
Abstract
Understanding the relationships between hydrology and salinity and plant community structure and production is critical to allow predictions of wetland responses to altered water management, changing precipitation patterns and rising sea-level. We addressed how salinity, water depth, hydroperiod, canal inflows, and local precipitation control marsh macrophyte aboveground net primary production (ANPP) and structure in the coastal ecotone of the southern Everglades. We contrasted responses in two watersheds - Taylor Slough (TS) and C-111 - systems that have and will continue to experience changes in water management. Based on long-term trajectories in plant responses, we found continued evidence of increasing water levels and length of inundation in the C-111 watershed south of the C-111 canal. We also found strong differentiation among sites in upper TS that was dependent on hydrology. Finally, salinity, local precipitation and freshwater discharge from upstream explained over 80 % of the variance in Cladium ANPP at a brackish water site in TS. Moreover, our study showed that, while highly managed, the TS and C-111 watersheds maintain legacies in spatial pattern that would facilitate hydrologic restoration. Based on the trajectories in Cladium and Eleocharis, shifts in plant community structure could occur within 5–10 years of sustained water management change.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In the southern Everglades, key drivers of ecosystem structure and plant production are duration and depth of inundation, P availability and salinity (Noe et al. 2001; Childers et al. 2006a). The influence of these drivers varies across spatial and temporal scales. For example, in freshwater marshes nearest to canals, concerns about P enrichment from inflowing water prevail (Surratt et al. 2012) and the effects of P enrichment have been well documented both in the field and experimentally (Childers et al. 2003, Daoust and Childers 2004). With respect to salinity however, while increased salinity has been observed in porewater of freshwater marshes of the Everglades upper ecotone, especially in the dry season (Troxler 2012) and mangrove transgression and sawgrass die-off also observed (Ross et al. 2000), the effects of salinity on freshwater species has been related neither with experiments nor explicitly with long-term trends. While, the relationship between salinity, hydrology and primary production is well documented for many types of coastal wetlands (Howard and Mendelsshohn 1999; Merino et al. 2010), long-term trends in salinity and hydrology (including precipitation and discharge) and relationships with structure and production of freshwater species in the southern coastal Everglades have not been evaluated.
Sea levels in south Florida are conservatively predicted to rise by 0.60 m by 2060 (Zhang et al. 2011). As sea levels rise, salinity and inundation will increase in fresh and brackish water areas of the Florida coastal zone (Florida Oceans and Coastal Council (FOCC) 2010; Pearlstine et al. 2010). South Florida is particularly vulnerable to sea-level rise given the low topographic incline from Florida Bay inland and large-scale hydrologic diversion from the coastal zone that has amplified salinity transgression in some areas (Ross et al. 2000). Thus, water delivery from upstream, driven by the interplay of water management and seasonal precipitation, is a key driver of marsh ecosystem structure and production throughout the southern Everglades. Large-scale restoration efforts aim to increase freshwater delivery to the coastal Everglades, with the potential to counter salinity transgression documented for the upper estuarine ecotone (Ross et al. 2000; Troxler 2012). Understanding the relationship between hydrologic variables (including water level, hydroperiod and salinity) and plant community structure and production is critical to allow predictions of wetland responses to altered water management, changing precipitation patterns and rising sea-level in the southern coastal Everglades.
Over a 10-year period, we have investigated how changes in hydroperiod, hydropattern, and salinity have influenced emergent macrophyte communities dominated by sawgrass (Cladium jamiacense) and with variable presence and densities of spike rush (Eleocharis cellulosa). Of these two freshwater species, given similar depth and inundation, Eleocharis shows lower mortality as compared with Cladium and found in areas with generally longer duration of inundation (Wetzel 2001) and were thus anticipated to be useful in characterizing long-term trends in hydrology and salinity. Our central objective was to evaluate how interannual variation in hydrologic and salinity drivers influenced Cladium aboveground net primary production (ANPP) and the presence of Eleocharis stems in the southern Everglades by evaluating these trends over site-specific and watershed (regional) scales. We tested the following hypotheses: 1) If increased freshwater inflows to the southern Everglades lead to longer marsh hydroperiods and deeper water, Cladium ANPP will decline and Eleocharis stem densities will increase, and; 2) If increased freshwater inflows to the southern Everglades lead to longer periods of fresh water (i.e. non-detectable salinity) in the upper estuarine ecotone, Cladium ANPP will increase and Eleocharis stem densities will decline.
Materials and Methods
Study Site
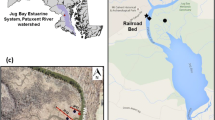
The southern Everglades study landscape includes the Taylor Slough and the C-111 Basin watersheds (Fig. 1). These two basins are the dominant contributors to water delivered to the freshwater and estuarine portions of eastern and southern Everglades National Park (ENP; Davis and Ogden 1994). These drainages, and the downstream mangrove estuaries that form the northern shore of Florida Bay (also included in the southern Everglades study area), will continue to experience significant hydrological changes as a result of anticipated Everglades restoration activities. The Taylor Slough watershed has experienced reduced freshwater inflow over the last 50 years some of which has been diverted to the C-111 Basin through the Barnes Sound and more recently (1997) as overland flow (Davis and Ogden 1994, Parker 2000). In 1998, the Southern Everglades Ecosystem Monitoring program began, collectively funded by the South Florida Water Management District (SFWMD) and ENP. This research program was designed to address the need for continued monitoring and adaptive assessment of key management drivers and to assess how hydrologic restoration has and will impact ecosystem dynamics in the Taylor Slough and C-111 basins of the southern Everglades. Water management practices result in markedly different hydrologic patterns in these two adjacent wetland drainages (Rudnick et al. 1999). In the C-111 Basin, past hydrologic modifications have included a levee that was removed from the C-111 canal in 1997 to restore water flow and increase hydroperiods in the downstream wetlands (Parker 2000). Since that time, water has entered the C-111 basin through overland flow across this levee removal zone once the canal reaches bankfull water level. In upper Taylor Slough, pump operations have been altered since 2001 with water now entering the system through a series of retention ponds across a small berm removal area (Fig. 1) just north of a now defunct pump structure. Anticipated hydrologic restoration actions in this region will direct more water to Taylor Slough, which is expected to result in less water delivery to the C-111 Basin (USACE 2011).

Map of Florida (inset) indicating the location of Everglades National Park ( ). The map shows sites monitored along Taylor Slough and the C-111 Basins with location of L31-W canal (dashed line) with levee breach indicated with dark line at southern end of L31-W. Water control structures labelled as S332-D, S18-C and S197. Sites W1 and W3 in the C111 Basin correspond to sites TS/Ph4 and TS/Ph5
). The map shows sites monitored along Taylor Slough and the C-111 Basins with location of L31-W canal (dashed line) with levee breach indicated with dark line at southern end of L31-W. Water control structures labelled as S332-D, S18-C and S197. Sites W1 and W3 in the C111 Basin correspond to sites TS/Ph4 and TS/Ph5
The study site encompasses the Taylor Slough watershed and C-111 Basin (Fig. 1). This landscape is made up largely of short hydroperiod marl marsh with estuarine mangrove wetlands to the south. Four sites in Taylor Slough form a long-term monitoring transect of the Florida Coastal Everglades LTER Program (FCE LTER; TS/Ph1, TS/Ph2, TS/Ph3, TS/Ph6; TS/Ph1 corresponds to Lat/Long 25.4, −80.6). The transect is anchored just north of the upstream source of canal inflow, where water enters the marsh through a breach in a levee that borders the eastern margin of Everglades National Park in this area (L31W levee; Fig. 1). The C-111 Basin sites are located both north (N1 and N2) and south (TS/Ph-4 also W1 and TS/Ph-5 also W3) of the C-111 canal, with one site east of both the canal and U.S. Highway 1 (USC1; Fig. 1). Seasonal variability in water levels and inundation in the C-111 Basin landscape are controlled by a combination of local rainfall, which dominates at N1, N2, and USC1, and activities associated with water management, which dominates at the TS/Ph-4 and TS/Ph-5 sites.
At each site, we quantified Cladium jamaicense aboveground live standing crop (biomass) and estimated ANPP and quantified Eleocharis cellulosa stem density. Macrophyte measurements were made every 2 months in triplicate 1 m2 plots at each of the sites. Cladium biomass and ANPP were quantified following the non-destructive phenometric method described in Daoust and Childers (1998) and using the biomass and mortality models described in Childers et al. (2006a) to estimate ANPP. All Eleocharis stems were counted bimonthly to obtain total stem density m−2. Annual Eleocharis stem density was assessed as average and maximum density of bimonthly estimates per year. Maximum Eleocharis stem density is a bimonthly maximum of the average of three replicate plots in a year. Due to changes in monitoring priorities and periodic funding gaps, data are not available for all sites in all years of the monitoring program. Water depth relative to the soil surface was recorded hourly with ultrasonic water level recorders (Infinity Instruments, Daytona Beach FL) at each of the sites. Discharge (m3 s−1) data from the S-332D pump and at the S-18C and S-197 structures were downloaded from DBHYDRO (SFWMD; http://www.sfwmd.gov/dbhydroplsql/) and used to estimate canal inflow rates for Taylor Slough and the C-111 Basin, respectively. We used precipitation data collected at each site or from ENP meteorological stations located in Taylor Slough (TSB=TS/Ph1, TS/Ph2, TS/Ph3; TR=TS/Ph6 and EVER8 = TS/Ph4, TS/Ph5, N1 and N2; ENP data not publically available) when site-specific data were not available. Water samples for salinity were collected every 18 h with ISCO automatic water samplers (autosampler) deployed at each of the sites (see Childers et al. 2006b for sampling and analytical details).
In this investigation of hydrology and salinity factors on Cladium ANPP and Eleocharis stem density, we quantified hydrologic parameters including mean annual water depth (cm), maximum annual water depth (cm), hydroperiod (days), Depth*Days (m*d; a hybrid variable that accounts for both mean water depth and hydroperiod; see Childers et al. 2006a for details), mean annual canal inflow (m3 s−1), and total annual canal inflow (m3 yr-1). Precipitation data were in annual totals (cm yr−1) and salinity variables at the TS/Ph6 site included mean annual salinity (ppt), maximum annual salinity (ppt), and the fraction of the year with salinity >0, <5, and >30 ppt (# days) based on divisions of no, measurable but low (with no cited effects; Macek and Rejmankova 2007) and high salinity. We used best-fit regression analysis to test the effects of time and hydrologic parameters on site-specific (at each site independently) responses of Cladium ANPP and Eleocharis stem density at all sites with the exception of the salinity parameters that were evaluated at TS/Ph6 only – the only site with measurable salinity. We also tested the effect of hydrologic variation on regional macrophyte patterns. For regional analyses, we grouped sites into freshwater Taylor Slough (TS/Ph 1, TS/Ph 2 and TS/Ph 3), C-111 south of the canal (TS/Ph 4 and TS/Ph 5) and C-111 north and east of the canal (N1, N2, USC1) based on geographic location and common hydrologic histories (Parker 2000; USACE 2011). Our approach was to determine trends in hydrology and salinity, and relate these trends to variation in site-specific and regional patterns of production and stem density of two, dominant freshwater species in the freshwater-coastal ecotone of the southern Everglades.
Results
Trends in Hydrology and Salinity
We monitored hydrologic variables, precipitation, and salinity for the eight sites in our study area (Table 1; hydrologic data for N2 were not available). The TS/Ph2 site, south of Main Park Road in Taylor Slough, had the highest mean and maximum annual water levels, longest hydroperiod, and highest values of Depth*Days. Sites with moderate inundation depths but long hydroperiods were TS/Ph3, TS/Ph4, TS/Ph5 and N1. The driest site was the TS/Ph6 estuarine site in lower Taylor Slough.
All sites but TS/Ph6 were characterized as freshwater marshes with no measurable salinity. However, at TS/Ph6 the inter annual variation in salinity was pronounced (Table 2). Over our 12 year period of record, mean annual salinity ranged from 1.5 to 16.4 ppt. Maximum salinities late in the dry season were often higher than oceanic salinities; the maximum reached nearly 50 ppt in 2005. In 2004, 2005 and 2008, salinity exceeded 30 ppt for roughly 25 % of the year while 2000, 2002 and 2003 were particularly wet years and salinity at TS/Ph6 never exceeded 30 ppt.
Temporal Patterns and Hydrologic Drivers of Eleocharis Density
At the sites south, north and east of the C-111, Eleocharis stem density varied significantly with time at the TS/Ph 4, TS/Ph5, and N2 sites (Fig. 2). There were significant increases in average annual stem density at TS/Ph4 and both average and maximum stem density at TS/Ph5. These relationships were described by: 1) a polynomial model for TS/Ph4 (r 2 = 0.83, F = 9.81, p = 0.029, y = 1.84x2-4.36x-8,715) and 2) linear relationships for TS/Ph5 (Average: r 2 = 0.81, F = 17.0, p = 0.015, y = 4.65x-9,293 and Maximum: r 2 = 0.70, F = 9.18, p = 0.039, y = 5.97x-11,931). We also found significant increases in average stem density at N2 that were described by linear relationships with time (r 2 = 0.91, F = 20.66, p = 0.045, y = 35.66x-71,545).

Interannual variability in average Eleocharis stem density at C-111 sites south (TS/Ph4, TS/Ph5), north (N1 and N2) and east (USC1) of the C-111 canal. Error bars are standard error
Unlike other sites in the C-111, we found that average water level and Depth*Days explained 84 and 78 % of the interannual variance in mean Eleocharis density at TS/Ph5 in negative relationships (Average: F = 21.05, p = 0.010, y = −2.73x + 80.69 and Depth*Days: F = 14.06, p = 0.020, y = −0.71x + 69.96). When sites within regions south of C-111 and north and east of C-111 were evaluated, there were no significant relationships between Eleocharis stem density and hydrologic drivers. Thus, there were site-specific responses in Eleocharis to hydrologic drivers at TS/Ph5 only and the sites within regions were not strongly contrasted by hydrologic condition.
Eleocharis stem density showed strong temporal declines in average and maximum annual values at TS/Ph6 but not at any of the freshwater TS sites (Fig. 3). At the TS/Ph6 site, non-linear temporal trends explained 90 and 95 % of the variance in average and maximum stem densities, respectively (F = 21.96, p = 0.003, y = 0.43x2 − 1.48x + 2,982 and F = 45.52, p = 0.0006, y = 0.59x2 − 2.23x + 4,494, respectively). Eleocharis density declined to a threshold in 2007–2008 and has maintained low mean and maximum densities since that time. Eleocharis has never been present at the TS/Ph1 site and at TS/Ph3 stem densities do not exceed average and maximum annual values of 8 and 15, respectively (Fig. 3). In relating hydrologic variation to site-specific Eleocharis stem density, there were no site-specific relationships in TS.

Interannual variability in average Eleocharis stem density at Taylor Slough sites TS/Ph2, TS/Ph3 and TS/Ph6. Eleocharis was not present at TS/Ph1. Error bars are standard error
Regional patterns in Eleocharis at our freshwater sites in the Taylor Slough (TS/Ph1, TS/Ph2 and TS/Ph3) included significant positive relationships between mean stem density and mean water level (r 2 = 0.78, F = 70.94, p < 0.0001, y = −9.29 + 0.99x; Fig. 4a), maximum water level (r 2 = 0.74, F = 56.45, p < 0.0001, y = −21.2 + 0.62x), hydroperiod (r 2 = 0.23, F = 6.00, p = 0.03, y = −23.5 + 0.14x), and Depth* Days (r 2 = 0.76, F = 63.86, p < 0.0001, y = 0.27x-5.97; Fig. 4b). We also found similar relationships between Eleocharis maximum stem density and these variables although relationships with mean stem density were more robust.

Regression models relating variation in a mean water level and b Depth*Days to Eleocharis stem density and c mean water level and d Depth*Days Cladium ANPP for the Taylor Slough freshwater watershed sites
Temporal Patterns and Hydrologic Drivers of Cladium ANPP
In contrast to trends for Eleocharis, we found significant temporal declines in Cladium ANPP at both sites that were south of the C-111 canal (Fig. 5a). However, time was a more important explanatory variable of Cladium ANPP at TS/Ph5 as compared with TS/Ph4 that described 63 and 33 % of the variance, respectively (TS/Ph5: F = 17.29, p = 0.002, y = −17.78x + 35,939 and TS/Ph4: F = 4.96, p = 0.050, y = −10.61x + 21,566). We found no temporal trends in ANPP for sites north (N1, N2) or east (USC1) of the C-111 canal, perhaps due to much shorter datasets at these sites (Fig. 5b). There were also no significant effects of hydrologic drivers on Cladium ANPP for any of our C-111 sites.

Cladium aboveground net primary production (g m−2 yr−1) at sites a south (TS/Ph4, TS/Ph5) and b north (N1 and N2) and east (USC1) of the C-111 canal. Error bars are standard error
We found different temporal trends in Taylor Slough Cladium ANPP depending on the site (Fig. 6). At TS/Ph1, there was a significant increase in ANPP with time (r 2 = 0.63, F = 17.02, p = 0.002, y = 16.44x + −32,679) while at TS/Ph2 and TS/Ph3 we found significant temporal declines in ANPP. At TS/Ph2, 36 % of the variance was explained by this trend (r 2 = 0.36, F = 5.75, p = 0.037, y = −12.75x + 25,801) and at TS/Ph3, we found that 54 % of the variation in ANPP was explained by change with time (r 2 = 0.54, F = 11.74, p = 0.006, y = −25.21x + 50,958). At the estuarine ecotone site, TS/Ph6, the relationship between time and ANPP was curvilinear. The decrease in ANPP between 2000 and 2005 was marginally significant (p = 0.06) but for 2006–2010 we found a significant temporal increase in ANPP (r 2 = 0.83, F = 19.06, p = 0.012, y = 59.05x + −118,262).

Cladium aboveground net primary production (g m−2 yr−1) at Taylor Slough marsh sites. Error bars are standard error
Tests of the relationship between hydrologic drivers and site-specific Cladium ANPP in freshwater sites in Taylor Slough were not significant. However, there were significant negative relationships between hydrologic drivers and Cladium ANPP within the Taylor Slough freshwater marsh region. At the freshwater sites, ANPP decreased with higher mean water level (r 2 = 0.37, F = 18.35, p = 0.0004, y = −3.64x + 418; Fig. 4c), maximum water level (r 2 = 0.40, F = 16.48, p = 0.0002, y = −2.34x + 471), and Depth*Days (r 2 = 0.33, F = 14.4, p = 0.0008, y = −0.95x + 402; Fig. 4d).
Salinity Effects on Cladium ANPP and Eleocharis Density
At TS/Ph6, we tested the relationship between salinity and precipitation and Cladium ANPP and Eleocharis stem density. There were no relationships for Eleocharis stem density. However, combinations of these drivers did explain trends in Cladium ANPP. Cladium ANPP decreased as both precipitation and the number of salinity days exceeding 30 ppt increased [ANPP = 708–2.67(Precip.) – 2.42(SalDays > 30); r 2 = 0.55, p = 0.03]. Including discharge from the S-332D pump described an additional 28 % of the variance in Cladium ANPP, with a positive relationship between this measure of water management inflow and production [ANPP = 712–4.67(Precip) – 4.66(SalDays > 30) + 0.006(InflowSum); r 2 = 0.83, p = 0.023].
Discussion
Evaluating changes in both Eleocharis and Cladium at sites over an extensive spatial scale and decadal time scales revealed important corollary trends and influence of hydrologic and climatic drivers. Additionally, these findings may be used to make more accurate predictions of how these regions of the wetland landscape may respond to future changes in water management associated with Everglades restoration efforts.
Since the hydrologic ranges of Eleocharis cellulosa and Cladium jamaicense can generally be differentiated under field conditions (Wetzel 2001), evaluating the direction of their responses simultaneously provides supporting evidence for wetland ecosystem changes in the Taylor Slough and C-111 regions of the southern Everglades. A recent experiment by Miao and Zou (2012) helps to identify more precise hydrologic tolerances for these species. In this study, the authors exposed the species to 9 months of inundation at water levels of 20 and 60 cm depth, respectively. They showed increased stem height of Eleocharis cellulosa plants exposed to deeper water levels, however this coincided with a 10 % increase in mortality and decreased biomass. At 20 cm water level, Eleocharis experienced 0 % mortality. Thus, a 9-month inundation period at 60 cm or greater water depth may approximate the upper hydrologic tolerance for Eleocharis. Further, experimental manipulation of water depth variation illustrated that Eleocharis cellulosa could adjust its biomass allocation to rapidly (< 10 weeks) respond to new water level conditions (Edwards et al. 2003). In the Miao and Zou (2012) experiment, the authors also showed 50 and 80 % mortality in Cladium jamaicense exposed to 20 and 60 cm water depths. This greater increase in mortality between 20 and 60 cm water depth also coincided with a nearly 50 % decrease in biomass (Miao and Zou 2012). Furthermore, Miao and Zou (2012) suggest that increased mortality in Cladium is related to an inability to shift aboveground/belowground allocation to adjust to changing hydrologic conditions. Consequently, these data suggest that, given the same duration of inundation, Eleocharis cellulosa is not only more water tolerant but also more tolerant of fluctuating water levels suggesting it is generally more robust within this range of hydrologic conditions. Moreover, these and other experimental data help to define the hydrologic conditions for these species and guide our interpretation of site-specific macrophyte responses, regional scale patterns and recent landscape change in the southern Everglades.
In the C-111 Basin, at sites south of the C-111 canal (TS/Ph4 and TS/Ph5), there were significant changes in both Eleocharis stem density and Cladium ANPP over time. Eleocharis responded with an increase in stem density over time while Cladium ANPP declined. In their analysis of hydrologic controls on Cladium ANPP rates, Childers et al. (2006a) also found a negative relationship, but with a shorter time-series of data. We expected to see similar relationships. While changes in Cladium ANPP in this area are consistent with increased water level and length of inundation, temporal changes in Eleocharis coincided with decreasing water level at TS/Ph5. Since 1998, overland flow into the C-111 south of the canal was reintroduced with species responses characteristic of an increasingly wetter environment. However, at the lower TS/Ph5 site, other factors may have influenced the relationship between hydrology and Eleocharis stem density. Nutrient availability and local redox intensity are also factors that have been shown to influence Eleocharis growth (Busch et al. 2004, Chen et al. 2005). It is plausible that conditions such as topographic position relative to the water table and connection to upstream surface water flow likely also modulate site-specific species responses and shifts in community structure. Regardless, the trajectory over time illustrates that despite the possible influence of other factors, both Eleocharis and Cladium are trending toward responses to a wetter environment.
In the C-111 region, north of the C-111 canal, we found an increase in Eleocharis over time at the southern N2 site without significant change at the upper site (N1) within this region. However, there was no relationship between macrophyte community response and variation in hydrologic parameters. Further, the conditions resulting in higher Eleocharis density at N2 have not coincided with changes in Cladium ANPP. Because hydrology data were not available for N2, we could not directly test its influence on Eleocharis response. It is not clear if active water management has contributed to this trend, but withholding water delivery at the S-18C structure would likely increase hydrologic head in this area. Given that the marsh is cut off from direct canal inflows and is largely impounded, the downstream levee might also increase ponding in this area just north of the C-111 levee road. Other localized hydrologic changes must also contribute to the trend we observed as the levee has been in place since the 1960’s and pronounced increase in Eleocharis stem density has only been observed to occur within the last 5 years.
Within Taylor Slough, we evaluated spatial trends based on differentiation between freshwater sites within upper TS as compared with TS/Ph6 in lower TS, as well as within the upper TS region. Among the freshwater TS sites, the uppermost site TS/Ph1 was the driest followed by TS/Ph3 with the wettest site just north of Main Park Road in ENP (TS/Ph2). Macrophyte patterns at TS/Ph1 were consistent with a drier hydrologic condition; Eleocharis was absent and the temporal trend illustrates that Cladium ANPP has increased at this site. This increase in Cladium ANPP over time could not be clearly linked with changes in hydrology probably due to missing data from earlier years. In central Taylor Slough (TS/Ph2 and TS/Ph3), there were no site-specific changes over time or response to hydrologic drivers in Eleocharis stem density. However, taken together, there was a regional trend of higher Eleocharis stem density with higher water level and Depth*Days (Fig. 4a and b). This relationship with hydrology is likely a function of hydrologic conditions that were contrasted by relatively dry sites and wet sites and illustrates the strong link between Eleocharis stem density and hydrology. Over time, there were site-specific declines in Cladium ANPP and these coincided with increasing water level and Depth*Days (Fig. 4c and d). Thus, conditions illustrate the initial shift in community structure that has been observed at other sites – initially a decline in Cladium ANPP that is often followed by an increase in Eleocharis stem density. This is illustrated by a trajectory of linear decline in Cladium ANPP followed by curvilinear increase in Eleocharis stem density in the range of 5–10 years.
In the estuarine ecotone, the TS/Ph6 site was the only site to have pronounced surface water salinity. Here, Eleocharis stem density has declined over time (from highest densities in 2003 and 2004) while Cladium ANPP has increased over the period between 2006 and 2010. In terms of the salinity parameters we tested, only the number of days in a given calendar year in which surface water salinity exceeded 30 ppt had a significant negative influence on Cladium ANPP in combination with other explanatory variables. Macek and Rejmankova (2007) found that low levels of salinity had no effect on Cladium jamaicense biomass, but the maximum salinity level used in that experiment was 5 ppt. Similarly, we found no relationship between Cladium ANPP and number of days with measurable salinity under 5 ppt. However, in a related study by Rejmankova and Macek (2008), the authors showed increased root phosphatase activity in Cladium jamaicense between 0.5 and 5 ppt and suggest this is a response to physiological stress. The effect of increased precipitation and freshwater discharge at TS/Ph6 also contributes to variation in plant community responses. We found that the effect of salinity >30 ppt was modulated by both increased freshwater flow from S-332D and local precipitation. Including these parameters in the model explained over 80 % of the variation in Cladium ANPP at the upper ecotone site TS/Ph6. This illustrates the significant effect that water management and climate exert on patterns of Cladium production in the marginal (i.e. ecotone) areas of the southern Everglades. A recent modeling study suggests the potential for dramatic changes in the structure and production of macrophyte communities across the Everglades with changes in atmospheric CO2 concentrations predicted to decrease precipitation and result in lower average water depths and reduced inundation length across the system (Todd et al. 2012).
Hydrologic restoration of the southern Everglades is incremental but underway. For instance, Phase I of the C-111 Spreader Canal project, set to be completed by 2012, is an effort to increase discharge into TS while minimizing pulsed flows from water management (USACE 2011). Thus, with water management operations that increase total discharge while decreasing discharge variability, further declines in Cladium ANPP would be predicted for TS freshwater sites in favor of more water-tolerant vegetation. At the upper ecotone of TS, Cladium ANPP could be expected to increase due to lower average salinity given otherwise similar or slightly wetter conditions in the range of 20 cm mean water depth and <9-months inundation based on experimental work (e.g. Miao and Zou 2012) and results of this study. This study further illustrates that while highly managed, the Taylor Slough and C-111 watersheds maintain legacy spatial patterns that would facilitate restoration of water depths and water delivery and that shifts in community structure, through changes in production and density of dominant species, could occur within 5–10 years.
Throughout the ecotone, observations of low or senescent sawgrass biomass is associated with a region of the lower coastal ecotone that is termed the “white zone” (Ross et al. 2000). While our extensive monitoring program has illustrated pronounced effects of salinity and hydrology across the Taylor Slough and C111 regions of the southern Everglades landscape, other factors known to control ANPP and species responses were not evaluated (i.e. soil phosphorus availability and redox intensity; Daoust and Childers 2004; Chen et al. 2005). Thus, a portion of the unexplained variance could be further resolved by a better understanding of these environmental drivers. While salinity is clearly an important driver of ecosystem production across the FCE coastal landscape (Childers et al. 2006a, b; Barr et al. 2010), the specific and relative influences of salinity, inundation and P availability require further evaluation with manipulative experiments. As sea level rise and salinity transgression continue to influence freshwater marsh macrophyte community structure, the influence of salinity, hydrology and P availability will be modulated by water delivery.
References
Barr JG, Engel VC, Fuentes JD, Zieman JC, O’Halloran TL, Smith TJ III, Anderson GH (2010) Controls on mangrove forest-atmosphere carbon dioxide exchanges in western Everglades National Park. Journal of Geophysical Research Biogeosciences 115:G02020. doi:10.1029/2009JG001186
Busch J, Mendelssohn IA, Lorenzen B, Brix H, Miao S (2004) Growth responses of the Everglades wet prairie species Eleocharis cellulose and Rhynchospora tracyi to water level and phosphate availability. Aquatic Botany 78:37–54
Chen H, Mendelssohn IA, Lorenzen B, Brix H, Miao S (2005) Growth and nutrient responses of Eleocharis cellulose (Cyperaceae) to phosphate level and redox intensity. American Journal of Botany 92:1457–1466
Childers DL, Doren RF, Noe GB, Rugge M, Scinto LJ (2003) Decadal change in vegetation and soil phosphorus patterns across the Everglades landscape. Journal of Environmental Quality 32:344–362
Childers DL, Iwaniec D, Rondeau D, Rubio G, Verdon E, Madden CJ (2006a) Responses of sawgrass and spikerush to variation in hydrologic drivers and salinity in Southern Everglades marshes. Hydrobiologia 569:273–292
Childers DL, Boyer JN, Davis SE, Madden CJ, Rudnick DT, Sklar FH (2006b) Relating precipitation and water management to nutrient concentrations in the oligotrophic “upside-down” estuaries of the Florida Everglades. Limnology and Oceanography 51:602–616
Daoust RJ, Childers DL (1998) Quantifying aboveground biomass and estimating productivity in nine Everglades wetland macrophytes using a non-destructive allometric approach. Aquatic Botany 62:115–133
Daoust RJ, Childers DL (2004) Ecological effects of low-level phosphorus additions on two plant communities in a neotropical freshwater wetland ecosystem. Oecologia 141(4):672–686
Davis SM, Ogden JC (1994) Everglades, the ecosystem and its restoration. St. Lucie Press, Florida
Edwards AL, Lee DW, Richards JH (2003) Responses to a fluctuating environment: effects of water depth on growth and biomass allocation in Eleocharis cellulosa Torr. (Cyperaceae). Canadian Journal of Botany 81:964–975
Florida Oceans and Coastal Council (FOCC) (2010) Climate change and sea-level rise in Florida: an update of the “The effects of climate change on Florida’s ocean and coastal resources.” [2009 report] Tallahassee, Florida, 26p
Howard RJ, Mendelsshohn IA (1999) Salinity as a constraint on growth of oligohaline marsh macrophytes. I. Species variation in stress tolerance. American Journal of Botany 86:785–794
Macek P, Rejmankova E (2007) Response of emergent macrophytes to experimental nutrient and salinity additions. Functional Ecology 21:478–488
Merino JH, Huval D, Nyman AJ (2010) Implication of nutrient and salinity interaction on the productivity of Spartina patens. Wetlands Ecology and Management 18:111–117
Miao SL, Zou CB (2012) Effects of inundation on growth and nutrient allocation of six macrophytes in the Florida Everglades. Ecological Engineering 42:10–18
Noe G, Childers DL, Jones RD (2001) Phosphorus biogeochemistry and the impacts of phosphorus enrichment: why are the Everglades so unique? Ecosystems 4:603–624
Parker III FM (2000) Changes in water inputs and nutrient loading after restoration of water flow to a Southern Ever- glades wetland landscape. MS Thesis, Florida International University, Miami, FL, 128 pp
Pearlstine LG, Pearlstine EV, Aumen NG (2010) A review of the ecological consequences and management implications of climate change for the Everglades. Journal of the North American Benthological Society 29:1510–1526
Rejmankova E, Macek P (2008) Response of root and sediment phosphatase activity to increased nutrients and salinity. Biogeochemistry 90:159–169
Ross MS, Meeder JF, Sah JP, Ruiz PL, Telesnicki GJ (2000) The southeast saline Everglades revisited: 50 years of coastal vegetation change. J. Vegetation Science 11:101–112
Rudnick DT, Chen Z, Childers DL, Boyer JN, Fontaine TD III (1999) Phosphorus and nitrogen inputs to Florida Bay: the importance of the Everglades watershed. Estuaries 22:398–416
Surratt D, Shinde D, Aumen N (2012) Recent cattail expansion and possible relationships to water management: changes in Upper Taylor Slough (Everglades National Park, Florida, USA). Environmental Management 49:720–733
Todd MJ, Muneepeerakul R, Miralles-Wilhelm F, Rinaldo A, Rodriguez-Iturbe I (2012) Possible climate change impacts on the hydrological and vegetative character of Everglades National Park, Florida. Ecohydrology 5:326–336
Troxler TG (2012) Ecological monitoring of southern Everglades wetlands, mangrove transition zone and “white zone” interactions with Florida Bay. Annual Report to the South Florida Water Management District, West Palm Beach, Fl. 65p
United States Army Corps of Engineers (USACE) (2011) Central and Southern Florida project, C-111 spreader canal western project: final project implementation report and environmental impact statement. http://www.evergladesplan.org/pm/projects/docs_29_c111_pir.aspx
Wetzel PR (2001) Plant community parameter estimates and documentation for the Across Trophic Level System Simulation (ATLSS). Institute for Environmental Modeling, University of Tennessee-Knoxville, TN
Zhang K, Dittmar J, Ross M, Bergh C (2011) Assessment of sea level rise impacts on human population and real property in the Florida Keys. Climate Change 107:129–146
Acknowledgments
This work would not have been possible without the herculean field efforts by Damon Rondeau, Greg Losada, Gustavo Rubio, Tim Grahl, Emilie Grahl, Adam Hines and Olga Sanchez. This work was funded by the South Florida Water Management District and Everglades National Park. ENP water level data were made available by ENP (South Florida Natural Resources Center (SFNRC) DataForEVER Dataset, Everglades National Park, Homestead, FL, Generated by D. Rondeau using Appaserver software (http://www.appaserver.com), Sacramento, CA, Public URL not currently available, please send data requests to EVER_data_request@nps.gov). We would like to further acknowledge the field assistance of numerous Florida Coastal Everglades LTER collaborators. This is SERC contribution # 625.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Troxler, T.G., Childers, D.L. & Madden, C.J. Drivers of Decadal-Scale Change in Southern Everglades Wetland Macrophyte Communities of the Coastal Ecotone. Wetlands 34 (Suppl 1), 81–90 (2014). https://doi.org/10.1007/s13157-013-0446-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-013-0446-5