
Abstract
Bird monitoring is frequent in wetlands; however, in the absence of information on other variables, trends in bird numbers are difficult to interpret. In this article we describe a methodology for bird’s habitat area assessment based on remote sensing. We calibrated the methodology to study the changes in the habitat area of Cygnus melancoryphus, the black-necked swan, at the Carlos Anwandter Sanctuary, a wetland located in Valdivia, Southern Chile. Swan habitat area was estimated by means of the Normalized Difference Vegetation Index (NDVI) based on Landsat images and calibrated through spectral photography by means of a portable Tetracam ADC Camera. Results show that calibrated NDVI values from Landsat images can be used to estimate habitat area but not to separate individual species of vegetation. We also show that the joint analysis of habitat area and swan count can indeed be used to separate some of the scales of variability of bird counts: those with “local-influence” associated with changes in habitat area from those of larger scales not related with habitat area.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Wetlands are considered havens for wildlife and sources of many ecosystem services; yet they are also under increasing threat (Fernández-Prieto and Finlayson 2009; Brinson and Eckles 2011; Fariña and Camaño 2012). Main threats are watershed modifications by agriculture, forestry and climate change (Marín et al. 2009; De Steven and Lowrance 2011; Marquet et al. 2012). In the year 2003 the European Space Agency and the Ramsar Convention Secretariat launched the GlobWetland project (http://www.globwetland.org) with the purpose of developing and demonstrating remote sensing technology to support wetland management. The project is currently in its second phase (GlobWetland II) aiming to the set up of a Global Wetlands Observing System. Indeed, remote sensing is likely to play an increasingly important role in wetland monitoring (Euliss Jr. et al. 2011).
Although medium-resolution sensors, such as Landsat TM (pixel size = 30 × 30 m) has historically being the most used platform for the remote sensing of the biosphere, small wetlands require higher resolutions (Klemas 2011). However, for large wetlands (e.g. 45 km2 and larger) Landsat images are still a good working solution, especially given its nearly 30 years of temporal coverage (http://glovis.usgs.gov).
The río Cruces wetland in southern Chile is an emblematic ecosystem, one of the largest wetland ecosystems in Chile (Marquet et al. 2012) hosting a large variety of bird species, including the highly noticeable black-necked swan, Cygnus melancoryphus (Corti and Schlatter 2002; AvesdeChile 2012). During the year 2004 the local swan population decreased dramatically from several thousand individuals to less than 500, mostly by emigration (Lopetegui et al. 2007; Delgado et al. 2009). Although most hypotheses pointed to the quasi-disappearance of its main food source, the invasive macrophyte Egeria densa (Hauenstein 2004; Ramírez et al. 2006; Lopetegui et al. 2007; Lagos et al. 2008; Marín et al. 2009), there has been no analysis of the temporal changes in macrophyte coverage within the wetland. The only pieces of information are the article by San Martín-Padovani et al. (2000) stating that in the spring/summer 1995/96 the area covered by E. densa was 23 km2 and the qualitative description by Jaramillo (2005) of its near-absence during November-December 2004. Indeed, Jaramillo (2005) when referring to E. densa abundance, previous to its disappearance in late 2004, only cites San Martín-Padovani et al. (2000). The Chilean Government conducts a monthly bird-monitoring program at the río Cruces wetland, especially after the 2004/05 event (CONAF 2012a, b). However, they have not considered other variables that may help explaining the trends of their bird counts. One improvement, used in other ecosystems, is to estimate bird’s habitat area by means of remote sensing (Connor and Gabor 2006; Guadagnin et al. 2009; Fleskes and Gregory 2010). However, the relationship between vegetation structure and satellite data may be contingent (sensu Schmitz 2010) to each ecosystem, requiring calibration and target analysis so results can be locally useful for monitoring and management purposes (Zomer et al. 2009). In this article we show a remote sensing methodology to monitoring black-necked swan habitat area at the río Cruces wetland in Southern Chile that incorporates those elements.
Methods
Study Site
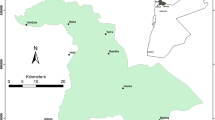
The Carlos Anwandter Sanctuary (area = 48.8 km2) is a wetland of international importance under the Ramsar Convention (Ramsar 2012), located in southern Chile (39°41′S 073°11′W, Fig. 1). Weather in the region is rainy-temperate, with dry, warm, summers (December through February), cold and rainy winters (June through August) and an annual mean precipitation of 1,791 mm (Marín et al. 2009). The Sanctuary is part of the río Cruces wetland, formed around the Cruces River by a 2 m tectonic subsidence due to one of the largest earthquakes recorded in recent history (9.6 on the Richter Scale on May 1960). Consequently, the depth distribution of the Sanctuary is one of a 4–6 m river surrounded by 2 m floodplains (Fig. 1). Egeria densa, Potamogeton Lucens and other macrophytes used to dominate floodplain’s vegetation also with an important presence of rushes of Scirpus californicus; the latter used as nesting area by birds (San Martín-Padovani et al. 2000). Thus, considering that macrophytes, especially E. densa, makes the bulk of the diet of black-necked swans (Erdman 2012) and rushes are nesting grounds, we studied their distribution and temporal changes in abundance by means of satellite images.

Geographic location of the Carlos Anwandter Sanctuary in the Chilean coast. The shaded areas in the image with geographic coordinates correspond to the wetland floodplains. The names in the figure correspond to the rivers draining the waters to the Sanctuary
Satellite Images: Acquisition and Pre-Processing
The objective of analyzing the habitat area of black-necked swans through satellite images was to gather the longest possible time-series, in the absence of direct measurements, in order to study its temporal fluctuations. The only archive of such images for the study area is available at the Earth Resources Observation and Science (EROS) Center belonging to the United States Geological Survey (USGS 2012). We searched the combined Landsat archive by means of the USGS Global Visualization Viewer (http://glovis.usgs.gov/) for the available time period (1984–2012) using the Path/Row 233/88 of the World Reference System 2 (WRS-2), with a central point at 40° 18′S, 72° 48′W. A total of 59 cloudless images were initially downloaded, with a considerable ten-year time gap between 1988 and 1998. However, after closer examination of the Sanctuary area, 12 were rejected due to excess haze leaving a total of 47 images (8 for the period 1985–1988 and 39 for the period 1998–2012).
Image’s pre-processing was done using the software ENVI 4.8. All images were first calibrated using the “Landsat Calibration” utility that converts Landsat digital numbers into exoatmospheric reflectance. On May 31st 2003 the Landsat 7 ETM sensor had a failure of the Scan Line Corrector. Since that time all Landsat 7 images have had wedge-shaped gaps on both sides of each scene, resulting in a 22 % data loss. Thus, we used the technique proposed by Scaramuzza et al. (2004) to fill the gaps, by means of the “Landsat Gapfill” utility, in all Landsat7 images after May 2003 using local histogram matching. Subsequently, we corrected all images from atmospheric effects using the Quick Atmospheric Correction (QUAC) routine, in order to generate surface reflectance values. Finally, we resized all images, both spatially and spectrally, by means of the Regions of interest (ROIs) tool. Spatially, we isolated all wetland pixels, changing the values of other areas (e. g. land, urban areas) to zero in all bands. Spectrally, we selected bands 2, 3 and 4 in order to fit those available from the Tetracam ADC Camera (see below), which correspond to the bands used to study vegetation.
Analysis of Target Species by Spectral Photography
We took, during November 2011 and January 2012, 29 spectral photographs of the main target species within the Sanctuary (E. densa [14], Potamogeton spp. [6] and Scirpus californicus [9]) by means of a portable Tetracam ADC camera. All photographs were taken from a small boat nearly on top of macrophyte patches and next to rushes of Sc. californicus. We also collected spectral information of water without any vegetation from all photographs.
The ADC camera has green, red and near-infrared (NIR) sensitivity with bands approximately equal to Landsat Thematic Mapper 2 (520–600 nm), Thematic Mapper 3 (630–690 nm) and Thematic Mapper 4 (760–900 nm) bands (Huang et al. 2010). All images were initially analyzed using the software PixelWrench2 provided with the camera. In order to calibrate images, we photographed a Teflon calibration tag during each session. Images of the calibration tag are used to teach the software about the spectral balance of sunlight in each photographic session. Calibrated images were exported as TIFF to be analyzed using the ENVI 4.8 software.
We obtained reflectance samples, from all spectral photographs, of the three bands from each target in order to analyze their spectral signatures. This procedure generated 71 samples for water, 364 for Sc. Californus, 295 for E. densa and 336 for Potamogeton spp. A preliminary analysis showed that spectral signatures did not differ significantly enough among species to be able to identify them using supervised classification techniques. However, reflectance in the near-infrared band differed significantly (p < 0.05) between vegetation targets and clear, no vegetation, water (Fig. 2). The literature (San Martín-Padovani et al. 2000) shows that the main components of the aquatic vegetation within the Sanctuary are Egeria densa (estimated area during spring/summer 1995/96 = 23 km2) and Scirpus californicus (6.7 km2). Thus, 60.9 % of the Sanctuary area used to be covered with vegetation that corresponded either to food or nesting grounds for black-necked swans. We decided, based on this information, to use the “Sanctuary’s vegetated area” as a compounded (nesting areas + food) proxy variable to quantify swan’s habitat area. Consequently, we calculated the Normalized Difference Vegetation Index (NDVI) for each target using the formula:

Corrected reflectance values for the near-infrared band from Tetracam ADC camera photographs for the studied targets: Egeria densa, Potamogeton spp., Sc. californicus and clear water
Where NIR and red correspond to two of the three bands available from ADC camera photographs (see above). Results of this analysis showed that indeed vegetation targets produced similar NDVI values, with a minimum of 0.1 and a maximum of 0.83, but significantly different (p < < 0.001) from water with no vegetation (Fig. 3). We restricted further analysis of NDVI values to the range 0.1 through 0.65; the first representing the low NDVI value for Potamogeton spp. and latter being one standard deviation (0.15) above the mean NDVI value (0.5) for E. densa.

Values for the Normalized Difference Vegetation Index (NDVI) based on Tetracam ADC Camera photographs for the studied targets: Egeria densa, Potamogeton spp., Sc. californicus and clear water
Landsat Images Processing: GIS Methods
We calculated, by means of ENVI 4.8, NDVI values for all corrected Landsat images. NDVI images were subsequently exported as ESRI grids to analyze them under ArcGIS 9.3. NDVI grids were resized, using a grid mask, to the Sanctuary area also selecting cells fulfilling the condition: 0.1 ≤ NDVI ≤ 0.65, based on spectral photography results (see above). Values for other cells were set to NO DATA. We then used those NDVI-Sanctuary grids to calculate the habitat area of the black-necked swan. Figure 4 shows a graphical example of the results for January 1987 (habitat area = 29.03 km2) and January 2004 (habitat area = 3.16 km2).

Examples of the habitat area (black areas) calculation based on Landsat images. The comparison shows a year with the highest area (January 1987) and a year with the smallest area (January 2004)
Ancillary Swan’s Data: CONAF’s Monitoring Program
The National (Chilean) Forestry Corporation (CONAF 2012a) is an organization governed by private law belonging to the Chilean Ministry of Agriculture. Its objective is to: “contribute to the conservation, increase, management and utilization of country’s forestry resources”. CONAF’s organization includes a Department of Wild Protected Areas, in charge of administering protected areas and nature sanctuaries such as the “Carlos Anwandter Nature Sanctuary”. One of CONAF activities in relation to the Sanctuary is a monthly bird census, since 1982 for swans. The census is carried out by boat and includes observations from 10 stations (towers) located inside the wetland. Results are freely available at CONAF’s web page (CONAF 2012b). We downloaded swan abundance data, with the exception of 1985 and 1986 that were not available, analyzing only those records corresponding to the months and years we had Landsat images in order to maintain a similar perspective on both datasets (i.e. habitat area and swan abundance). We further conducted a swan survey from a boat during our photographic sampling on November 23rd, 2011 to compare it with CONAF’s data. Total swan count (632) was near the mean value (635) of CONAF counts for November (753) and December 2011 (516), (CONAF 2012b).
Results
Temporal and Spatial Changes in Swan’s Habitat
Swan’s habitat showed a dramatic decrease from a maximum area (29.03 km2) during January 1987 to a minimum (0.53 km2) during August 2004 (Fig. 5). Afterwards, there was an increase up to 8.36 km2 during January 2012. The maximum, 1987, area is close to the value (29.7 km2) estimated by San Martín-Padovani et al. (2000) during spring/summer 1995/96.

Interannual changes in habitat area (black dots) and swan abundance (white dots). Lines correspond to two-point running means for habitat area (continuous line) and swan abundance (dashed line)
If habitat area is arranged by month, independent of year, results show a seasonal signal with high values during the spring/summer season (November–February; Fig. 6). When this signal is removed, analyzing only spring/summer values, results showed an interannual decreasing trend in habitat area since 1987 (Fig. 7). Still, if we include San Martín-Padovani et al. (2000) value, it is likely that the decreasing trend may have indeed started a decade later. Thus, we also analyzed spring/summer values from 1998 forward (Fig. 5) and found a significant (p = 0.002) linear decreasing trend but with a lower coefficient of determination (r 2 = 0.32).

Monthly distribution of swan habitat area

Interannual changes in habitat area based for spring-summer season values. The line corresponds to the result of a linear regression analysis
We analyzed spatial changes in habitat area dividing the Sanctuary in three subareas, similar to those used by Lagos et al. (2008): south, central and north (Fig. 8). The south sector is the closest to urban areas (e.g. city of Valdivia and Punucapa); the central sector receives the input from two rivers that drain the cattle-feeding prairies of the watershed and the north sector corresponds to the main river input (the Cruces River; Fig. 1). In order to decrease the seasonal effect, we analyzed the same month (January) in all years assuming that the area for January 1987 corresponded to 100 %. We also have included the data from San Martín-Padovani et al. (2000) assuming that the spatial distribution was the same as that from 1987 (Fig. 8). The first decrease in habitat area, down to 26.9 %, appeared in the south sector during January 1999; but by January 2000 the south and central sectors showed areal values lower than 13 %, while the north sector was at 32.3 % of the 1987 area. Later, during 2002 the south and central sectors showed some increase in area but only to a 31 % of 1987, while the northern sector declined to 17.6 %. During 2004 south and central sectors showed the lowest percentages of habitat (<7.5 %) with the north sector remaining in values similar to 2002. Habitat area recovery started slowly during 2011 for the south sector and continued to January 2012 reaching 47.5 % of 1987. The north sector also showed an increase in habitat area (34.5 %) while the central sector has remained below 20 % of 1987 area. This recovery of swan’s habitat during 2012 can indeed be supported by a different, indirect, type of data. The dominant presence of macrophytes in shallow water ecosystem such as wetlands, is commonly associated with the so-called “clear water regime” (Tironi 2012). CONAF (2012b), in all its monthly censuses since October 2010, has incorporated comments about water transparency (either clear or turbid) in a section under the title “Other observations made by the Rangers”. An analysis of those comments showed that during the year 2011, 6 out of 10 (60 %) censuses stated that water conditions were clear, increasing to 100 % (5 out of 5 comments) for 2012 (February–June). As a comparison, during the year 2010 although only 5 censuses registered water transparency, 80 % of them corresponded to turbid waters. Thus, the spatial analysis showed that changes in habitat area of the central sector has been the slowest both during reduction as in recovery but, at the same time, has experienced the highest reductions during the 2004–2011period. The relationship between these temporal/spatial changes in habitat and the abundance of swans is analyzed in the next section.

Changes in the spatial distribution of habitat area based on January values. The upper figure shows the divisions of the Sanctuary corresponding to the data in the lower figure
Relationship Between Changes in Habitat Area and Swan’s Abundance
Swan’s abundance in the Carlos Anwandter Sanctuary has changed from 1,500 individuals in the 1980s to 6000–8000 between 1999 and 2004, decreasing afterward to less than a thousand between 2005 and 2011 to finally increase to 1,800 during January 2012 (Fig. 5). If indeed the “Sanctuary’s vegetated area” is a compounded (nesting areas + food) proxy variable for swan’s habitat, we should find a relationship with the changes in swan abundance. We analyzed such relationship in several ways. First, we generated smoother series calculating a two-point running mean for both habitat area and swan’s abundance (Fig. 5). Results from this analysis, especially from 1999 forward, showed that changes in swan abundance were preceded by changes in habitat area. Second, we calculated the cross-correlation between both variables using data from February 1998 forward in order to avoid problems with the ten-year gap between 1988 and 1998. Results showed that there is a positive, significant (p < 0.05), correlation at a lag of 3 to 6 periods (Fig. 9) between both variables (i.e. habitat area preceding swan’s abundance). However, since we did not have images for all months, this lag only shows that vegetation changes precede changes in swan abundance but we couldn’t calculate a precise time, in months, for such a lag. Finally, we plotted changes in habitat area and swan’s abundance as a trajectory in a phase-space plot for all spring/summer data (Fig. 10). This result showed that the large habitat areas for 1987 and 1996 indeed corresponded to dramatically different swan abundances, with considerable fluctuations between 1988 and 1999. Since some authors (e.g. Schlatter et al. 2002) have related the abundance of swans at río Cruces with the different phases of the El Niño Southern Oscillation (ENSO) climatic phenomenon, we searched for information regarding the climatic conditions for the years 1987 and 1996 (NOAA 2012; DGF 2012). Both sources agree that 1987 was a warm, El Niño, period while 1996 was a cold, La Niña, period (Fig. 10). After 1999 and before January 2005 both habitat area and swan’s abundance reduced the fluctuation range; between 5 km2 and 13 km2 for area and between 4400 and 6000 for swans. During 2004/2005 and up to 2011 both variables shifted to a rather reduced fluctuation range, with low swan abundances. Finally, during January 2012 both variables showed an increase approaching values close to those registered during November 1988 (Fig. 10).

Result of the cross-correlation analysis for the habitat area and swan abundance between February 1998 and January 2012. Red line corresponds to p < 0.05. Significant values corresponded to habitat area preceding swan abundance

Phase-space trajectories based on habitat area and swan abundance. The black line separates the condition before (Pre-2005) and after (Post-2005) the year 2005. Numbers inside the graph correspond to month/year for key changes in the trajectory
Spatially, our November 2011 swan survey showed that 58 % of the population was located in the central sector of the Sanctuary (Fig. 8) and 30 % in the south sector. Furthermore, an analysis of CONAF Ranger’s descriptions on swan’s distribution (i.e. locations where the largest concentration of swan were observed), available between January 2009 and January 2012 (CONAF 2012b) showed that the central sector was the one holding the largest part of the population. Thus, from a spatial perspective, the largest reduction of habitat area between 2004 and 2011 (Fig. 8) occurred in the sector where most of the swan population was located.
Discussion
Wetlands are critical habitats for a large variety of wildlife (Klemas 2011; RAMSAR 2012). Earth observation technology or remote sensing is vital for the study, monitoring and management of wetlands worldwide (Fernández-Prieto and Finlayson 2009; Adam et al. 2010; Huang et al. 2012). It can provide information on multiple time and spatial scales and on inaccessible habitats (Liu et al. 2010). The latter is especially important when analyzing the habitat area of waterbirds, one of the most important ecosystem services from wetlands (Brinson and Eckles 2011).
Cygnus melancoryphus, the black-necked swan, is a bird with an extremely large, native, geographic range in South America; including Argentina, Brazil, Chile, Falkland Islands (Malvinas) and Uruguay. The IUCN in its “Red list of threatened species” (IUCN 2012), classifies it in the “least concern” category, due to its large range and its stable population trend. In Chile, its range extends between 30°S (Coquimbo) down to Cape Horns (55°S), nearly 2800 km, inhabiting lakes, wetlands and river areas (AvesdeChile 2012). Still, at a local scale (e.g. the Carlos Anwandter Sanctuary in Valdivia; Fig. 1) the sudden fluctuations of its population (from 8000 in April 2004 to 249 in August 2005; Source: CONAF 2012b) generated public concern (Jaramillo 2005; Lopetegui et al. 2007; Lagos et al. 2008; Marín and Delgado in press), increasing the need for information about the ecological state of the Sanctuary. However, one of the problems of concentrating exclusively on one ecosystem component, birds, especially one with such a long range as the black-necked swan, is that it is almost impossible to disentangle effects from different scales based on bird counts alone. That is, changes in abundance may either be due to local effects (e.g. decrease in food supply) or to larger scale phenomena, such as ENSO (Schlatter et al. 2002) or other transient climatic events (Marín et al. 2009). The alternative, from a local point of view, is to incorporate additional information that may serve to separate, or at least to suggest, effects from different scales. Other strategy, such as simultaneous bird counts in several wetlands covering the species range, is challenging since for the case of the black-necked swan it crosses over at least four different countries. One alternative, based on ecological theory and practice (Connor and Gabor 2006; Guadagnin et al. 2009; Fleskes and Gregory 2010), is to estimate the area of swan’s habitat, by means of remote sensing, in order to analyze its trends along those of the swan population. In this article we have shown that it is possible to estimate the habitat area of the black-necked swan using corrected Landsat TM images by means of the Normalized Difference Vegetation Index (NDVI). Although Landsat TM is a medium-resolution sensor (pixel size = 30 × 30 m), the size of the Sanctuary (48.8 km2) allows enough elements (54000) to be analyzed in each image. This result agrees with that from Jakubauskas et al. (2000) showing a good relationship between macrophyte cover (Nuphar polysepalum) and NDVI values in Swan Lake (Wyoming, USA) and those of Behera et al. (2012) that used Landsat images to monitor wetlands in India. However, the use of remote sensing technology for monitoring wetland vegetation requires a careful analysis of the target species through, for example, spectral photography. In our case, we selected, due to several considerations including cost and portability in small boats, the Tetracam ADC camera. The analysis of the main target species showed, in agreement with the work of Silva et al. (2008) and Zomer et al. (2009), that when dealing with wetland vegetation Landsat images are useful for mapping macrophytes communities but not necessarily individual species. We found that the analyzed species (Egeria densa, Potamogeton spp. and Scirpus californicus) generate similar spectral signatures when using Landsat TM bands 2, 3 and 4 but with a significant difference from clear water (Figs. 2 and 3). Other researchers, using sensors with higher spatial and spectral resolutions have indeed reported specie-specific spectral signatures (e.g. Zomer et al. 2009). Nevertheless, using specie-specific spectral libraries or an index (e.g. NDVI) to estimate the area of a community will depend upon the objective of the work and the available resources. In our case, considerations also included having a long-enough record of images in order to use it to disentangle the various scales included in the temporal changes of the black-necked swan at the Carlos Anwandter Sanctuary.
Our results of the trends of habitat area (Figs. 5, 7 and 8) and of swan abundance (Figs. 5, 9 and 10) show that: (1) habitat area has an interannual decreasing, linear, trend since at least 1996 (Fig. 7), superimposed to (2) smaller scale fluctuations which generated a very low area during August 2004 (Figs. 5 and 8). On the other hand, swan abundance show large fluctuations: (1) related with changes in habitat (Figs. 5 and 9) and (2) to larger scale processes (Fig. 10). Thus, using a mixture of direct bird counts and habitat area estimation through remote sensing it is possible to have a suit of hypotheses that may serve to guide both monitoring and management actions. The methodology is based on the idea that the “local-influence” is to be accepted as the primary, null, hypothesis and its failure to explain trends in bird’s abundance should lead into considering larger scales as possible explanations. For example, it is clear from our analysis that changes in swan abundance between 1999 and 2012 falls into the local-influence hypothesis with changes in habitat area preceding changes in bird abundance (Figs. 5 and 9). However the local explanation fails in the case of 1987 and 1996 (Fig. 10). Indeed, the habitat area for 1987 and 1996 were almost the same (Fig. 10) yet swans were 5 times more abundant in the latter year. Schlatter et al. (2002) and Vilina et al. (2002) have related local changes in swan abundance to the climatological phenomenon known as El Niño Southern Oscillation. In one case (río Cruces wetland, Schlatter et al. 2002) they propose that swans are more abundant during cold, La Niña, years (e.g. 1996). In the other, (El Yali wetland, located 640 km north of río Cruces; Vilina et al. 2002) they propose swans are more abundant during warm, El Niño, years (e.g. 1987). Thus, the proposed explanation for the 1987–1996 difference is in this case related to the large migratory capacity of swans; migrating north (El Yali and other northern wetland systems) during warm years and south (río Cruces) during cold, dry, years. Such a hypothesis will of course require further analysis; however, the incorporation of data on habitat area enables it for a follow-up in terms both of monitoring and eventually for management purposes.
Ecosystems can be defined as medium-number open systems (Jørgensen 1992). That is, their components cannot simply be averaged out in order to simplify its analysis and, at the same time, they cannot be analyzed in its entirety. Thus, there will always be difficult decisions to make when choosing what components should be monitored, with public pressure exerting a powerful influence (Marín and Delgado in press). The black-necked swan population of the Carlos Anwandter Sanctuary in southern Chile is one of those examples. We propose that in such cases, science should play an important role by proposing additional, cost-efficient, variables that may help disentangling the many scales affecting the temporal fluctuations of ecosystem variables. In this context, the coupling of remote sensing-GIS technologies and local sampling could indeed go a long way toward achieving the goal of more comprehensive monitoring.
References
Adam E, Mutanga O, Ruege D (2010) Multiespectral and hyperespectrl remote sensing for identification and mapping of wetland vegetation: a reiew. Wetlands Ecology and Management 18:281–296
AvesdeChile (2012) Aves de Chile. Cisne de Cuello Negro. http://www.avesdechile.cl/050.htm. Accessed 15 August 2012
Behera MD, Chitale VS, Shaw A, Roy PS, Murthy MSR (2012) Wetland monitoring, serving as an index of land use change- A study in Samasputt wetlands, Uttar Pradesh, India. Journal of the Indian Society of Remote Sensing 40:287–297
Brinson MM, Eckles SD (2011) U.S. Department of Agriculture conservation program and practice effects on wetland ecosystem services: a synthesis. Ecological Applications 21:S116–S127
CONAF (2012a) Corporación Nacional Forestal de Chile. http://www.conaf.cl. Accessed 2 August 2012
CONAF (2012b) Plan Gestión Ambiental Río Cruces. http://www.conaf.cl/parques/seccion-plan-gestion-ambiental-rio-cruces.html. Accessed 8 July 2012
Connor KJ, Gabor S (2006) Breeding waterbird wetland availability and response to water-level management in Saint John River floodplain wetlands, New Brunswick. Hydrobiologia 567:169–181
Corti P, Schlatter RP (2002) Feeding ecology of the black-necked swan Cygnus melancoryphus in two wetlands of Southern Chile. Studies on Neotropical Fauna and Environment 37:9–14
De Steven D, Lowrance R (2011) Agricultural conservation practices and wetland ecosystem services in the wetland-rich Piedmont-Coastal Plain region. Ecological Applications 21:S3–S17
Delgado LE, Marín VH, Bachmann PL, Torres-Gomez M (2009) Conceptual models for ecosystem management through the participation of local social actors: the Río Cruces wetland conflict. Ecology and Society 14 (1): 50. http://www.ecologyandsociety.org/vol14/iss1/art50
DGF (2012) Boletín climático. Departamento de Geofísica, FCFM, Universidad de Chile. http://met.dgf.uchile.cl/clima/. Accessed 8 July 2012
Erdman L (2012) Black necked swan website. http://bioweb.uwlax.edu/bio203/s2009/erdman_laur/#. Accessed 10 July 2012
Euliss NH Jr, Smith LM, Liu S, Duffy WG, Faulkner SP, Gleason RA, Eckles SD (2011) Integrating estimates of ecosystem services from conservation programs and practices into models for decision makers. Ecological Applications 21:S128–S134
Fariña JM, Camaño A (2012) Humedales costeros de Chile. Aportes científicos a su gestión sustentable. Ediciones Universidad Católica de Chile, Santiago de Chile
Fernández-Prieto D, Finlayson CM (2009) Earth observation and wetlands. Journal of Environmental Management 90:2119–2120
Fleskes JP, Gregory CJ (2010) Distribution and dynamics of waterbird habitat during spring in southern Oregon-Northeastern California. Western North American Naturalist 70:26–38
Guadagnin DL, Maltchik L, Fonseca CR (2009) Species-area relationship of Neotropical waterbird assemblages in remnant wetlands: looking at the mechanisms. Diversity and Distributions 15:319–327
Hauenstein E (2004) Antecedentes sobre Egeria densa (Luchecillo), hidrófita importante en la alimentación del cisne de cuello negro. Gestión Ambiental 10:89–95
Huang Y, Thomson SJ, Lan Y, Mass SJ (2010) Multispectral imaging systems for airborne remote sensing to support agricultural production management. Int J Agric & Biol Eng 3:50–62
Huang LB, Bai J, Yan DH, Chen B, Xiao R, Gao HF (2012) Changes of wetland landscape patterns in Dadu river catchment from 1985 to 2000, China. Frontiers of Earth Science 6:237–249
IUCN (2012) The IUCN Red list of threatened species. 2012.1 http://www.iucnredlist.org/. Accessed 10 August 2012
Jakubauskas M, Kindscher K, Fraser A, Debinski D, Price KP (2000) Close-range remote sensing of aquatic macrophyte vegetation cover. Int J Remote Sensing 21:3533–3538
Jaramillo E (2005) Final report: “Study about the origin of mortalities and population reduction of waterbirds in the Nature Sanctuary Carlos Anwandter, at the Valdivia Province.” Complementary Agreement Number 1210-1203/2004-12-14. National Environmental Agency (CONAMA) – Austral University of Chile. http://www.sinia.cl/1292/articles-31665_segundoinformeCisnes.pdf. Accessed 8 July 2012
Jørgensen SE (1992) Integration of ecosystem theories: a pattern. Kluwer Academic Publishers, London
Klemas V (2011) Remote sensing of wetlands: case studies comparing practical techniques. Journal of Coastal Research 27:418–427
Lagos NA, Paolini P, Jaramillo E, Lovengreen C, Duarte C, Contreras H (2008) Environmental processes, water quality degradation, and decline of waterbird populations in the río Cruces wetland, Chile. Wetlands 28:938–950
Liu C, Jiang H, Hou Y, Zhang S, Su L, Li X, Pan X, Wen Z (2010) Habitat changes for breeding waterbirds in Yancheng National Nature Reserve, China: A remote sensing study. Wetlands 30:879–888
Lopetegui EJ, Vollman RS, Cifuentes HC, Valenzuela CD, Herbach EP, Huepe JU, Jaramillo GV, Leischner BP, Riveros RS (2007) Emigration and mortality of black-necked swans (Cygnus melancoryphus) and disappearance of the macrophyte Egeria densa in a Ramsar wetland site of Southern Chile. Ambio 36:607–609
Marín VH, Delgado LE (in press) From ecology to society and back: the (in)convenient hypothesis syndrome. International Journal of Sustainable Development
Marin VH, Tironi A, Delgado LE, Contreras M, Novoa F, Torres-Gómez M, Garreaud R, Vila I, Serey I (2009) On the sudden disappearance of Egeria densa from a Ramsar wetland site of Southern Chile: A climatic event trigger model. Ecological Modelling 220:1752–1763
Marquet PA, Abades S, Barría I (2012) Distribución y conservación de humedales costeros: una perspectiva geográfica. In: Fariña JM, Camaño A (eds) Humedales costeros de Chile. Aportes científicos a su gestión sustentable. Ediciones Universidad Católica de Chile, Santiago de Chile, pp 2–19
NOAA (2012) Changes to the oceanic Niño Index. National Weather Service, Climate Prediction Center. http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ensoyears.shtml.. Accessed 8 July 2012
Ramírez C, Carrasco E, Mariani S, Palacios N (2006) La desaparición del Luchecillo (Egeria densa) del Santuario del Río Cruces (Valdivia, Chile): una hipótesis plausible. Ciencia & Trabajo 20:79–86
RAMSAR (2012) The Ramsar Convention on Wetlands. http://www.ramsar.org/. Accessed 8 July 2012
San Martín-Padovani C, Contreras-Fernández D, Ramírez-García C (2000) El recurso vegetal del Santuario de la naturaleza “Carlos Andwanter” (Valdivia, Chile). Revista Geográfica de Valparaíso 31:225–235
Scaramuzza P, Micijevic E, Chander G (2004) SLC gap-filled products. Phase one methodology. United States Geological Survey, U.S. Department of Interior. http://landsat.usgs.gov/documents/SLC_Gap_Fill_Methodology.pdf. Accessed 10 April 2012
Schlatter RP, Navarro RA, Corti P (2002) Effects of El Nino southern oscillation on numbers of black-necked swans at Rio Cruces Sanctuary, Chile. Waterbirds 25(Sup. 1):114–122
Schmitz OJ (2010) Resolving ecosystem complexity. Monographs in Population Biology 47. Princenton University Press, Oxford
Silva TSF, Costa MPF, Melack JM, Novo EMLM (2008) Remote sensing of aquatic vegetation: theory and applications. Environmental Monitoring and Assessment 140:131–145
Tironi A (2012) Propuesta teórica para el análisis topológico de redes ecológicas: en la búsqueda de la resiliencia ecosistémica. Tesis para optar al grado académico de Doctor en Ciencias con mención en Ecología y Biología Evolutiva. Facultad de Ciencias, Universidad de Chile. ftp://ecosistemas.uchile.cl/pub/publicaciones/2012/Tesis_Doctoral_A_Tironi.pdf. Accessed 8 July 2012
USGS (2012) Earth Resources Observation and Science (EROS) Center. United States Geological Survey, U.S. Department of Interior. http://eros.usgs.gov. Accessed 8 July 2012
Vilina YA, Cofre HL, Silva-García C, Cargía MD, Perez-Friedenthal C (2002) Effects of El Niño on abundance and breeding of Black-necked Swans on El Yali wetland in Chile. Waterbirds 25(Sup. 1):123–127
Zomer RJ, Trabucco A, Usin SL (2009) Building spectral libraries for wetlands land cover classification and hyperspectral remote sensing. Journal of Environmental Management 90:2170–2177
Acknowledgments
This research was funded by a grant from FONDECYT-Chile (N° 1110077). We thank the help of Carlos Ibañez and Antonio Tironi with bird counts. We also thank Captain Carlos Cisterna and the crew of the “Catalina” for the logistic support during fieldwork.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Delgado, L.E., Marín, V.H. Interannual Changes in the Habitat Area of the Black-Necked Swan, Cygnus melancoryphus, in the Carlos Anwandter Sanctuary, Southern Chile: A Remote Sensing Approach. Wetlands 33, 91–99 (2013). https://doi.org/10.1007/s13157-012-0354-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-012-0354-0