
Abstract
Increased nitrogen (N) input to ecosystems could alter soil organic carbon (C) dynamics, but the effect still remains uncertain. To better understand the effect of N addition on soil organic C in wetland ecosystems, a field experiment was conducted in a seasonally inundated freshwater marsh, the Sanjiang Plain, Northeast China. In this study, litter production, soil total organic C (TOC) concentration, microbial biomass C (MBC), organic C mineralization, metabolic quotient (qCO2) and mineralization quotient (qmC) in 0–15 cm depth were investigated after four consecutive years of N addition at four rates (CK, 0 g N m−2 year−1; low, 6 g N m−2 year−1; moderate, 12 g N m−2 year−1; high, 24 g N m−2 year−1). Four-year N addition increased litter production, and decreased soil organic C mineralization. In addition, soil TOC concentration and MBC generally increased at low and moderate N addition levels, but declined at high N addition level, whereas soil qCO2 and qmC showed a reverse trend. These results suggest that short-term N addition alters soil organic C dynamics in seasonally inundated freshwater marshes of Northeast China, and the effects vary with N fertilization rates.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Nitrogen (N) is recognized as a key nutrient limiting net primary production in most terrestrial and aquatic ecosystems (Vitousek and Howarth 1991). In past decades, anthropogenic activities have altered global N cycle, and substantially increased the rate of N input into ecosystems at regional and global scales (Vitousek et al. 1997; Galloway et al. 2004), especially in Asia (Galloway et al. 2004). Increased N availability has the potential to enhance plant productivity and biomass accumulation (Vitousek and Howarth 1991), and hence affect ecosystem carbon (C) dynamic and global C cycle (Vitousek et al. 1997).
Increased N input could enhance net primary productivity, leading to increased vegetation C pool in most ecosystems (LeBauer and Treseder 2008; Xia and Wan 2008). However, the effect of N enrichment on soil organic C dynamics is inconsistent and remains remarkable uncertain (Neff et al. 2002; Lu et al. 2011). N fertilization caused an increase in soil organic C in some ecosystems (Turunen et al. 2004; Pregitzer et al. 2008) but a substantial decline in other ecosystems (Mack et al. 2004; Khan et al. 2007; Allison et al. 2010). In addition, some studies observed that increased N input to ecosystems had a negligible effect on soil organic C dynamics (Nadelhoffer et al. 1999; Zeng et al. 2010). Therefore, the response of soil organic C to increased N loading may vary with plant species composition and ecosystem types. Moreover, there is limited information on the effect of N addition on soil organic C in wetlands, although similar studies have been widely conducted in forests, grasslands and agricultural lands (Lu et al. 2011). Wetland ecosystems, especially those in mid-high northern latitudes, store a large amount of C in soils and thus play an important role in global C cycle (Post et al. 1982; Gorham 1991). Considering that wetland soils hold much higher C density than other ecosystem types (Post et al. 1982), a better knowledge of the response of wetland soil organic C to N addition is needed to predict how changes in N availability influence soil organic C dynamics in terrestrial ecosystems.
The Sanjiang Plain, formerly the largest freshwater marsh region in China, has been extensively cultivated for agricultural production in the past 50 years (Huang et al. 2010). Due to fertilizer application during agricultural activities, the still uncultivated marshes in this region often receive increasingly exogenous N input (Zhang et al. 2007a, b). In the previous studies, N addition has promoted plant net primary productivity in freshwater marshes in the Sanjiang Plain, Northeast China (Zhang et al. 2007a; Liu and Song 2008). However, the effect of N enrichment on soil organic C is still unclear, given that soil organic C mainly depends on the balance between plant production and organic matter decomposition. Will N enrichment influence soil organic C in freshwater marsh ecosystems? To answer the question, this study investigated soil total organic C (TOC) concentration in a seasonally inundated freshwater marsh after four consecutive years of N addition. For a better knowledge of soil organic C dynamic under N enrichment, the present study also examined the effect of N addition on soil microbial biomass C (MBC), organic C mineralization, metabolic quotient (qCO2) and mineralization quotient (qmC) in this freshwater marsh ecosystem.
Materials and methods
Study site
The study was conducted in a typical seasonally inundated freshwater marsh near the Sanjiang Experimental Station of Wetland Ecology (47°35′N, 133°31′E), which is located in the Sanjiang Plain, Heilongjiang Province, Northeast China. The station is administered by the Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, and is one of the members of the Chinese Ecological Research Network. Freshwater marsh is the main wetland type in the study site, and Carex lasiocapa and Deyeuxia angustifolia are the dominant species in permanently inundated and seasonally inundated wetlands, respectively. The climate of the study site belongs to the temperate continental monsoon of a seasonal frozen zone, with a mean annual temperature of 2.5 °C, precipitation of 556 mm (about 50 % fall in July and August) and frost-free period of 125 days. The soils of study sites were marsh soil, meadow soil and peat soil, with high soil organic matter content and pH of 5–6.
Experimental design
During autumn 2003, 12 1 m × 1 m plots were established in a D. angustifolia-dominated seasonally inundated freshwater marsh. D. angustifolia-dominated freshwater marsh is one of the main wetland types in the Sanjiang Plain, and occupies approximately 31 % of the wetland area in this region (Zhao 1999). In the study plots, the ground was very flat, and the vegetation was relative uniform. The dominant species in the plots were D. angustifolia and Glyceria spiculosa, which were perennial, cold-temperate mesophytes and accounted for about 90 % of the total aboveground biomass. The soil of the plots was developed from alluvial deposit, and was meadow soil according to the Chinese Classification, or Inceptisols according to the American Classification. Given that all the plots shared similar soil type, microtopography and vegetation, all plots were assumed to have similar soil properties prior to the establishment of this experiment. Before the experiment began, the initial soil TOC concentration, total nitrogen, pH, bulk density and C:N ratio at 0–15 cm layer in the study site were 71.3 mg g−1, 6.4 mg g−1, 5.73, 0.85 g cm−3 and 11.1, respectively.
In autumn 2003, each plot was separated by a 1-m wide buffer strip. Polyvinyl chloride frames (0.5 m depth) were installed to prevent horizontal movement and lateral diffusion of the added N. Board walks giving access to the whole experimental area were installed in 2004 to minimize disturbance on the plots. Four N fertilization levels (CK, 0 g N m−2 year−1; low, 6 g N m−2 year−1; moderate, 12 g N m−2 year−1; high, 24 g N m−2 year−1) were randomly assigned to the 12 plots, and each treatment had three replicate plots. From 2005 to 2008, N was added as ammonium nitrate (NH4NO3) during the growing season. In each year, N fertilizer was divided into nine equal doses and applied biweekly from May to September. Fertilizer was weighed, mixed with 1 L surface marsh water, and applied to each plots. At the same time, the CK plots received 1 L surface marsh water without N fertilizer. Low N addition level was approximately equal to the annual N input to wetlands through leaching and runoff during agricultural activities (5.8 g N m−2) in this region, and moderate and high N addition levels were used to study the response of this ecosystem to N enrichment that may occur in the future (Zhang et al. 2007a).
Sample collection and analyses
At the end of the growing seasons (mid-September) of 2005, 2007 and 2008, litter was collected by using a 0.25 m2 quadrat in each plot, oven-dried to a constant mass at approximately 65 °C for 48 h, and then weighed. After that, plant litter was returned to the corresponding plots.
Soil samples from 0 to 15 cm layer were collected in late July 2008. In each plot, five soil cores (2.5 cm in diameter) were randomly collected following removal of surface litter, and thoroughly mixed to homogenize a sample. After removing the plant roots, fauna and debris by hand, the soil was sieved through a 2-mm mesh screen and divided into two subsamples. One subsample was stored at 4 °C until analysis for soil MBC and organic C mineralization, and the second subsample was air-dried at room temperature around 20 °C, ground and then passed through a 0.25-mm sieve for measurement of soil TOC concentration.
Soil TOC concentration was determined using the K2Cr2O7–H2SO4 wet oxidation method from Walkley and Black (Nelson and Sommers 1996).
Soil MBC was measured by the chloroform fumigation-extraction method of Vance et al. (1987). Briefly, three portions of fresh soil (equivalent to 20 g oven-dried soil) were weighed into 100 mL beakers and fumigated under dark at 25 °C for 24 h with ethanol-free chloroform. After fumigant removal, the soil was extracted with 50 mL 0.5 mol L−1 K2SO4 for 30 min. Three portions of unfumigated soil were extracted simultaneously when fumigation commenced. Organic C in the extracts was measured by the dichromate oxidation method. Soil MBC was calculated by dividing the difference in organic C concentration between the fumigated and unfumigated soils by a correction factor of 0.38 (Vance et al. 1987).
Soil organic C mineralization was measured by the method described in Goberna et al. (2006). Field-moist soil equivalent to 15 g oven-dried soil was placed in 600 mL glass jars and aerobic incubated for 27 days at 25 °C and 60 % water-holding capacity. Additionally, three jars without soil were considered as blanks. After 1, 3, 8, 14, 20 and 27 days of incubation, the evolved CO2 was measured by extracting 10 mL gas from the jars by using a syringe and injecting it into a gas chromatograph (Agilent 4890D, Agilent CO., Santa Clara, CA, USA). And then, the jars were left open for 20 min to allow the air in the flasks to be replenished. In order to maintain at 60 % water-holding capacity throughout the incubation, soil moisture content was checked by weighing the flask every week and adjusted by adding distilled water when necessary. Soil organic C mineralization was calculated as the difference between the CO2 produced from the soil and that from the blank, and expressed as μg CO2–C g−1 soil. Soil qCO2, respiration rate per unit of MBC, was calculated by dividing the mean values of soil organic C mineralization during the entire incubation period by the corresponding MBC. Soil qmC was calculated as the portion of TOC that was mineralized during the entire incubation period (CO2–C(27D)/TOC) (Pinzari et al. 1999).
Statistical analyses
Data were statistically analyzed using procedures of SPSS (v. 13.0), and the accepted significance level was α = 0.05. Data were tested for normality using the Kolmogorov–Smirnov test, and all data were conformed to a normal distribution (data not shown). Fisher’s least significant difference analysis was used to determine significant difference in litter production, soil TOC, MBC, organic C mineralization, qCO2, qmC and MBC/TOC among all treatments. In addition, the effects of N addition on soil TOC, MBC, MBC/TOC were assessed by calculating the N-induced changes (%) = (k N − k C)/k C × 100, where k N is the soil properties in the N addition plots, k C is the soil properties in the CK plots.
Results
In 2005 and 2007, only high N addition treatments had greater litter production than the CK treatments (Fig. 1). However, low, moderate and high N addition treatments had higher litter production than the CK treatments in 2008 (Fig. 1). On average, N addition generally increased litter production (Fig. 1). Moreover, low and moderate N addition treatments had lower litter production than the high N addition treatment (Fig. 1).

Effect of N addition on litter production in freshwater marshes of Northeast China. Values are means and errors bars represent standard deviation (n = 3). Means with different lowercase letters are significantly different at P < 0.05. CK, 0 g N m−2 year−1; Low, 6 g N m−2 year−1; Moderate, 12 g N m−2 year−1; High, 24 g N m−2 year−1
Four-year N addition significantly altered soil TOC concentration, MBC and MBC/TOC, and the effects varied with fertilization rates (Fig. 2). Soil TOC concentration increased by 14 and 10 % at low and moderate N addition levels, respectively, and declined by 24 % at high N addition level (Fig. 2). In addition, low N addition caused a 17 % increase in soil MBC, while high N addition caused a 39 % reduction (Fig. 2). However, soil MBC/TOC decreased only at high N addition level, and did not change at low and moderate N addition levels (Fig. 2).

Effects of N addition on soil total organic carbon (TOC), microbial biomass carbon (MBC) and MBC/TOC in freshwater marshes of Northeast China. Values are means and errors bars represent standard deviation (n = 3). Means with different lowercase letters are significantly different at P < 0.05. CK, 0 g N m−2 year−1; Low, 6 g N m−2 year−1; Moderate, 12 g N m−2 year−1; High, 24 g N m−2 year−1
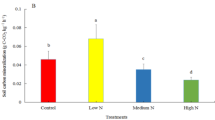
N addition significantly decreased soil organic C mineralization during the entire incubation periods, albeit there were no significant differences in soil organic C mineralization among low, moderate and high N addition levels (Fig. 3). In addition, soil qCO2 and qmC decreased initially with increased N addition level, and subsequently increased (Fig. 4). Soil qCO2 and qmC in both low and moderate N addition treatments were significantly lower than those in the CK and high N addition treatments, respectively (Fig. 4).

Effect of N addition on soil organic C mineralization in freshwater marshes of Northeast China. Values are means and errors bars represent standard deviation (n = 3). CK, 0 g N m−2 year−1; Low, 6 g N m−2 year−1; Moderate, 12 g N m−2 year−1; High, 24 g N m−2 year−1

Effects of N addition on soil metabolic quotient (qCO2) and mineralization quotient (qmC) in freshwater marshes of Northeast China. Values are means and errors bars represent standard deviation (n = 3). Means with different lowercase letters are significantly different at P < 0.05. CK, 0 g N m−2 year−1; Low, 6 g N m−2 year−1; Moderate, 12 g N m−2 year−1; High, 24 g N m−2 year−1
Discussion
In the present study, both low and moderate N addition levels increased soil TOC concentration, whereas high N addition level decreased soil TOC concentration. Indeed, previous studies also found varying responses of soil TOC to N fertilization, including positive (Turunen et al. 2004; Pregitzer et al. 2008), negative (Mack et al. 2004; Khan et al. 2007; Allison et al. 2010), and negligible (Nadelhoffer et al. 1999; Zeng et al. 2010). Nevertheless, the results of the present study clearly demonstrate that the effect of N addition on soil organic C varies with fertilization rates, and suggest that N enrichment alters surface soil organic C in natural wetlands in the Sanjiang Plain, and the effect varies with N loading rates.
Soil TOC depends on the balance between organic C input and output (Mack et al. 2004). In the present study, low and moderate N addition increased plant productivity, leading to increased organic matter inputs to soils, and hence increased soil TOC concentration (Zak et al. 1994). Moreover, N addition may enhance soil organic matter quality (low C/N ratio) (Song et al. 2011) and improve soil microbial utilization efficiency (low qCO2 and qmC) (Fig. 4), and thus decreased soil organic C mineralization (Fig. 3). In addition, the decrease in soil TOC concentration under high N addition level may be related to the following mechanisms. First, high N addition rates may exceed the N-demand of plants, and lead to N-saturation in freshwater marshes (McNulty et al. 1996). Actually, Liu and Song (2008) found no significant difference in aboveground biomass among low, moderate and high N addition treatments. Second, according to the resource optimization hypothesis, increased N availability generally alters plant allocation between shoots and roots, and decreases root biomass (Ågren and Franklin 2003). High N addition level may decrease root biomass and reduce belowground C input to the soil in this freshwater marsh. Third, high N addition level may increase soil dissolved organic C leaching (Treseder 2008). Since soil dissolved organic C is very critical in controlling C accumulation in soils (Bolan et al. 2011), increased dissolved organic C outputs would cause a decline in soil TOC.
In this study, N addition caused a change in soil MBC in freshwater marsh ecosystems, and the effect varied with N addition levels. However, Treseder (2008) reviewed the effect of N enrichment on soil microbial biomass and found that soil microbial biomass declined 15 % on average under N addition. Compared to CK treatment, a greater soil MBC under low N addition level may be due to the increased litter production and thus soil organic C input (Zak et al. 1994). However, greater N loads may lead to the soil acidification by nitrification (Li et al. 2010), N toxicity (Treseder 2008) and reduced belowground C sources (Bowden et al. 2004; Mo et al. 2008). Therefore, soil MBC did not change at moderate N addition level, and even declined at high N addition level, although there were increases in plant litter production. Moreover, the decrease of soil MBC/TOC with increased N addition rates suggests that, increased N inputs declines soil C availability in wetland ecosystems.
N addition decreased soil organic C mineralization, and substantially altered soil qCO2 and qmC in freshwater marshes. Treseder (2008) also observed that N addition could decrease soil CO2 emission due to reduced microbial biomass. Interestingly, N addition decreased soil organic C mineralization at low and moderate N addition levels, although there was no reduction in soil MBC. At low and moderate N addition rates, decreased soil organic C mineralization may be caused by the increased soil microbial C-utilization efficiency (low qCO2 and qmC) (Mao et al. 2010), resulting from the enhanced quality of soil organic matter input (low C/N ratio) (Song et al. 2011). At high N addition rate, reduced soil microbial biomass may be responsible for the decline in soil organic C mineralization (Treseder 2008). In addition, an increase in soil qCO2 may be explained by the increased stress induced by high N addition and decreased C use efficiency for growth by soil microorganisms (Fisk and Fahey 2001).
In conclusion, N addition over 4 years have altered litter production, soil TOC concentration, MBC, organic C mineralization, qCO2 and qmC in freshwater marshes of Northeast China. Both low and moderate N addition increased soil TOC concentration and MBC, and decreased soil qCO2 and qmC, whereas high N addition had reverse effects on soil TOC, MBC and qCO2. Moreover, N addition increased litter production, and decreased soil organic C mineralization, irrespective of N fertilization rates. These results suggest that short-term N addition alters soil organic C dynamics in wetland ecosystems of Northeast China, and the effects vary with N addition rates.
References
Ågren GI, Franklin O (2003) Root:shoot ratios, optimization and nitrogen productivity. Ann Bot 92:795–800
Allison SD, Gartner TB, Mack MC, McGuire KL, Treseder KK (2010) Nitrogen alters carbon dynamics during early succession in boreal forest. Soil Biol Biochem 42:1157–1164
Bolan NS, Adriano DC, Kunhikrishnan A, James T, McDowell R, Senesi N (2011) Dissolved organic matter: biogeochemistry, dynamics and agro-environmental significance in soils. Adv Agron 110:1–75
Bowden RD, Davidson E, Savage K, Arabia C, Steudler P (2004) Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard forest. Forest Ecol Manag 196:43–56
Fisk MC, Fahey TJ (2001) Microbial biomass and nitrogen cycling responses to fertilization and litter removal in young Northern hardwood forests. Biogeochemistry 53:201–223
Galloway JN, Dentener FJ, Capone DG, Boyer EW, Howarth RW, Seitzinger SP, Asner GP, Cleveland CC, Green PA, Holland EA, Karl DM, Michaels AF, Porter JH, Townsend AR, Vorosmarty CJ (2004) Nitrogen cycles: past, present, and future. Biogeochemistry 70:153–226
Goberna M, Sanchez J, Pascual JA, Garcia C (2006) Surface and subsurface organic carbon, microbial biomass and activity in a forest soil sequence. Soil Biol Biochem 38:2233–2243
Gorham E (1991) Northern peatlands: role in the carbon cycle and probable responses to climatic warming. Ecol Appl 1:181–195
Huang Y, Sun WJ, Zhang W, Yu YQ, Su YH, Song CC (2010) Marshland conversion to cropland in northeast China from 1950 to 2000 reduced the greenhouse effect. Global Change Biol 16:680–695
Khan SA, Mulvaney RL, Ellsworth TR, Boast CW (2007) The myth of nitrogen fertilization for soil carbon sequestration. J Environ Qual 36:1821–1832
LeBauer DS, Treseder KK (2008) Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 89:371–379
Li LJ, Zeng DH, Yu ZY, Fan ZP, Mao R (2010) Soil microbial properties under N and P additions in a semi-arid, sandy grassland. Biol Fert Soils 46:653–658
Liu DY, Song CC (2008) Responses of marsh wetland plant Calamagrostis angustifolia to exogenous nitrogen input. Chin J Appl Ecol 19(12):2599–2604 (in Chinese)
Lu M, Zhou X, Luo Y, Yang Y, Fang C, Chen J, Li B (2011) Minor stimulation of soil carbon storage by nitrogen addition: a meta-analysis. Agr Ecosyst Environ 140:234–244
Mack MC, Schuur EAG, Bret-Harte MS, Shaver GR, Chapin FS (2004) Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization. Nature 431:440–443
Mao R, Zeng DH, Ai GY, Yang D, Li LJ, Liu YX (2010) Soil microbiological and chemical effects of a nitrogen-fixing shrub in poplar plantations in semi-arid region of Northeast China. Eur J Soil Biol 46:325–329
McNulty SG, Aber JD, Newman SD (1996) Nitrogen saturation in a high elevation New England spruce-fir stand. Forest Ecol Manag 84:109–121
Mo J, Zhang W, Zhu W, Gundersen P, Fang Y, Li D, Wang H (2008) Nitrogen addition reduces soil respiration in a mature tropical forest in southern China. Global Change Biol 14:403–412
Nadelhoffer KJ, Emmett BA, Gundersen P, Kjonaas OJ, Koopmans CJ, Schleppi P, Tietema A, Wright RF (1999) Nitrogen deposition makes a minor contribution to carbon sequestration in temperate forests. Nature 398:145–148
Neff JC, Townsend AR, Gleixner G, Lehman SJ, Turnbull J, Bowman WD (2002) Variable effects of nitrogen additions on the stability and turnover of soil carbon. Nature 419:915–917
Nelson DW, Sommers LE (1996) Total carbon, organic carbon, and organic matter. In: Sparks DL, Page AL, Helmke PA, Loeppert RH, Soltanpour PN, Tabatabai MA, Johnston CT, Sumner ME (eds) Methods of soil analysis. Part 3. Chemical methods. Soil Science Society of America Book Series, Number 5. Wisconsin, USA, pp 961–1010
Pinzari F, Trinchera A, Benedetti A, Sequi P (1999) Use of biochemical indices in the Mediterranean environment: comparison among soils under different forest vegetation. J Microbiol Meth 36:21–28
Post WM, Emanuel WR, Zinke PJ, Stangenberger AG (1982) Soil carbon pool and world life zones. Nature 298:156–159
Pregitzer KS, Burton AJ, Zak DR, Talhelm AF (2008) Simulated chronic nitrogen deposition increases carbon storage in Northern Temperate forests. Global Change Biol 14:142–153
Song C, Liu D, Yang G, Song Y, Mao R (2011) Effect of nitrogen addition on decomposition of Calamagrostis angustifolia litters from freshwater marshes of Northeast China. Ecol Eng 37:1578–1582
Treseder KK (2008) Nitrogen additions and microbial biomass: a meta-analysis of ecosystem studies. Ecol Lett 11:1111–1120
Turunen J, Roulet NT, Moore TR (2004) Nitrogen deposition and increased carbon accumulation in ombrotrophic peatlands in eastern Canada. Global Biogeochem Cycles 18, GB3002. doi:10.1029/2003GB002154
Vance ED, Brookes PC, Jenkinson DS (1987) An extraction method for measuring soil microbial biomass C. Soil Biol Biochem 19:703–707
Vitousek PM, Howarth RW (1991) Nitrogen limitation on land and in the sea: how can it occur? Biogeochemistry 13:87–115
Vitousek PM, Aber JD, Howarth RW, Likens GE, Matson PA, Schindler DW, Schlesinger WH, Tilman DG (1997) Human alteration of the global nitrogen cycle: sources and consequences. Ecol Appl 7:737–750
Xia J, Wan S (2008) Global response patterns of terrestrial plant species to nitrogen addition. New Phytol 179:428–439
Zak DR, Tilman D, Parmenter RR, Rice CW, Fisher FM, Vose J, Milchunas D, Martin CW (1994) Plant production and soil microorganisms in late-successional ecosystems: a continental-scale study. Ecology 75:2333–2347
Zeng DH, Li LJ, Fahey TJ, Yu ZY, Fan ZP, Chen FS (2010) Effects of nitrogen addition on vegetation and ecosystem carbon in a semi-arid grassland. Biogeochemistry 98:185–193
Zhang L, Song C, Wang D, Wang Y (2007a) Effects of exogenous nitrogen on freshwater marsh plant growth and N2O fluxes in Sanjiang Plain, Northeast China. Atmos Environ 41:1080–1090
Zhang L, Song C, Zheng X, Wang D, Wang Y (2007b) Effects of nitrogen on the ecosystem respiration, CH4 and N2O emissions to the atmosphere from the freshwater marshes in northeast China. Environ Geol 52:529–539
Zhao KY (1999) Mires in China. Science Press, Beijing (in Chinese)
Acknowledgments
This work was supported by “Strategic Priority Research Program-Climate Change: Carbon Budget and Related Issues” of the Chinese Academy of Sciences (No. XDA05050508), National Natural Science Foundation of China (Nos. 40930527 and 41125001) and National Key Basic Research and Development projects of China (No. 2009CB421103). The authors thank James W. LaMoreaux and two anonymous reviewers for their suggestions.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Song, C., Liu, D., Song, Y. et al. Effect of nitrogen addition on soil organic carbon in freshwater marsh of Northeast China. Environ Earth Sci 70, 1653–1659 (2013). https://doi.org/10.1007/s12665-013-2252-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12665-013-2252-z