
Abstract
We used tree-ring analysis to evaluate the combined effects of rising water levels and 13 years of municipal wastewater addition on growth of baldcypress in a subsiding swamp forest in southern Louisiana. Trees in the treatment, downstream outflow, and adjacent untreated control areas all experienced increased growth coinciding with a period of widespread rapid subsidence and water-level increases in the late 1960s. Tree growth in the treatment and outflow sites began to decrease before wastewater application began in 1992, and afterward was apparently unaffected by treatment. In contrast, trees in the control site have not experienced growth declines. Hydrological changes caused by subsidence have apparently overwhelmed any effect of wastewater treatment on baldcypress growth. Increasing inundation may have increased growth initially by eliminating competition from species less tolerant of inundation; however, after a decade of sustained higher water, growth declined steadily. Release of baldcypress from competition continues at the topographically higher control site, but growth will likely subsequently decrease as ongoing subsidence and eustatic sea-level rise cause more prolonged inundation. We conclude that short-term increases in water level stimulated growth of baldcypress, but long-term increased inundation was a net stressor and was more important than nutrient limitations in controlling growth at this site.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Tertiary treatment of municipal wastewater has been increasingly used as an inexpensive and effective way of simultaneously improving effluent water quality and increasing productivity of wetland vegetation (Day et al. 2004). Application of municipal wastewater has been used as a treatment and restoration practice in cypress (Taxodium spp.)-tupelo (Nyssa spp.) swamps, with the typical result being enhanced growth rates by increasing nutrients in otherwise dystrophic swamps (Mitsch and Ewel 1979; Brown 1981; Nessel et al. 1982; Hesse et al. 1998; Hunter et al. 2009). However, experience with this practice remains limited.
One place where productivity of cypress-tupelo swamps has been degraded is the delta of the Mississippi River, where swamp forests dominated by baldcypress (Taxodium distichum (L.) Rich) and water tupelo (Nyssa aquatica L.) occupy about 3450 km2 (Keim et al. 2006). Hydrologic engineering throughout the delta has eliminated riverine flooding and distributary flows by an extensive network of artificial levees and other water control structures. The result has been that consolidation of recent alluvium has not been balanced by continued deposition from overbank flooding and in situ organic matter production, and rapid subsidence has occurred nearly throughout the delta (Penland and Ramsey 1990) resulting in increased stagnant inundation, low nutrient mobility, accumulated phytotoxins, low dissolved oxygen, and high rates of wetland loss (Day et al. 2007).
Baldcypress is a species typical of riverine swamps with extended but not permanent flooding. In contrast to periodic flooding common in floodplains, stagnant and semipermanent flooding causes reduced growth, crown dieback, increased susceptibility to pests, decreased root mass, greater susceptibility to blow down during hurricanes, and increased tree mortality (Conner et al. 1981; Keeland et al. 1997; Amos 2006; Shaffer et al. 2009). In the delta of the Mississippi River, multiple stressors are often present (Allen et al. 1996; Effler and Goyer 2006). Often, the stressor most responsible for degradation is not clear, so the effect of implementing restoration measures such as municipal wastewater application is uncertain.
The objective of this research was to evaluate the combined effects of municipal wastewater treatment and rising water levels on growth of a swamp forest in southern Louisiana. We used tree rings to evaluate previous growth rates of baldcypress at a site where wastewater additions began 13 years prior to sampling. This site has been the subject of intensive research since its establishment. Previous work has quantified nutrient dynamics (Zhang et al. 2000), denitrification (Crozier et al. 1996; Boustany et al. 1997), floating marsh establishment (Izdepski et al. 2009), and organic and inorganic accretion (Rybczyk et al. 1998, 2002) at the site. Short-term measurements of litter production have suggested that wastewater addition has not increased tree productivity at this site (Rybczyk et al. 1996). Elsewhere in the same interdistributary basin, dendrochronology has revealed that baldcypress are generally declining in response to increased long-term inundation (Amos 2006), but Hesse et al. (1998) reported increased diameter growth in response to wastewater additions in a well-drained swamp forest in south Louisiana. The specific objective of this work was therefore to test whether wastewater additions were sufficient to counteract expected declines in tree growth caused by increased inundation.
Methods
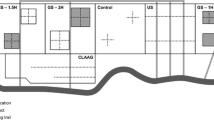
The study site (Fig. 1) is in Lafourche Parish, Louisiana, near Bayou Lafourche, which was an active distributary of the Mississippi River until it was disconnected for flood control in 1904. Bayou Lafourche carried the majority of Mississippi River flow about 1000 years ago, but this had declined to less than 20% of flow by the time of European colonization in the early 1700s. The site was isolated from upland runoff in the 1930s by the construction of the Terrebonne-Lafourche drainage canal. The spoil bank of the canal blocks any surface drainage into the study area.

Study location in Louisiana, USA. Stars indicate tree-ring collection sites. Effluent is released from a canal along the northern border of the treatment wetland, and flows down-gradient to the southwest. Flow is restricted to an area between a small, raised road and a low ridge formed by a former distributary channel
Discharge of treated effluent at the site began in 1992, and an average of 7500 m3 d−1 have been discharged since. Loading rates of N and P from the wastewater effluent to the treatment site average 3.1 g m−2 y−1 and 0.6 g m−2 y−1, respectively. Effluent is released from a canal along the northern border of the treatment wetland, and flows down-gradient to the southwest (Fig. 1). Flow is restricted to an area between a small, raised road and a low ridge formed by a former distributary channel. Wastewater drains out of the treatment area at an outflow site located about 2000 m at the down-gradient end of the swamp. There is substantial uptake of nitrogen and phosphorus in the forested wetlands. Nitrate, the main form of inorganic nitrogen, often exceeds concentrations of 10 mg l−1 in the effluent, but these are reduced to near zero at the outflow site (Zhang et al. 2000).
In addition to the treatment and outflow sites, there is an adjacent untreated site used as a control (Fig. 1). The control area is hydrologically separated from the treatment site by a low natural ridge in the swamp whose mean elevation is 1.16 m above mean sea level (MSL) and about 30 cm higher than the adjacent forested wetlands, which are at mean elevation of 0.76 m above MSL. The control site is about 20–25 cm higher than the treatment site.
In the treatment site, the most common tree species in addition to baldcypress are ashes (Fraxinus pennsylvanica Marsh. and F. profunda (Bush) Bush), most of which germinated earlier than the 1980s and are now mostly dead. In the outflow site, baldcypress is the most common species but others present include water tupelo, Drummond’s red maple (Acer rubrum var. drummondii (Hook. & Arn. ex Nutt.) Sarg.), and dwarf palmetto (Sabal minor (Jacq.) Pers.). In the control site, baldcypress is growing in a mix with black willow (Salix nigra Marsh), ashes, Nuttall oak (Q. texana Buckl.), and Drummond’s red maple; the site also has substantial understory regeneration of baldcypress. Aside from baldcypress, the ridge is vegetated primarily with oaks (Quercus nigra L. and Q. laurifolia Michx.), sweetgum (Liquidambar styraciflua L.), American elm (Ulmus americana L.), dwarf palmetto, and boxelder (Acer negundo L.).
In each site (Treatment, Outflow, and Control), 10–11 overstory baldcypress trees were sampled with a 5.15-mm Haglöf increment borer. This sample size is sufficient given the relatively high inter-series correlation and high sensitivity of the trees to environmental variation (Table 1, Fritts 1976). At least two (maximum four) cores were extracted from each tree. The cores were extracted at least 3 m above the ground to avoid the deformed rings in the buttressed base (Young et al. 1995; Keeland and Young 1997). Cores were dehydrated at 40°C for approximately 2 weeks, then glued onto wooden holders and sanded with progressively finer sandpaper (100–600 grit; 125–16 μm (Orvis and Grissino-Mayer 2002)) to prepare surfaces for reading.
All ring widths were measured to the nearest 0.01 mm using a Velmex sliding stage under a dissecting microscope. Cores were crossdated to assign exact calendar year to each ring using skeleton plots (Stokes and Smiley 1968) and marker rings of unusually good or poor growth years from established baldcypress chronologies from the area (Amos 2006). Crossdating was verified using the cross-dating software COFECHA (Holmes 1983; Grissino-Mayer 2001). False and locally missing rings are common in baldcypress (Ewel and Parendes 1984; Young et al. 1993), but the crossdating procedure sufficiently reduces errors in ring dating so that baldcypress is one of the most useful trees for dendrochronology in the region (Stahle et al. 1985, 1988; Stahle and Cleaveland 1992). Once all cores were crossdated, we averaged all measurements of ring width by year for each tree to obtain a single time series of mean ring width for each tree (as recommended by Speer 2010), then constructed chronologies from these mean measurements. Chronologies for all three sites were truncated to 1922–2005, which is the period over which sample size was sufficient.
Investigating competing wastewater treatment and inundation effects is complicated by the fact that there are multiple controlling factors on tree growth. Thus it was important to account for variance in growth not related to the treatments or inundation. We modeled ring widths modified from Cook (1987), following Amos (2006), as
where R is ring width in year t; A is the expected ring width based on the age-related decrease in width; C is the effect of annual variations in climate; H is the effect of variations in water level; and ε is error not accounted for in other terms, including stand disturbance.
Tree-ring series are autocorrelated because tree growth rates integrate response to climatic conditions for several years (Fritts 1976). Some environmental variables from Eq. 1 are also autocorrelated (i.e., H and some stand-disturbance elements of ε) but some are not (C); we used separate modeling approaches for each. For analysis of the effects of serially uncorrelated environmental variables (C) on growth, we used the ARSTAN model (Cook 1985; Cook and Holmes 1986) to create a tree ring chronology with no autocorrelation. In ARSTAN, each tree ring series is individually detrended by both (1) a model for combined A and long-timescale variance originating in H and ε and (2) an autoregressive-moving average (ARMA) model.
For analysis of the effects of serially correlated environmental variables (H and stand disturbance) on growth, we created a chronology by the Regional Curve Standardization (RCS) method (Briffa et al. 1992; Esper et al. 2003). The RCS method detrends tree ring time series by cambial age as a ratio of observed ring width to the average ring width for a large number of trees from the region. Data for the regional curve (RC) were 62 baldcypress from a study in the same region by Amos (2006) as well as all trees in this study, for a total of 93 trees. We formed the RC by fitting a negative exponential curve to these data. The RCS chronology preserved low-frequency variance and autocorrelation in the chronology.
To assess effects of annual climate variation and short-term water-level variation on growth, we used Pearson correlations between the chronologies and several climate and hydrological variables. Climate variation was examined for correlation with the ARSTAN growth index, using climatic data from Louisiana Region 9 of the National Oceanic and Atmospheric Administration National Climatic Data Center (NCDC) database. Available variables were monthly temperature, monthly precipitation, monthly Palmer’s Drought Severity Index (PDSI), and monthly Palmer’s Hydrological Drought Index (PHDI). The PDSI is designed to model soil moisture, and the PHDI is designed to model longer-term water balance in groundwater and larger water bodies. Positive values for both indices indicate wetter than average conditions. There are no long-term hydrologic data available in the immediate vicinity of the treatment site; however, water levels do not vary substantially spatially because this is a large estuarine swamp within which correlations among gauges are generally high (Amos 2006). Therefore, for analysis of hydrological data, we used water-level (stage) data from the nearest gauged site at Lake Verret (Attakapas Landing), which is 25.5 km west-northwest of the treatment site. The effects of short-term hydrologic variation on annual growth were analyzed using the RCS growth index.
As at many locations in the subsiding Mississippi River delta in southern Louisiana, the Attakapas Landing gauge has recorded an increase in water depth (due to subsidence and perhaps backwater flooding from the Atchafalaya River) since its establishment in 1955. Amos (2006) found that 10-year running average water levels were statistically significant predictors of long-term changes in baldcypress growth, so we compared the 10-year running average water levels to 10-year running average growth in the RCS chronology.
Results
Tree-Ring Chronologies
All three sites had mean inter-series correlation (mean of all correlations between individual-tree series and the stand-average series after detrending and correcting for autocorrelation by COFECHA) greater than 0.52, which indicates proper crossdating, and mean sensitivity (mean ratio of inter-ring difference to mean ring width for all pairs of rings in all trees) substantially greater than the threshold of 0.3 that indicates trees are highly sensitive to environmental variation (Fritts 1976; Grissino-Mayer 2001) (Table 1). Problem segments (50-year time spans during which an individual tree was correlated to the stand-average series at r < 0.3281) were less than 10% of all series at all sites, which also corroborates proper crossdating.
The date of the earliest ring at the height of coring ranged from 1835 to 1922, and 20 of the 31 trees first reached the height of coring between 1908 and 1919. Assuming it required 5–10 years for trees to grow to the 3 m height of coring, these dates indicate most trees at the site became established between about 1898 and 1914. For trees that predated the 1898–1914 cohort, the RCS chronology shows a slow decrease in growth through 1906 followed by a growth release (Fig. 2a); these trees averaged 9 cm diameter inside bark at 3 m height in 1899. Thus, it appears the site was logged about 1906, but small trees were left. This history is typical for cypress-tupelo swamps in southern Louisiana, which were nearly all logged from about 1890–1925 (Conner and Toliver 1990).

Tree ring chronologies (a and b) and sample sizes (c) from the wastewater effluent site (treatment), downstream outflow site, and adjacent control site. Tree-ring crossdating was only performed for the period 1922–2005. The vertical line indicates the onset of effluent addition to the treatment site in 1992
Growth Relationships with Treated Wastewater Additions
The 13 years following wastewater additions were the poorest growth of the previous half century in the treatment and outflow sites, while the control site grew at rates equal to or higher than any in the past 50 years (Fig. 3). It was difficult to ascribe these changes to the treatment, however, because post-treatment growth in the treatment and outflow sites followed a trend of declining growth that began at least a decade before treatment (Figs. 2a, 3).

Decadal means of tree ring chronologies; error bars indicate one standard deviation. The first four decades were prior to the onset of effluent additions in 1992
Growth Relationships with Climate and Inundation
Several monthly climatic and hydrological variables were significantly correlated with the ARSTAN (detrended, non-autocorrelated) tree-ring chronology (Fig. 4). Correlations were similar for all three stands (not shown), so that controls by climate and short-term hydrological variation on annual growth were not strongly affected by elevation-driven hydrological regime (deepest inundation at the lowest, outflow site and shallowest inundation at the highest, control site). Correlations were also similar for the treatment stand both before and after initiation of wastewater addition (Fig. 4).

Pearson correlations of ARSTAN tree ring chronologies (combined across all three sites) with monthly climatic and hydrologic variables. Months presented range from March of the year prior to ring formation through August of the year of ring formation. Large squares indicate correlations significant at p = 0.05, and small squares indicate correlations significant at p = 0.10. The pre-treatment period is 1922–1991, and the post-treatment period is 1993–2005
The greatest annual growth occurred in cool, wet growing seasons, particularly during the months of April and May (Fig. 4). High precipitation and low temperature during this period were individually correlated with greater growth, but their combination in the form of drought and hydrological indices (PDSI and PHDI) were more consistently significantly correlated with growth. Positive correlations with PDSI and PHDI persisted later into the summer in part because of temporal correlation in those variables.
The strongest correlations between annual tree growth and all short-term variables were with swamp water levels (stage); high water levels were positively correlated with growth beginning in the winter preceding the growing season and through the growing season of ring formation (Fig. 4). When analyzing the average growth response across all sites both before and after treatment, the month with greatest simple correlation with growth was May of the current growing season (r = 0.38), and the multiple-month period of greatest correlation was March-May of the current growing season (r = 0.40). Other variables highly correlated with all-site growth were maximum water level during the year of ring formation (r = 0.39) and mean stage throughout the growing season (r = 0.36).
In contrast with the positive correlation between growth and short-term water level, there was a negative correlation between growth and long-term water level. Growth rates at all three sites increased strongly (Fig. 5) in conjunction with a rapid increase in water level by about 20 cm in the early 1970s related to floods on the Atchafalaya River (which dominates the hydrology of the area, including Lake Verret, because of substantial backwater effects; Swarzenski 2003). However, after about the first decade of sustained higher water, the growth in the treatment and outflow sites declined steadily. This growth pattern follows a regional trend in the large area of wetlands west of the study area, in which three other low-elevation baldcypress stands surrounding Lake Verret also experienced short-term increases in growth followed by long-term decline (Amos 2006) (Fig. 5). Growth in the control site experienced the same increase following water-level increases, but unlike the other sites did not experience a subsequent decline. Because the control site is about 20 cm higher than the treatment and outflow sites, it has not experienced the deeper inundation and still occasionally dries out. At all three sites, the relationship between long-term water levels and growth rates was hysteretic (path-dependent) (Fig. 6), indicating non-unique relationships between long-term water levels and growth rates. Initially, increased long-term water level was correlated with increased growth, but growth declined after long periods of continued high water.

Ten-year average tree ring chronologies (top) and water levels at nearby gauges (bottom). The Lake Verret chronology is from three sites adjacent to nearby Lake Verret reported by Amos (2006). The Lake Verret and Atchafalaya River gauges are US Army Corps of Engineers gauges at Attakapas Landing and Morgan City, respectively. The vertical line indicates the onset of effluent addition to the treatment site

Relationship between long-term water levels and tree ring chronologies. The arrows indicate direction of the progression through time. The large dot is 1992, which was the onset of effluent addition to the treatment site
Discussion
The expected positive relationship between effluent additions and tree growth did not exist at the study site. There was no demonstrable effect of the effluent discharge; instead there was a strong relationship between growth and inundation. Hydrological differences between the study site and other sites where added nutrients have increased growth may explain the lack of the expected response. Elsewhere, Hesse et al. (1998) and Hunter et al. (2009), also working in the Mississippi River deltaic plain, found baldcypress increased growth in response to the addition of treated effluent. However, that site was subject only to seasonal inundation, whereas our study site has remained flooded continuously for at least the entire length of treatment (13 years) and likely for about two decades prior to treatment. This hydrological regime has dominated growth responses and prevents robust conclusions regarding the effects of nutrient additions from wastewater on tree growth at our site.
There was a growth increase apparent in the baldcypress trees at our research site that predated the effluent application (Fig. 2a). The most likely cause of this is a release from competition from trees of other species; their mortality appears have been an important mechanism affecting growth responses to hydrological change. Increasing inundation causes mortality first in the species less tolerant of inundation, and stands eventually lose tree density under semipermanent inundation (Conner et al. 1981; Conner and Day 1992; Visser and Sasser 1995; Conner et al. 2002). Before effluent discharge began in 1992, bottomland hardwood species in the treatment area such as green ash were already dying and by 2008 the only living adult trees left were cypress and tupelo. Long-term increased inundation is almost certainly the cause for the widespread mortality of these trees. The presence of a few scattered, large, dwarf palmettos in the outflow site suggests that it, too, was once much less flooded than presently because dwarf palmetto cannot germinate in flooded conditions. Even in the control site (the highest elevation of the three), the species that are least flood tolerant have nearly all died. There are many young black willows and baldcypress in the control site, which are replacing other species less tolerant of inundation. We expect baldcypress at the control site to eventually experience the growth declines of the other two sites as water levels continue to rise.
The differences in hysteretic relationships between water level and growth among sites are likely related to thresholds in inundation effects on different species. As water levels increase, species that are not favored by the higher water drop out of the stand, and the remaining species experience a growth release. If water levels do not increase beyond the tolerance of baldcypress, they show sustained increased growth as in the control site. On the other hand, a hydrological threshold appears to have been exceeded in the treatment and outflow stands, leading to long-term decline of all tree species. Baldcypress may persist at these sites for some time, but the long-term successional pathway is likely to be to open water, shrub-scrub, or treeless marsh. This is consistent with findings for baldcypress in general in south Louisiana that are subject to permanent inundation (Keim et al. 2006).
The reason for the steeper decline in baldcypress growth at the outflow site compared to the treatment site (Fig. 2a) is unclear. Nutrient loading in the effluent may have directly prevented decline in the treatment site but less so in the outflow site because much of the inorganic nitrogen has been assimilated by the time water reaches the outflow site. The effluent may also have prevented deeper inundation by increasing production of aquatic vegetation that caused sufficient accretion to offset subsidence (Rybczyk et al. 2002; Izdepski et al. 2009). However, differentially greater decline in the outflow site began about 15 years prior to initiation of treatment, which suggests the differential decline is not likely related to the effluent. Stand density effects may explain part of the difference: trees in the treatment site are growing nearly without competition from adjacent trees because the original stand consisted of many individuals of species less tolerant of inundation. In contrast, the outflow site consists mostly of baldcypress, so that mortality of competing species with increased water level may have freed relatively fewer resources.
It may seem counterintuitive that wetter weather increased growth (Fig. 4) in this forested wetland where water is plentiful, but tree physiology and its relationship with redox processes may explain the paradox. First, baldcypress and pondcypress (T. ascendens Brongn.) in flooded conditions develop canopies with low leaf area (Brown 1981; Oren et al. 1999) and use water efficiently (Brown 1981). These characteristics likely relate to the high cost of water uptake in anaerobic soils; growth rates and water use are normally higher when roots are in aerobic or less-reduced conditions (Dickson and Broyer 1972; Pezeshki and DeLaune 1998; Pezeshki and Santos 1998). Although trees can adapt physiologically to improve oxidation of the rhizosphere (Megonigal and Day 1992; Kozlowski 1997), phytotoxins may be present in reduced conditions that inhibit free uptake of water (Kozlowski 1997; Pezeshki 2001). In cool, rainy weather, incident precipitation may partially aerate standing water, and cooler water increases the solubility of oxygen. Davidson et al. (2006) found that surface water penetrated to the rhizosphere in continuously flooded baldcypress during periods of rainfall, and speculated that attendant oxidation was responsible for increased growth in baldcypress when rainfall is high. Vapor pressure deficit and evaporative demand is lower during cooler weather with higher precipitation and humidity so that limits to growth originating in water uptake are relaxed.
Similar to the way cool, rainy growing seasons increased growth, short-term high water events increased growth. This relationship may be caused by the same processes that increased growth during cool, rainy weather because rain events also tend to increase water levels. However, high water levels in the winter also increased growth in the subsequent growing season even though photosynthesis is not active in winter. The most logical explanation is that high water events moved the hydrological regime away from stagnant and toward a flow-through system. Trees in swamps with flowing water that brings nutrients, flushes phytotoxins, and oxygenates the rhizosphere typically grow more rapidly than dystrophic, stagnant swamps (Conner and Day 1976; Mitsch and Ewel 1979; Brown 1981; Shaffer et al. 2009).
Tertiary treatment of municipal wastewater in natural wetlands is an increasingly applied tool to improve quality of receiving waters and reduce costs of treatment, while simultaneously increasing wetland productivity and restoring ecosystems degraded because of low nutrients or saltwater intrusion. At our study site, wastewater applications have increased overall productivity and organic accretion (Rybczyk et al. 1998, 2002). However, the increases have been in the incipient floating marsh at the site (Izdepski et al. 2009) rather than in the trees. Thus, adding wastewater as ecosystem restoration at this site has not benefitted the swamp forest ecosystem because rising water levels are creating limiting factors other than nutrient deprivation.
References
Allen JA, Pezeshki SR, Chambers JL (1996) Interaction of flooding and salinity stress on baldcypress (Taxodium distichum). Tree Physiology 16:307–313
Amos JB (2006) Dendrochronological analysis of productivity and hydrology in two Louisiana swamps. Thesis, Louisiana State University
Boustany RG, Crozier CR, Rybczyk JM, Twilley R (1997) Denitrification in a south Louisiana wetland forest receiving treated sewage effluent. Wetlands Ecology and Management 4:273–283
Briffa KR, Jones PD, Bartholin TS, Eckstein D, Schweingruber FH, Karlén W, Zetterberg P, Eronen M (1992) Fennoscandian summers from AD 500: temperature changes on short and long timescales. Climate Dynamics 7:111–119
Brown SL (1981) A comparison of the structure, primary productivity, and transpiration of cypress ecosystems in Florida. Ecological Monographs 51:403–427
Conner WH, Day JW Jr (1976) Productivity and composition of a baldcypress–water tupelo site and a bottomland hardwood site in a Louisiana swamp. American Journal of Botany 63:1354–1364
Conner WH, Day JW Jr (1992) Diameter growth of Taxodium distichum (L.) Rich. and Nyssa aquatica L. from 1979–1985 in four Louisiana swamp stands. American Midland Naturalist 127:290–299
Conner WH, Toliver JR (1990) Long-term trends in the baldcypress (Taxodium distichum (L.) Rich.) resource in Louisiana. Forest Ecology and Management 33(34):543–557
Conner WH, Gosselink JG, Parrondo RT (1981) Comparison of the vegetation of three Louisiana swamp sites with different flooding regimes. American Journal of Botany 68:320–331
Conner WH, Mihalia I, Wolfe J (2002) Tree community structure and changes from 1987 to 1999 in three Louisiana and three South Carolina forested wetlands. Wetlands 22:58–70
Cook ER (1985) A time series analysis approach to tree-ring standardization. Dissertation, University of Arizona
Cook ER (1987) The decomposition of tree-ring series for environmental studies. Tree-Ring Bulletin 47:37–59
Cook ER, Holmes RL (1986) Guide for program ARSTAN. Laboratory of Tree-Ring Research, University of Arizona
Crozier CR, Rybczyk JM, Patrick WH (1996) Spatial gradients of dissolved nitrate and nitrous oxide in a wetland forest receiving treated sewage effluent. In: Flynn K (ed) Proceedings of the Southern forested wetlands ecology and management conference. Clemson University, South Carolina, pp 65–68
Davidson GR, Laine BC, Galicki SJ, Threlkeld ST (2006) Root zone hydrology: why bald cypress in flooded wetlands grow more when it rains. Tree-Ring Research 62:3–12
Day JW Jr, Ko J-Y, Rybczyk J, Sabins D, Bean R, Berthelot G, Brantley C, Cardoch L, Conner W, Day JN, Englande AJ, Feagley S, Hyfield E, Lane R, Lindsey J, Mitsch J, Reyes E, Twilley R (2004) The use of wetlands in the Mississippi Delta for wastewater assimilation: a review. Ocean & Coastal Management 47:671–691
Day JW Jr, Boesch DF, Clairain EJ, Kemp GP, Laska SB, Mitsch WJ, Orth K, Mashriqui H, Reed DJ, Shabman L, Simenstad CA, Streever BJ, Twilley RR, Watson CC, Wells JT, Whigham DF (2007) Restoration of the Mississippi Delta: lessons from hurricanes Katrina and Rita. Science 315:1679–1684
Dickson RE, Broyer TC (1972) Effect of aeration, water supply, and nitrogen source on growth and development of tupelo gum and bald cypress. Ecology 53:626–634
Effler RS, Goyer RA (2006) Baldcypress and water tupelo sapling response to multiple stress agents and reforestation implications for Louisiana swamps. Forest Ecology and Management 226:330–340
Esper J, Cook ER, Krusic PJ, Peters K, Schweingruber FH (2003) Tests of the RCS method for preserving low-frequency variability in tree-ring chronologies. Tree-Ring Research 59:81–98
Ewel KC, Parendes LA (1984) Usefulness of annual growth rings of cypress trees (Taxodium distichum) for impact analysis. Tree-Ring Bulletin 44:39–43
Fritts HC (1976) Tree rings and climate. Academic, San Diego
Grissino-Mayer HD (2001) Evaluating crossdating accuracy: a manual and tutorial for the computer program COFECHA. Tree-Ring Research 57:205–221
Hesse ID, Day JW Jr, Doyle TW (1998) Long-term growth enhancement of baldcypress (Taxodium distichum) from municipal wastewater application. Environmental Management 22:119–127
Holmes RL (1983) Computer assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin 43:69–78
Hunter RG, Day JW Jr, Lane RR, Lindsey J, Day JN, Hunter MG (2009) Impacts of secondarily treated municipal effluent on a freshwater forested wetland after 60 years of discharge. Wetlands 29:363–371
Izdepski CW, Day JW Jr, Sasser CE, Fry B (2009) Early floating marsh establishment and growth dynamics in a nutrient amended wetland in the lower Mississippi delta. Wetlands 29:1004–1013
Keeland BD, Young PJ (1997) Long-term growth trends of baldcypress (Taxodium distichum (L.) Rich.) at Caddo Lake, Texas. Wetlands 17:559–566
Keeland BD, Conner WH, Sharitz RR (1997) A comparison of wetland tree growth response to hydrologic regime in Louisiana and South Carolina. Forest Ecology and Management 90:237–250
Keim RF, Chambers JL, Hughes MS, Nyman JA, Miller CA, Amos JB, Conner WH, Day JW Jr, Faulkner SP, Gardiner ES, King SL, McLeod KW, Shaffer GP (2006) Ecological consequences of changing hydrological conditions in wetland forests of coastal Louisiana. In: Xu YJ, Singh VP (eds) Coastal environment and water quality. Water Resource Publications, Highlands Ranch, Colo, pp 383–395
Kozlowski TT (1997) Responses of woody plants to flooding and salinity. Tree Physiology Monograph 1.
Megonigal JP, Day FP (1992) Effects of flooding on root and shoot production of bald cypress in large experimental enclosures. Ecology 73:1182–1193
Mitsch WJ, Ewel KC (1979) Comparative biomass and growth of cypress in Florida wetlands. American Midland Naturalist 101:417–426
Nessel JK, Ewel KC, Burnett MS (1982) Wastewater enrichment increases mature pondcypress growth rates. Forestry Sciences 28:400–403
Oren R, Phillips N, Ewers BE, Pataki DE, Megonigal JP (1999) Sap-flux-scaled transpiration responses to light, vapor pressure deficit, and leaf area reduction in a flooded Taxodium distichum forest. Tree Physiology 19:337–347
Orvis KH, Grissino-Mayer HD (2002) Standardizing the reporting of abrasive papers used to surface tree-ring samples. Tree-Ring Research 58:47–50
Penland S, Ramsey KE (1990) Relative sea-level rise in Louisiana and the Gulf of Mexico: 1908–1988. Journal of Coastal Research 6:323–342
Pezeshki SR (2001) Wetland plant responses to flooding. Environmental and Experimental Botany 46:299–312
Pezeshki SR, DeLaune RD (1998) Responses of seedlings of selected woody species to soil oxidation-reduction conditions. Environmental and Experimental Botany 40:123–133
Pezeshki SR, Santos MI (1998) Relationships among rhizosphere oxygen deficiency, root restriction, photosynthesis, and growth in baldcypress (Taxodium distichum L.) seedlings. Photosynthetica 35:381–390
Rybczyk JM, Day JW Jr, Hesse ID, Delgado-Sanchez P (1996) An overview of forested wetland wastewater treatment projects in the Mississippi River delta region. In: Flynn K (ed) Proceedings of the southern forested wetlands ecology and management conference. Clemson University, South Carolina, pp 78–82
Rybczyk RM, Callaway JC, Day JW Jr (1998) A relative elevation model for a subsiding coastal forested wetland receiving wastewater effluent. Ecological Modelling 112:23–44
Rybczyk RM, Day JW Jr, Conner WH (2002) The impact of wastewater effluent on accretion and decomposition in a subsiding forested wetland. Wetlands 22:18–32
Shaffer GP, Wood WB, Hoeppner SS, Perkins TE, Zoller J, Kandalepas D (2009) Degradation of baldcypress-water tupelo swamp to marsh and open water in southeastern Louisiana, U.S.A: an irreversible trajectory? Journal of Coastal Research 54:152–165
Speer JH (2010) Fundamentals of tree-ring research. University of Arizona Press, Tucson
Stahle DW, Cleaveland MK (1992) Reconstruction and analysis of spring rainfall over the southeastern United-States for the past 1000 Years. Bulletin of the American Meteorological Society 73:1947–1961
Stahle DW, Cook ER, White JWC (1985) Tree-ring dating of baldcypress and the potential for millenia-long chronologies in the southeast. American Antiquity 50:796–802
Stahle DW, Cleaveland MK, Hehr JG (1988) North Carolina climate changes reconstructed from tree rings: A.D. 372 to 1985. Science 240:1517–1519
Stokes MA, Smiley TL (1968) An introduction to tree-ring dating. University of Chicago Press.
Swarzenski CM (2003) Surface-water hydrology of the Gulf Intracoastal Waterway in South-Central Louisiana, 1996–99. USGS Professional Paper 1672
Visser JM, Sasser CE (1995) Changes in tree species composition, structure and growth in a bald cypress–water tupelo swamp forest, 1980–1990. Forest Ecology and Management 72:119–129
Young PJ, Megonigal P, Sharitz RR, Day FP (1993) False ring formation in baldcypress (Taxodium distichum) saplings under two flooding regimes. Wetlands 13:293–298
Young PJ, Keeland BD, Sharitz RR (1995) Growth-response of baldcypress (Taxodium distichum (L) Rich) to an altered hydrologic regime. American Midland Naturalist 133:206–212
Zhang X, Feagley S, Day JW Jr, Conner W, Hesse I, Rybczyk J, Hudnall W (2000) A water chemistry assessment of wastewater remediation in a natural swamp in Louisiana. Journal of Environmental Quality 29:1960–68
Acknowledgments
This study was funded by CSREES/USDA under project number LAB93735 and supported by funds from the City of Thibodaux. This is manuscript 2009-241-4066 of the Louisiana Agriculture Experiment Station. Richard Hartman, Ben Hartman, Jason Zoller, and Blake Amos assisted with field data collection and laboratory analysis.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Keim, R.F., Izdepski, C.W. & Day, J.W. Growth Responses of Baldcypress to Wastewater Nutrient Additions and Changing Hydrologic Regime. Wetlands 32, 95–103 (2012). https://doi.org/10.1007/s13157-011-0248-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13157-011-0248-6