
Abstract
The preservation of endangered species requires clarifying habitat preferences through survival, growth and competitive ability. The determinants of habitat differentiation between the endangered species, Drosera anglica, and a widespread congener, D. rotundifolia, were compared. The effects of water level, Sphagnum mats and overstory vascular plants on Drosera distribution, recruitment and survival were monitored at a previously mined Sphagnum peatland. Seedling transplant experiments were conducted using different water levels. Seed-sowing experiments were conducted using different light intensities in three habitat-types: bare ground, Sphagnum mat and waterlogged surface. Distributions of D. anglica and D. rotundifolia were determined using survival at the seedling stage. D. anglica seedling recruitment and survival occurred more at lower water levels and/or lower plant cover, while D. rotundifolia seedlings established independent of these factors. In the greenhouse the seedlings of both species survived better at lower water levels but grew more slowly. D. anglica seedlings reduced their growth under shade more than D. rotundifolia. D. anglica showed low competitive light and nutrient ability on Sphagnum mats. Therefore, D. anglica was pushed to areas of high water levels where few competitors could establish. The habitat differentiation between D. anglica and D. rotundifolia originated from the interactions with Sphagnum mats.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The conservation of endangered plant species is critical for maintaining biodiversity (IUCN 2013). Endangered species often establish with common congener (s), but endangered species develop smaller populations and/or narrower distributional ranges compared with more common species (Brook et al. 2008). Species comparisons allow us to detect the mechanisms and determinants of population dynamics, including extinction (Young et al. 2007; Verhulst et al. 2008; Zietsman et al. 2008). The main factors influencing endangered species are their relatively small habitats and distributions derived from high self-compatibility, small seed size, low dispersal ability, high vegetative reproduction and/or low competitive ability (Murray et al. 2002; Römermann et al. 2008).
In boreal regions, broadly distributed bogs show significant biodiversity loss from human activities, represented by changes in land use by drainage construction for agriculture, resulting in the extinction of rare species (Keddy 2010). Drosera anglica Hudson and D. rotundifolia L. are carnivorous plants with overlapping distributional ranges, occurring in Sphagnum bogs in the circum-boreal regions of the Northern Hemisphere (Crowder et al. 1990). D. anglica has speciated from hybridizing between D. rotundifolia and D. linearis (Rivadavia et al. 2003). D. rotundifolia is a relatively widespread species compared with D. anglica (Wolf et al. 2006). D. anglica populations are declining even within undisturbed bogs in various regions where there are no direct anthropogenic disturbances because of climate changes (Huntke 2007; Jennings and Rohr 2011). D. anglica has been assigned as a vulnerable species in the Red List, Japan (Ministry of the Environment Government of Japan 2013).
The establishment of D. anglica requires higher water-table levels compared with D. rotundifolia (Nordbakken 1996; Nordbakken et al. 2004). However, seed germination and seedling survival determine the habitat preference and distribution (Poorter 2007; ten Brink et al. 2013), with ecologically ‘safe’ sites showing differences between seed germination and seedling growth (Dalling et al. 2001; Eriksson 2002). In wetlands, seed germination rates are often determined by water levels (Keddy 2010), whereas seedling growth is inhibited by environmental stress, such as drought and inundation (Coops and van der Velde 1995; Anderson et al. 2009). Therefore, these two environmental factors were separately examined, clarifying the effects of water levels on D. anglica and D. rotundifolia.
The habitat preferences of wetland plants are determined by water levels and interspecific interactions, including light and nutrient acquisition (Keddy et al. 1998). Sphagnum mats, commonly found in association with D. anglica and D. rotundifolia, promote the establishment of vascular plants through increasing moisture in peat and reducing light-attenuating feather mosses in ombrotrophic bogs (Fenton and Bergeron 2006). However, the Sphagnum mats enhance nutrient competition with vascular plants (Heijmans et al. 2002; Malmer et al. 2003). These results suggest that Sphagnum mats may mediate habitat differentiation between D. anglica and D. rotundifolia.
Sphagnum mats develop heterogeneous habitats through increased microtopographical diversity such as the hollow-hummock complex (Gore 1983). The hummocks are covered with vascular plants while the hollows have less vegetation (Charman 2002). We hypothesized that Sphagnum mat directly differentiated the habitats between D. anglica and D. rotundifolia, where the resulting factors are indirectly related to habitat differentiations. To test our hypothesis, we monitored the seedling recruitment and survival of D. anglica and D. rotundifolia species for three years, experimentally testing the effects of water level, shade and habitat on the seedling survival and growth.
Materials and Methods
Study Site
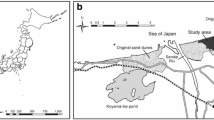
The field census was conducted at a previously mined peatland in the Sarobetsu mire (45°06’N, 141°42’E, 7 m elevation), northern Hokkaido, Japan. Peat mining was conducted from 1970 to 2003 and thereafter terminated. During 2008 and 2011, the mean annual temperature was 6.4 °C, with a mean annual precipitation of 962.4 mm at the Toyotomi weather station 6 km from the field site (Japan Meteorological 2012. The original peatland was dominated by Sphagnum mosses mainly S. papillosum Lindb. (Nishimura et al. 2009). The most common vascular plant species on the mined peatland were Scheuchzeria palustris L., Rhynchospora alba (L.) Vahl, Vaccinium oxycoccus L. and Carex middendorffii Fr. Schm. However, the plant species cover was relatively low.
Field Census and Seed Preparation
D. rotundifolia and D. anglica became established on the peatland where peat-mining was conducted ca. 39 years ago (Hoyo and Tsuyuzaki 2013). Five 300 × 40 cm plots divided into thirty 20 × 20 cm cells in areas where the two Drosera species were established. The elevation and water level were measured at the four corners of each cell on 20 November 2010, a few days after rainfall. Level lines were established using nylon string stretched along the two long sides for each plot. The distances between the lines and ground surface and between the lines and water surface were measured for each cell grid. The elevation from the ground surface was calculated using the difference between the water and ground surface. For this survey the water surface was defined as 0 cm elevation. The water elevation for each cell was calculated using the mean difference between the water and ground surface from each of the cell’s four corners.
Three categories of growth stages were assigned to each shoot. These growth stages consisted of the seedling, daughter, and non-seedling. Each growth stage was defined as: the seedling stage was an annual shoot; the daughter was a ramet produced by vegetative reproduction; the non-seedling was the over-wintered stage of the annual shoot. The location and length of the largest leaf were recorded from late July to early August for 3 years (2009–2011). The leaf length was measured with a ruler. The vegetation in each cell was recorded using Braun-Blanquet classes (Braun-Blanquet 1964).
The seeds used for the transplant experiments were collected from ≥ 30 individuals per plant species in the wetland during August and September of the previous year. The individuals that were visually matured were selected for sampling. The seeds were air-dried in paper bags for 5 days and stored at 3 °C until required. The seeds were cold-stratified in Petri dishes placed in an incubator at 5/1 °C (12 h/12 h, day/night cycle, respectively) for eight weeks to break seed dormancy (Baskin et al. 2001). The seeds were then allowed to germinate in an incubator at 25/15 °C (day/night cycle) for 5 weeks.
The effects of water level on seedling survival and growth
The effects of water level on seedling survival and growth were examined under greenhouse conditions. Twenty-four plastic pots (12 × 10 cm) were filled with commercially available peat moss and placed into four plastic containers (42 × 31 cm surface area; 16 cm depth). The water level was controlled using holes drilled in the sides of the containers at 1, 3, 5 or 9 cm, giving water levels of 9, 5, 3, and 1 cm. Water was supplied via sprinklers 10 times daily, with plants receiving natural sunlight and temperatures ranged from 20 to 32 °C. One hundred seedlings were grown in each pot during early August, 2009. Seedling survival was monitored weekly. The water content in the peat moss was measured using time domain reflectometry (TDR) (Hydrosense, Campbell Scientific, Logan, UT) with a 12-cm probe inserted vertically to the peat. All seedlings were grown for 63 days, oven dried at 70 °C for 48 h and weighed. Each treatment contained three replicates.
The Effects of Shade on Seedling Recruitment and Growth
The effects of shade on seedling recruitment and growth were investigated using a factorial-designed seed-sowing experiment in an area mined during 1972. The three shade levels were: a 30 % reduction using a single white sheer net; 50 % using a single black net; 70 % using a doubled black net (Dio-Kanreisya, Dio Chemicals, Ltd. Tokyo, Japan). For each Drosera species, one hundred seeds were individually sown in degradable pot (100 cm2 surface area; 7.5 cm depth) filled with Sphagnum peat collected from the study site. The seeds were buried in the field on 15 June 2011 and monitored monthly until 10 September, 2011. Six replicates were prepared per treatment per species. Two pots were not used for the analysis because of mammalian trampling. In 2012, the light intensity was measured on one pot per treatment at hourly intervals from June 15 to September 10, using an automatic data logger (HOBO, UA-002-XX, Onset Computer Corporation, Bourne, MA). The seedlings were excavated 87 days after transplanting, and oven dried at 70 °C for 48 h. The samples were weighed after cooling for 24 h in a desiccator.
The Effect of Habitat on Seedling Establishment
To establish habitat preferences, three different habitats were selected for the seed-sowing experiment: bare ground, Sphagnum mat, and waterlogged surface. Three replications of 100 × 30 cm plots were established in each habitat in late June, 2010. The distances separating the habitats were: 30 m between the bare ground and Sphagnum mat and 200 m between the bare ground and waterlogged surface habitats. Following the removal of all Drosera shoots prior to the experiment, a total of 1,000 seeds were sown per plot.
To compare seedling growth and survival between habitats, one hundred seeds were sown in 30 plastic cups for each plot (8 cm × 12 cm; 10 cups for each species plus the control, n = 30 cups). The cups were buried to a depth of 3 cm in each habitat on 3 July, 2010. Ten blank cups were used as the control in each habitat to estimate the number of seedlings that contaminated the plots. On 30 July (2010) most of the seeds germinated. Wire frames (8.5 × 8.5 cm) were established to mark the location on each shoot following cup removal. The seedlings were monitored monthly during July and September 2010 and mid-summer in 2011/2012. On each plant, the length of the longest leaf was measured in late July 2011/2012. The seedlings were removed on July 25 (2012) and the biomass was oven dried at 60 °C for 48 h. Thee samples were weighed after cooling for 24 h in a desiccator. Preliminary measurements confirmed that biomass did not differ significantly between oven dried at 60 °C and 70 °C over 48 h.
Volumetric water content was measured in July and October 2010 using a TDR (Hydrosense, Campbell Scientific). The light intensity was measured at the three ground levels in each plot at 1 h intervals from June 2010 to July 2012 using automated data loggers (HOBO, UA-002-XX, Onset Computer Corporation, Bourne, MA). Where the sensors were inundated or covered with litter material, the data was not used in the analysis. The light intensity was converted to photosynthetic photon flux density (PPFD) by comparing the light intensity and PPFD measured using a sensor established on a bare ground (HOBO, S-LIA-M003, Onset, USA).
Statistical Analysis
For seedling survival and growth analysis, seedlings where the maximum leaf length >3 mm were used, following seedling identification (Hoyo and Tsuyuzaki 2013). Hierarchical Bayesian models (HBMs) estimated the determinants on density, recruitment and survival for the two species. The parameters of HBMs were estimated with a Markov chain Monte Carlo (MCMC) simulation using WinBUGS 1.4.3 through R2WinBUGS (Spiegelhalter et al. 2003). Seedling density, recruitment and survival were estimated for each species per annum. The response variable, y i , is the probability of the number of individual or recruitment in cell i, following a Poisson distribution, y i ~ P (λ), where λ is the observed number in cell i. The log-link function was applied to connect λ and the linear predictor (logλ i ) as:
where β 1 is the intercept and β 2 and β 3 the coefficients; E i elevation; V i overstory vegetation cover; and S i Sphagnum cover. The random effects, r i and r j are for cell i and plot j, respectively. The prior distribution of β is non-informative, following a normal distribution, N (0, 104). The inverse-variance parameters r i and r j are assumed to follow a priori distributions, N (0, σ c ) and N (0, σ p ). Here, σ c and σ p represent the standard deviation of individual characteristics under the linear predictor of λ i , assuming they follow a gamma distribution, 1/σ ~ G (10−4, 10−4). The spatial autocorrelation of the distribution of individuals was not considered, because a preliminary analysis detected weak autocorrelations.
To investigate the effects of elevation, Sphagnum cover and overstory vegetation on seedling survival were examined for each species. The response variable, p i , is the number of seedlings observed in cell i, following a binomial distribution, p i ~ B (q i , N i ). The survival probability (q i ) represents survival from the previous year in cell i, and N i is the number of individuals in cell i from the previous year. The logit-link function connects p i and the linear predictors as:
where D i was the density in cell i, and β, E, V and r are shown in Eq. 1. The prior distribution of β is non-informative following N (0, 102). The posterior samples were obtained using the combination of three independent MCMC chains with 1,000 samplings at 20-step intervals after 20,000 burn-in MCMC steps. The ^R < 1.1 for all the parameters showed the Markov chains converged (Gelman et al. 2003).
The effects of water level, shade and habitat on seedling survival were analyzed using a generalized linear mixed-effects model (GLMM) with a binomial distribution. The cell code was used as the random effect. The seedling biomass was compared between species, between treatments (water level, shade or habitat) and between their interactions, using a generalized linear model (GLM) with a gamma distribution and log-link function. Akaike’s information criteria (AIC) were used to select the best models given the data. The statistical analysis was conducted using the statistical program R (v. 2.15.1, R Development Core Team 2012; R Foundation for Statistical Computing, Vienna, AT).
Results
Vegetation and Habitat
Drosera anglica cover ranged from 1 to 15 % in the five plots for the 3 years surveyed, with D. rotundifolia covering 2 to 7 % of the five plots. In the five plots, the dominant species was Rhynchospora alba, covering < 25 %, followed by Carex middendorffii and Moliniopsis japonica (Table 1). The Sphagnum cover varied between the plots ranging from 5 to 69 %, with areas of open water ranging from 10 to 67 %. Elevation showed no correlation with overstory vegetation cover (r 2 = 0.11, P = 0.18), but was positively correlated with Sphagnum cover (r 2 = 0.69, P <0.001).
The number of shoots per plot fluctuated annually between 30 and 554 for D. anglica and between 82 and 546 for D. rotundifolia (Table 1.). For D. rotundifolia, The percentage in the different growth stages for seedling, daughter and non-seedling was 43, 1 and 56 % respectively, with D. anglica showing 40, 9 and 51 % in the seedling, daughter and non-seedling (respectively) growth stages. Shoots in the daughter growth stage were nine times higher for D. anglica than for D. rotundifolia, but with lower densities.
Temporal Changes in Density, Recruitment and Survival
The density intercepts for HBMs were significant for D. rotundifolia in 2010 and 2011. The remaining intercepts were non-significant (Fig. 1). D. anglica plants had the highest densities at low elevations (2010 and 2011), while D. rotundifolia plants had the highest density at high elevations. Under high overstory cover, D. anglica had relatively low densities in 2009. The Sphagnum mat area reduced D. anglica, but not D. rotundifolia density. The density of D. anglica was determined by overstory plants, the Sphagnum mat and water level, while D. rotundifolia density was limited only by the water level.

Determinants for the densities of Drosera anglica and D. rotundifolia, estimated using a hierarchical Bayesian model. The coefficients are shown by medians (symbols) with 95 % Bayesian credible intervals (vertical bars) in 2009, 2010 and 2011. Asterisks show the variables do not overlay zero at 95 % Bayesian credible intervals
For 2010 and 2011, the D. rotundifolia seedling recruitment intercept was significantly different from zero, the other three intercepts were non-significant (Fig. 2). D. rotundifolia seedling recruitment was greater at high elevation in 2010, but D. anglica recruitment was low under high overstory cover (2010) and on Sphagnum mats in 2010 and 2011. D. anglica seedlings had high survival at high elevation, but low survival where the overstory cover was high. D. anglica seedling survival declined with increasing seedling density, but D. rotundifolia did not show this pattern. Therefore, the number of sites for seedling establishment was more limited for D. anglica compared with D. rotundifolia. Elevation, overstory plant cover, plant density and the Sphagnum mat showed no effect on non-seedling survival of both Drosera species, indicating that individual plant survival was mostly determined before overwintering, i.e., at the annual seedling stage.

Determinants of Drosera anglica and D. rotundifolia on the recruitment rates of seedlings and daughters, and on the survival rates of seedlings and non-seedlings in 2010 and 2011, estimated using a hierarchical Bayesian model. Asterisks show the variables do not overlay zero at 95 % Bayesian credible intervals
All the intercepts of the daughter production growth-stage showed a significant difference (Fig. 2). Overstory vascular plant cover reduced daughter production of D. anglica but did not alter the daughter production on D. rotundifolia. D. anglica produced more daughters at low elevations in 2010, while D. rotundifolia daughter production showed no association with water level. The Sphagnum mat did not affect daughter production of the two Drosera species. Overall, the water level and overstory plant cover influenced daughter production on D. anglica.
The Effect of Water Level on Seedling Survival and Growth
The water content in peat decreased slightly with decreasing water levels, i.e., 68, 63, 61 and 56 % at 9, 5, 3 and 1 cm depth, respectively. The seedling survival was lower at water levels of 3 and 1 cm compared with 9 cm 2 months after seed-sowing. Seedling survival was higher for D. anglica compared with D. rotundifolia (Fig. 3a). The interaction between treatments and species was not significant (Table 2). For D. anglica and D. rotundifolia seedling survival was lowest (40 %), at the lowest water level.

The effects of water level on seedling survival and biomass in the greenhouse experiment. (a) Temporal changes in the mean survival of Drosera anglica and D. rotundifolia shown with standard errors (vertical bars) at four different water levels. (b) Seedling biomass for D. anglica and D. rotundifolia
The seedling biomass was higher on D. anglica (0.472 ± 0.273 mg, mean ± standard deviation) compared with D. rotundifolia (0.272 ± 0.136 mg; Fig. 3b). The optimal water levels for seedling growth were between 1 and 5 cm when the water level was stable. The two species developed larger seedlings when the water levels were 1 and 5 cm (Fig. 3b). However, the interaction effect indicated that biomass increase of D. rotundifolia was slower at water levels of 3 and 5 cm compared with D. anglica (Table 2).
The Effects of Shade on Seedling Survival and Growth
The number of seedlings increased 3 weeks after the seeds were sown, stabilised or showed a slight decline (Fig. 4a). Shading did not affect the number of seedlings (Table 2). D. anglica produced larger seedlings (0. 365 ± 0.527 mg) compared with D. rotundifolia (0. 220 ± 0.174 mg; Fig. 4b). Shading reduced D. anglica seedling growth but not for D. rotundifolia (Table 2). Therefore, the shading of D. anglica resulted in a higher number of seedling recruitment, survival and growth but reduced plant biomass, while shading had no influence on D. rotundifolia seedling recruitment, mortality and growth.

Seedling survival and biomass of Drosera anglica and D. rotundifolia in the seed-sowing experiments at Sarobetsu mire in relation to shade cover of 30, 50 and 70 %. (a) Changes in numbers of seedlings (shown with standard errors by vertical bars). (b) Seedling biomass
Seedling Survival and Growth in Three Different Habitats
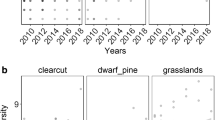
The mean water content for the bare ground, Sphagnum mat and waterlogged surface was 93, 105 and 101 %, respectively. Before plant growth began in June, the photosynthetic photon flux density (PPFD) was high with no significant differences among the three habitats (Fig. 5). After June the PPFD decreased in all three habitats, in particular on Sphagnum mat, as the PPFD was reduced by the growth of overstory plant and the Sphagnum mat. The PPFD on the Sphagnum mat was equivalent to a 50 % of full shade from the sheer net material.

Temporal changes in the daily maximum photosynthetic photon flux density (PPFD) in 2011 on the three habitats: bare ground (solid lines), Sphagnum mat (dotted lines) and waterlogged surface (interrupted lines). Artificial shade that reduced 50 % of PPFD is also shown (grey lines)
Most seedlings originated from the seeds sown in the first year of the field experiment, because ten, three and eight seedlings (bare ground, Sphagnum mat and waterlogged surface, respectively) emerged in the blank controls. The seedlings of the two Drosera species showed higher recruitment on the Sphagnum mat compared with bare ground and the waterlogged surface 1 month after seed sowing (Fig. 6a). However, until late September 2010, D. anglica and D. rotundifolia seedlings usually died with the highest mortality on the Sphagnum mat. In 2011, most unidentified Drosera seedlings occurred on the Sphagnum mat (Fig.6b). The number of survivors did not differ between the two Drosera species in the third year (2012), except for a large number of D. rotundifolia surviving on the Sphagnum mat (Table 2). More D. anglica seedlings survived on bare ground, where three plants flowered during 2012.

Fluctuations in the number of seedlings for Drosera anglica and D. rotundifolia in three habitats: bare ground, Sphagnum mat and waterlogged surface. (a) The number of seedlings per cell from July to September, 2010. (b) The number of seedlings per plot from September 2010 to July 2012. The squares in (b) indicate the number of unidentified seedlings where leaves were < 3 mm long. The number is shown using the mean ± standard errors. (c) Seedling biomass of D. anglica and D. rotundifolia
When the seedlings were harvested in July 2012, D. anglica had larger seedlings and higher biomass compared with D. rotundifolia. The seedlings were largest in the bare ground habitat for both species (D. anglica, 15.35 mg ± 17.95; D. rotundifolia, 9.17 ± 9.62 mg; Fig. 6c). The biomass of D. anglica and D. rotundifolia was 1.81 ± 1.16 mg and 0.39 ± 0.10 mg, respectively, on the waterlogged surface and 0.18 ± 0.23 mg and 0.22 mg ±0.31 (D. anglica and D. rotundifolia, respectively) on the Sphagnum mat. The waterlogged surface reduced the seed germination and growth of the two species, but allowed higher establishment levels of D. anglica compared with D. rotundifolia
Discussion
Water Level and Habitat Differentiation
This study indicated that water level, overstory plants and the Sphagnum mat restricted D. anglica densities more than those of D. rotundifolia. However, the overwintering shoot survival of these two species was unaffected by the water level, overstory plant cover and the Sphagnum mat. At the Sarobetsu mire site, the results indicated that the distribution patterns of D. anglica and D. rotundifolia were determined mainly at the annual seedling stage, including the dominant species, R. alba and M. japonica (Egawa et al. 2009; Koyama and Tsuyuzaki 2010).
The water level was significantly associated with annual seedling survival and growth of the two Drosera species. The greenhouse experiments suggested that the optimal water level range was relatively narrow (i.e., a few centimetres), for seedling growth. In the seeding experiment in the three habitats, the seedling biomass was highest on bare ground where the two Drosera species did not normally establish. D. anglica and D. rotundifolia grew faster on bare ground, as these two species are usually found in full-sun habitats (Crowder et al. 1990). However, inundation and drought often occur on bare-ground habitats (Koyama and Tsuyuzaki 2013). Therefore, seedling establishment is reduced on bare ground because of the fluctuations in water levels over the short and long term.
Habitat Differentiation Between D. Anglica and D. Rotundifolia
The shade effect from overstory vegetation decreased the number of D. anglica shoots but not the number of D. rotundifolia shoots. The experimental shading effect did not decrease seedling survival for both species but reduced D. anglica biomass. This species was more shade-intolerant compared with D. rotundifolia, although these two species are categorized as shade-intolerant species. Sites for seedling recruitment are not necessarily preferable sites for seedling growth and survival, because of site exclusion from biotic and/or abiotic factors (Poorter 2007). The waterlogged surface had a relatively low level of vascular plant cover and Sphagnum mosses. In addition, D. anglica grew faster on the waterlogged surface than on the Sphagnum mat. This suggests that reduced interspecific competition promoted D. anglica growth on waterlogged surfaces even though D. anglica seedling survival was low.
The highest recruitment of the two Drosera species on the Sphagnum mat infers that the Sphagnum mat was the most suitable environment for seed germination. However, the seedling biomass in the sowing experiment was higher on the bare ground and waterlogged surface than on the Sphagnum mat for both Drosera species, particularly D. anglica. Therefore, the Sphagnum mat provided seedling beds but inhibit the seedling growth and survival, particularly for D. anglica. This species showed relatively slow growth and high mortality on the Sphagnum mat, with decreasing biomass when under shade, suggesting that growth and mortality were regulated by competition with Sphagnum. Sphagnum mosses absorb nutrients in the water through their epidermis, outcompeting vascular plants in nutrient capture (van Breemen 1995; Svensson 1995). An exception occurs when Sphagnum mosses transport excess water to shallow-rooted vascular plants, such as Drosera species (Adlassnig et al. 2005), where the roots overlap the Sphagnum layer (Malmer et al. 1994). Drosera shoots should show strong competition with Sphagnum mosses compared with deep-rooted vascular plants. D. anglica seedlings grew faster than D. rotundifolia on the bare-ground habitat where other plant species did not establish as quickly. The growth of the two Drosera species was conspicuously high 3 years after seed sowing. These results suggested that D. anglica was more susceptible to light and nutrient competition than D. rotundifolia. In contrast, D. rotundifolia established better on the Sphagnum mat, because it was susceptible to inundation, and more tolerant to shade and low nutrient levels compared with D. anglica.
Reproductive Strategies
The reproductive strategies differed between the two species. D. anglica used vegetative reproduction by producing daughters’ more than D. rotundifolia. The establishment of vegetation in inundated habitats using vegetative reproduction is one plant strategy (Griffith and Forseth 2003; Sosnová et al. 2010). The D. anglica population persisted in inundated habitats using daughter production as an anchoring mechanism, although the anchor did not allow long-distance dispersal. Following the exclusion of D. anglica by Sphagnum mosses on the waterlogged surface, D. anglica persistent using a vegetative reproduction strategy in unsuitable habitats.
Scale-Dependent Determinants on the Establishment of D. Anglica
The establishment of D. anglica was restricted to the Sphagnum mat at the habitat scale, and displaced to the waterlogged surface where inter-specific competition was decreased with vascular plants. At the wetland-ecosystem scale, Sphagnum mats develop a diverse topography, such as hummocks and palsas, in boreal regions (Charman 2002; Tsuyuzaki et al. 2008). Waterlogged surfaces develop within the hollows which were created in raised Sphagnum mats. Waterlogged surfaces do not develop when Sphagnum mats develop homogenously. Therefore, Sphagnum is a keystone taxon for habitat restoration in bog wetlands because of ecosystem engineering and their role as seedbeds for vascular plant species (Svensson 1995; Rochefort 2000). This factor is a prerequisite for developing waterlogged surfaces where D. anglica may establish.
In conclusion, D. anglica is endangered because of low competitive ability. In habitats where inter-specific competition was low, suitable habitat for D. anglica was provided for seedling germination and establishment on waterlogged surface created by Sphagnum mats. However, the waterlogged surface was not significantly preferable for seedling survival and growth. To conserve D. anglica populations, the management of overstory vascular plants and Sphagnum moss cover is critical to reduce inter-specific competition to solar energy and nutrient availability at the habitat scale.
References
Adlassnig W, Peroutka M, Lambers H, Lichtscheidl IK (2005) The roots of carnivorous plants. Plant and Soil 274:127–140
Anderson JT, Landi AA, Marks PL (2009) Limited flooding tolerance of juveniles restricts the distribution of adults in an understory shrub (Itea virginica; Iteaceae). American Journal of Botany 96:1603–1611
Baskin CC, Milberg P, Andersson L, Baskin JM (2001) Seed dormancy-breaking and germination requirements of Drosera anglica, an insectivorous species of the Northern Hemisphere. Acta Oecologica-International Journal of Ecology 22:1–8
Braun-Blanquet J (1964) Pflanzensoziologie. Springer Verlag, Wien, New York
Brook BW, Sodhi NS, Bradshaw CJA (2008) Synergies among extinction drivers under global change. Trends in Ecology & Evolution 23:453–460
Charman D (2002) Peatlands and Environmental Change. John Wiley, Hoboken
Coops H, van der Velde G (1995) Seed dispersal, germination and seedling growth of six hetelophyte species in relation to water-level zonation. Freshwater Biology 34:13–20
Crowder AA, Pearson MC, Grubb PJ, Langlois PH (1990) Biological flora of the British Isles. Drosera L Journal of Ecology 78:233–267
Dalling JW, Winter K, Nason JD, Hubbell SP, Murawski DA, Hamrick JL (2001) The unusual life history of Alseis blackiana: a shade-persistent pioneer tree? Ecology 82:933–945
Egawa C, Koyama A, Tsuyuzaki S (2009) Relationships between the developments of seedbank, standing vegetation and litter in a post-mined peatland. Plant Ecology 203:217–228
Eriksson O (2002) Ontogenetic niche shifts and their implications for recruitment in three clonal Vaccinium shrubs: Vaccinium myrtillus, Vaccinium vitis-idaea, and Vaccinium oxycoccos. Canadian Journal of Botany 80:635–641
Fenton NJ, Bergeron Y (2006) Facilitative succession in a boreal bryophyte community driven by changes in available moisture and light. Journal of Vegetation Science 17:65–76
Gelman A, Carlin JB, Stern HS, Rubin DB (2003) Bayesian data analysis. Chapman and Hall, London, UK
Gore AJP (ed) (1983) Mires: swamp, bog, fen and moor. Ecosystems of the World 4A. Elsevier, Amsterdam
Griffith AB, Forseth IN (2003) Establishment and reproduction of Aeschynomene virginica (L.) (Fabaceae) a rare, annual, wetland species in relation to vegetation removal and water level. Plant Ecology 167:117–125
Heijmans MMPD, Klees H, Berendse F (2002) Competition between Sphagnum magellanicum and Eriophorum angustifolium as affected by raised CO2 and increased N deposition. Oikos 97:415–425
Hoyo Y, Tsuyuzaki S (2013) Characteristics of leaf shapes among two parental Drosera species and a hybrid examined by canonical discriminant analysis and a hierarchical Bayesian model. American Journal of Botany 100:817–823
Huntke T (2007) The distribution of Drosera anglica Huds. in lower Saxony past and present–the extent of the decline of a raised bog specialist and its causes. Tuexenia: 241–253
IUCN Red List of Threatened Species. Version 2013. http://www.iucnredlist.org/
Japan Meteorological Agency (2012). http://www.jma.go.jp/jma/(in Japanese)
Jennings DE, Rohr JR (2011) A review of the conservation threats to carnivorous plants. Biological Conservation 144:1356–1363
Keddy PA, Fraser LH, Wisheu IC (1998) A comparative approach to examine competitive responses of 48 wetland plant species. Journal of Vegetation Science 9:777–786
Keddy PA (2010) Wetland ecology: Principles and Conservation. Cambridge University Press, Cambridge, UK
Koyama A, Tsuyuzaki S (2010) Effects of sedge and cottongrass tussocks on plant establishment patterns in a post-mined peatland, northern Japan. Wetlands Ecology and Management 18:135–148
Koyama A, Tsuyuzaki S (2013) Facilitation by tussock-forming species on seedling establishment collapses in an extreme drought year in a post-mined Sphagnum peatland. Journal of Vegetation Science 24:473–483
Malmer N, Svensson BM, Wallén B (1994) Interactions between Sphagnum mosses and field layer vascular plants in the development of peat-forming systems. Folia Geobotanica et Phytotaxonomica 29:483–496
Malmer N, Albinsson C, Svensson BM, Wallén B (2003) Interferences between Sphagnum and vascular plants: effects on plant community structure and peat formation. Oikos 100:469–482
Ministry of the Environment Government of Japan (2013) Japan integrated biodiversity information system. Red list of Threatened Plant of Japan, revisedth edn. Ministry of the Environment Government of Japan, Tokyo, Japan
Murray BR, Thrall PH, Gill AM, Nicotra AB (2002) How plant life-history and ecological traits relate to species rarity and commonness at varying spatial scales. Austral Ecology 27:291–310
Nishimura A, Tsuyuzaki S, Haraguchi A (2009) A chronosequence approach for detecting revegetation patterns after Sphagnum-peat mining, northern Japan. Ecological Research 24:237–246
Nordbakken JF (1996) Plant niches along the water-table gradient on an ombrotrophic mire expanse. Ecography 19:114–121
Nordbakken JF, Rydgren K, Økland RH (2004) Demography and population dynamics of Drosera anglica and D. rotundifolia. Journal of Ecology 92:110–121
Poorter L (2007) Are species adapted to their regeneration niche, adult niche, or both? The American Naturalist 169:433–442
Rivadavia F, Kondo K, Kato M, Hasebe M (2003) Phylogeny of the sundews, Drosera (Droseraceae), based on chloroplast rbcL and nuclear 18S ribosomal DNA sequences. American Journal of Botany 90:123–130
Rochefort L (2000) New frontiers in bryology and lichenology–Sphagnum–A keystone genus in habitat restoration. Bryologist 103:503–508
Römermann C, Tackenberg O, Jackel AK, Poschiod P (2008) Eutrophication and fragmentation are related to species’ rate of decline but not to species rarity: results from a functional approach. Biodiversity and Conservation 17:591–604
Sosnová M, van Diggelen R, Klimešová J (2010) Distribution of clonal growth forms in wetlands. Aquatic Botany 92:33–39
Spiegelhalter D, Thomas A, Best N, Lunn D (2003) WinBUGS version 1.4 User manual. http://www.mrc-bsu.cam.ac.uk/bugs
Svensson BM (1995) Competition between Sphagnum fuscum and Drosera rotundifolia: a case of ecosystem engineering. Oikos 74:205–212
ten Brink D-J, Hendriksma HP, Bruun HH (2013) Habitat specialization through germination cueing: a comparative study of herbs from forests and open habitats. Annals of Botany 111:283–292
Tsuyuzaki S, Sawada Y, Kushida K, Fukuda M (2008) A preliminary report on the vegetation zonation of palsas in the Arctic National wildlife Refuge, northern Alaska, USA. Ecological Research 23:787–793
van Breemen N (1995) How Sphagnum bogs down other plants. Trends in Ecology and Evolution 10:270–275
Verhulst J, Montana C, Mandujano MC, Franco M (2008) Demographic mechanisms in the coexsistence of two closely related perennials in a fluctuating environment. Oecologia 156:95–105
Wolf E, Gage E, Cooper DJ (2006) Drosera anglica Huds. (English sundew): A technical conservation assessment. Report prepared for the USDA Forest Service, Rocky Mountain Region, Species Conservation Project [online]. Website http://www.fs.usda.gov/ Internet/FSE_DOCUMENTS/stelprdb5250872.pdf./[accessed 30 August 2012]
Young AS, Chang SM, Sharitz RR (2007) Reproductive ecology of a federally endangered legume, Baptisia arachnifer and its more widespread congener, B. lanceolata (Fabaceae). American Journal of Botany 94:228–236
Zietsman J, Dreyer LL, Esler KJ (2008) Reproductive biology and ecology of selected rare and endangered Oxalis L. (Oxalidaceae) plant species. Biological Conservation 141:1475–1483
Acknowledgments
We wish to thank members of the plant ecology laboratory for their support, and T. Kubo for his statistical analyses. We are grateful to staff from the Ministry of Environment of Japan and the Toyotomi Town Office for research permission and support. This work is partly supported by grants from Japan Society for the Promotion of Science, Expo '90 Foundation and Global COE grants to Hokkaido University.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Hoyo, Y., Tsuyuzaki, S. Habitat Differentiation Between Drosera anglica and D. rotundifolia in a Post-Mined Peatland, Northern Japan. Wetlands 34, 943–953 (2014). https://doi.org/10.1007/s13157-014-0555-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-014-0555-9