
Abstract
Shrubs are important components of alpine mountain ecosystems in terms of productivity and diversity. An estimation of shrub biomass using allometric equations represents a non-destructive option for obtaining useful quantitative data. However, species-specific allometric equations for alpine shrubs have yet to be developed in sufficient detail. This study proposed allometric equations that can be used to estimate aboveground biomass of alpine shrubs. These are based on easily acquired descriptive parameters of plant height (H), crown area (C), basal diameter (D), and D 2 H (basal diameter squared × total H) for four of the most abundant alpine shrub species in the upstream ecosystems of China’s Heihe River Basin: Salix cupularis, Salix oritrepha, Potentilla fruticosa, and Caragana jubata. The results show that several equations relating biomass categories to H, C, and D 2 H provided significant (P < 0.01) data for measuring aboveground biomass; the determination coefficients (R 2) varied from 0.95 to 0.97 and fit indices (FIs) varied from 0.96 to 0.97. The form and variables comprising the allometric equations differed among species-specific models. Aboveground shrub biomass increased with D 2 H for S. cupularis and C. jubata, whereas it increased with C and H for P. fruticosa. For S. oritrepha, an exponential function provided the best-fit model with C. However, the results could provide useful approximations of aboveground biomass of similar shrubs currently lacking such equations. The allometric equations also provide a useful and non-destructive method of estimating aboveground shrub biomass in alpine ecosystems.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Accurately estimating forest biomass is essential to understanding how plant growth is relate to the commercial production of wood fiber (Morgan and Moss 1985), energy transfer (David et al. 2006), nutrient flows (Rapp et al. 1999), and the global carbon cycle (Houghton 2007; Bombelli et al. 2009). The characterization of variability and trends in forest biomass at local to national scales is needed for monitoring the sources and sinks in the global carbon cycle (Ganguly et al. 2012). Therefore, the study of aboveground biomass in rapidly changing environments has been a central theme in ecological and hydrological research (Dong et al. 2011; Wang et al. 2014). Shrublands form a major component of vegetation in alpine ecosystems worldwide and are a widely distributed biome type in China (Hu et al. 2006). Shrublands cover 19 % of the Qinghai-Tibet Plateau and approximately 20 % of China (Hou 2001; Hu et al. 2006). Recent research using repeated photography, long-term ecological monitoring, and dendrochronology have documented shrub expansion in some cold regions, which could cause major changes in biodiversity and result in increased shrub cover and biomass in recent decades (Tape et al. 2006; Myers-Smith 2011). Rapid climatic warming has allowed the rapid increasing of shrublands in China over the past two decades (Fang et al. 2007). Piao et al. (2009) identified shrub increasing as the most uncertain factor contributing to carbon sinks of forest ecosystems in China. A need exists to improve the accuracy of the estimation of alpine shrub biomass because biomass data provide valuable information for explaining the functional role of shrub ecosystems as part of the carbon and hydrological balances in the alpine regions of China (Yashiro et al. 2011; Liu et al. 2014).
Compared with arboreal vegetation, shrubs have often been neglected during biomass estimation research. Shrubs come in a wide variety of forms, both single and multiple stemmed. Thus, accurate estimation of shrub biomass has proved to be relatively difficult owing to the mixture of species typically present in some shrubland ecosystems (Sah et al. 2004). A large amount of research has delved into biomass estimation of individual shrub species in the some temperate and tropical regions using allometric equations (Murray and Jacobson 1982; Hierro et al. 2000; Paton et al. 2002; Návar et al. 2004). However, little research has explored the development of an accurate method to assess shrub biomass in alpine ecosystems (Elzein et al. 2011).
Traditionally, researchers have performed biomass estimation in forest ecosystems using direct destructive or extractive methods, or using indirect methods such as allometric equations (Keller et al. 2001). Undoubtedly, direct methods provide the most accurate biomass estimates; however, they are time-consuming and laborious (Sironen et al. 2003; Case and Hall 2008; Liu and Westman 2009). Allometric equations use mathematical models to estimate vegetative biomass based on various morphological variables such as basal diameter of stems and branches, canopy area, and H (Návar et al. 2004; Elzein et al. 2011). Indirect methods have been developed to estimate some dimensional variables by fitting specific equations for individual plant species or community models (Murray and Jacobson 1982; Návar et al. 2004). Therefore, researchers have developed allometric equations designed for use with specific to forest types (Basuki et al. 2009; Xiang et al. 2011) and shrublands ecosystems (Murray and Jacobson 1982; Uso et al. 1997; Elzein et al. 2011; Hierro et al. 2000; Návar et al. 2004; Foroughbakhch et al. 2005; Sah et al. 2004). Predictive mathematical models have normally been tested using simple linear or multiple regression models (Murray and Jacobson 1982). The relationship between biomass and measured variables has usually been determined using a nonlinear function (Foroughbakhch et al. 2005). Different types of variables and functions have been used for predictive model equations (Návar et al. 2004). Singh et al. (2011) estimated shrub aboveground biomass using a destructive harvesting method in the central Himalayan region of Terai; Luo et al. (2002) estimated biomass and natural vegetation productivity based on forest survey data and field sampling of the Tibetan Plateau. These results indicated that allometric equations provide useful tools that can be used to estimate biomass for shrub ecosystems in cold regions, because they offer a non-destructive and time-efficient method of estimating biomass (Uso et al. 1997; Návar et al. 2004).
The Heihe River originates from the Qilian Mountains, scatters itself across the landscape in the middle reaches oasis region, and disappears into the desert lower reaches. Located between 97.24–102.10°E and 37.41–42.42°N, the Heihe River forms the second largest inland river in Northwestern China and covers about 130,000 km2 (Chen et al. 2007). Well-developed vegetation thrives at elevations from 1,600 to 5,500 m a.s.l. in the mountainous catchment, where the annual average precipitation is about 400 mm. Shrublands cover 1,352 km2, or about 13.7 % of the upper reaches area, in the mountain region of Heihe River Basin (Hou 2001). Developing accurate allometric equations is essential for shrub biomass estimation in this region for two reasons. First, alpine shrubs form the dominant vegetation type in the alpine valleys; this ecosystem has rather extreme conditions making it difficult for arbor species to survive; therefore, shrubs play an important role in water conservation and in the carbon cycle of the mountain environment. Second, the coverage area and average patch area of shrubs have increased by 6.8 and 49 %, respectively, over the past 15 years in a process recently described as “shrubification” in this region (Liu et al. 2011). However, biomass estimation using allometric equations for individual shrub species in the Heihe River Basin has received little attention (Zhang and Han 2008).
This study primarily attempts to establish correlation and regression relationships between aboveground biomass and easily measured parameters (H: height, D: basal diameter, C: crown area, and D 2 H: basal diameter squared × total H) of the alpine shrubs in the upper reaches of Heihe River Basin in Northwestern China. It also attempts to develop general allometric equations designed specifically for the four most common shrub species found here.
Materials and methods
Site description
The study area, located in the Hulu watershed, covers 23.1 km2 in the Qilian Alpine Ecology and Hydrology Research Station within the Qilian Mountains, Qinghai Province, China (38°12′–38°17′N, 99°49′–99°54′E; Fig. 1). The elevation of the Hulu watershed varies from 2,960 to 4,800 m a.s.l; the average annual temperature is about 0.2 °C, with a low mean of −18.4 °C in January and a high reaching 19.0 °C in August. The mean annual precipitation is 495.1 mm, with 85 % falling from June to October, based on 4 years of manual and electronic rain gauge data collected from 2008 to 2012 (Liu et al. 2014).

Locations of the sampling sites in the Hulu Watershed of the Heihe River Basin, Qinghai Province, China
The growing season for vascular plants lasts from early June to mid-September, with vegetation in the Hulu watershed including tree Picea crassifolia and Sabina przewalskii, alpine shrubs and meadows. Shrubs occur mainly on shady, semi-shady, and semi-sunny slopes in the Qilian Mountains. The most dominant shrub species, Salix cupularis, Salix oritrepha, Potentilla fruticosa, Caragana jubata, and Hippophae rhamnoides, occur mainly from 3,000 m to 3,700 m in the Qilian Mountains (Wang et al. 2002). The dominant herbs include Polygonum viviparum, Poa pratensis, Stipa capillata, and Stellera chamaejasme. Mountain drab, chestnut, and subalpine shrub meadow soils cover most of the Hulu watershed (Liu et al. 2014).
Shrub sampling
While considering the characteristics of the distributions of alpine shrubs, four shrubs species, S. cupularis, P. fruticosa, C. jubata, and S. oritrepha, were selected for analysis. Three 10 m × 10 m quadrates were established for each shrub species. Twenty shrubs per quadrate were randomly selected as sample individuals. Therefore, 60 shrubs per species were used to calculate correlation and regression relationships. The shrubs were monitored in mid-September 2012 during the peak season of increasing biomass, when biophysical parameters such as foliage biomass and canopy cover are relatively stable. The elevation, aspect, slope angle, and shrub density of the plots were documented, and the density was obtained by quadrat method (Xiang et al. 2011). The shrubs were harvested using the selective destruction technique (Lodhiyal and Lodhiyal 1997). Prior to harvest, the following data were measured for all sampled shrubs using a digital caliper (XL.10-160, Beijing Zhuochuan Co., Beijing, China): (1) basal diameter (D: diameter at 10 cm above ground, cm); (2) height (H: height from ground level for the tallest point in the canopy, cm); and (3) crown area (C: crown width × crown length, measured at right angles, cm2).
In the laboratory, the fresh weight of all aboveground sections of shrubs was determined to the nearest 0.01 g using a balance (MP21001; Shanghai Hengping Co., Shanghai, China). The samples were then placed in envelopes and dried for 48 h in an oven at 80 °C to a constant weight. Finally, at room temperature, the dry weights of the samples were recorded as the biomass.
Data analysis
First, prior to establishing the allometric equations, scatter plots between the dependent and independent variables were used to determine whether the relationships were linear. Scatter plots were also used to determine the types of relationships existing between shrub biomass and each of the measured and calculated variables. Second, several allometric relationships between the dependent and independent variables were subsequently tested. The independent variables, represented by X, included D, H, D 2 H, and C. Aboveground shrub biomass was the dependent variable, W. The following six models were used in this study: (1) W = a + bX; (2) W = a + bX 1 + cX 2; (3) W = a + b ln X; (4) W = a + bX + cX 2; (5) W = aX b; and (6) ln (W) = ln (a) + bX.
Correlation and regression analyses between total aboveground biomass and field measured parameters (D, H, D 2 H, and C) were conducted using the SPSS 16.0 statistical package. The regression model with the highest R 2 and the lowest root mean square error (RMSE) with a significant fit (P < 0.05) was selected as the best model (Davison and Hinkley 1997). However, the results of R 2 statistical analysis could not be directly compared among equations with untransformed and log-transformed independent variables (Draper and Smith 1998). To compare such equations, a Fit Index (FI) that had a value of unity for an optimum fit similar to R 2 was calculated (Schlaegel 1981). The FI was based on the residuals in the measured units (Brand and Smith 1985; Crow and Schlaegel 1988) based on Eq. (1):
where \(Y_{i}\) is the ith observed value, \(\hat{Y}_{i}\) is the ith predicted value for \(Y_{i}\), and \(\bar{Y}\) is the mean observed value of \(Y_{i}\). If the FI calculated for a logarithmic model was closer to the R 2 for a linear model, the former was preferred, given that the logarithmic transformation increased the statistical validity of the analysis by homogenizing the variance over the range of sample data. The logarithmic model was suitable for data in which variance increased with an increase in the magnitude of the observation (Sah et al. 2004).
When more than one model presented a similarly good fit with the data, the regression equation with the fewest parameters was selected as the best model. Although logarithmic models are often used to predict shrub biomass, logarithmic transformations could sometimes result in biased biomass estimates. Therefore, when a logarithmic model was selected as the best model, the model was transformed into the corresponding nonlinear model.
Results
Correlation relationships between aboveground biomass and variables
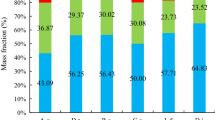
To determine the shrub biomass and develop the allometric equations, the independent variables had to be determined. The variables of four shrub species in this study exhibited no homogeneous distribution between different quadrats (Fig. 2).

Dependent and independent variables of the shrubs (Mean ± SD). D basal diameter (at 10 cm above the ground), H height from ground level to the tallest point in the canopy, C crown area (crown width × crown length, measured at a right angle), W aboveground biomass
Table 1 illustrates the correlation between W and D, H, D 2 H, and C for four shrubs. The correlation coefficient values indicated that a positive correlation existed among all dependent variables and independent variables. The correlation coefficient R values varied from 0.560 for H in S. cupularis to 0.977 for D 2 H in S. oritrepha among species-specific correlations. The measured parameters allowed an effective estimation of shrub biomass. However, the correlations of D, D 2 H, and C with W were significantly higher than that of H with W. Moreover, the indicators of multicollinearity [tolerance, variance inflation factor (VIF)] were statistically examined: the three variables D, D 2 H, and C were correlated. These correlations suggested that a combination of these three variables could provide the best regression model for biomass estimation. Compared with the other variables, the correlation between D 2 H and W was higher for C. jubata, S. cupularis, and S. oritrepha. As shown in the following model, the best parameters for biomass estimation using an allometric equation were determined based on the correlation results (Table 2).
The scatter plot diagrams between each dependent and independent variable could allow advance determination of the expected sign of the coefficient of a particular variable in the regression model (Fig. 3). Despite showing the evident correlations between dependent and independent variables, an effective model could not be developed by employing all measured variables simultaneously because of the multicollinearity among measured independent variables. The tolerances were 0.001, 0.018, and 0.021 and the VIFs were 1.23, 1.54, and 3.15 for D, D 2 H, and C, respectively. Therefore, these three variables could not be included in an equation at the same time.

The scatter plot diagrams between each dependent and independent variable. a–d represent data for P. fruticosa, C. jubata, S. cupularis, and S. oritrepha, respectively. D basal diameter (at 10 cm above the ground), H height from ground level to the tallest point in the canopy, C crown area (crown width × crown length, measured at a right angle)
Models that include all measured variables could result in an unexpected negative sign on the coefficient, subsequently giving rise to high values of RMSE. Therefore, to avoid this issue, stepwise regression was used to establish the allometric equations of shrub biomass. This caused these models to improve significantly; they had the fewest parameters with the highest R 2 and FIs values as well as the lowest RMSE values.
Developing allometric equations
The biomass equations based on the variables measured in the field proved an excellent fit for the estimated biomass (Table 2). All of the species-specific regression equations relating biomass to the measured variables were statistically significant (P < 0.01) with equations proposed for each species able to satisfy the requirement of the equations. No self-correlation or heteroscedasticity was observed in the distribution of residues. Notably, all equations had an R 2 higher than 0.95 and an FI higher than 0.96. R 2 values varied from 0.95 for C. jubata and S. cupularis to 0.97 for S. oritrepha among species-specific best-fit models (Table 2). However, the forms of the best-fit model differed among shrub species (Table 2). The power equations were the ideal models for C. jubata and S. cupularis, having the highest R 2 and FI and the lowest RMSE. Exponential equations were chosen as the ideal models for S. oritrepha. Linear models showed the best fit for P. fruticosa (Table 2).
Species-specific allometric equations used different variables and combinations of variables to estimate biomass. When the R 2, FI, and RMSE of the regression equations were compared, D 2 H was found to be the best parameter for predicting C. jubata and S. cupularis biomass. In contrast, equations with C as the independent variable were the best for estimating S. oritrepha biomass. Moreover, H and C were the best parameters for estimating P. fruticosa biomass (Table 2).
Discussion
Direct harvesting techniques for the estimation of shrub biomass are labor intensive and time-consuming. Researchers commonly use allometric equations as a nondestructive alternative, in which biomass is estimated based on the easily measured parameters of shrubs (Návar et al. 2004; Elzein et al. 2011). Mathematical forecasting models for the biomass productivity and carbon cycling in forest ecosystem research could provide an estimation tool and data in support of large-scale models. The relationships between aboveground biomass and measured variables were determined for the four most dominant alpine shrubs in the upper reaches of Heihe River Basin (Wang et al. 2002). The measured variables showed a typical linear correlation with dependent variables, and allometric equations were obtained to estimate the aboveground biomass of the four alpine shrubs. Generally, aboveground biomass is best determined through the analysis of D, H, C, and other variables; these types of variables are typically used as independent variables in shrub biomass regression equations (Murray and Jacobson 1982). However, some researchers continue to establish biomass equations for individual shrub species by relating biomass variables to total shrub characteristics. Stem diameter (D) alone or in combination with H is the best predictor of biomass for some shrub species (Haase and Haase 1995; Paton et al. 2002; Liu et al. 2006; He et al. 2011). In this study, D 2 H provided the most accurate prediction of aboveground biomass in two shrub species, C. jubata and S. cupularis, which was consistent with previous research in other alpine regions (Elzein et al. 2011). The canopy morphology of shrubs serves as an important factor influencing prediction accuracy of model and variables selection. The canopies of C. jubata and S. cupularis form elliptical cones, and height and basal diameter can replace the distribution of branches and leaves during biomass estimation. Zhang and Han (2008) estimated the aboveground biomass of shrubs in the Eastern Qilian Mountains and developed a model with a similar type of equations. However, their number of sampled individual shrubs and the distribution of the canopy size were undocumented.
Previous studies have used crown area to estimate shrub biomass (Peek 1970; Sah et al. 2004), particularly for low-growing shrubs with a multiple-stemmed growth form, which was not amenable to the measurement of individual diameters (Elzein et al. 2011). Harrington (1979) found that the weakest relationships and highest estimation errors were obtained when D was used as the independent variable in small multi-stemmed shrubs. Elzein et al. (2011) used crown area and mean height to develop equations for shrub biomass estimation in the western Alps. In this study, C could be used to accurately predict biomass for the multi-stemmed shrub species S. oritrepha (Table 2). Some studies have not commonly included canopy height into biomass regression equations (Peek 1970; Harrington 1979; Murray and Jacobson 1982; Zhang and Han 2008). However, the addition of height in the models developed for the shrubs was reasonable although the correlation between the height and biomass was lower than that of the other three variables (Tables 1 and 2). Halpern et al. (1996) reported that models considering crown area did not explain variations associated with extension growth, although variation associated with lateral branching was explained. Therefore, the addition of H increased the precision in estimating biomass for P. fruticosa in this study.
In contrast to previous studies using allometric equations to estimate alpine shrub biomass, the present study applied the stepwise regression and used R 2, FI, and RMSE to evaluate the accuracy of the models. In this study, the power, linear, and exponential models provided the most appropriate equations for estimating and describing the relationships between measured variables and aboveground biomass. The best-fit models proposed in the results were consistent with those reported in previous studies of shrubs in the Gannan Grasslands (Zhang and Han 2008). That study estimated biomass using linear regression models to determine the relationships between shrub biomass and crown width. Power and linear equations have commonly been used to model shrub biomasses in subtropical regions (Cai et al. 2006; Zeng et al. 2010), such as the Jinggang and Balangshan Mountains of China (Lin et al. 2010; Liu et al. 2006, respectively), western Andalusia of Spain (Oyonarte and Cerrillo 2003), and northeastern Mexico (Návar et al. 2004). Li et al. (2006) assessed the accumulated stem biomass of P. fruticosa using a linear regression equation. Shrub biomasses in the western Alps were also estimated using logarithmic equations (Elzein et al. 2011). The best-fit equation for S. oritrepha in the current study was a logarithmic equation. However, the equation was transformed to a nonlinear form.
Plant functional types (PFTs) were different from species-specific methods in biomass estimation models (Ustin and Gamon 2010). PFTs were derived from traits based on species morphology, physiology, and/or life history, depending on the aims and scale of the research. Some traits could be measured in the field, and others required more detailed laboratory measurement and experimentation (Duckworth et al. 2000). PFTs bridged the gap between plant physiology and both community and ecosystem processes, thus providing a powerful tool in carbon and biomass research (Díaz and Cabido 1997). PFTs were more important than specific species in modeling the estimating biomass at the ecosystem scale (Manzoni et al. 2011). However, for each PFT, a number of key parameters needed to be defined, such as fecundity, competitiveness, and resorption. The value of each parameter is determined or inferred from observable characteristics such as plant height and leaf area (Lavorel et al. 2007). PFTs will be most useful if the same grouping of species describes a response to, and effects on, multiple environmental factors and ecosystem processes (Chapin et al. 1996). In alpine regions, the microclimate induced by topographical change is very large at the watershed scale. Thus, the parameters of PFTs varied with the changes in elevation and slope (Spadavecchia et al. 2008). The biomass of PFTs varied along a climatic gradient because of the heterogeneous nature of the environment (Liu et al. 2014). In this study, the specific species method was selected to estimate the biomass of shrubs rather than using PFTs because this method is more suitable for accurately modeling shrub biomass in this heterogeneous mountainous area. Species-specific equations could provide the essential data and method needed for estimating biomass of PFTs in a large-scale biomass model. Leaf area index (LAI) is an important variable in many ecological and environmental applications and is usually used in biomass estimation models at the ecosystem scale (Luo et al. 2002; Ganguly et al. 2012). An allometric relationship has been observed between leaf area and stem diameter, crown dimensions, and biomass (Spadavecchia et al. 2008). Specific leaf area (SLA) has been used as an indicator of the relative growth rate (RGR), which represented the productivity and biomass of an ecosystem (Díaz and Cabido 1997). In that study, RGR represented the productivity and biomass of ecosystem while height of the crown base affected SLA. Generally, the LAI increased from the tree top downward to the crown base and decreased significantly with increasing tree size (Seidel et al. 2011). Ganguly et al. (2012) presented an approach based on remote sensing for mapping live forest aboveground biomass, using a model that calculated LAI derived from remote sensing and canopy maximum height. Bartelink (1997) found a strong linear relationship between leaf area and crown area. Therefore, some parameters used in allometric equations for estimating biomass could allow the calculation of a LAI. The species-specific and PFT estimation methods could be combined with environmental variables in future research. The combination will allow researchers to overcome the biases in biomass estimation caused by inconsistencies in field inventories and offers a high-resolution and up-scale approach for forest carbon monitoring and productivity estimation in larger scale ecosystem models.
Conclusions
Researchers have recognized the usefulness of allometric equations for decades, but few equations have been developed for alpine shrubs. The equations presented in this work could be useful in quantifying aboveground biomass in the upper reaches area of the Heihe River Basin. Empirical models related to measured parameters were proposed for alpine shrubs. These models could be applied in biomass estimation based on the strong correlations between each variable and aboveground biomass. The proposed models may also be applicable to other alpine shrubland ecosystems by re-determining the coefficient for the statistical relationships between biomass and the measured variables. Ideally, all the proposed equations should be tested with larger data sets encompassing ecological contexts that were not considered in the present study, including gradients in elevation and slope. Such equations will provide useful data for studies estimating aboveground biomass and allow researchers to test the contribution of shrubs to overall productivity of an alpine ecosystem. The equations provided here could also significantly improve the accuracy of biomass estimation and allow researchers to develop methods designed to increase carbon sequestration in alpine ecosystems. However, care should be taken in applying the allometric models developed in this study to other sites when the structure and size of shrubs is unknown. For example, Zhang and Han (2008) found a shrub height of more than 2.1 m resulted in a loss of model accuracy. The models should also undergo further tests to determine their accuracy for estimating shrub biomass in other alpine regions.
References
Bartelink HH (1997) Allometric relationships for biomass and leaf area of beech (Fagus sylvatica L.). Ann For Sci 54(1):39–50
Basuki TM, Van Laake PE, Skidmore AK, Hussinl YA (2009) Allometric equations for estimating the above-ground biomass in tropical lowland Dipterocarp forests. Forest Ecol Manag 257:1684–1694
Bombelli A, Henry M, Castaldi S, AduBredu S, Arneth A, de Grandcourt A, Grieco E, Kutsch WL, Lehsten V, Rasile A, Reichstein M, Tansey K, Weber U, Valentini R (2009) An outlook on the Sub-Saharan Africa carbon balance. Biogeosciences 6:2193–2205
Brand GJ, Smith WB (1985) Evaluating allometric shrub biomass equationsfit to generated data. Can J Bot 63:63–67
Cai Z, Liu QJ, Ouyang QL (2006) Estimation model for biomass of shrub in Qianyanzhou experiment station. J Cent South Forest Univ 26:15–23 (in Chinese with English Abstract)
Case BS, Hall RJ (2008) Assessing prediction errors of generalized tree biomass and volume equations for the boreal forest region of west-central Canada. Can J Forest Res 38:878–889
Chapin FS, Bret-Harte MS, Hobbie SE, Zhong HL (1996) Plant functional types as predictors of transient responses of arctic vegetation to global change. J Veg Sci 7(3):347–358
Chen RS, Kang ES, Ji XB, Yang JP, Wang JH (2007) An hourly solar radiation model under actual weather and terrain conditions: a case study in Heihe river basin. Energy 32:1148–1157
Crow TR, Schlaegel BE (1988) A guide to using regression equations for estimating tree biomass. North J Appl For 5:15–22
David V, Sautour B, Galois R, Chardy P (2006) The paradox high zooplankton biomass low vegetal particulate organic matter in high turbidity zones: what way for energy transfer? J Exp Mar Biol Ecol 333:202–218
Davison AC, Hinkley DV (1997) Bootstrap methods and their application. Cambridge University Press, Cambridge
Díaz S, Cabido M (1997) Plant functional types and ecosystem function in relation to global change. J Veg Sci 8(4):463–474
Dong J, Tao F, Zhang G (2011) Trends and variation in vegetation greenness related to geographic controls in middle and eastern Inner Mongolia, China. Environ Earth Sci 62(2):245–256
Draper NR, Smith H (1998) Applied regression analysis, 2nd edn. Wiley, New York
Duckworth JC, Kent M, Ramsay PM (2000) Plant functional types: an alternative to taxonomic plant community description in biogeography? Prog Phys Geog 24(4):515–542
Elzein TM, Blarquez O, Gauthier O, Carcaillet C (2011) Allometric equations for biomass assessment of subalpine dwarf shrubs. Alpine Bot 121:129–134
Fang JY, Guo ZD, Piao SL, Chen AP (2007) Terrestrial vegetation carbon sinks in China, 1981–2000. Sci China Earth Sci 50:1341–1350
Foroughbakhch R, Reyes G, Alvarado-Vázquez MA, Hernández-Piñero J, Alejandra Rocha-Estrada A (2005) Use of quantitative methods to determine leaf biomass on 15 woody shrub species in northeastern Mexico. Forest Ecol Manag 216:359–366
Ganguly S, Zhang G, Nemani R, Miles C, Wang W, Saatchi S, Yu Y, Myneni R (2012) A simple parametric estimation of live forest aboveground biomass in california using satellite derived metrics of canopy height and leaf area index. AGU Fall Meet Abstr 1:0433
Haase R, Haase P (1995) Above-ground biomass estimates for invasive trees and shrubs in the Pantanal of Mato Grosso, Brazil. Forest Ecol Manag 73:29–35
Halpern CB, Miller EA, Geyer MA (1996) Equations for predicting aboveground biomass of plant species in early successional forests of the western Cascade Range, Oregon. Northwest Sci 70:306–320
Harrington G (1979) Estimation of above-ground biomass of trees and shrubs in a Eucalyptus populaca F. Muell woodland by regression of mass on trunk diameter and plant height. Aust J Bot 27:135–143
He LY, Kang XG, Fan XL, Gao Y, Feng QX (2011) Estimation and analysis of understory shrub biomass in Changbai Mountains. J Nanjing Forest Univ (Nat Sci Edition) 35:45–50 (in Chinese with English Abstract)
Hierro JL, Branch LC, Villareal D, Clark KL (2000) Predictive equations for biomass and fuel characteristics of Argentine shrubs. J Range Manag 53:617–621
Hou XY (2001) Vegetation Atlas of China (1: 1 000 000). Science Press, Beijing (in Chinese)
Houghton RA (2007) Balancing the global carbon budget. Annu Rev Earth Planet Sci 35:313–347
Hu HF, Wang ZW, Liu GH, Fu BJ (2006) Vegetation carbon storage of major shrubland in china. J Plant Ecol 60:539–544 (in Chinese with English Abstract)
Keller M, Palace M, Hurtt G (2001) Biomass estimation in the Tapajos National Forest, Brazil: examination of sampling and allometric uncertainties. Forest Ecol Manag 154:371–382
Lavorel S, Díaz S, Cornelissen JHC, Garnier E, Harrison SP, McIntyre S, Urcelay C (2007) Plant functional types: are we getting any closer to the Holy Grail? In: Canadell J, Pitelka LF, Pataki D (eds) Terrestrial ecosystems in a changing World. Springer, New York, pp 149–164
Li YN, Zhao L, Wang QX, Du MY, Gu S, Xu SX, Zhang FW, Zhao XQ (2006) Estimation of biomass and annual turnover quantities of Potentilla froticosa shrub. Acta Agrestia Sinica 14:72–76 (in Chinese with English Abstract)
Lin W, Li JS, Zheng BF, Guo JM, Hu LL (2010) Models for estimating biomass of twelve shrub species in Jinggang Mountain nature reserve. J Wuhan Bot Res 28:725–729 (in Chinese with English Abstract)
Liu C, Westman CJ (2009) Biomass in a Norway spruce-Scots pine forest: a comparison of estimation methods. Boreal Environ Res 14:875–888
Liu XL, Liu SR, Su YM, Cai XH, Ma QY (2006) Aboveground biomass of Quercus aquifolioides shrub community and its responses to altitudinal gradients in Balangshan Mountain, Shichuan Province. Cientia Silvae Sinicae 42:1–7 (in Chinese with English Abstract)
Liu J, Liu XL, Hou LM (2011) Changes and ecological vulnerability of landscape pattern in Eastern Qilian Mountains. Arid Land Geogr 35:795–805 (in Chinese with English Abstract)
Liu Z, Chen R, Song Y, Han C (2014) Distribution and estimation of aboveground biomass of alpine shrubs along an altitudinal gradient in a small watershed of the Qilian Mountains, China. J Mt Sci Engl. doi:10.1007/s11629-013-2854-7
Lodhiyal LS, Lodhiyal N (1997) Variation in biomass and net primary productivity in short rotation high density central Himalayan poplar plantations. Forest Ecol Manag 98:167–179
Luo T, Li W, Zhu H (2002) Estimated biomass and productivity of natural vegetation on the Tibetan Plateau. Ecol Appl 12:980–997
Manzoni S, Vico G, Katul G, Philip A, Wayne P, Sari P, Amilcare P (2011) Optimizing stomatal conductance for maximum carbon gain under water stress: a meta-analysis across plant functional types and climates. Funct Ecol 25(3):456–467
Morgan WB, Moss PA (1985) Biomass energy and urbanisation: commercial factors in the production and use of biomass fuels in tropical Africa. Biomass 6:285–299
Murray RB, Jacobson MQ (1982) An evaluation of dimension analysis for predicting shrub biomass. J Range Manag 35:451–454
Myers-Smith IH (2011) Shrub encroachment in arctic and alpine tundra: mechanisms of expansion and ecosystem impacts. Ph.D. thesis, University of Alberta
Návar J, Méndez E, Nájera A, Graciano J, Dale V, Parresol B (2004) Biomass equations for shrub species of Tamaulipan thornscrub of North-eastern Mexico. J Arid Environ 59:657–674
Oyonarte PB, Cerrilo RMN (2003) Aboveground phytomass for major species in shrub ecosystem of western Andalusia. Investigación Agraria: Sistemas y Recursos Forestales 12:47–55
Paton D, Nuñez J, Bao D, Muñoz A (2002) Forage biomass of 22 shrub species from Monfragüe Natural Park (SW Spain) assessed by log-log regression models. J Arid Environ 52:223–231
Peek JM (1970) Relation of canopy area and volume to production of three woody species. Ecology 51:1098–1101
Piao SL, Fang JY, Ciais P (2009) The carbon balance of terrestrial ecosystems in China. Nature 458:1009–1013
Rapp M, Santa Regina I, Rico M, Gallego HA (1999) Biomass, nutrient content, litterfall and nutrient return to the soil in Mediterranean oak forests. Forest Ecol Manag 119:39–49
Sah JP, Ross MS, Koptur S, Snyder JR (2004) Estimating aboveground biomass of broadleaved woody plants in the understory of Florida Keys pine forests. Forest Ecol Manag 203:319–329
Schlaegel B (1981) Testing, reporting, and using biomass estimation models. In: Proceedings of 1981 Southern Forest Biomass Workshop. The Belle W. Baruch Forest Science Institute, Clemson University, SC, USA, pp. 95–112
Seidel D, Fleck S, Leuschner C, Hammett T (2011) Review of ground-based methods to measure the distribution of biomass in forest canopies. Ann For Sci 68(2):225–244
Singh V, Tewari A, Kushwaha SPS, Dadhwalb VK (2011) Formulating allometric equations for estimating biomass and carbon stock in small diameter trees. Forest Ecol Manag 261:1945–1949
Sironen S, Kangas A, Maltamo M, Kangas J (2003) Estimating individual tree growth with nonparametric methods. Can J Forest Res 33:444–449
Spadavecchia L, Williams M, Bell R, Stoy P, Huntley B, van Wijk MT (2008) Topographic controls on the leaf area index and plant functional type of a tundra ecosystem. J Ecol 96(6):1238–1251
Tape KEN, Sturm M, Racine C (2006) The evidence for shrub expansion in northern Alaska and the Pan-Arctic. Global Change Biol 12:686–702
Uso JL, Mateu J, Karjalainen T, Salvador P (1997) Allometric regression equations to determine aerial biomasses of Mediterranean shrubs. Plant Ecol 132:59–69
Ustin SL, Gamon JA (2010) Remote sensing of plant functional types. New Phytol 186(4):795–816
Wang GH, Zhou GS, Yang LM, Li Z (2002) Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slopes of Qilianshan Mountains, Gansu, China. Plant Ecol 165:169–181
Wang P, Xie D, Zhou Y, Youhao E, Zhu Q (2014) Estimation of net primary productivity using a process-based model in Gansu Province, Northwest China. Environ Earth Sci 71(2):647–658
Xiang WH, Liu SH, Deng XW, Shen AH, Lei XD, Tian DL, Zhao MF, Peng CH (2011) General allometric equations and biomass allocation of Pinus massoniana trees on a regional scale in southern China. Ecol Res 26:697–711
Yashiro Y, Shizu Y, Hirota M, Shimono A, Ohtsuka T (2011) The role of shrub (Potentilla fruticosa) on ecosystem CO2 fluxes in an alpine shrub meadow. J Plant Ecol 3:89–97
Zeng HQ, Liu QJ, Feng ZW (2010) Biomass equations for four shrub species in subtropical China. J Veg Res 15:83–90
Zhang ZM, Han TH (2008) A mathematical model for aboveground biomass in the Gannan, Gansu province. Pratacultural Sci 25:10–13 (in Chinese with English Abstract)
Acknowledgments
Thanks to Dr. LaMoreaux and anonymous reviewers for their valuable comments. The National Natural Science Foundation of China (Grant Nos. 91025011 and 91125013) and the National Science Fund for Excellent Youth Scholars of China (Grant No.41222001) supported this research.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Liu, Z., Chen, R., Song, Y. et al. Estimation of aboveground biomass for alpine shrubs in the upper reaches of the Heihe River Basin, Northwestern China. Environ Earth Sci 73, 5513–5521 (2015). https://doi.org/10.1007/s12665-014-3805-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12665-014-3805-5