
Abstract
Soil organic carbon (OC) is sensitive to climatic change, and it can be expected to manifest measurable responses to global warming. Globally, nitrogen (N) and phosphorus (P) are the most common nutrients limiting plant growth and soil carbon storage. We collected soil samples at 17 marsh sites in August 2012 across Northeast China. These samples were analysed for variations in soil organic carbon (OC), total nitrogen (TN) and total phosphorus (TP) levels, and multiple controlling of environmental and biotic factors. Results showed the means to be as follows: 16,850.7 mmol kg−1 (OC), 540.5 mmol kg−1 (TN), 30.0 mmol kg−1 (TP), 29.9 (C:N), 516.5 (C:P) and 16.8 (N:P). The OC, TN, TP and C:N:P ratios decreased with increases in the mean annual temperature (MAT) and flooding depth, whereas the C:N ratio did not change significantly with the flooding depth. Quadratic relationships were observed between the OC, TN and TP and soil pH. Linear mixed-effect models showed that climate exerted great influences on soil nutrients. These results will improve our understanding of the ecological patterns of nutrient fluxes and the biogeochemical mechanisms of the response of vegetation to climate changes.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
From cellular metabolism to ecosystem structure, carbon (C), nitrogen (N) and phosphorus (P) are biologically coupled through their influences on the biochemical reactions that control photosynthesis, respiration and decomposition in terrestrial ecosystems (Sterner and Elser 2002; Vitousek et al. 2010; Finzi et al. 2011; Melillo et al. 2011; Schlesinger et al. 2011; Peñuelas et al. 2012; Rivas-Ubach et al. 2012; Penuelas et al. 2013). Soil organic carbon (OC) is sensitive to climatic change, and it can be expected to manifest measurable responses to global warming (Xiao 1999). Globally, nitrogen (N) and phosphorus (P) are the most common nutrients limiting plant growth and soil carbon storage (Vitousek and Howarth 1991; Aerts and Chapin 2000). These elements are required in strict proportions for living organisms, in order to synthesize essential compounds with specific ratios of C:N:P (Sterner and Elser 2002; Finzi et al. 2011). Hence, a better knowledge of the soil C, N, and P ecological stoichiometries would enable predictions of how the terrestrial ecosystems would respond to future climate change.
Soil nutrient traits have received considerable attention in the past decade. However, in terrestrial systems, as soil has high spatial heterogeneity caused by both local-scale (e.g., fires, landslides and land use change) and regional-scale factors (e.g., glacial history, climate, topography, and biotic diversity) (Jenny 1941; McGroddy et al. 2004), there are large differences in research outcomes (Cleveland and Liptzin 2007; Tian et al. 2010; Bui and Henderson 2013; Qu et al. 2014). Therefore, studies at different scales are needed to understand the spatial heterogeneity of the soil.
Wetlands can be sources or sinks of C, N and P (Reddy et al. 1995; Mitsch and Gosselink 2007; Ramsar 2013) and are very sensitive to environmental change. Some environmental factors have been confirmed or assumed to be critical drivers, such as climate change (Yang et al. 2005; Novak et al. 2008), flooding regimes (Steinman et al. 2012; Wang et al. 2015), soil pH (Craft et al. 1988; Guo et al. 2008), and community type (Shang et al. 2013). However, most of these studies deal with the relationship between soil elements and environmental variables, especially at a small region. Little effort has been invested in seeking multivariable controlling of climate, flooding regime, soil pH and community type on soil chemical traits due to the high spatial heterogeneity in soil properties.
Northeast China has a large area of wetlands of around 1.02 × 105 km2, which accounts for 26.5% of the natural wetland area in China (Liu 2005). The wetlands are mostly located adjacent to the rivers and noteworthy for their rich biodiversity, however, they are suffering from a decline in area and deterioration in quality due to changes in climate and land use over recent decades. Although concern about global climate change is increasing, few studies of wetlands in Northeast China have paid attention to the impacts of these climatic factors on soil nutrient traits and C:N:P ratios (Zhang et al. 2012, 2013; Xiao et al. 2014).
Therefore the objective of this study was to explore the responses of OC, TN, TP and C:N:P ratios in Northeast China to the environmental factors in riverine wetlands. Specifically, the purposes of this study were to: 1) characterize the patterns of OC, TN, TP and C:N:P ratios across Northeast China, 2) evaluate variations in the patterns of soil element content along environmental gradient and 3) reveal how the environmental and biotic factors are associated with these patterns.
Materials and Methods
Study Area
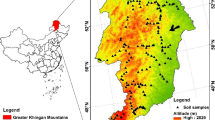
The study area, located in Northeast China, is a broad plain bordered on three sides by mountains with the Bohai Sea to the south (Fig. 1). Mean annual precipitation (MAP) varies from 59 mm in the west to 1090 mm in the east, and mean annual temperature (MAT) ranges from −4 °C to more than 11 °C. Winter is typically long and cold with predominantly northwest winds, whereas the summer is short, warm and wet with predominantly southeast winds (Lu 2008). Many rivers and streams run across the plain, including the Heilong River, Songhua River, Liao River, Ussuri River, Razdolnaya River, and Tumen River. The dominant soil types of the study region are azonal soils such as meadow soil, boggy soil and albic bleached soil. The typical marsh plants are species of the genera Carex, Glyceria, Deyeuxia, Phragmites, Cyperus and Echinochloa.

Distribution map of plots used in this study
Soil Sampling and Measurements
A total of 127 soil samples were collected across 17 marsh sites (Table 1) in August 2012. We selected large wetlands with minimal anthropogenic disturbances. At each wetland, at least three samples were collected using a metal auger to a depth of 10 cm. The samples were stored in polyethylene plastic bags. The soil was air-dried and then finely ground to pass through 1 mm nylon screens prior to chemical analysis.
OC concentrations were measured by the K2Cr2O7-H2SO4 oxidation method. TN and TP concentrations were measured by the Kjeldahl and molybdenum-antimony anti-spectrophotometric methods (Sparks et al. 1996), with a continuous flow analyzer (SKALAR San++, Netherlands) after pretreatment. Soil pH was determined by a Leici pH meter (PHS-3C, Shanghai).
Climate Data Collection
MAT and MAP estimates were derived using spatial interpolation with a resolution of 10 × 10 km in the ESRI ArcGIS 9.3 software package. Data for the estimates were obtained from 95 climate stations in Northeast China (from 1983 to 2012), which was sourced from China meteorological data sharing service system (cdc.cma.gov.cn/home.do).
Statistical Analyses
A square-root-transformed (sqrt) was first applied to the contents of OC, TN and TP and C:N:P ratios in order to improve the normality of the data for later statistical analyses. All means and standard errors were then transformed back into the original units. An analysis of the Spearman linear correlation coefficients was provided to reflect the relationship between soil stoichiometry and environmental factors. Analysis-of-variance followed by Student–Newman–Keuls (S–N–K) post hoc tests were used to examine potential differences in the means of sqrt(OC), sqrt(TN), sqrt(TP), sqrt(C:N), sqrt(N:P), sqrt(C:P) and sqrt(C:N:P) ratios for different regions and community types.
Linear mixed-effects models (LMMs) were used to calculated the variance explained by fixed effects only (marginal R2) and the variance explained by both fixed and random effects (conditional R2; Nakagawa and Schielzeth 2013). Climate, soil pH, flooding depth and community type were treated as fixed factors and site was treated as random factor. Akaike information criterion (AIC) was used to select competing models: model with the lowest AIC valuewas chosen as the final model. All the above-mentioned analyses were carried out using SPSS 18.0, 2010 and R 3.3.1 (R Development Core Team 2016).
Results
Statistics of Soil OC, TN, TP
Across all data, the OC contents varied from 111.9 mmol kg−1 to 51,390.5 mmol kg−1, and the highest coefficient of variation was approximately 84%. The TN values ranged from 11.6 mmol kg−1 to 1809.1 mmol kg−1, and the TP values varied from 2.5 mmol kg−1 to 115.7 mmol kg−1. The soil C:N ratio varied between 9.3 and 72.4, and the soil N:P ratio ranged from a low of 2.7 to a high of 52.0. The observed soil C:P ratio exhibited the widest range (from 26.5 to 2102.1) and the highest coefficient of variation of 67% (Table 2).
The Spearman linear correlation indicated that the results were significant, positive associations among the OC, TN and TP concentrations (overall, P < 0.01 for all models; Fig. 2). The strength of the relationship varied between elements, although the R2-values ranged from 0.688 (soil C:P) to 0.899 (soil C:N; Fig. 2).

Relationships among the SOC, TN and TP contents in wetland soils of Northeast China
Variations of Soil OC, TN and TP along Environmental Gradients
Negative linear correlations were found between the OC, TN, TP and C:N:P ratios and environmental variables including MAT and flooding depth, where only the C:N ratio did not significantly differ with flooding depth (Figs. 3a–f and 4a–f). Soil pH showed negative linear correlations with C:N, C:P and N:P, while quadratic regression relationships with OC, TN and TP (Fig. 4g–l). As to MAP, soil elements exhibited no significant correlations with it (Fig. 3g–l).

Relationships between soil stoichiometry and MAT and MAP in wetland soils of Northeast China

Relationships between soil stoichiometry and flooding depth and soil pH in wetland soils of Northeast China
Variation among Community Types
Large differences in the soil nutrient concentrations were found among community types(p < 0.05). The OC, TN and TP of Carex, Glyceria, and Deyeuxia were markedly higher than that of the others. The C:N of Echinochloa and C:P of Cyperus were significantly lower than that of Carex and Glyceria. The N:P of Cyperus were significantly lower than that of the other communities (Table 5).
Multivariate Relationships of Soil OC, TN and TP versus Environmental and Biotic Factors
The LMMs analysis showed that for soil OC, TN and TP, the best model included MAT, MAP and site as predictors, with a marginal R2 of 0.31, 0.28 and 0.45 and conditional R2 of 0.45, 0.45 and 0.45, respectively (Table 3). For C:N and C:P, MAT and site were included in the final model, with a marginal R2 of 0.13 and 0.13 and conditional R2 of 0.24 and 0.33, respectively (Table 3). The best model included site for N:P (Table 3), with a conditional R2 of 0.27.
Discussion
The spatial distributions of soil C, N and P are important for understanding the underlying patterns of nutrient fluxes along with the biogeochemical mechanisms of the response of vegetation to climate change. Quantifying the response of environmental and biotic factors to soil nutrient traits is important for ecologists to understand the driving mechanism of these patterns. This study is the first description of wetland soil C, N and P in Northeast China, and will provide the scientific base for management of this region.
Variation of Soil C, N and P
Our analysis showed that the contents of OC, TN, and TP in the wetland soils were higher than values reported from those in grasslands and forests (Table 4). Similar results have also been demonstrated by the other studies (Jiao et al. 2013; Shang et al. 2013; Gao et al. 2014; Fan et al. 2015). This may be related to the anaerobic condition in wetlands, which inhibits litter decomposition. To a certain degree, this explains the fact that wetland ecosystems, which occupy a relatively small land surface area, play an important role in the global storage capacity of C, N and P.
Our results also demonstrated that OC and P contents in wetland soils of Northeast China were higher and lower, respectively, when compared to those in other regions of China (Table 4). Meanwhile, the two elements in grasslands and forests of Northeast China were also higher and lower than those in other regions of China (Wang 2007; Wang et al. 2014; Zhao et al. 2014; Fan et al. 2015). This may be due to the unique climate and geologic parent material in our research area. Consequently, this finding indicates that we should pay more attention to the research on the interaction between soil OC, P content and plants in this region in the near future.
Cleveland and Liptzin (2007) observed that in the soil, similar to marine ecosystems, the atomic C:N:P ratios are constrained and suggested that close interactions between organisms and the environment drive the observed similarities in their element ratios. Although the concentrations of soil elements were highly variable (Fig. 2), our analysis suggests a similar pattern in the wetland soils of the study region. The C:N, C:P and N:P ratios were well-constrained throughout the dataset, as demonstrated by the relatively high correlation coefficients among the OC, TN and TP concentrations, where R2 = 0.899, 0.601, 0.688, respectively (Fig. 2). This might indicate that the soil C:N:P ratio was remarkably well-constrained, at a value of approximately 517:17:1. However, since this value is different to the 186:13:1 value reported in Cleveland and Liptzin (2007), this demonstrates that the C:N:P ratio might change with variations in environmental factors.
Influence of Environment Factors on C, N and P
We observed a negative association between spatial distributions of soil nutrients and MAT (Fig. 3a–f). This pattern agreed with the observations found by previous studies in China (Zhao et al. 2005; Yang et al. 2010). Temperature influences the accumulation and mineralization of organic matter by controlling the activity of soil microbes (Craft et al. 1988; Ajwa et al. 1998). Paul (2014) reported that a C:N > 25 on a mass basis implies that organic matter accumulation is occurring faster than decomposition and a C:P < 200 implies net mineralization. In our research, the C:N ratio was greater than 25 in Sanjiang Plain and Daxinganling where the MAT was relatively low, indicating net immobilization; in contrast, the C:P ratio was less than 200 in Liaohe Estuary and Songnen Plain where the MAT was relatively high, thus indicating net mineralization (Fig. 1, Table 4). Hence, this demonstrates that the OC, TN and TP contents decreased with increasing temperature.
Flooding depth is the most significant factor that controls production and turnover of organic matter in wetlands (Brinson et al. 1984; Sun and Liu 2007). Our results showed that the flooding depth clearly decreased the SOC, TN and TP concentrations. These decreasing concentrations were consistent with those observed in previous studies (Guo et al. 2008; Zhao et al. 2014). Low microbial activity due to the low oxygen levels in persistently flooded systems inhibited decomposition, which tended to produce low-nutrient soil conditions (Crawford et al. 2007; Guo et al. 2008). Furthermore, high water levels make litter not easily returned to the soil directly, which reduces the input of OC, N and P (van Oorschot et al. 2000; Bai et al. 2005; Zhao et al. 2014) that cause strongly anaerobic conditions in surficial soils, which could induce the release of phosphorus (Patrick and Khalid 1974). When soluble phosphorus is released into a flooded wetland it may flow out of the wetland.
Soil pH is an important chemical factor that directly affects the existing forms, bioavailability and migration of various elements (Yu et al. 2002). Our results showed that the OC, TN and TP had quadratic relationships with the soil pH. Similar trends were found in other wetlands (Bai et al. 2003; Gao et al. 2008). Perhaps it could be attributed to bacterial diversity, richness and activity which were highly inhibited in acid or alkaline environments (Huang 1994; Li et al. 2001; Fierer and Jackson 2006). Hence, when microorganisms decompose less litter, lower levers of OC, TN and TP exist in the soils.
Plant production inputs are the major source of carbon and nitrogen in wetland soils (Gorham 1991; Updegraff et al. 1995; Bridgham et al. 1998). We found significant changes in OC, TN and TP among community types (Table 5), which can partly be explained by higher relative belowground biomass for Carex, Glyceria, and Deyeuxia than for Phragmites, Cyperus and Echinochloa (He 2003; Jia et al. 2006; Qin et al. 2006; Zhang et al. 2006). Below-ground plant residues can be decomposed easier, however wetlands are covered by water, so litter is not easily returned to the soil.
Multivariate Relationships of Soil C, N and P versus Flooding Depth, Vegetation Type and Climate
The LMMs analysis showed that climate was the major environmental factor influencing soil nutrients, which was in accordance with previous studies (Jobbágy and Jackson 2000; Xu et al. 2013). Climate explained large proportions of variance in soil OC, TN and TP (31, 28 and 45%, respectively, Table 3), indicating that soil elements may decrease in response to future increases in temperature. In addition, climate had stronger influences on soil TP than on soil OC and TN, which can be explained by the stronger response of P cycling induced by climate (Elser et al. 2010). In contrast to the adequate C and N mainly deriving from plants, soil P is mainly derived from primary rock minerals which are strongly influenced by shifts in climate (Chadwick et al. 1999; Aerts and Chapin 2000). Therefore, soil P is more sensitive to climate.
In the final LMMs, site explained large proportions of variation in OC, TN and C:N:P ratios (Table 3). The variation represented the among-site variability that had not been captured by the selected environmental factors. Unexplained variances may be due to various other factors we did not measure, such as micro-environment, geologic parent material and topography (Jenny 1941; Xu et al. 2013).
Conclusions
Our findings showed that soil C, N, P and C:N:P ratios were highly sensitive to environmental change, especially climate. This should encourage increased efforts to utilize the potential capacity of wetland ecosystems to fix carbon and decrease the emission of greenhouse gases. Our results suggest that there may be a single Redfield-like ratio in the wetland soils of Northeast China of approximately 566:18:1. However, further research is needed with even larger sample sizes to cover all the wetlands in the region.
References
Aerts R, Chapin F III (2000) The mineral nutrition of wild plants revisited. Advances in Ecological Research 30:1–67
Ajwa H, Rice C, Sotomayor D (1998) Carbon and nitrogen mineralization in tallgrass prairie and agricultural soil profiles. Soil Science Society of America Journal 62(4):942–951
Bai J, Deng W, Zhu Y, Luan Z, Zhang Y (2003) Spatial distribution characteristics and ecological effects of carbon and nitrogen of soil in Huolin River catchment wetland. The Journal of Applied Ecology 14(9):1494–1498 (in Chinese)
Bai J, Ouyang H, Deng W, Zhu Y, Zhang X, Wang Q (2005) Spatial distribution characteristics of organic matter and total nitrogen of marsh soils in river marginal wetlands. Geoderma 124(1):181–192
Bridgham SD, Updegraff K, Pastor J (1998) Carbon, nitrogen, and phosphorus mineralization in northern wetlands. Ecology 79(5):1545–1561
Brinson MM, Bradshaw HD, Kane ES (1984) Nutrient assimilative capacity of an alluvial floodplain swamp. Journal of Applied Ecology:1041–1057
Bui EN, Henderson BL (2013) C: N: P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant and Soil 373(1–2):553–568
Chadwick OA, Derry L, Vitousek PM, Huebert BJ, Hedin LO (1999) Changing sources of nutrients during four million years of ecosystem development. Nature 397(6719):491–497
Cleveland CC, Liptzin D (2007) C: N: P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 85(3):235–252
Craft C, Broome S, Seneca E (1988) Nitrogen, phosphorus and organic carbon pools in natural and transplanted marsh soils. Estuaries 11(4):272–280
Crawford ER, Day FP, Atkinson RB (2007) Influence of environment and substrate quality on root decomposition in naturally regenerating and restored Atlantic white cedar wetlands. Wetlands 27(1):1–11
Elser JJ, Fagan WF, Kerkhoff AJ, Swenson NG, Enquist BJ (2010) Biological stoichiometry of plant production: metabolism, scaling and ecological response to global change. The New Phytologist 186(3):593–608
Fan H, Wu J, Liu W, Yuan Y, Hu L, Cai Q (2015) Linkages of plant and soil C: N: P stoichiometry and their relationships to forest growth in subtropical plantations. Plant and Soil 392(1–2):127–138
Fierer N, Jackson RB (2006) The diversity and biogeography of soil bacterial communities. Proceedings of the National Academy of Sciences of the United States of America 103(3):626–631
Finzi AC, Austin AT, Cleland EE, Frey SD, Houlton BZ, Wallenstein MD (2011) Responses and feedbacks of coupled biogeochemical cycles to climate change: examples from terrestrial ecosystems. Frontiers in Ecology and the Environment 9(1):61–67
Gao J, Xu X, Zhang F, Wang C (2008) Distribution characteristics of soil labile carbon along water table gradient of alpine wetland soils. Journal of Soil and Water Conservation 22(3):126–131 (in Chinese)
Gao Y, Chen H, Zeng X (2014) Effects of nitrogen and sulfur deposition on CH4 and N2O fluxes in high-altitude peatland soil under different water tables in the Tibetan plateau. Soil Science & Plant Nutrition 60(3):404–410
Gorham E (1991) Northern peatlands: role in the carbon cycle and probable responses to climatic warming. Ecological Applications 1(2):182–195
Guo X, Lu X, Tong S, Dai G (2008) Influence of environment and substrate quality on the decomposition of wetland plant root in the Sanjiang plain, Northeast China. Journal of Environmental Sciences 20(12):1445–1452
He C (2003) Dynamics of litter and under-ground biomass in Carex lasiocarpa wetland on Sanjiang plain. The Journal of Applied Ecology 14(3):363–366 (in Chinese)
Huang R (1994) Environment Pedology. Advanced Education Press, Beijing, pp. 145–146
Jenny H (1941) Factors of soil formation. McGraw-Hill Book Co. Inc, New York, NY
Jia Q, Zhou L, Xie Y, Zhou G (2006) Study on biomass dynamics of Phragmites communis community in Panjin wetland. Journal of Meteorology and Environment 22:25–29 (in Chinese)
Jiao F, Wen Z-M, An S-S, Yuan Z (2013) Successional changes in soil stoichiometry after land abandonment in Loess Plateau, China. Ecological Engineering 58:249–254
Jobbágy EG, Jackson RB (2000) The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecological Applications 10(2):423–436
Li Z, Sun B, Lin X (2001) Density of soil organic carbon and the factors controlling its turnover in East China. Geographical. Science 21(4):301–301 (in Chinese)
Liu X (2005) Wetlands in Northern China. Science Press, Beijing, p. 9 (in Chinese)
Lu X (2008) Wetland ecosystem study in China. Hebei Sience and Technology Publishing House, Hebei, p. 338
McGroddy ME, Daufresne T, Hedin LO (2004) Scaling of C:N:P stoichiometry in forests worldwide: implications of terrestrial redfield-type ratios. Ecology 85(9):2390–2401. doi:10.1890/03-0351
Melillo JM, Butler S, Johnson J, Mohan J, Steudler P, Lux H, Burrows E, Bowles F, Smith R, Scott L (2011) Soil warming, carbon–nitrogen interactions, and forest carbon budgets. Proceedings of the National Academy of Sciences 108(23):9508–9512
Mitsch WJ, Gosselink JG (2007) Wetlands, 4th edn. Wiley, Hoboken
Nakagawa S, Schielzeth H (2013) A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods in Ecology and Evolution 4(2):133–142
Novak M, Brizova E, Adamova M, Erbanova L, Bottrell SH (2008) Accumulation of organic carbon over the past 150 years in five freshwater peatlands in western and Central Europe. Science of the Total Environment 390(2):425–436
Patrick W, Khalid R (1974) Phosphate release and sorption by soils and sediments: effect of aerobic and anaerobic conditions. Science 186(4158):53–55
Paul EA (2014) Soil microbiology, ecology and biochemistry. Academic Press, New York
Peñuelas J, Sardans J, Rivas-ubach A, Janssens IA (2012) The human-induced imbalance between C, N and P in Earth’s life system. Global Change Biology 18(1):3–6
Penuelas J, Poulter B, Sardans J, Ciais P, van der Velde M, Bopp L, Boucher O, Godderis Y, Hinsinger P, Llusia J (2013) Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe. Nature Communications 4:2934
Qin S, Liu J, Sun Z (2006) Dynamics of phosphorus and biomass accumulation of Calamagrostis angustifolia in Sanjiang plain. Journal of Applied Ecology 25(6):646–651 (in Chinese)
Qu F, Yu J, Du S, Li Y, Lv X, Ning K, Wu H, Meng L (2014) Influences of anthropogenic cultivation on C, N and P stoichiometry of reed-dominated coastal wetlands in the Yellow River Delta. Geoderma 235:227–232
R Development Core Team (2016) R: a language and environment for statistical computing, R version 3.3.1. R Foundation for Statistical Computing, Vienna. http://www.R-project.org/
Ramsar (2013) The Ramsar manual, 6th-ed edn. Ramsar Convention Secretariat, Switzerland, p. 110
Reddy K, Diaz O, Scinto L, Agami M (1995) Phosphorus dynamics in selected wetlands and streams of the Lake Okeechobee Basin. Ecological Engineering 5(2):183–207
Rivas-Ubach A, Sardans J, Pérez-Trujillo M, Estiarte M, Peñuelas J (2012) Strong relationship between elemental stoichiometry and metabolome in plants. Proceedings of the National Academy of Sciences 109(11):4181–4186
Schlesinger WH, Cole JJ, Finzi AC, Holland EA (2011) Introduction to coupled biogeochemical cycles. Frontiers in Ecology and the Environment 9(1):5–8
Shang Z, Feng Q, Wu G, Ren G, Long R (2013) Grasslandification has significant impacts on soil carbon, nitrogen and phosphorus of alpine wetlands on the Tibetan Plateau. Ecological Engineering 58:170–179
Sparks DL, Page A, Helmke P, Loeppert R, Soltanpour P, Tabatabai M, Johnston C, Sumner M (1996) Methods of soil analysis. Part 3-Chemical methods. Soil Science Society of America Inc.
Steinman AD, Ogdahl ME, Weinert M, Thompson K, Cooper MJ, Uzarski DG (2012) Water level fluctuation and sediment–water nutrient exchange in Great Lakes coastal wetlands. Journal of Great Lakes Research 38(4):766–775
Sterner RW, Elser JJ (2002) Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton University Press, Princeton
Sun Z, Liu J (2007) Nitrogen cycling of atmosphere-plant-soil system in the typical Calamagrostis angustifolia wetland in the Sanjiang plain, Northeast China. Journal of Environmental Sciences 19(8):986–995
Tian H, Chen G, Zhang C, Melillo JM, Hall CA (2010) Pattern and variation of C: N: P ratios in China’s soils: a synthesis of observational data. Biogeochemistry 98(1–3):139–151
Updegraff K, Pastor J, Bridgham SD, Johnston CA (1995) Environmental and substrate controls over carbon and nitrogen mineralization in northern wetlands. Ecological Applications 5(1):151–163
van Oorschot M, van Gaalen N, Maltby E, Mockler N, Spink A, Verhoeven JT (2000) Experimental manipulation of water levels in two French riverine grassland soils. Acta Oecologica 21(1):49–62
Vitousek PM, Howarth RW (1991) Nitrogen limitation on land and in the sea: how can it occur? Biogeochemistry 13(2):87–115
Vitousek PM, Porder S, Houlton BZ, Chadwick OA (2010) Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions. Ecological Applications 20(1):5–15
Wang W (2007) Spatical parterns of soil nutrients and their response to grazing disturbance over the Songnen meadow steppes of NE China. Northeast Normal University, pp. 59 (in Chinese)
Wang F, Song M, Huang M, Zhang J (2014) The spatial distribution of soil nutrients and the controlling factors of temperate forest and steppe in Northeastern China. Ecology and. Environmental Sciences 23(8):1280–1285 (in Chinese)
Wang W, Wang C, Sardans J, Tong C, Jia R, Zeng C, Peñuelas J (2015) Flood regime affects soil stoichiometry and the distribution of the invasive plants in subtropical estuarine wetlands in China. Catena 128:144–154
Xiao H (1999) Climate change in relation to soil organic matter. Soil and Environmental Sciences 4:014
Xiao Y, Shang L, Huang Z, Zhang W, Xue Z, Zhang Z, Lu X (2014) Ecological stoichiometry characteristics of soil carbon, nitrogen and phosphorous in mountain swamps of eastern Jilin province. Scientia Geographica Sinica 34(8):994–1001 (in Chinese)
Xu X, Thornton PE, Post WM (2013) A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Global Ecology and Biogeography 22(6):737–749
Yang G, Zhang W, Tong C, Wu J (2005) Effects of temperature on the mineralization of organic carbon in sediment of wetland. Acta Ecologica Sinica 25(2):343–348 (in Chinese)
Yang Y, Fang J, Guo D, Ji C, Ma W (2010) Vertical patterns of soil carbon, nitrogen and carbon: nitrogen stoichiometry in Tibetan grasslands. Biogeosciences Discussions 7:1–24
Yu J, Wang J, Liu J, Qi X, Wang Y (2002) Effect of soil pH value variation on effective content of trace elemments in typical black soil. Journal of Soil and Water Conservation 16(2):93–95 (in Chinese)
Zhang Y, Yang Y, Wang L (2006) Studies on seasonal dynamics of production and allocation in Phragmites australis population in Sanjiang wetland plain. Chinese Journal of Grassland 28(4):1–5 (in Chinese)
Zhang Z, Lu X, Song X, Guo Y, Xue Z (2012) Soil C, N and P stoichiometry of Deyeuxia angustifolia and Carex lasiocarpa wetlands in Sanjiang plain, Northeast China. Journal of Soils and Sediments 12(9):1309–1315
Zhang ZS, Song XL, Lu XG, Xue ZS (2013) Ecological stoichiometry of carbon, nitrogen, and phosphorus in estuarine wetland soils: influences of vegetation coverage, plant communities, geomorphology, and seawalls. Journal of Soils and Sediments 13(6):1043–1051
Zhao K (1999) Chinese marsh. Science Press, Beijing (in Chinese)
Zhao S, Yu W, Zhang L, Shen S (2005) Latitudinal differentiation of phosphorus in black soil in Northeast of China. Chinese society of. Agricultural Engineering 21:34–37 (in Chinese)
Zhao H, Liu W, Wang X, Cai Y, Du Z (2014) Distribution of soil carbon and nitrogen under different water conditionsin alpine salty wetlands, northern Tibet Plateau. Mountain. Research 32(4):431–437 (in Chinese)
Acknowledgements
This research was supported by the National Natural Science Foundation of China (Nos. 41371107, 41271106) and the National Key Technology R & D Program (2012BAC19B05). We thank Dr. Andrew Revill for language edit and Dr. Zhi Ding for a drawing of a map.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
About this article

Cite this article
Liu, Y., Jiang, M., Lu, X. et al. Carbon, Nitrogen and Phosphorus Contents of Wetland Soils in Relation to Environment Factors in Northeast China. Wetlands 37, 153–161 (2017). https://doi.org/10.1007/s13157-016-0856-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-016-0856-2