
Abstract
Artificially drained commercial forests are hydrologically novel ecosystems, where the array of aquatic habitats consists of ditches and remnant pools. In general the network of ditches has been found to have longer hydroperiod, the knowledge, however, about aquatic invertebrates in this system is scarce. We examined which environmental factors are impacting the biomass and abundance of functional feeding groups. Scrapers and shredders were aggregated to ditches and gatherers to pools. Filterers’ distribution pattern suggested that the function of filtering is carried out by different taxa in pools and ditches. Ditches were rather more suitable for feeding groups that rely on autochthonous resources. Acidity was a major driver of functional community composition, for example, one of the causes for higher scraper frequency in ditches. Predators exhibited greater quantities in extensive macrophyte cover regardless of water body type. Our results suggest that the trophic organization in ditches and pools is different because habitat factors select the feeding groups directly through food resources, but also because of the environmental filter on the other biological traits of the organisms. To support complex ecosystems with several trophic levels also in commercial forests, we suggest to avoid destroying macrophyte rich pools and ditches during silvicultural management.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Forestry drainage has been regionally widespread practice for at least 40 years, transforming large areas of (semi)open wetlands and swamps into productive forests (Paavilainen and Päivänen 1995). Forestry drainage is accomplished by excavation of open ditches that lead away excess surface water, lower the ground water table and as a result, improve conditions for tree growth (Laine et al. 1995). Such practices have long-term and largely irreversible repercussions, such as changes in hydrological and disturbance regime, homogenization of landscape elements, disappearance of wetland-dependent species, reduction of peat soils, and alteration of stand structure and composition (Lõhmus et al., 2015; Paavilainen and Päivänen, 1995). Such novel landscapes are presently encompassing 20–25% of all the forest areas in top-drained regions: Finland, the Baltic States, and parts of Sweden (Paavilainen and Päivänen 1995); in Canada and the US State of North Carolina (Skaggs et al. 2016). Even more, the effects of modified hydrology extend beyond the drained sites to the ecosystems that receive the runoff water (Vuori et al. 1998).
Although forestry drainage does not unambiguously decrease the habitat availability for aquatic organisms, it shortens the hydroperiod of natural pools (Remm et al. 2015; Suislepp et al. 2011) and partly replaces natural water bodies with linear ditch networks (Remm et al. 2015). Ditches have relatively longer hydroperiod (Remm et al. 2015) and connectivity adds the possibility to escape from local drying. In the context of seminatural forestry, the taxonomic composition of macroinvertebrates is similar in ditches and in water bodies in undrained forests; but the gamma diversity is much lower in pools that remain in drained forests (Vaikre et al. 2015). Apart from this, the knowledge about aquatic invertebrates in artificially drained forests is scarce and studies on functional communities non-existent (Lõhmus et al. 2015).
Functional traits (biological traits) of organisms – well-defined, measurable characteristics that influence species performance in given environment, reflect the response of biota to the environment or its effects to ecosystem processes (McGill et al. 2006; Nock et al. 2016) and could be a useful tool aiding conservation and restoration activities (Diaz and Cabido 2001; Nock et al. 2016). Hydroperiod length may determine the distribution of functional feeding groups that rely on autochthonous resources and thus the differences in functional communities between ditches and pools in a drained forest ecosystem. The abundance and species richness of periphytic algae may increase with shorter hydroperiod, but the abundance of diatoms – critical nutritional component of periphyton (Anderson and Cummins 1979) – is higher in water bodies with longer hydroperiod (Gottlieb et al. 2006; Rober et al. 2013) as is the amount of phytoplankton (Boven et al. 2008).
However, certain environmental factors may be more important than hydroperiod in shaping functional communities. For example, in sun exposed ponds and ditches macrophyte beds favor shredders and scrapers, providing food such as plant litter or algae growing on vascular plants, whereas gathering-collectors predominate in sediments (Bazzanti 2015; Bazzanti et al. 2009; Leslie and Lamp 2017; Sychra et al. 2010). In forest streams, shredders’ habitat is mainly formed in concurrence of tree cover with boulders and logs, the latter increasing retentiveness (Haapala et al. 2003; Houghton et al. 2011; Masese et al. 2014), whereas increases in sediment loads, originating from ditching in catchment area, may shift the community to dominance of collector-filterers (Vuori and Joensuu 1996).
In current study, we explore functional feeding groups of macroinvertebrates inhabiting ditches and pools in artificially drained commercial forests, where the whole ecosystem, including the pools, are hydrologically altered. Specifically we aim to:
1) Compare functional feeding communities between ditches and pools. We hypothesize that feeding groups that benefit from autochthonously produced resources (scrapers and filterers) are mostly aggregated to ditches.
2) Measure the responses of functional feeding groups to environmental gradients and find out which contemporary and local habitat factors could explain the possible differences in functional communities between ditches and pools.
Material and Methods
Study Area and Design
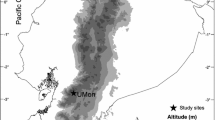
Our study was conducted in Estonia, where large-scale systematized drainage was carried out in the 1950–1980s and resulted in approximately 25% of forests impacted by drainage (Torim and Sults 2005). Data originate from three artificially drained commercial forest plots (total area 239 ha) (Fig. 1) located on Gleysols and to a smaller extent on shallow peat soils. The tree stand consists of native species: birch (dominant on 39% of the area), pine (32%) and spruce (17%). Prevalent forest site types (Lõhmus 1984) were Vaccinium, Oxalis, Aegopodium, Filipendula, mesotrophic bog, and stagnant water swamp. Clear cuts with stands younger than five years covered 2.6% of the area and stands older than 60 years 31%. Forestry drainage ditches comprised 98% of the network of lotic water bodies (76 m/ha) and have not been cleaned or reconstructed since the digging. Ditches were mostly overgrown with Sphagnum mosses and macrophytes and in respect of minor flow velocity resembled pools rather than streams. Only three ditches from our study plots were visibly flowing. For this reason we assumed that hydroperiod would be the main factor causing differences in functional communities between ditches and pools.

Locations of study plots in Estonia. Left panel shows the sampling locations (triangles for ditches and squares for pools) in one of the study plots
Macroinvertebrates were sampled in May 2013 and 2014, 26 days after the mean air temperature rose over +5 °C, i.e., the beginning of growing season. From each study plot we selected 10 ditches and 10 pools (natural floodings, wheel rut pools etc.; with depth > 15 cm), from different forest site types according to the relative area of the site type. We dip-netted an area of 4 m2 for 20 s with 0.5 mm mesh D-frame net (17 × 19 cm) in each water body. Samples were preserved on-site in 96% ethanol. We recorded 12 environmental variables from each water body that could influence the arrangement of macroinvertebrates’ functional communities (see Table 1). We used 57 of the sampled water bodies from both year for analyses.
Invertebrates were sorted from the detritus, counted, and identified in laboratory. We estimated the abundance of very numerous taxa (> 100 or > 10 specimens in a 1/25 subsample) using 3/25 or 5/25 of the sample respectively. Individuals were identified to lowest possible taxonomic level, mostly to species or genus, except for Psychodidae, which were identified to family level. We assigned each taxon to functional feeding group (Appendix) following Merritt et al. (1996), Heino (2000), Paunović et al. (2006), Zilli et al. (2008) and Bazzanti et al. (2009). In case the adults and larvae of the same taxon exhibiting different feeding modalities, we assigned them to different functional groups. As species of Chironomidae represent many different feeding strategies and we were not able to determine them further than to family level, we excluded this group from the analyses.
To describe the quantity of functional feeding groups in community level, we used abundance (the number of individuals) and biomass per sample. Abundance has been the most applied metrics in functional group analyses and is believed to reflect the interspecific patterns of resource division (Magurran 1988). However, in case of species with wide variety of body masses, the relationship between abundance and functional impact may be over- or underestimated. Also, energy flow through trophic levels are more strongly connected to the total biomass than the abundance (Brown et al. 2004; Saint-Germain et al. 2007). Moreover, abundance and biomass have shown to have different responses to environmental conditions (Laux and Torgan 2015).
The total biomass of each taxon in a water body was calculated from individual lengths or head widths using equations, preferably from similar and geographically close habitats (Baumgärtner and Rothhaupt 2003; Benke et al. 1999; Edwards et al. 2009; Haas et al. 2007; Mährlein et al. 2016; Méthot et al. 2012). Dry mass of the individuals of a species in a water body was estimated using all the collected specimens (if N < 20) or 20 specimens and extrapolated to others. Dry mass of molluscs was considered without the shell. For Oligochaeta we estimated the dry mass using cylindrical volume of pieces or whole individuals according to Schwoerbel (1994) and Haas et al. (2007). In case the individuals were accidentally discarded after determination (23%), we used constant species specific weights or constant lengths with equation (according to Timm (1999) or consulting with Tarmo Timm).
Data Analysis
Differences in functional group composition were evaluated using two-way factorial permutation-based nonparametric MANOVA (PerMANOVA) analysis with Sørensen dissimilarity and ‘water body type’ (fixed) and ‘plot’ as factors. Biomass and richness data from both years were summed up. Community matrices were then relativized dividing the quantity of a certain feeding group in a sample with the total quantity of the feeding group in all water bodies. Whereas PerMANOVA requires balanced design, we randomly selected some water bodies to be deleted and ran the analysis 50 times using batch command with sample size of 54.
To find out whether the abundance and biomass of feeding groups differ between water body types and which habitat factors influence given group, we used general linear models (GLM). Abundance and biomass data were log-transformed to fit the normality assumption. Models were built according to the following procedure: (1) test for the significance (GLM: p < 0.15) of each habitat factor and water body type; (2) search for correlating habitat factors (Spearman correlation: p < 0.05); (3) multifactorial GLM with significant and non-correlating factors. The best model was defined as the one with lowest Akaike’s information criterion value and all the habitat factors significant (p < 0.05) in multifactorial GLM. Finally, water body type, when significant in first step, was added to the best model to see whether it loses its significance, i.e. habitat factors explain the difference between ditches and pools. Year and plot were included in all models as independent factors, but to find the correlating factors (step 2) all the observations were merged. Scrapers were absent in a large part of samples, therefore we built models for two subsets: for the samples containing scrapers (N = 70) and, by logistic regression with occurrence data for the whole dataset (N = 114). Statistical analyses were carried out using PC-ord 6.07 (McCune and Mefford 2011) and Statistica 7.
Results
The biomass, abundance and taxa richness varied among feeding groups (Fig. 2; Appendix). We observed two distribution patterns resulting in large total biomass of a feeding group: (i) very large individuals abundant only in a few and absent from majority of water bodies; (ii) individuals with medium biomass numerous in majority of the water bodies. First distribution pattern was typical to scrapers in ditches, e.g., gastropods Planorbis planorbis, Aplexa hypnorum and Lymnaea stagnalis, whereas the overall taxa richness of scrapers was small. Second pattern characterizes shredders in ditches, including generalistic species such as waterlouse Asellus aquaticus, stonefly Nemoura cinerea and caddisflies Limnephilus stigma and Trichostegia minor. Similarly abundant and ubiquitous were filterers, though they had high biomass only in ditches. There 70% of the filterer individuals were bivalves (mostly Pisidium spp.), but in pools 66% of individuals belonged to Nematocera (e.g., Aedes, Culex, Culiseta and Dixella sp). Feeding groups with lowest total biomass — predators and gatherers — had highest taxa richness and were moderately abundant.

Mean abundance (number of individuals) and biomass with standard errors of functional feeding groups per sample collected during 20 s from 4 m2 in each water body. For abbrevations see Table 2
Functional group composition differed between water body types and plots in regards of biomass (PerMANOVA: F = 7.8–2.5; p < 0.006) and abundance (F = 7.6–2.6; p < 0.004). Scrapers occurrence (GLZ first step: χ2 = 4.3; p = 0.037), abundance (GLM first step: F = 14.9; p < 0.001), and biomass (F = 23.7; p < 0.001) was significantly higher in ditches. Ditches had also higher mean abundance (F = 20.3; p < 0.001) and biomass (F = 8.0; p = 0.005) of shredders and biomass of filterers (F = 7.1; p = 0.008), while pools had higher biomass (F = 5.9; p = 0.016) of gatherers. Neither abundance nor biomass of predators differed between water body types (Table 2, Fig. 2). Ditches and pools did not differ conspicuously in respect of habitat factors (Table 1). Better than water body type, water depth and graminoid litter explained the biomass of gatherers; and pH and shade explained the frequency of scrapers. In other cases, water body type remained significant in multifactorial model (Table 2). In case of scraper abundance, pH lost its significance (in first step p = 0.001) if together with water body type in the model, which suggests that it is probably one of the reasons for greater abundance of scrapers in ditches. This also occurred with water depth in case of gatherers abundance (in first step p = 0.045).
Discussion
Our study shows that the variability of macroinvertebrate functional communities and thus functioning of aquatic ecosystems, including the difference between the networks of ditches and isolated pools, is remarkable in artificially drained forests. Ditches and pools had distinctly different feeding group composition – scrapers, filterers (and less contrastingly shredders) were aggregated to ditches and gatherers to pools. This supports our hypothesis that the longer hydroperiod of ditches (Remm et al. 2015; Suislepp et al. 2011) favors feeding groups that rely on autochthonously produced recourses. Surprisingly, water depth did not explain this difference, and thus is not a good proxy for the length of hydroperiod in such small water bodies.
Acidity was a major driver of functional community composition and also one of the causes for higher scraper frequency in ditches. The variation in pH probably is caused both by mosaic of soil types, and soil disturbance during artificial drainage. Higher pH in ditches compared to small forest pools is common in Estonia (Remm et al. 2015). This was also evident in our study plots (though statistically not significant), where the cover (15%) and depth (< 1 m; according to soil map provided by the Estonian Land Board) of peat layer was small, hence, ditches were easily penetrated down to mineral soil during digging. The positive scraper-pH correlation is probably not derived from the amount of food as increasing water acidity does not necessarily limit periphyton production (Greenwood & Lowe 2006). Low pH imposes respiratory and osmoregulatory challenges to aquatic invertebrates in general (Harrison 2001). Scrapers were further limited in our study system, because gastropods made up more than 99% of their quantity. Shell formation and reproduction are depending on calcium-rich habitat with high pH (Dillon 2000). In the context of our study plots snails were mostly absent from water bodies with pH lower than 5.
We found more gatherers in shallow water bodies, especially pools, where the bottom was covered with litter from terrestrial or semiaquatic plants: tree leaves and graminoids. Although long hydroperiod supports rich autochthonous food base (Boven et al. 2008), gatherers are not dependent on that and are instead adept to use allochthonous litter (Kraus and Vonesh 2012). Shallow waters could favor this group, as litter from trees and herbaceous plants may decompose faster and have higher protein content when exposed to air (Bärlocher et al. 1978). Additionally, invertebrates from this group (e.g., Cloeon dipterum, Anacaena lutescens) often exhibited traits that allow them to exploit temporary water bodies.
Filterers attained higher biomass in ditches, while mean abundance was equally distributed, suggesting that the function of filtering is carried out by different taxa in different water bodies or biodiversity based redundancy (Schmera et al. 2017) ensuring the stability of the effect. Indeed, relatively heavy bivalves were mostly aggregated to ditches, and were replaced with Nematocera in pools. Mosquitoes often exhibit life history strategies that allow them to utilize temporary water bodies and some taxa even relay on drought for oviposition (Wiggins et al. 1980). Water acidity affected only filterers’ biomass, suggesting it controls foremost bivalves that are pH-sensitive also in lakes and rivers (Mackie and Flippance 1983; Saunders and Kling 1990).
The quantities of Shredders, which were greater in ditches, correlated positively with pH, water depth, graminoids and macrophyte cover. Surprisingly, the shade or amount of leaf litter had no effect on this group. Experiments in forest stream mesocosms have shown higher abundances of several shredder species following increased leaf litter input, but not necessarily increase in their total abundance (Melody and Richardson 2004; Richardson 1991). It might be supposed that (1) the nutritional quality of leaf litter is more limiting for shredders than the quantity; and (2) periphyton (i.e., essential food resource for shredders; Cummins and Klug 1979) abundance and nutritional quality is enhanced by high water pH (Mulholland et al. 1991) and permanent inundation (Aspbury and Juliano 1998; Inkley et al. 2008; but see (Bärlocher et al. 1978). These correlations may be even stronger because the presence of leaf litter from overstory vegetation can increase acidity by leaching of the humic acids while decomposing (Stoler and Relyea 2011).
The abundance of predators was somewhat higher in pools yet biomass did not differentiate. Such pattern emerged because this group obtained high numbers only in some pools, which functioned as breeding places for Chaoboridae — a Family of small-bodied Diptera. Top-down control (predators as a proportion of total of all other functional groups; (Merritt et al. 1996) may be lower in ditches, considering the higher biomass of other feeding groups there. Predators exhibited greater quantities in water bodies with extensive macrophyte cover, which seems to be more important than water body type. Macrophyte beds are preferred by invertebrates that avoid dry phase via migration to permanent water bodies (such as many species of Coleoptera and Hemiptera; (Bazzanti 2015; Bazzanti et al. 2009). Their dispersal ability allows them to benefit from three dimensional structurally complex mesohabitat (Bazzanti et al. 2009). Thus, to support complex ecosystems with several trophic levels also in commercial forests, we suggest to carefully avoid draining, cleaning, and destroying the macrophyte rich pools and ditches.
Our study showed that ditches and pools in commercial forests support various ecosystem function providers. The trophic organization in ditches and pools is different because habitat factors select feeding groups directly through food resources, but also because of the environmental filter on other biological traits of the organisms. The possible results of ditch maintenance: reduction of food amount during ditch cleaning and increase of sun exposure by removing ditch-side vegetation, would probably lead to shifts in functional communities. These, together with the comparison of the functioning of water-bodies in drained forest with natural wetland ecosystems, remain the issues for further studies.
References
Anderson NH, Cummins KW (1979) The influences of diet on the life histories of aquatic insects. Journal of the Fisheries Research Board of Canada 36:335–342
Aspbury AS, Juliano SA (1998) Negative effects of habitat drying and prior exploitation on the detritus resource in an ephemeral aquatic habitat. Oecologia 115:137–148
Bärlocher K, Mackay RJ, Wiggins GB (1978) Detritus processing in a temporary vernal pool in southern Ontario. Arch Hydrobiol 1:269–295
Baumgärtner D, Rothhaupt K-O (2003) Predictive length-dry mass regressions for freshwater invertebrates in a pre-alpine lake littoral. International Review of Hydrobiology 88:453–463
Bazzanti M (2015) Pond macroinvertebrates of the Presidential Estate of Castelporziano (Rome): a review of ecological aspects and selecting indicator taxa for conservation. Rendiconti Lincei 26:337–343
Bazzanti M, Della Bella V, Grezzi F (2009) Functional characteristics of macroinvertebrate communities in Mediterranean ponds (Central Italy): influence of water permanence and mesohabitat type. Annales de Limnologie-International Journal of Limnology 45:29–39
Benke AC, Huryn AD, Smock LA, Wallace JB (1999) Length-mass relationships for freshwater macroinvertebrates in North America with particular reference to the South-eastern United States. Journal of the North American Benthological Society 18:308–343
Boven L, Stoks R, Forró L, Brendonck L (2008) Seasonal dynamics in water quality and vegetation cover in temporary pools with variable hydroperiods in Kiskunság (Hungary). Wetlands 28:401–410
Brown JH, Gillooly JF, Allen AP, Savage VM, West GB (2004) Toward a metabolic theory of ecology. Ecology 85:1771–1789
Cummins KW, Klug MJ (1979) Feeding ecology of stream invertebrates. Annual Review of Ecology and Systematics 10:147–172
Diaz S, Cabido M (2001) Vive la difference: plant functional diversity matters to ecosystem processes. Trends Ecol Evol 16:646–655
Dillon RT (2000) The ecology of freshwater molluscs. Cambridge University Press
Edwards FK, Lauridsen RB, Armand L, Vincent HM, Jones JI (2009) The relationship between length, mass and preservation time for three species of freshwater leeches (Hirudinea). Fundamental and Applied Limnology 173:321–327
Gottlieb AD, Richards JH, Gaiser EE (2006) Comparative study of periphyton community structure in long and short-hydroperiod Everglades marshes. Hydrobiologia 569:195–207
Greenwood JL, Lowe RL (2006) The effects of pH on a periphyton community in an acidic wetland, USA. Hydrobiologia 561:71–82
Haapala A, Muotka T, Laasonen P (2003) Distribution of benthic macroinvertebrates and leaf litter in relation to streambed retentivity: implications for headwater stream restoration. Boreal Environment Research 8:19–30
Haas K, Köhler U, Diehl S, Köhler P, Dietrich S, Holler S, Jaensch A, Nidermaier M, Vilsmeier J (2007) Influence of fish on habitat choice of water birds: a whole system experiment. Ecology 88:2915–2925
Harrison JF (2001) Insect acid-base physiology. Annual Review of Entomology 46:221–250
Heino J (2000) Lentic macroinvertebrate assemblage structure along gradients in spatial heterogeneity, habitat size and water chemistry. Hydrobiologia 418:229–242
Houghton DC, Berry EA, Gilchrist A, Thompson J, Nussbaum MA (2011) Biological changes along the continuum of an agricultural stream: influence of a small terrestrial preserve and use of adult caddisflies in biomonitoring. Journal of Freshwater Ecology 26:381–397
Inkley MD, Wissinger SA, Baros BL (2008) Effects of drying regime on microbial colonization and shredder preference in seasonal woodland wetlands. Freshwater Biology 53:435–445
Kraus JM, Vonesh JR (2012) Fluxes of terrestrial and aquatic carbon by emergent mosquitoes: a test of controls and implications for cross-ecosystem linkages. Oecologia 170:1111–1122
Laine J, Vasander H, Sallantaus T (1995) Ecological effects of peatland drainage for forestry. Environmental Reviews 3:286–303
Laux M, Torgan LC (2015) Which metric to choose? Differences between abundance and biomass responses to environmental conditions in a planktonic diatom community. Hydrobiologia 744:63–76
Leslie AW, Lamp WO (2017) Taxonomic and functional group composition of macroinvertebrate assemblages in agricultural drainage ditches. Hydrobiologia 787:99–110
Lõhmus E (1984) Estonian Forest Site Types. Estonian SSR Agro-Industrial Consortium Information Board. In: Tallinn
Lõhmus A, Remm L, Rannap R (2015) Just a ditch in forest? Reconsidering draining in the context of sustainable forest management. BioScience 65:1066–1076
Mackie GL, Flippance LA (1983) Intra- and interspecific variations in calcium content of freshwater Mollusca in relation to calcium content of the water. Journal of Molluscan Studies 49:204–212
Magurran AE (1988) Ecological diversity and its measurement. Croom Helm, London
Mährlein M, Pätzig M, Brauns M (2016) Length-mass relationships for lake macroinvertebrates corrected for back-transformation and preservation effects. Hydrobiologia 768:37–50
Masese FO, Kitaka N, Kipkemboi J, Gettel GM, Irvine K, McClain ME (2014) Macroinvertebrate functional feeding groups in Kenyan highland streams: Evidence for a diverse shredder guild. Freshwater Science 33:435–450
McCune B, Mefford MJ (2011) PC-ORD. Multivariate analysis of ecological data. Version 6.0. MjM Software, Oregon
McGill BJ, Enquist BJ, Weiher E, Westoby M (2006) Rebuilding community ecology from functional traits. Trends in Ecology & Evolution 21:178–185
Melody KJ, Richardson JS (2004) Responses of invertebrates and algae of a boreal coniferous forest stream to experimental manipulation of leaf litter inputs and shading. Hydrobiologia 519:197–206
Merritt RW, Wallace JR, Higgins MJ, Alexander MK, Berg MB, Morgan WT, Cummins KW, Vandeneeden B (1996) Procedures for the functional analysis of invertebrate communities of the Kissimmee river-floodplain ecosystem. Florida Scientist 59:216–274
Méthot G, Hudon C, Gagnon P, Pinel-Alloul B, Armellin A, Poirier AMT (2012) Macroinvertebrate size-mass relationships: how specific should they be? Freshwater Science 31:750–764
Mulholland PJ, Steinman AD, Palumbo AV, Elwood JW, Kirschtel DB (1991) Role of nutrient cycling and herbivory in regulating periphyton communities in laboratory streams. Ecology 72:966–982
Nock CA, Vogt RJ, Beisner BE (2016) Functional Traits. John Wiley & Sons Ltd, Chichester
Paavilainen E, Päivänen J (1995) Peatland forestry ecology and principles. Springer-Verlag, New York
Paunović M, Csányi B, Simić V, Stojanović B, Cakić P (2006) Distribution of Anodonta (Sinanodonta) woodiana (Rea, 1834) in inland waters of Serbia. Aquatic Invasions 1:154–160
Remm L, Lõhmus A, Rannap R (2015) Temporary and small water bodies in human-impacted forests: an assessment in Estonia. Boreal Environment Research 20:603–619
Richardson JS (1991) Seasonal food limitation of detritivores in a montane stream: an experimental test. Ecology 72(3):873–887
Rober AR, Wyatt KH, Turetsky MR, Stevenson RJ (2013) Algal community response to experimental and interannual variation in hydrology in an Alaskan boreal fen. Freshwater Science 32(1):1–11
Saint-Germain M, Buddle CM, Larrivée M, Mercado A, Motchula T, Reichert E, Sackett TE, Sylvain Z, Webb A (2007) Should biomass be considered more frequently as a currency in terrestrial arthropod community analyses? Journal of Applied Ecology 44:330–339
Saunders JF, Kling GW (1990) Species distributions and shell characteristics of Pisidium (Mollusca. Bivalvia) in the Colorado Front Range: The role of abiotic factors. Freshwater Biology 24:275–285
Schmera D, Heino J, Podani J (2017) Functional diversity: a review of methodology and current knowledge in freshwater macroinvertebrate research. Hydrobiologia 87:27–44
Schwoerbel J (1994) Methoden der Hydrobiologie. Süßwasserbiologie. Stuttgart, Jena, New York, Fischer
Skaggs RW, Tian S, Chescheir GM, Amatya DM, Youssef MA (2016) Forest drainage. In: Amatya DM, Williams TM, Bren L, de JC (eds) Forest hydrology: processes, management and assessment. CABI, pp 124–140
Stoler AB, Relyea RA (2011) Living in the litter: the influence of tree leaf litter on wetland communities. Oikos 120(6):862–872
Suislepp K, Rannap R, Lõhmus A (2011) Impacts of artificial drainage on amphibian breeding sites in hemiboreal forests. Forest Ecology and Management 262:1078–1083
Sychra J, Adámek Z, Petřivalská K (2010) Distribution and diversity of littoral macroinvertebrates within extensive reed beds of a lowland pond. Annales de Limnologie-International Journal of Limnology 46:281–289
Timm T (1999) A guide to the Estonian Annelidae. Estonian Naturalists´ Society, Tartu
Torim T, Sults Ü (2005) Main environmental problems of forest drainage. In: Arukuusk A, Etverk I (eds) 55 years of mechanized forest drainage in Estonia. Estonian Forest Society, pp 55–57
Vaikre M, Remm L, Rannap R (2015) Macroinvertebrate diversity and community structure in woodland pools and ditches and their response to artificial drainage. Hydrobiologia 762:157–168
Vuori K-M, Joensuu I (1996) Impacts of forest draining on the macroinvertebrates of a small boreal headwater stream: do buffer zones protect lotic biodiversity? Biological Conservation 77:87–95
Vuori K-M, Joensuu I, Latvala J, Jutila E, Ahvonen A (1998) Forest drainage: a threat to benthic biodiversity of boreal headwater streams? Aquatic Conservation: Marine and Freshwater Ecosystems 8:745–759
Wiggins GB, Mackay RJ, Smith IM (1980) Evolutionary and ecological strategies of animals in annual temporary pools. Archiv fur Hydrobiologie 58:97–206
Zilli FL, Montalto L, Marchese MR (2008) Benthic invertebrate assemblages and functional feeding groups in the Parana river floodplain. Limnologica 38:159–171
Acknowledgements
We are grateful to Tarmo Timm for his help in faunal identification. The research was funded by the European Union through the European Regional Development Fund (program 3.2.0802.11-0043) and the Estonian Research Council (grants no 9051 and IUT 34-7).
Author information
Authors and Affiliations
Corresponding author
Electronic Supplementary Material
ESM 1
(DOCX 33 kb)
Rights and permissions
About this article

Cite this article
Vaikre, M., Remm, L., Rannap, R. et al. Functional Assemblages of Macroinvertebrates in Pools and Ditches in Drained Forest Landscape. Wetlands 38, 957–964 (2018). https://doi.org/10.1007/s13157-018-1045-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-018-1045-2