepatic steatosis, causes various hepatobiliary diseases, and obesity is independently associated with liver fibrosis due to pathophysiological changes in immune system [6, 7]. Thus, obesity itself and immune response, such as chronic low-grade inflammation caused by obesity, are important factors causing various diseases. Obesity causes chronic low-grade inflammation because adipocytokines such as IL-1, IL-6, IL-8, IFN-γ, TNF-α and adipose tissue-secreted hormones such as leptin and resistin, induce the inflammation to become chronic [8]. Leptin promote Th1 responses which disrupt the Th1/Th2 immunity balance [9, 10]. Toll-like receptor 4 signaling pathway has been recognized as one of the main triggers in increasing the obesity-induced inflammatory response [11]. These obesity-induced inflammatory condition can lead to develop the cardiovascular diseases, type 2 diabetes mellitus, and certain types of cancer [12,13,14].
Various factors are involved in the pathophysiology of obesity, such as energy balance, genetic factors, epigenetic factors, and gut microbiota [15,16,17]. Gut microbiota influences nutrient absorption, energy balance, and the immunologic system, which contribute to modulation of body homeostasis and metabolism [18,19,20]. Associations of obesity and gut microbial imbalance have been extensively studied in human and animal models [21,22,23]. Gut microbiome has a potential role in predicting and validating the personalized responses to diet and variability of human metabolism in precision nutrition research field [24, 25]. Recent study reported the combined application of inorganical prebiotics and probiotics demonstrating a great clinical potential to treat hypercholesterolemia in affected individuals [26]. Cholesterol metabolism is sensitive to microbiota profiles and the setup of appropriate probiotic, immunobiotic, and prebiotic is crucial for the personalized clinical set [26]. Modulation of the gut microbiota and probiotic therapy can be important strategies in predictive, preventive, and personalized medicine to prevent the development of obesity and metabolic syndrome.
Probiotics are defined as “live microorganisms that, when administered in adequate amounts, confer a health benefit on the host” and provide health benefits such as inhibiting colonization of pathogens, modulating cholesterol level and treatment of diseases (e.g., obesity, diabetes, non-alcoholic fatty liver disease) [27,28,29,30]. Lactobacillus and Bifidobacterium are genus containing representative probiotics. Lactic acid bacteria including Lactobacillus have been reported to have functions related with inhibition of pathogens colonization, cholesterol metabolism, liver function, and fat accumulation [31,32,33,34,35], and Bifidobacterium have been reported to promote human health by decreasing weight and cholesterol, restoring liver morphology, modulating the gut microbiota, utilizing the human milk oligosaccharides, and enhancing the immune system [31, 36,37,38].
Human gut microbiota has different community structures depending on various factors such as diet, age, and geography [39, 40]. It has been suggested that human gut microbial communities are mainly grouped into so-called enterotypes, depending on the dominant genus, such as Prevotella-dominant enterotype and Bacteroides-dominant enterotype [41, 42]. Enterotypes dominated by Ruminococcus or Enterobacteriaceae also occur, depending on the country or diet, but these are rare or ambiguous [43, 44]. There are some studies on the association of enterotype with cardiometabolic risk factors [45, 46] and the differential response to obesity-related markers, such as weight and fat mass [47, 48]. Therefore, selection of adequate probiotics for each person per their enterotype can increase their health promotion or disease preventing effects.
The aim of this study was to evaluate whether multi-strain probiotics improved obesity-related markers of subjects and to investigate the different responses per human gut microbial enterotype. The results of this study could be used for predictive diagnostics based on each individual’s enterotype information and establishing a targeted prevention strategy by predicting the progression of obesity for each enterotype. It is also expected to be utilized for personalized medical services according to enterotypes including medical care and medical foods.
Materials and methods
Study design
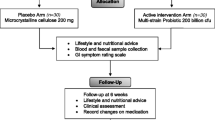
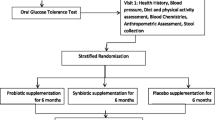
Fifty healthy obese men and women participated in a 12-week double-blind, placebo-controlled, randomized clinical trial. This study investigated the effect of probiotic supplementation for body weight control. Subject recruitment and intervention proceeded from July 23, 2016 to February 14, 2017. Subjects received either probiotics or placebo for 12 weeks. Every 3 weeks, subjects visited for anthropometrical measurements and received probiotic or placebo capsules. At every visit, subjects were thoroughly surveyed for possible adverse events.
Subject eligibility criteria
Regarding inclusion and exclusion criteria, fifty subjects between 20 and 60 of age were recruited through advertisements in posters and local newspapers. In this trial, obesity was defined as those with a body mass index (BMI) over 25 kg/m2, according to the Asia-Pacific Obesity guidelines [49]. Subjects who agreed to follow the study protocol for 12 weeks and voluntarily signed informed consent were included in this study.
Subjects with the following conditions were excluded from this study: (1) weight change more than 10% during the past 6 months; (2) weight-influencing diseases such as hypothyroidism and Cushing syndrome; (3) chronic diseases such as heart diseases, cancer, pulmonary diseases, cholelithiasis, narrow angle glaucoma, and psychogenic diseases; (4) serious functional failure of liver and kidney (aspartate aminotransferase (AST) and alanine aminotransferase (ALT) concentrations more than 2.5 times the normal values, creatinine above 2.0 mg/dl); (5) undergone surgery for weight loss, such as bypass surgery; (6) history of stroke or ischemic heart diseases; (7) history of eating disorders such as anorexia nervosa or sitomania; (8) taking drugs such as hormonal drugs, selective serotonin reuptake inhibitors, central nervous system simulants, or herbal medicine within 4 weeks; (9) history of taking anorectic agents, laxatives, steroids, thyroid hormones, amphetamine, or drugs that may affect body weight within 3 months; (10) history of taking probiotics for antibiotics within a month; (11) smokers or non-smokers who recently quit smoking within 3 months; (12) pregnant women, or who are planning to have a baby; or (13) participated in another clinical trial during the last 4 weeks.
Treatments
Following more than 10 days of run-in period, subjects took probiotic capsules or placebo capsules for 12 weeks. Subjects consumed two capsules per day after breakfast. The probiotic capsules, GP2 (Cell Biotech Co. Ltd., Gimpo, Republic of Korea), contained a formulation consisting of B. breve CBT BR3 isolated from Korean infant feces (15 billion viable cells/2 capsules), L. plantarum CBT LP3 isolated from Korean fermented vegetable product kimchi (15 billion viable cells/2 capsules), fructo-oligosaccharide, and magnesium stearate. Placebo capsules contained only fructo-oligosaccharide and magnesium stearate, but were identical in appearance.
Randomization and blinding strategies
Subjects were randomly allocated to either probiotic group or placebo group by a random code issued by an independent statistician. Random code was generated using Random Allocation Software (M. Saghaei, Iran) and sent to the manufacturer for packing. To maintain blindness on both the assessors and the subjects, probiotic and placebo capsules were packed and labeled with a provided random number before delivery to the hospital. Probiotic and placebo capsules were manufactured by Cell Biotech Co. (Gimpo, Republic of Korea), following the Korea Good Manufacturing Practice (KGMP). Placebo capsules had the same size and color as the probiotic capsules, so subjects could not differentiate the capsules. The pharmacist supplied the drug sequentially according to the random code. The blinding maintained until all the subjects had completed the study to minimize the risk of bias.
Energy intake measurements
Dietary guidelines suggested that subjects maintain a caloric restriction diet throughout the study, limiting caloric intake to 20–25 kcal per subject weight (kg). Although the subjects were not forced to maintain a caloric restriction, diets representing daily life were examined to determine whether there have been any changes during the study. Dietary intake was assessed before and after the trial, using the 24-h dietary recall method. Total energy consumption and nutrient intake were calculated using CAN-Pro 4.0 nutrient analysis software (The Korean Nutrition Society, Republic of Korea). Subjects were instructed to maintain their usual exercise program throughout the intervention. In order to rule out weight loss by exercise, people who did not exercise normally instructed to maintain their lifestyle.
Anthropometric parameters and body compositions measurements
Anthropometrical measurements were performed at every visit (Table 1). Body weight and height were measured with an automatic scale (G-tech, Uijeongbu, Republic of Korea). BMI was calculated by weight in kilograms divided by height in meters squared. Body composition measurements were determined using InBody 3.0 (Biospace, Seoul, Republic of Korea), a bioelectrical impedance analysis device to measure fat percentage, fat mass, and lean body mass. Blood pressure and pulse rate were measured using an automatic digital sphygmomanometer. To reduce errors, waist circumference (WC) and hip circumference (HC) were measured three times in a row by the same person and mean value was used for statistical analysis. WC was measured according to WHO instructions [50]. HC was measured at the widest portion of the buttocks.
Abdominal fat measurements
Abdominal adiposity and visceral fat adiposity were measured before and after the study by computed tomography. Total abdominal fat area and visceral fat area were measured in a single transverse image taken at the level of L4–5 intervertebral space [51]. Subcutaneous fat area was obtained by subtracting visceral fat area from total abdominal fat area.
Blood chemistry
After overnight fasting, blood samples were collected before and after the intervention. Fasting glucose, insulin, triglyceride (TG), total cholesterol (TC), high-density lipoprotein cholesterol (HDL-C), C-reactive protein, AST, and ALT were measured using Cobas 8000 modular analyzer (Roche, Branford, CT, USA). Low-density lipoprotein cholesterol (LDL-C) was estimated using the Friedewald equation [52]: LDL-C = (TC – HDL-C) – (TG/5).
Sequence-based microbiota analysis
Fecal samples were collected before and after the intervention and immediately stored at − 80 °C. Metagenomic DNA was isolated from the fecal samples using QIAamp DNA Stool Mini Kit (Qiagen, Hilden, Germany), per manufacturer’s instructions with an additional bead-beating step [53]. The V3–V4 regions of the 16S rRNA genes were amplified using the Illumina universal primers (341F and 806R). Amplified samples were sequenced on Illumina MiSeq platform using 2 × 300 bp MiSeq Reagent Kit V3 (Illumina, San diego, CA, USA). Raw sequence reads were quality-filtered using Trimmomatic software [54], and read pairs passing the quality filter were merged using PEAR software [55]. Chimeras were identified using USEARCH software and removed from the sequence data [56]. Quality-controlled reads were processed to alpha and beta diversity analysis using QIIME2 pipelines [57]. All raw sequencing data described in this study are available from the DNA Data Bank of Japan with the accession number DRA008223.
Serum metabolite profiling
We analyzed serum metabolites using ultra-performance liquid chromatography/quadrupole time-of-flight mass spectrometry (Synapt G2Si, Waters, USA). A total of 200 μL of isopropanol was added to 50 μL of serum samples. Supernatants were obtained and diluted in half with deionized water; then, they were injected into an ultra-performance liquid chromatography/quadrupole time-of-flight mass spectrometer (UPLC-Q/TOF-MS, Synapt G2Si, Waters, USA). Acquity UPLC BEH C18 column (2.1 mm × 100 mm, 1.7 μm, Waters) was used for separation. Mobile phase A contained 0.1% formic acid in acetonitrile:water (6:4) and 10 nM ammonium formate. Mobile phase B consisted of 0.1% formic acid in isopropanol:acetonitrile (9:1) and 10 mM ammonium formate. Samples were eluted in 40 to 53% B for 2 min, 2.1 min to 50% A, 12 min to 54% B, 12.1 min to 70% B, 18 min to 1% B, 18.1mi n to 40% B, and equilibrated for additional 2 min with the flow rate of 0.4 ml/min. Column temperature was retained at 55 °C. Mass acquisition was performed on a Q/TOF-MS (Synapt HDMS system, Waters) operating in both positive (ESI+) and negative (ESI–) electrospray ionization mode with following conditions: cone voltage of 10 V; capillary voltage of 2.0 kV; desolvation temperature was 550 °C; source temperature was 110 °C; and desolvation gas flow was 900 L/h. The range of m/z was 60 to1400, mass data were collected with a scan time of 0.25 s, and interscan time of 0.02 s for 12 min. Information from mass data, including retention time, ion intensity, and m/z, were extracted using Progenesis QI software (Waters). After alignment and normalization, the multivariate data matrix was exported into SIMCA-P ver. 14.1+ (Umetrics, Umea, Sweden) for multivariate statistical analysis. The ions contributing to the separation of probiotic and placebo groups among female subjects over age 45 were further investigated by searching the METLIN database (http://metlin.scripps.edu/) and human metabolomics database (http://www.hmdb.ca/).
Statistical analyses
For statistical analyses of clinical parameters, intention-to-treat (ITT) analysis was used with the last observation carried forward method. Fisher’s exact test was used for categorical data, such as gender. For continuous data, statistical analyses were done after verifying normality. Independent t test or Mann-Whitney test were used for the comparative analyses between two groups. We used paired t test or Wilcoxon signed rank test to compare pre-post results within groups. Statistical analyses were performed with SPSS statistical software package 18.0 (SPSS Inc., Chicago, IL, USA). A p value less than 0.05 was considered statistically significant.
Results
Study participants and adverse reactions
We screened and enrolled 50 subjects (Fig. 1), and they were randomly allocated to either probiotic group (n = 25) or placebo group (n = 25). There were no differences in baseline characteristics between the two groups (Table 2). During the study, four subjects from the probiotic group and two from the placebo group withdrew consent for personal reasons.

Participant flowchart
Adverse events were reported from two patients in the probiotic group. One patient experienced dyspepsia and diarrhea, but recovered completely without any treatment. The other patient reported wrist sprain which is irrelevant to this study.
Dietary analysis
The investigators did not strictly encouraged participants to maintain caloric restriction nor give them specific guidelines for food, and our result is difficult to infer how eating habits have changed. But 24-h dietary recall analysis was investigated before and after the intervention to confirm whether subjects maintained a caloric restriction diet (Table 3). Results showed that there was no significant difference in energy intake between the two groups. Although energy intake did not change throughout the study, fat consumption significantly increased in both groups (mean difference of 2.35 g in the probiotic group and 3.77 g in the placebo group, p < 0.0001 for both groups). No significant difference in mineral intake was observed between two groups (Table 4). In both groups, calcium intake significantly increased (13.65 mg in the probiotic group and 27.48 mg in the placebo group), while iron intake significantly decreased (0.21 mg in the probiotic group and 0.64 mg in the placebo group, p < 0.0001). There was no significant change in vitamin intake, except for vitamin C (Table 5), which increased significantly in the probiotic group (mean difference of 24.49 mg, p = 0.027).
Body weight and composition changes
The primary outcome of this study was measured by body weight and body composition changes after 12 weeks of intervention (Table 6). The probiotic group showed a significant decrease in waist circumferences (mean difference of 1.88 cm, p = 0.049), while the fat percentage of the placebo group significantly increased (mean difference of 1.14%, p = 0.005). Internal fat area and ratio of visceral to subcutaneous fat area (V/S ratio) significantly decreased in both groups. Internal fat area decreased by an average of 6.47 cm2 in the probiotic group (p < 0.001) and 4.98 cm2 in the placebo group (p = 0.021). The V/S ratio dropped by 0.022 in the probiotic group (p < 0.001) and 0.015 in the placebo group (p = 0.013). The pulse rate of the probiotic group significantly decreased (mean difference of 3.4, p = 0.009).
Biochemical analyses
After 12 weeks of intervention, blood analyses indicated that there was no significant difference between two groups (Table 7). However, a few minor changes occurred. Fasting glucose concentration increased in the placebo group (mean difference of 3.88, p = 0.023). HDL-C concentration significantly increased in both the probiotic and placebo groups (mean differences of 2.72 and 5.28, respectively). Due to increased HDL-C concentration, total cholesterol/HDL-C ratio significantly increased in both groups. However, there were no significant differences between the two groups.
Gut microbial community analysis
Although our clinical results were based on ITT analysis, we performed a gut microbial analysis on pre- and post-intervention stool samples. Due to failure to acquire a fecal sample, one subject from the placebo group was excluded from this analysis.
Compositional changes in gut microbiota by probiotic or placebo supplementation were analyzed using 16S rRNA gene sequence analysis (Fig. 2). The Chao1 indices slightly decreased after administration of probiotic (p = 0.04) or placebo capsule (p = 0.0078). The Shannon and Simpson indices did not change significantly (Fig. 2a). Non-metric multidimensional scaling (NMDS) based on the Bray–Curtis distance matrices showed no obvious separation between the groups (Fig. 2b). There were no significant differences between groups or before–after comparison at phylum and family levels (Fig. 2c).

Diversity analysis between groups and their changes during intervention. a Alpha diversity (Chao1, Shannon, and Simpson metrics). Lines in box represent median alpha diversities. The box ranges from the lower to the upper quartiles. Statistical significance was determined by Wilcoxon paired t test. b Non-metric multidimensional scaling (NMDS) beta diversity plots using Bray–Curtis distance matrices. c Relative abundance of the top 5 most abundant phyla and the top 14 most abundant families in each group
When clustering gut microbiota before intervention, it was divided into two enterotypes according to Prevotella/Bacteroides ratio (Fig. 3a). Enterotype 1 (Ent1) had a relatively higher proportion of Prevotella (average = 24.18%, SD = 15.70%) than Bacteroides (average = 10.33%, SD = 5.97%). In the other hand, Enterotype 2 (Ent2) had relatively lower Prevotella (average = 0.55%, SD = 1.57%) than Bacteroides (average = 32.43%, SD = 14.43%). The Chao1 richness estimator significantly decreased in Ent1 after administering probiotic (p = 0.044) or placebo capsule (p = 0.0035) (Fig. 3b). While NMDS based on the Bray–Curtis distance matrices showed no obvious separation between the groups, the change in the Bray–Curtis distance of Ent1 after administering probiotics was significantly higher than the other group (Fig. 3c). Firmicutes and Bacteroidetes, as well as other rare phyla, were not significantly different between the probiotic–placebo, before–after, or Ent1–Ent2 comparisons (Fig. 3d). However, the relative abundance of Prevotellaceae in the Ent1 of the probiotic group was diminished—from an average of 35.66% (SD = 15.29%) before intervention to 20.99% (SD = 17.73%) after—but it was not significant.

Changes in microbial community according to enterotype. a Identified enterotypes and abundances of the main contributors for each enterotype. b Alpha diversity (Chao1, Shannon, and Simpson metrics). Lines in box represent median alpha diversities. The box ranges from the lower to the upper quartiles. Statistical significance was determined by Wilcoxon paired t test. c Non-metric multidimensional scaling (NMDS) beta diversities using Bray–Curtis distance matrices. Statistical significance was determined by Wilcoxon paired t test. d Relative abundance of the top 5 most abundant phyla and the top 14 most abundant families in each group
Differences in probiotic response per microbial enterotype
The differences in response per enterotype were identified by changes in waist circumferences and body fat composition (Table 8). Total fat area was significantly less in the Ent1 group with probiotics (mean difference of 19.46 cm2, p = 0.040), whereas fat percentage was significantly greater in the Ent1 of the placebo group (mean difference of 1.27%, p = 0.015). Internal fat area and V/S ratio significantly decreased in both Ent1 and Ent2 of the probiotic group. However, comparing Ent1 and Ent2 within probiotics group showed that internal fat area reduced much more in the Ent1 group, by an average of 12.50 cm2 (p = 0.002), compared with the Ent2 group, by an average of 4.10 cm2 (p = 0.029). The differential response according to enterotype was also identified by changes in biochemical markers (Table 9). TG significantly decreased in Ent1 of the probiotic group (mean difference of 16.22 mg/dL, p = 0.022). TC/HDL significantly decreased in Ent1 of the probiotic group (mean difference of 7.92, p = 0.0.036). Glucose significantly increased in Ent1 of the placebo group (mean difference of 5.37 mg/dL, p = 0.010). Finally, insulin significantly increased in Ent1 of the placebo group (mean difference of 2.47 μIU/mL, p = 0.0.039).
Regardless of the allocated group, baseline clinical parameters were compared to determine if there were any biomarkers to distinguish between Ent1 and Ent2. There were statistically significant differences between two groups in baseline glucose and insulin level. However, the differences were not clinically noticeable.
Changes in plasma metabolites
Multivariate statistical analysis from the metabolic profiling was performed with plasma samples obtained from the subjects before and after the study. The principal component analysis of plasma spectral data did not show clear clustering. Therefore, further analysis applied the orthogonal projections to latent structures discriminant analysis (OPLS-DA) model to distinctly demonstrate metabolite profiles (Fig. 4). The OPLS-DA score plots revealed that the probiotic group was not statistically distinguishable from the placebo group (R2X = 0.0803, R2Y = 0.0409 in the negative mode and R2X = 0.154 and R2Y = 0.32 in the positive mode). However, the probiotic and placebo groups after the study showed distinct clusters in the negative mode of OPLS-DA score plots, but there were no distinct clusters between the groups before the study.

Orthogonal projections to latent structures discriminant analysis (OPLS-DA) score plots based on serum spectral samples, obtained before and after taking either probiotic or placebo capsules for 12 weeks. a Operated in negative ion mode. b Operated in positive ion mode
Discussion
In this double-blinded, placebo-controlled, randomized clinical trial, we found that a 12-week administration of a multispecies probiotic significantly changed obesity-related markers, such as waist circumference, total fat area, visceral fat, V/S ratio, and biochemical markers, such as TG, TC.HDL, glucose, and insulin. These changes were more remarkable in the Prevotella-rich enterotype than in the Bacteroides-rich enterotype.
Obesity-related markers in response to probiotics treatment
Although the effect on weight loss was not significant, the results showed an improvement in other factors related to obesity, such as WC and total fat area. Our result is notable, because among many obesity-related parameters suggested to explain risk factors of metabolic diseases, WC is associated with an increased risk for metabolic diseases when the BMI is between 25 and 34.9 [58, 59]. A similar result was observed in a study providing Lactobacillus gasseri BNR17 supplementation for 12 weeks—significant reductions in WC and HC in the probiotic group—while a slight increase in fat percentage was observed in the placebo group [60].
Considerable evidence has shown that amount of visceral fat is a key etiological factor of cardiovascular diseases [61, 62]. Focusing on fat distribution, V/S ratio has emerged as a potential risk factor [63, 64]. In the current study, prominent reductions in visceral fat area and V/S ratio were found in both the probiotic and placebo groups. The mechanisms involved in reduction of visceral fat are not clear, but a few studies have reported the possible mechanisms. There was a human study that demonstrated a cholesterol- and glucose-lowering effect of L. plantarum [65]. The result was accompanied by a significant reduction in interleukin 6. Another study reported that Lactobacillus plantarum strain No.14 had a beneficial effect on obese mice by reducing adipocyte size [66]. This may explain our study result that showed reduction in WC was accompanied by a significant decrease in visceral fat. A similar study using a 12-week administration of Bifidobacterium animalis subsp. lactis GCL2505 for obese subjects found a significant decrease in abdominal visceral fat, which was accompanied by an increase in the total number of Bifidobacterium in fecal samples [67]. Another study—L. gasseri SBT2055 treatment for 12 weeks—showed beneficial effects on visceral fat area and WC, while the placebo group showed increasing body fat percentage and body fat mass [68].
Results from the 24-h dietary recall showed that there was no statistical difference between the two groups in energy intake. Interestingly, the intake of fat increased in both groups after the intervention. Despite the increased consumption of fat, obesity-related markers improved in the probiotic group, while the placebo group showed an increase in body fat percentage. This result is probably due to GP2, which presents a probiotic effect on obesity. For some reason, both groups showed significant decrease in iron intake and increase in calcium intake. Although our data from the 24-h dietary recall does not reflect habitual changes in diet, the changes of mineral intake are worthy of notice. Probiotics is known to have beneficial effect on host by increasing in the absorption of micronutrients such as calcium, iron, and magnesium [69]. An in vivo study found that a low-iron diet triggers iron-dependent microbiome, such as Lactobacillus species to absorb iron from the food and increase iron-storage protein ferritin [70]. In human studies, it is known that commensal bacteria do not need iron from the intestinal lumen, and rather excessive intake of iron leads to pathogenic intestinal flora. However, iron deficiency can cause a decrease in bacteria such as Bacteroides and Roseburia and an increase in Lactobacillus and Enterobacteriaceae [71]. In this context, low intake of iron in both groups might have affected bacterial homeostasis and Lactobacillus supplementation might have helped the modulation in probiotics group but not in the placebo group which might have consequently affected overall clinical conditions. However, our data from the dietary recall is not directly related to amount of iron absorbed from the food. Future study to evaluate whether probiotic intake affects mineral absorption is needed.
Bifidobacterium breve and Lactobacillus plantarum strains as a probiotics
The GP2, which we used in this study, is a multiple strain probiotic composed of Bifidobacterium breve and Lactobacillus plantarum. Bifidobacterium is capable of producing conjugated linoleic acids that have anti-obesity, anti-diabetic, anti-carcinogenic, and immune regulation properties [72,73,74]. Particularly, Bifidobacterium breve have effect on metabolic syndrome-suppressing and anti-obesity [75, 76]. Lactobacillus is classified as a lactic acid producing bacteria that play a major role in modulating gut ecology [77, 78]. Particularly, L. plantarum showed effect on treatment of vaginal disorders and reducing the body weight and cholesterol level [79,80,81,82]. Many experimental studies have clearly shown the anti-obesity effects of probiotics belonging to Lactobacillus and Bifidobacterium strains [83, 84].
Various probiotic characteristics such as gastric and bile acids tolerance and epithelial cell adhesion ability of currently used Lactobacillus plantarum CBT LP3 were previously reported [85]. Probiotic functions of L. plantarum CBT LP3 related to energy metabolism in obesity and reduction of atopic dermatitis symptoms were also previously reported [86, 87]. Currently used Bifidobacterium breve CBT BR3 strain was isolated from the feces of a healthy infant and several kinds of genetic factors associated with health promotion of the human host such as oligosaccharide degradation and vitamin biosynthesis were identified by complete genome sequence analysis of this strain [88]. Health promoting probiotic functions of B. breve CBT BR3 such as antimicrobial, anti-inflammatory, and anti-obesity activities also have been reported [87, 89,90,91]. For the development of anti-obesitic probiotics, we evaluated anti-obesity effect of various probiotic strains including L. plantarum CBT LP3 and B. breve CBT BR3 onto cell and animal model [92]. L. plantarum CBT LP3 and B. breve CBT BR3 showed higher efficacy than other tested strains and treatment of complex probiotics by these two strains was most effective. For the clinical trial, each volunteers served two probiotic capsules formulated with L. plantarum CBT LP3 BR3 (1.5 × 109 viable cells/2 capsules) and B. breve CBT BR3 (1.5 × 109 viable cells/2 capsules), and this daily dosage of Lactobacillus and Bifidobacterium spp. is accepted as probiotics when delivered in food at a level of 1 × 109 colony-forming units (CFU) per serving [93].
GP2 and placebo capsule contain fructo-oligosaccharide (FOS) and magnesium stearate. Magnesium stearate is commonly used as an inactive ingredient in the production of pharmaceutical tablets, capsules, and powders [94]. Prebiotics are defined as “a substrate that is selectively utilized by the host microorganisms conferring a health benefit” by the dedicated International Scientific Association for Probiotics and Prebiotics (ISAPP) expert group [95]. Fructans (FOS and inulin) and galactans (galactooligosaccharides or GOS) have dominated the prebiotic category and recently, nanoceria can be a new prebiotics to reduce cholesterol levels and restore gut microbiota in obese mice when composed with probiotics [26]. Whereas probiotics use live microorganisms, prebiotics are non-viable substrates that serve as nutrients for administered probiotic strains and indigenous microorganisms [95, 96]. In addition, prebiotics can be used together with specified probiotic treatments by synergetic effect to treat obesity and other diseases in view of predictive, preventive, and personalized concepts [26]. Prediction and optimization of the pre- and probiotic composition from individualized patient profiles enable the personalized treatment.
Immunity systems: relevant pathways
It has been well established that the immune system plays an important role in the pathogenesis of obesity [97]. Disrupted gut barrier function in obesity increases bacterial translocation such as LPS and LPS-binding protein, which stimulate the inflammation in adipose tissue [98, 99]. T cells are involved primarily in the cell-mediated immune response and T helper (Th) cells are key regulators [100]. Th1 and Th2 cytokine imbalance are reported in various disease states such as autoimmune disorders, asthma, and adipose tissue inflammation [101,102,103]. Lean adipose tissue contains balanced level of Th2 and regulatory T (Treg) cells that maintain macrophage activity and inhibit proinflammatory cells through cytokines such as IL-10, whereas obese adipose tissue contains many infiltrated proinflammatory cells such as Th1 cell and M1 macrophages [103]. Proinflammatory immune cells produce high amounts of inflammatory cytokines such as TNF-α, IL-6, and IL-1β and adipocytes produce high amounts of leptin, which promotes Th1 cells and inhibits Treg cells [103]. Adipose tissue inflammation by Th1/Th2 imbalance contributes to adipocyte hypertrophy and obesity-induced insulin resistance [104].
Probiotics are known to maintain a balance of Th1/Th2 immune at the level of receptor-ligand interactions by modulating immune response with bacterial cell wall components [105,106,107]. Lactobacillus and Bifidobacterium are Gram-positive bacteria whose cell wall is comprised of lipoprotein (LP) and lipoteichoic acid (LTA) embedded in thick layer of peptidoglycan (PGN) [108]. Previous study reported that strains in the genus Lactobacillus or Bifidobacterium induce the production of opposite cytokines IL-12 or IL-10, but combinational use of Lactobacillus and Bifidobacterium induced the production of IL-10 and promoted Treg cell polarizing through interaction of LP and LTA with TLR2 [109]. It is previously reported that currently used two strains, L. plantarum CBT LP3 and B. breve CBT BR3, have an anti-inflammatory effect by increasing IL-10 and Treg cell and decreasing IL-6 and TNF-α [86, 90]. Therefore, it is assumed that L. plantarum CBT LP3 and B. breve CBT may modulate Th1/Th2 balance and adipose tissue inflammation. Although this study did not directly confirm whether immune regulation of current probiotics strains affected to the reduction of obesity symptoms, we suggested Th1/Th2 balance recovery by current probiotics as a possible mechanism.
Although probiotics are believed to help the activation of host immune system and to maintain intestinal barrier function, there can be some risks of using probiotics. Risks associated with probiotics treatment include minor gastrointestinal discomfort such as gas and bloating. In rare cases, probiotics can cause infections but cases of infection due to Lactobacillus and Bifidobacterium are extremely rare in immunocompromised subjects [110]. In our study, only one patient reported mild dyspepsia and diarrhea, and there was no sign for immunological problems. So far, concerns for immune deviation or excessive immune stimulation caused by probiotics are particularly relevant in the field of pregnant women and neonates who are vulnerable in immune-related diseases [111]. Currently, it is impossible to propose risk of autoimmune diseases and who is likely to have side effects. Since it is difficult to know which probiotics strains are suitable for each individual, dividing people by enterotype can be one strategy for prevention of autoimmune diseases after long-term probiotic consumption.
Enterotype as a potential microbiome biomarker
Recognizing that physicians should consider individual variability, precision medicine has shifted the focus on clinical trials from thousands-people to single-person measurement, so called N-of-1 trial [112]. True personalized medicine is based on the individual patient profile [113]. Although studies on individual genetic variation have been conducted to increase treatment efficiency, genetic variation alone is not sufficient to account for individual differences [114]. Considering meta-analysis studies, it is difficult to derive irrefutable conclusions about the anti-obesity effects of probiotics in clinical trials [115,116,117]. In clinical studies, various factors, such as diet, ethnicity, and individual differences in gut microbiome profiles, make it difficult to acquire consistent results from probiotics intake [84, 115]. To minimize individual differences and obtain effective results, it is necessary to evaluate by enterotypes to consider the subjects’ gut microbiome profile.
Enterotype is influenced by diverse factors such as heredity and living environment. Among them, it is mostly affected by food, and enterotypes are not easily altered by short-term dietary changes [42]. Advances in gut microbiome study have found that different metabolic responses appear with the same treatment depending on the enterotype [118, 119], and stratified analysis per enterotype is proposed for obesity management [120]. In this study, we identified positive responses of obesity-related parameters in the Prevotella-rich enterotype. Visceral fat showed a significant decrease in both enterotypes but a much larger decline in the Prevotella-rich enterotype than in Bacteroides-rich enterotype. Additionally, the Prevotella-rich enterotype showed decreased TG and total fat area after administering probiotics, while blood glucose, insulin, and fat ratios increased in the placebo group. Consistently in other studies, the Prevotella-rich enterotype exhibited an effective response to weight loss [47, 48]. However, other studies have shown that acarbose, an anti-diabetic drug, and capsaicin, an active compounds of chili pepper, improved metabolic parameters better in the Bacteroides-rich enterotype than in the Prevotella-rich enterotype [119, 121]. In our study, the inflammation level, C-reactive protein, was elevated in the placebo group of the Bacteroides-rich enterotype. It may be appear that the specific enterotype was not sensitive to response, but different enterotypes react according to different treatments. Due to the small sample size of our study, it was difficult to subdivide the enterotypes. However, in addition to the enterotypes represented by Prevotella and Bacteroides, enterotypes represented by Ruminococcus or Enterobacteriaceae have also been reported [43, 44], so detailed analysis of the enterotype in various and large-scale models is necessary.
Although we have found that probiotics can have a beneficial effect on obese subjects with Prevotella-rich enterotype, our result showed rather opposite results within placebo group. Subjects with Prevotella-rich enterotype showed increased glucose and insulin, while subjects with Bacteroides-rich enterotype did not show significant changes in glucose homeostasis. Because the placebo capsules we used in this study contain FOS, it might have affected the results itself. Both Prevotella and Bacteroides are well known as fiber-utilizing bacteria, and that the dominance of Prevotella or Bacteroides can differentially impact short-chain fatty acid (SCFA) productions. There was a study comparing fiber-utilizing capacity of Prevotella and Bacteroides with different fibers including FOS and they showed Prevotella-dominated microbiota produces more SCFA than Bacteroides-dominated microbiota [122]. Although it is well known that SCFA produced by microbiome have important effects on glucose homeostasis by range of mechanisms including improvement of gut barrier function and improved pancreatic β-cell function [123, 124], our result shows that just because there are a lot of SCFA-producing microbiota does not directly result in lowering the blood glucose and insulin levels. Moreover, our results may emphasize the importance of probiotics that it is better to take the prebiotics with the probiotics even if the subjects have abundant SCFA-producing bacteria in their gut.
Predictive, preventive, and personalized medicine
The recent progress in predictive, preventive, and personalized medicine (PPPM) has led to a paradigm shift from the uniform treatment for each disease to optimal health care taking account the characteristics of each patient [113]. Great potential for application of microbiome has been studied and implemented in personalized diets and disease prevention areas [125,126,127]. Enterotype classification speculates the reactivity and provides a personalized treatment for the individuals through the differential responses to drugs and diets [128,129,130]. We suggested that Lactabacillus and Bifidobacterium strains might be a supplementary therapy for preventing obesity in specific enterotype. Development of next probiotics for each enterotype is necessary to prevent obesity in personalized approach. Moreover, enterotype is potential microbiome biomarker. Enterotype-specific responses to the probiotics can be useful for predicting probiotics effect and developing personalized probiotics in predictive medicine approach. Obesity-related markers based on enterotype can help to stratify patients with obesity by predicting clinically relevant developments and facilitate to personalized individualized treatments and preventive measures. Enterotype-based PPPM might be applicable to the prevention and treatment of various diseases as well as obesity.
Study limitations
Some limitations should be addressed to interpret the findings of this study. Firstly, because our study aims to evaluate effect of GP2 on general adult population with obesity, the inclusion criteria was broad. It is well known that gender, age, ethnicity, and menopausal status can affect obesity prevalence as well as composition of gut microbiome [131,132,133]. To minimize confounding factors that influence both obesity and gut microbiota, individual clinical studies by age and gender groups are needed. Secondly, there may be limitations in the verification of the precise effects of probiotics strains. When making probiotics commercially, prebiotics such as FOS are usually added. We also add FOS when we make placebo capsules, so we can exclude bias by prebiotics. However, we think we should have performed clinical trials with probiotics with only strains in order to verify the effect of GP2. Thirdly, the study did not explain which pathways are involved with the effects. We suggested that the immune system plays an important role in anti-obesity effect of probiotics. Further researches of the molecular mechanisms underlying the effects of probiotics are needed to clarify the effects of probiotics and provide guidance for probiotic treatment as personalized and precision medicine.
Conclusions
In conclusion, this double-blinded, placebo-controlled, randomized clinical trial showed that GP2, a probiotic formulation containing L. plantarum and B. breve, changed obesity-related markers and biochemical markers. These changes differed per enterotype and were more pronounced in the Prevotella-rich enterotype than in the Bacteroides-rich enterotype. Nonetheless, we did not elucidate the exact mechanism for these enterotype-specific reactions. However, our study first reported a different response to the probiotics according to the enterotype, as well as the improvement of obesity-related markers by probiotics supplementation. Our findings provide important information for the use of probiotics to ameliorate the symptoms of obesity and for the application of probiotics in consideration of gut microbial enterotypes to treat obesity in a personalized way. In the future, gut microbial enterotype should be carefully examined to develop predictive diagnostics methods, targeted prevention, and personalized of medical services for obesity and other diseases.
References
Agha M, Agha R. The rising prevalence of obesity: part A: impact on public health. Int J Surg Oncol (N Y). 2017;2(7):e17. https://doi.org/10.1097/IJ9.0000000000000017.
Van Gaal LF, Mertens IL, De Block CE. Mechanisms linking obesity with cardiovascular disease. Nature. 2006;444(7121):875–80. https://doi.org/10.1038/nature05487.
Chan JM, Rimm EB, Colditz GA, Stampfer MJ, Willett WC. Obesity, fat distribution, and weight gain as risk factors for clinical diabetes in men. Diabetes Care. 1994;17(9):961–9.
Global BMIMC, Di Angelantonio E, Bhupathiraju S, Wormser D, Gao P, Kaptoge S, et al. Body-mass index and all-cause mortality: individual-participant-data meta-analysis of 239 prospective studies in four continents. Lancet. 2016;388(10046):776–86. https://doi.org/10.1016/S0140-6736(16)30175-1.
Anandacoomarasamy A, Fransen M, March L. Obesity and the musculoskeletal system. Curr Opin Rheumatol. 2009;21(1):71–7. https://doi.org/10.1097/bor.0b013e32831bc0d7.
Harris R, Card TR, Delahooke T, Aithal GP, Guha IN. Obesity is the most common risk factor for chronic liver disease: results from a risk stratification pathway using transient elastography. Am J Gastroenterol. 2019;114(11):1744–52. https://doi.org/10.14309/ajg.0000000000000357.
Dietrich P, Hellerbrand C. Non-alcoholic fatty liver disease, obesity and the metabolic syndrome. Best Pract Res Clin Gastroenterol. 2014;28(4):637–53. https://doi.org/10.1016/j.bpg.2014.07.008.
Castro AM. Macedo-de la concha LE, Pantoja-Meléndez CA. Low-grade inflammation and its relation to obesity and chronic degenerative diseases. Revista Médica del Hospital General de México. 2017;80(2):101–5. https://doi.org/10.1016/j.hgmx.2016.06.011.
Lord GM, Matarese G, Howard JK, Baker RJ, Bloom SR, Lechler RI. Leptin modulates the T-cell immune response and reverses starvation-induced immunosuppression. Nature. 1998;394(6696):897–901. https://doi.org/10.1038/29795.
Youssef DM, Elbehidy RM, Shokry DM, Elbehidy EM. The influence of leptin on Th1/Th2 balance in obese children with asthma. J Bras Pneumol. 2013;39(5):562–8. https://doi.org/10.1590/S1806-37132013000500006.
Rogero MM, Calder PC. Obesity, inflammation, Toll-like receptor 4 and fatty acids. Nutrients. 2018;10(4):432. https://doi.org/10.3390/nu10040432.
Galli C, Calder PC. Effects of fat and fatty acid intake on inflammatory and immune responses: a critical review. Ann Nutr Metab. 2009;55(1–3):123–39. https://doi.org/10.1159/000228999.
Simopoulos AP, DiNicolantonio JJ. The importance of a balanced ω-6 to ω-3 ratio in the prevention and management of obesity. Open Heart. 2016;3(2):e000385–e. https://doi.org/10.1136/openhrt-2015-000385.
Roberts CK, Barnard RJ. Effects of exercise and diet on chronic disease. J Appl Physiol (Bethesda, Md : 1985). 2005;98(1):3–30. https://doi.org/10.1152/japplphysiol.00852.2004.
Feinberg AP, Irizarry RA, Fradin D, Aryee MJ, Murakami P, Aspelund T, et al. Personalized epigenomic signatures that are stable over time and covary with body mass index. Sci Transl Med. 2010;2(49):49ra67. https://doi.org/10.1126/scitranslmed.3001262.
Alonso R, Farías M, Alvarez V, Cuevas A. Chapter 7 - the genetics of obesity. In: Rodriguez-Oquendo A, editor. Translational cardiometabolic genomic medicine. Boston: Academic Press; 2016. p. 161–77.
Romieu I, Dossus L, Barquera S, Blottiere HM, Franks PW, Gunter M, et al. Energy balance and obesity: what are the main drivers? Cancer Causes Control : CCC. 2017;28(3):247–58. https://doi.org/10.1007/s10552-017-0869-z.
Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444(7122):1027–31. https://doi.org/10.1038/nature05414.
Turnbaugh PJ, Gordon JI. The core gut microbiome, energy balance and obesity. J Physiol. 2009;587(Pt 17):4153–8. https://doi.org/10.1113/jphysiol.2009.174136.
Thaiss CA, Zmora N, Levy M, Elinav E. The microbiome and innate immunity. Nature. 2016;535:65–74. https://doi.org/10.1038/nature18847.
Turnbaugh PJ, Hamady M, Yatsunenko T, Cantarel BL, Duncan A, Ley RE, et al. A core gut microbiome in obese and lean twins. Nature. 2008a;457:480. https://doi.org/10.1038/nature07540 https://www.nature.com/articles/nature07540#supplementary-information.
Turnbaugh PJ, Bäckhed F, Fulton L, Gordon JI. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe. 2008b;3(4):213–23. https://doi.org/10.1016/j.chom.2008.02.015.
Turnbaugh PJ, Ridaura VK, Faith JJ, Rey FE, Knight R, Gordon JI. The effect of diet on the human gut microbiome: a metagenomic analysis in humanized gnotobiotic mice. Sci Transl Med. 2009;1(6):6ra14–6ra. https://doi.org/10.1126/scitranslmed.3000322.
Hughes RL, Marco ML, Hughes JP, Keim NL, Kable ME. The role of the gut microbiome in predicting response to diet and the development of precision nutrition models—part I: overview of current methods. Adv Nutr. 2019a;10(6):953–78. https://doi.org/10.1093/advances/nmz022.
Hughes RL, Kable ME, Marco M, Keim NL. The role of the gut microbiome in predicting response to diet and the development of precision nutrition models. Part II: Results. Adv Nutr. 2019b;10(6):979–98. https://doi.org/10.1093/advances/nmz049.
Bubnov R, Babenko L, Lazarenko L, Kryvtsova M, Shcherbakov O, Zholobak N, et al. Can tailored nanoceria act as a prebiotic? Report on improved lipid profile and gut microbiota in obese mice. EPMA J. 2019;10(4):317–35. https://doi.org/10.1007/s13167-019-00190-1.
Markowiak P, Śliżewska K. Effects of probiotics, prebiotics, and Synbiotics on human health. Nutrients. 2017;9(9):1021. https://doi.org/10.3390/nu9091021.
Wang L, Guo M-J, Gao Q, Yang J-F, Yang L, Pang X-L, et al. The effects of probiotics on total cholesterol: a meta-analysis of randomized controlled trials. Medicine. 2018;97(5):e9679–e. https://doi.org/10.1097/MD.0000000000009679.
Collado MC, Meriluoto J, Salminen S. Role of commercial probiotic strains against human pathogen adhesion to intestinal mucus. Lett Appl Microbiol. 2007;45(4):454–60. https://doi.org/10.1111/j.1472-765X.2007.02212.x.
Hill C, Guarner F, Reid G, Gibson GR, Merenstein DJ, Pot B, et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol. 2014;11(8):506–14. https://doi.org/10.1038/nrgastro.2014.66.
Bubnov RV, Babenko LP, Lazarenko LM, Mokrozub VV, Demchenko OA, Nechypurenko OV, et al. Comparative study of probiotic effects of Lactobacillus and Bifidobacteria strains on cholesterol levels, liver morphology and the gut microbiota in obese mice. EPMA J. 2017;8(4):357–76. https://doi.org/10.1007/s13167-017-0117-3.
Park D-Y, Ahn Y-T, Park S-H, Huh C-S, Yoo S-R, Yu R, et al. Supplementation of Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032 in diet-induced obese mice is associated with gut microbial changes and reduction in obesity. PLoS One. 2013;8(3):e59470. https://doi.org/10.1371/journal.pone.0059470.
Huang Y, Wang J, Quan G, Wang X, Yang L, Zhong L. <span class="named-content genus-species" id="named-content-1">Lactobacillus acidophilus</span> ATCC 4356 prevents atherosclerosis via inhibition of intestinal cholesterol absorption in apolipoprotein e-knockout mice. Appl Environ Microbiol. 2014;80(24):7496. https://doi.org/10.1128/AEM.02926-14.
Aronsson L, Huang Y, Parini P, Korach-André M, Håkansson J, Gustafsson J-Å, et al. Decreased fat storage by Lactobacillus paracasei is associated with increased levels of angiopoietin-like 4 protein (ANGPTL4). PLoS One. 2010;5(9):e13087. https://doi.org/10.1371/journal.pone.0013087.
Ma X, Hua J, Li Z. Probiotics improve high fat diet-induced hepatic steatosis and insulin resistance by increasing hepatic NKT cells. J Hepatol. 2008;49(5):821–30. https://doi.org/10.1016/j.jhep.2008.05.025.
Hidalgo-Cantabrana C, Delgado S, Ruiz L, Ruas-Madiedo P, Sánchez B, Margolles A. Bifidobacteria and their health-promoting effects. Microbiol Spectr. 2017;5(3). https://doi.org/10.1128/microbiolspec.BAD-0010-2016.
Thomson P, Medina DA, Garrido D. Human milk oligosaccharides and infant gut bifidobacteria: molecular strategies for their utilization. Food Microbiol. 2018;75:37–46. https://doi.org/10.1016/j.fm.2017.09.001.
Ruiz L, Delgado S, Ruas-Madiedo P, Sánchez B, Margolles A. Bifidobacteria and their molecular communication with the immune system. Front Microbiol. 2017;8:2345. https://doi.org/10.3389/fmicb.2017.02345.
Yatsunenko T, Rey FE, Manary MJ, Trehan I, Dominguez-Bello MG, Contreras M, et al. Human gut microbiome viewed across age and geography. Nature. 2012;486(7402):222–7. https://doi.org/10.1038/nature11053.
Moschen AR, Wieser V, Tilg H. Dietary factors: major regulators of the gut’s microbiota. Gut Liver. 2012;6(4):411–6. https://doi.org/10.5009/gnl.2012.6.4.411.
Wu GD, Chen J, Hoffmann C, Bittinger K, Chen YY, Keilbaugh SA, et al. Linking long-term dietary patterns with gut microbial enterotypes. Science. 2011;334(6052):105–8. https://doi.org/10.1126/science.1208344.
Lim MY, Rho M, Song Y-M, Lee K, Sung J, Ko G. Stability of gut enterotypes in Korean monozygotic twins and their association with biomarkers and diet. Sci Rep. 2014;4:7348. https://doi.org/10.1038/srep07348 https://www.nature.com/articles/srep07348#supplementary-information.
Liang C, Tseng H-C, Chen H-M, Wang W-C, Chiu C-M, Chang J-Y, et al. Diversity and enterotype in gut bacterial community of adults in Taiwan. BMC Genomics. 2017;18(1):932. https://doi.org/10.1186/s12864-016-3261-6.
Arumugam M, Raes J, Pelletier E, Le Paslier D, Yamada T, Mende DR, et al. Enterotypes of the human gut microbiome. Nature. 2011;473(7346):174–80. https://doi.org/10.1038/nature09944.
de Moraes AC, Fernandes GR, da Silva IT, Almeida-Pititto B, Gomes EP, Pereira AD, et al. Enterotype may drive the dietary-associated cardiometabolic risk factors. Front Cell Infect Microbiol. 2017;7:47. https://doi.org/10.3389/fcimb.2017.00047.
Roager HM, Licht TR, Poulsen SK, Larsen TM, Bahl MI. Microbial enterotypes, inferred by the prevotella-to-bacteroides ratio, remained stable during a 6-month randomized controlled diet intervention with the new nordic diet. Appl Environ Microbiol. 2014;80(3):1142–9. https://doi.org/10.1128/aem.03549-13.
Hjorth MF, Roager HM, Larsen TM, Poulsen SK, Licht TR, Bahl MI, et al. Pre-treatment microbial Prevotella-to-Bacteroides ratio, determines body fat loss success during a 6-month randomized controlled diet intervention. Int J Obes. 2017;42:580–3. https://doi.org/10.1038/ijo.2017.220.
Hjorth MF, Blaedel T, Bendtsen LQ, Lorenzen JK, Holm JB, Kiilerich P, et al. Prevotella-to-Bacteroides ratio predicts body weight and fat loss success on 24-week diets varying in macronutrient composition and dietary fiber: results from a post-hoc analysis. Int J Obes (Lond). 2019;43(1):149–57. https://doi.org/10.1038/s41366-018-0093-2.
Organization WH. The Asia-Pacific perspective: redefining obesity and its treatment. Sydney: Health Communications Australia; 2000.
Organization WH. Waist circumference and waist-hip ratio: report of a WHO expert consultation, Geneva, 8-11 December 2008. 2011.
Perry AC, Applegate EB, Jackson ML, Deprima S, Goldberg RB, Ross R, et al. Racial differences in visceral adipose tissue but not anthropometric markers of health-related variables. J Appl Physiol (Bethesda, Md : 1985). 2000;89(2):636–43. https://doi.org/10.1152/jappl.2000.89.2.636.
Friedewald WT, Levy RI, Fredrickson DS. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin Chem. 1972;18(6):499–502.
Lim MY, Song E-J, Kim SH, Lee J, Nam Y-D. Comparison of DNA extraction methods for human gut microbial community profiling. Syst Appl Microbiol. 2018;41(2):151–7. https://doi.org/10.1016/j.syapm.2017.11.008.
Bolger AM, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics. 2014;30(15):2114–20. https://doi.org/10.1093/bioinformatics/btu170.
Zhang J, Kobert K, Flouri T, Stamatakis A. PEAR: a fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics. 2014;30(5):614–20. https://doi.org/10.1093/bioinformatics/btt593.
Edgar RC, Haas BJ, Clemente JC, Quince C, Knight R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics. 2011;27(16):2194–200. https://doi.org/10.1093/bioinformatics/btr381.
Bolyen E, Rideout JR, Dillon MR, Bokulich NA, Abnet C, Al-Ghalith GA, et al. QIIME 2: reproducible, interactive, scalable, and extensible microbiome data science. Peer J Preprints. 2018;6:e27295v2. https://doi.org/10.7287/peerj.preprints.27295v2.
Janssen I, Katzmarzyk PT, Ross R. Body mass index, waist circumference, and health risk: evidence in support of current National Institutes of Health guidelines. Arch Intern Med. 2002;162(18):2074–9.
Janssen I, Katzmarzyk PT, Ross R. Waist circumference and not body mass index explains obesity-related health risk. Am J Clin Nutr. 2004;79(3):379–84.
Jung SP, Lee KM, Kang JH, Yun SI, Park HO, Moon Y, et al. Effect of Lactobacillus gasseri BNR17 on overweight and obese adults: a randomized, double-blind clinical trial. Korean J Fam Med. 2013;34(2):80–9. https://doi.org/10.4082/kjfm.2013.34.2.80.
Matsuzawa Y, Nakamura T, Shimomura I, Kotani K. Visceral fat accumulation and cardiovascular disease. Obes Res. 1995;3(Suppl 5):645s–7s.
Despres JP. Cardiovascular disease under the influence of excess visceral fat. Crit Pathw Cardiol. 2007;6(2):51–9. https://doi.org/10.1097/HPC.0b013e318057d4c9.
Fukuda T, Bouchi R. Ratio of visceral-to-subcutaneous fat area predicts cardiovascular events in patients with type 2 diabetes. J Diabetes Investig. 2018;9:396-402. https://doi.org/10.1111/jdi.12713.
Kaess BM, Pedley A, Massaro JM, Murabito J, Hoffmann U, Fox CS. The ratio of visceral to subcutaneous fat, a metric of body fat distribution, is a unique correlate of cardiometabolic risk. Diabetologia. 2012;55(10):2622–30. https://doi.org/10.1007/s00125-012-2639-5.
Barreto FM, Colado Simão AN, Morimoto HK, Batisti Lozovoy MA, Dichi I, Helena da Silva Miglioranza L. Beneficial effects of Lactobacillus plantarum on glycemia and homocysteine levels in postmenopausal women with metabolic syndrome. Nutrition 2014;30(7):939–942. doi:https://doi.org/10.1016/j.nut.2013.12.004.
Takemura N, Okubo T, Sonoyama K. Lactobacillus plantarum strain No. 14 reduces adipocyte size in mice fed high-fat diet. Exp Biol Med (Maywood, NJ). 2010;235(7):849–56. https://doi.org/10.1258/ebm.2010.009377.
Takahashi S, Anzawa D, Takami K, Ishizuka A, Mawatari T, Kamikado K, et al. Effect of Bifidobacterium animalis ssp. lactis GCL2505 on visceral fat accumulation in healthy Japanese adults: a randomized controlled trial. Biosci Microbiota Food Health. 2016;35(4):163–71. https://doi.org/10.12938/bmfh.2016-002.
Kadooka Y, Sato M, Imaizumi K, Ogawa A, Ikuyama K, Akai Y, et al. Regulation of abdominal adiposity by probiotics (Lactobacillus gasseri SBT2055) in adults with obese tendencies in a randomized controlled trial. Eur J Clin Nutr. 2010;64(6):636–43. https://doi.org/10.1038/ejcn.2010.19.
Kortman GAM, Raffatellu M, Swinkels DW, Tjalsma H. Nutritional iron turned inside out: intestinal stress from a gut microbial perspective. FEMS Microbiol Rev. 2014;38(6):1202–34. https://doi.org/10.1111/1574-6976.12086.
Das NK, Schwartz AJ, Barthel G, Inohara N, Liu Q, Sankar A, et al. Microbial metabolite signaling is required for systemic iron homeostasis. Cell Metabolism. 2019;S1550–4131(19):30560–1. https://doi.org/10.1016/j.cmet.2019.10.005.
Skrypnik K, Suliburska J. Association between the gut microbiota and mineral metabolism. J Sci Food Agric. 2018;98(7):2449–60. https://doi.org/10.1002/jsfa.8724.
Raimondi S, Amaretti A, Leonardi A, Quartieri A, Gozzoli C, Rossi M. Conjugated linoleic acid production by Bifidobacteria: screening, kinetic, and composition. Biomed Res Int. 2016;2016:8654317. https://doi.org/10.1155/2016/8654317.
Kim JH, Kim Y, Kim YJ, Park Y. Conjugated linoleic acid: potential health benefits as a functional food ingredient. Annu Rev Food Sci Technol. 2016a;7:221–44. https://doi.org/10.1146/annurev-food-041715-033028.
Patterson E, Wall R, Lisai S, Ross RP, Dinan TG, Cryan JF, et al. Bifidobacterium breve with α-linolenic acid alters the composition, distribution and transcription factor activity associated with metabolism and absorption of fat. Sci Rep. 2017;7:43300. https://doi.org/10.1038/srep43300 https://www.nature.com/articles/srep43300#supplementary-information.
Kondo S, Kamei A, Xiao JZ, Iwatsuki K, Abe K. Bifidobacterium breve B-3 exerts metabolic syndrome-suppressing effects in the liver of diet-induced obese mice: a DNA microarray analysis. Benefic Microbes. 2013;4(3):247–51. https://doi.org/10.3920/BM2012.0019.
Kondo S, Xiao J-z, Satoh T, Odamaki T, Takahashi S, Sugahara H, et al. Antiobesity effects of Bifidobacterium breve strain B-3 supplementation in a mouse model with high-fat diet-induced obesity. Biosci Biotechnol Biochem. 2010;74(8):1656–61. https://doi.org/10.1271/bbb.100267.
Molly K, Smet ID, Nollet L, Woestyne MV, Verstraete W. Effect of lactobacilli on the ecology of the gastro-intestinal microbiota cultured in the SHIME reactor. Microb Ecol Health Dis. 1996;9(2):79–89. https://doi.org/10.3109/08910609609166446.
Walter J. Ecological role of lactobacilli in the gastrointestinal tract: implications for fundamental and biomedical research. Appl Environ Microbiol. 2008;74(16):4985–96. https://doi.org/10.1128/AEM.00753-08.
Wu C-C, Weng W-L, Lai W-L, Tsai H-p, Liu W-H, Lee M-H et al. Effect of Lactobacillus plantarum strain K21 on high-fat diet-fed obese mice. Evid Based Complement Alternat Med. 2015;2015:391767. doi:DOI:https://doi.org/10.1155/2015/391767.
Million M, Angelakis E, Paul M, Armougom F, Leibovici L, Raoult D. Comparative meta-analysis of the effect of Lactobacillus species on weight gain in humans and animals. Microb Pathog. 2012;53(2):100–8. https://doi.org/10.1016/j.micpath.2012.05.007.
Michael DR, Davies TS, Moss JWE, Calvente DL, Ramji DP, Marchesi JR, et al. The anti-cholesterolaemic effect of a consortium of probiotics: an acute study in C57BL/6J mice. Sci Rep. 2017;7(1):2883. https://doi.org/10.1038/s41598-017-02889-5.
Mastromarino P, Macchia S, Meggiorini L, Trinchieri V, Mosca L, Perluigi M, et al. Effectiveness of Lactobacillus-containing vaginal tablets in the treatment of symptomatic bacterial vaginosis. Clin Microbiol Infect. 2009;15(1):67–74. https://doi.org/10.1111/j.1469-0691.2008.02112.x.
Kobyliak N, Conte C, Cammarota G, Haley AP, Styriak I, Gaspar L, et al. Probiotics in prevention and treatment of obesity: a critical view. Nutri Metab. 2016;13:14. https://doi.org/10.1186/s12986-016-0067-0.
Brusaferro A, Cozzali R, Orabona C, Biscarini A, Farinelli E, Cavalli E, et al. Is it time to use probiotics to prevent or treat obesity? Nutrients. 2018;10(11). https://doi.org/10.3390/nu10111613.
X-M L, HA L, M K, E-S P, K-Y P. Probiotic effects of Lactobacillus plantarum strains isolated from Kimchi. J Korean Soc Food Sci Nutr. 2016;45(12):1717–24.
Kim HW, Hong R, Choi EY, Yu K, Kim N, Hyeon JY, et al. A probiotic mixture regulates T cell balance and reduces atopic dermatitis symptoms in mice. Front Microbiol. 2018a;9(2414). https://doi.org/10.3389/fmicb.2018.02414.
Lee SJ, Bose S, Seo J-G, Chung W-S, Lim C-Y, Kim H. The effects of co-administration of probiotics with herbal medicine on obesity, metabolic endotoxemia and dysbiosis: a randomized double-blind controlled clinical trial. Clin Nutr. 2014;33(6):973–81. https://doi.org/10.1016/j.clnu.2013.12.006.
Kwak M-J, Yoon J-K, Kwon S-K, Chung M-J, Seo J-G, Kim JF. Complete genome sequence of the probiotic bacterium Bifidobacterium breve KCTC 12201BP isolated from a healthy infant. J Biotechnol. 2015;214:156–7. https://doi.org/10.1016/j.jbiotec.2015.09.035.
Shin J-H, Chung M-J, Seo J-G. A multistrain probiotic formulation attenuates skin symptoms of atopic dermatitis in a mouse model through the generation of CD4(+)Foxp3(+) T cells. Food Nutr Res. 2016;60:32550. https://doi.org/10.3402/fnr.v60.32550.
Kim MS, Byun JS, Yoon YS, Yum DY, Chung MJ, Lee JC. A probiotic combination attenuates experimental colitis through inhibition of innate cytokine production. Benefic Microbes. 2016b;8(2):231–41. https://doi.org/10.3920/BM2016.0031.
J-S L, M-J C, J-G S. In vitro evaluation of antimicrobial activity of lactic acid bacteria against Clostridium difficile. Toxicological Research. 2013;29(2):99–106.
Chung M-J. Efficacy and safety evaluation of anti-obesity probiotics. Unpublished raw data. 2017.
Canada Go. Accepted claims about the nature of probiotic microorganisms in food. 2019. http://www.hc-sc.gc.ca/fn-an/label-etiquet/claims-reclam/probiotics_claims-allegations_probiotiques-eng.php%20.
Hobbs CA, Saigo K, Koyanagi M, Hayashi S-M. Magnesium stearate, a widely-used food additive, exhibits a lack of in vitro and in vivo genotoxic potential. Toxicol Rep. 2017;4:554–9. https://doi.org/10.1016/j.toxrep.2017.10.003.
Gibson GR, Hutkins R, Sanders ME, Prescott SL, Reimer RA, Salminen SJ, et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat Rev Gastroenterol Hepatol. 2017;14(8):491–502. https://doi.org/10.1038/nrgastro.2017.75.
Reid G, Abrahamsson T, Bailey M, Bindels LB, Bubnov R, Ganguli K, et al. How do probiotics and prebiotics function at distant sites? Benefic Microbes. 2017;8(4):521–33. https://doi.org/10.3920/BM2016.0222.
Andersen CJ, Murphy KE, Fernandez ML. Impact of obesity and metabolic syndrome on immunity. Adv Nutr. 2016;7(1):66–75. https://doi.org/10.3945/an.115.010207.
Cheru L, Saylor CF, Lo J. Gastrointestinal barrier breakdown and adipose tissue inflammation. Curr Obes Rep. 2019;8(2):165–74. https://doi.org/10.1007/s13679-019-00332-6.
Chang C-C, Sia K-C, Chang J-F, Lin C-M, Yang C-M, Huang K-Y, et al. Lipopolysaccharide promoted proliferation and adipogenesis of preadipocytes through JAK/STAT and AMPK-regulated cPLA2 expression. Int J Med Sci. 2019;16(1):167–79. https://doi.org/10.7150/ijms.24068.
Moss RB, Moll T, El-Kalay M, Kohne C, Soo Hoo W, Encinas J, et al. Th1/Th2 cells in inflammatory disease states: therapeutic implications. Expert Opin Biol Ther. 2004;4(12):1887–96. https://doi.org/10.1517/14712598.4.12.1887.
Charlton B, Lafferty KJ. The Th1/Th2 balance in autoimmunity. Curr Opin Immunol. 1995;7(6):793–8. https://doi.org/10.1016/0952-7915(95)80050-6.
Mazzarella G, Bianco A, Catena E, De Palma R, Abbate GF. Th1/Th2 lymphocyte polarization in asthma. Allergy. 2000;55(Suppl 61):6–9. https://doi.org/10.1034/j.1398-9995.2000.00511.x.
Han JM, Levings MK. Immune regulation in obesity-associated adipose inflammation. J Immunol. 2013;191(2):527. https://doi.org/10.4049/jimmunol.1301035.
Richardson VR, Smith KA, Carter AM. Adipose tissue inflammation: feeding the development of type 2 diabetes mellitus. Immunobiology. 2013;218(12):1497–504. https://doi.org/10.1016/j.imbio.2013.05.002.
Savcheniuk OA, Virchenko OV, Falalyeyeva TM, Beregova TV, Babenko LP, Lazarenko LM, et al. The efficacy of probiotics for monosodium glutamate-induced obesity: dietology concerns and opportunities for prevention. EPMA J. 2014;5(1):2. https://doi.org/10.1186/1878-5085-5-2.
Tymoshok NO, Lazarenko LM, Bubnov RV, Shynkarenko LN, Babenko LP, Mokrozub VV, et al. New aspects the regulation of immune response through balance Th1/Th2 cytokines. EPMA J. 2014;5(1):A134. https://doi.org/10.1186/1878-5085-5-S1-A134.
Мokrozub VV, Lazarenko LM, Sichel LM, Babenko LP, Lytvyn PM, Demchenko OM, et al. The role of beneficial bacteria wall elasticity in regulating innate immune response. EPMA J. 2015;6(1):13–5. https://doi.org/10.1186/s13167-015-0035-1.
Shockman GD, Barren JF. Structure, function, and assembly of cell walls of gram-positive bacteria. Annu Rev Microbiol 1983;37(1):501–527. doi:https://doi.org/10.1146/annurev.mi.37.100183.002441.
Zeuthen LH, Fink LN, Frøkiær H. Toll-like receptor 2 and nucleotide-binding oligomerization domain-2 play divergent roles in the recognition of gut-derived lactobacilli and bifidobacteria in dendritic cells. Immunology. 2008;124(4):489–502. https://doi.org/10.1111/j.1365-2567.2007.02800.x.
Borriello SP, Hammes WP, Holzapfel W, Marteau P, Schrezenmeir J, Vaara M, et al. Safety of probiotics that contain lactobacilli or bifidobacteria. Clin Infect Dis. 2003;36(6):775–80. https://doi.org/10.1086/368080.
Boyle RJ, Robins-Browne RM, Tang MLK. Probiotic use in clinical practice: what are the risks? Am J Clin Nutr. 2006;83(6):1256–447. https://doi.org/10.1093/ajcn/83.6.1256.
Schork NJ. Personalized medicine: time for one-person trials. Nature. 2015;520(7549):609–11. https://doi.org/10.1038/520609a.
Golubnitschaja O, Baban B, Boniolo G, Wang W, Bubnov R, Kapalla M, et al. Medicine in the early twenty-first century: paradigm and anticipation - EPMA position paper 2016. EPMA J. 2016;7(1):23. https://doi.org/10.1186/s13167-016-0072-4.
Nebert DW, Zhang G, Vesell ES. From human genetics and genomics to pharmacogenetics and pharmacogenomics: past lessons. Future Directions Drug Metabolism Reviews. 2008;40(2):187–224. https://doi.org/10.1080/03602530801952864.
John GK, Wang L, Nanavati J, Twose C, Singh R, Mullin G. Dietary alteration of the gut microbiome and its impact on weight and fat mass: a systematic review and meta-analysis. Genes. 2018;9(3). https://doi.org/10.3390/genes9030167.
Park S, Bae J-H. Probiotics for weight loss: a systematic review and meta-analysis. Nutr Res. 2015;35(7):566–75. https://doi.org/10.1016/j.nutres.2015.05.008.
Borgeraas H, Johnson LK, Skattebu J, Hertel JK, Hjelmesaeth J. Effects of probiotics on body weight, body mass index, fat mass and fat percentage in subjects with overweight or obesity: a systematic review and meta-analysis of randomized controlled trials. Obes Rev. 2018;19(2):219–32. https://doi.org/10.1111/obr.12626.
Korem T, Zeevi D, Zmora N, Weissbrod O, Bar N, Lotan-Pompan M, et al. Bread affects clinical parameters and induces gut microbiome-associated personal glycemic responses. Cell Metab. 2017;25(6):1243–53.e5. https://doi.org/10.1016/j.cmet.2017.05.002.
Kang C, Zhang Y, Zhu X, Liu K, Wang X, Chen M, et al. Healthy subjects differentially respond to dietary capsaicin correlating with specific gut enterotypes. J Clin Endocrinol Metab. 2016;101(12):4681–9. https://doi.org/10.1210/jc.2016-2786.
Christensen L, Roager HM, Astrup A, Hjorth MF. Microbial enterotypes in personalized nutrition and obesity management. Am J Clin Nutr. 2018a;108(4):645–51. https://doi.org/10.1093/ajcn/nqy175.
Gu Y, Wang X, Li J, Zhang Y, Zhong H, Liu R, et al. Analyses of gut microbiota and plasma bile acids enable stratification of patients for antidiabetic treatment. Nat Commun. 2017a;8(1):1785. https://doi.org/10.1038/s41467-017-01682-2.
Chen T, Long W, Zhang C, Liu S, Zhao L, Hamaker BR. Fiber-utilizing capacity varies in Prevotella- versus Bacteroides-dominated gut microbiota. Scientific reports. 2017;7(1):2594. https://doi.org/10.1038/s41598-017-02995-4.
Morrison DJ, Preston T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes. 2016;7(3):189–200. https://doi.org/10.1080/19490976.2015.1134082.
Kim YA, Keogh JB, Clifton PM. Probiotics, prebiotics, synbiotics and insulin sensitivity. Nutr Res Rev. 2018b;31(1):35–51. https://doi.org/10.1017/S095442241700018X.
Zmora N, Zeevi D, Korem T, Segal E, Elinav E. Taking it personally: personalized utilization of the human microbiome in health and disease. Cell Host Microbe. 2016;19(1):12–20. https://doi.org/10.1016/j.chom.2015.12.016.
Bubnov RV, Babenko LP, Lazarenko LM, Mokrozub VV, Demchenko OA, Nechypurenko OV, et al. Comparative study of probiotic effects of Lactobacillus and Bifidobacteria strains on cholesterol levels, liver morphology and the gut microbiota in obese mice. EPMA J. 2017;8(4):357–76. https://doi.org/10.1007/s13167-017-0117-3.
Bubnov RV, Spivak MY, Lazarenko LM, Bomba A, Boyko NV. Probiotics and immunity: provisional role for personalized diets and disease prevention. EPMA J. 2015b;6(1):14. https://doi.org/10.1186/s13167-015-0036-0.
Christensen L, Roager HM, Astrup A, Hjorth MF. Microbial enterotypes in personalized nutrition and obesity management. Am J Clin Nutr. 2018b;108(4):645–51. https://doi.org/10.1093/ajcn/nqy175.
Costea PI, Hildebrand F, Arumugam M, Bäckhed F, Blaser MJ, Bushman FD, et al. Enterotypes in the landscape of gut microbial community composition. Nat Microbiol. 2018;3(1):8–16. https://doi.org/10.1038/s41564-017-0072-8.
Gu Y, Wang X, Li J, Zhang Y, Zhong H, Liu R, et al. Analyses of gut microbiota and plasma bile acids enable stratification of patients for antidiabetic treatment. Nat Commun. 2017b;8(1):1785. https://doi.org/10.1038/s41467-017-01682-2.
de la Cuesta-Zuluaga J, Kelley ST, Chen Y, Escobar JS, Mueller NT, Ley RE, et al. Age- and sex-dependent patterns of gut microbial diversity in human adults msystems. 2019;4(4):e00261–19. https://doi.org/10.1128/mSystems.00261-19.
Zhang Q, Wang Y. Socioeconomic inequality of obesity in the United States: do gender, age, and ethnicity matter? Soc Sci Med. 2004;58(6):1171–80. https://doi.org/10.1016/s0277-9536(03)00288-0.
Santos-Marcos JA, Rangel-Zuñiga OA, Jimenez-Lucena R, Quintana-Navarro GM, Garcia-Carpintero S, Malagon MM, et al. Influence of gender and menopausal status on gut microbiota. Maturitas. 2018;116:43–53. https://doi.org/10.1016/j.maturitas.2018.07.008.
Funding
This research was supported by Main Research Program (E0170601-03) of the Korea Food Research Institute (KFRI) funded by the Ministry of Science and ICT.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Ethical approval
This study was conducted according to the principles of Declaration of Helsinki and good clinical practice guidelines. The study protocol was approved by the Institutional Review Board of Ilsan Dongguk University Hospital (approval number 2016-02) and registered in Clinical Research Information Service (CRIS identifier: KCT0002292). Written informed consent has also been obtained from all the participants.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Song, EJ., Han, K., Lim, TJ. et al. Effect of probiotics on obesity-related markers per enterotype: a double-blind, placebo-controlled, randomized clinical trial. EPMA Journal 11, 31–51 (2020). https://doi.org/10.1007/s13167-020-00198-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13167-020-00198-y