
Abstract
Ecological restoration of the Wangaloa coal mine in southern New Zealand is hindered by a range of geoecological factors. The site has some substrate acidification (down to pH 1) and acid rock drainage with discharge waters initially down to pH 4, although this has since risen to ca. pH 6. Surface and ground waters develop elevated sulfate (up to 700 mg/kg) during oxidation of pyrite in coal and quartz in waste rock. Coal has elevated boron content (up to 450 mg/kg) and surface waters on coal-rich waste rock have up to 6 mg/L dissolved boron. Evaporation causes formation of salt encrustations dominated by gypsum with minor boron salts. Boron is bioavailable and may be at toxic levels (>200 mg/kg) in some plants. Quartz-rich waste rock is readily eroded, and develops a cm-scale low-nutrient quartz pebble armouring layer with low water retention properties. All waste rocks including loess siltstone have low nutrient contents, and low moisture retention properties, that are barely sufficient for plant establishment. Native plants introduced to the site during rehabilitation have grown on loess substrate (up to fivefold increase in height over 3 years), with poor or no growth on coal-rich and quartz-rich substrates. In contrast, natural colonisation of manuka (Leptospermum scoparium) has been most effective at revegetation on even the most hostile substrates. This natural revegetation has been facilitated by islands of manuka established accidentally during 60 years of mining history. Manuka from local genetic stock is most viable for this revegetation, and introduced manuka seedlings have had a 70% mortality rate. Natural plant colonisation is the key step in overall ecosystem recovery, and invertebrates rapidly colonise beneath new shrubs irrespective of the nature of the substrate from vegetation islands that have high invertebrate numbers and species richness.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Mining inevitably causes disturbances of the land surface and associated biota. In the past, most mine sites were abandoned, and natural rehabilitation processes were relied upon to develop new ecosystems on the disturbed land (Roberts et al. 1981; Bradshaw 1983; Russell and La Roi 1986; Bell 2001; Prach and Pyšek 2001; Wanner and Dunger 2002). All disturbed land will develop new and sustainable ecosystems eventually, but considerable erosion and dispersal of mine site materials may take place before that occurs (Munshower 1994; Wong 2003; Pensa et al. 2004). In addition, nearby human populations are becoming progressively less tolerant of long-term disturbed sites in their midst. Hence, rapid development of new ecosystems on mine sites is now a high priority for any mining venture, and commonly a regulatory condition imposed before mining starts (Bell 2001; Prach and Pyšek 2001; Pensa et al. 2004). Nowadays, planning of the rehabilitation stage is part of the overall mine plan before the mine opens and simple steps, such as stockpiling topsoil for subsequent plant establishment, can facilitate rapid ecosystem restoration (Carroll et al. 2000; Holmes 2001; Bell 2001; Pensa et al. 2004). Planning of the rehabilitation activity can be considerably enhanced by a close knowledge of the natural processes of rehabilitation that occur on disturbed lands, because the natural processes are normally the most efficient and effective for a particular site (Russell and La Roi 1986; Holmes 2001; Bell 2001). This study examines natural ecosystem recovery in the context of subsequent and related anthropogenic restoration activity, all on the same mine site.
The nature and speed of natural ecosystem recovery depends on the physical and chemical constraints imposed by the geology of the mine site. Sites may be affected by a combination of many factors, such as low-nutrient rocks, elevated concentrations of metals, the presence of mine-related toxins, steep and unstable slopes, and harsh climatic conditions (Russell and La Roi 1986; Mench et al. 2003; Neel et al. 2003; Widerlund et al. 2004; Craw et al. 2006; Mains et al. 2006). In addition, natural ecosystem recovery depends on availability of suitable colonising species that can contribute to the required biological succession (Roberts et al. 1981; Russell and La Roi 1986; Prach and Pyšek 2001; Wanner and Dunger 2002). There is therefore a complex set of inter-relationships between geology and ecology that arises during mine site rehabilitation and these control the success and rate of new ecosystem development (Roberts et al. 1981; Davis et al. 1997; Wanner and Dunger 2002; Craw et al. 2006). Knowledge of these geoecological inter-relationships is key in developing a management strategy for rapid and successful anthropogenic mine-site rehabilitation.
This study describes a wide range of geological site factors that contribute to, or restrict, ecosystem recovery at an abandoned coal mine site. Most of the ecosystem recovery was natural, and site rehabilitation has been interrupted and locally enhanced by recent anthropogenic activity facilitating accelerating ecosystem recovery. A key aim of this paper is to identify and compare the different geological factors at the site that affect ecosystem recovery, so that the relative significance of these various factors can be quantified. In-depth accounts of the nature of several individual geoecological factors at this site have been published previously (Craw et al. 2006, 2007; Begbie et al. 2007), but this paper is the first attempt at linking and integrating all these factors. Knowledge of the inter-relationships and relative significance of the various geoecological issues at a mine site is important for management of anthropogenic site restoration, so that key issues can be addressed rapidly and resources are not wasted on less important or irrelevant issues.
General setting
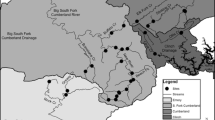
The coal mine site used for this study (Fig. 1) is at Wangaloa in southern New Zealand (169°54′E; 46°17′S), 3 km from the east coast (Pacific Ocean). The site occurs in rolling hills with locally steep relief (ca. 120 m), with a cool temperate maritime climate. Rainfall is 700–1,000 mm/year, and mean annual temperature is ca. 12°C. Rainfall is distributed evenly through the year, but periodic dry spells occur for up to a month at a time, especially in summer. These dry spells result in abundant evaporational drying of bare land surface and formation of mineral efflorescences (Begbie et al. 2007).

Maps of the Wangaloa mine site, showing geoecological features relevant to the rehabilitation programme. a Hydrogeology map, showing the main surface and ground water features and drainage directions, and the wetlands, and main areas of waste rock within the drainage system. b Paste pH results for 229 substrate points over the mine site, measured in 2003. c Principal vegetation cover in and around the site, with the main patches of naturally regenerated native shrubs indicated. Dashed box shows the location of the eastern portion of Fig. 3
The mine was developed in the Late Cretaceous Taratu Formation, which consists mainly of conglomerate with minor sandstone, siltstone and mudstone, and numerous coal seams ca. 10-m thick (Harrington 1958). The Taratu Formation at Wangaloa has been extensively eroded over the past 5 million years, as the area lies between two active faults that are causing localised uplift. The mined coal seam at Wangaloa is in the hinge of a shallow northeast plunging anticline (Fig. 1a). Low sea levels during the last glacial period (ca. 14 ky) exposed glacial silt from the nearby Clutha River near the shoreline, and this was blown onshore to form a blanket of silt (loess) over the topography (Bruce 1973). This loess veneer is up to 5-m thick, especially on leeward slopes and where local reworking has concentrated it into colluvial channels incised into the underlying Taratu Formation. Loess consists of quartz, albite, muscovite and chlorite derived from the Otago Schist basement inland from Wangaloa. Coal exposed at the surface went on fire at several localities, probably because of spontaneous combustion of piles of eroded fine coal at outcrops. Remnants of resultant baked sediments and paralavas are widespread in the Wangaloa area, and form a significant part of the waste rock locally.
Loess and other surficial materials were removed as overburden when the mine was opened in the 1940s, and deposited in waste dumps on site. As the mine developed, most waste rock at the mine site consisted of Taratu Formation conglomerate made up of quartz pebbles (up to 2 cm) with a matrix of finer-grained quartz, muscovite, and kaolinite (Fig. 2a). This material was deposited on top of the loess in places, and waste dumps rich in quartz pebbles now cover several hectares (Fig. 1a, b). Coal in the main seam at Wangaloa is rich in sulfur (typically 4–5 wt%) that is partly dispersed through the organic matter (Suggate 1959), and is partly present as pyrite (Begbie et al. 2007). Pyrite also cements quartz conglomerate immediately above the coal seam (Fig. 2b). Pyrite oxidation products include gypsum, jarosite, and iron oxyhydroxide, and these commonly occur as surface coatings on coal fragments and associated quartz pebbles in the vadose zone of waste rock piles. Unsaleable high-ash coal was also deposited in waste dumps with some admixed quartz pebbles (Fig. 2c), and mineral efflorescences are common on this waste rock.

Photographs of the Wangaloa site, showing some key features of geoecological significance. a Poorly vegetated quartz-rich substrates (white, centre) that have not been recontoured in the Eastern waste rock pile (Fig. 1a), showing erosion rills. Wetland in foreground has tall (4 m) manuka that has colonised from a forest remnant nearby, and slopes in the background have loess-rich substrate that has been cleared of weeds and planted in native shrubs. b Quartz-rich waste rock on the acid slope (Fig. 1a), with a cement of pyrite in one piece (centre). Acid from oxidation of this pyrite has killed grass in the immediate surrounds. c Coal-rich waste rock with scattered quartz pebbles, forming a substrate hostile to plant growth. Native plantings have little growth or have died, but colonisation of manuka from a nearby bush patch (rear) is occurring. d Healthy native plantings (Hebe spp. and Griselinia littoralis) on loess substrate, with adventive grasses suppressed by herbicide
Mining activity created wetlands on the site, as waste rock dumps grew progressively across the natural drainage system (Fig. 1a). The largest of these, the upper wetland and southern wetland (Fig. 1a) have formed behind quartz-rich waste rock piles. The lower wetland lies at the foot of a major waste rock pile (eastern overburden; Fig. 1a) and this wetland formed behind a natural landslide downstream of the site.
The mine was closed in 1989, leaving a depression partially filled with waste rock piles, and cliffs (high walls) of quartz conglomerate to the north and south (Fig. 1a). Low points in the site became lakes, with the lowest part of the mine forming the largest lake (Main Lake; Fig. 1a) that discharges excess water to a wetland (Lower wetland; Fig. 1a). By that time, some of the waste rock dumps had become partially revegetated by natural colonisation (Figs. 2a, c, 3; Craw et al. 2007). In addition, in preparation for closure some of the site and surrounds were planted as pine plantation forest (Pinus radiata; Fig. 1c) in the 1980s. Pine trees on the waste rock piles became stunted and poorly formed, and some died, over the following 10 years. Minor acid rock drainage had developed by 2000, resulting in the Main Lake having water with pH between 4 and 5 (Black and Craw 2001).

Map of natural revegetation by predominantly manuka colonisation on a range of waste rock substrates at the western edge of the mine site (Fig. 1c)
A more concerted effort for rehabilitation of the site began in 2000, with the aim of re-establishment of native vegetation to form a recreational area. Pine trees were removed, waste rock was partially recontoured, and adventive weedy shrubs (mainly gorse, Ulex europaeus) were removed (Fig. 2a). Patches of naturally regenerated native bush were retained in this process (Figs. 1c, 3). More than 70,000 New Zealand native plants were emplaced over the unvegetated parts of the site in individual peat-rich blocks containing slow-release fertilizer, beginning in 2003 (Fig. 2c, d). Weeds were controlled with herbicides where necessary, to limit competition with the plantings (Fig. 2d).
Methods
Observations and field and laboratory data were collected for a wide range of geoecological factors at the site between 2002 and 2007, in order to evaluate the success of the anthropogenic rehabilitation, and to compare these rehabilitation results to natural regeneration of ecosystems that had been occurring on the site beginning in the 1940s. Details of methods for most of the data collection and analysis have been published previously (Craw et al. 2006; Rufaut et al. 2006; Begbie et al. 2007), and a summary is provided here.
Water samples from a range of streams, lakes, boreholes, and wetlands (Fig. 1a) were collected for major ion analysis and limited trace metal suites (Begbie et al. 2007). Dissolved sulfate was analysed via ion chromatography, and alkalinity was determined by potentiometric titration. Chloride analysis was done with a FOSS Tecator Fluid-Injection Analyser (FIAstar 5000). Cations were analysed using a Varian SpectrAA 220 FS atomic absorption spectrophotometer. Detection limits for cations and sulfate were 1 mg/L, for alkalinity was 1.2 mg/L, and for chloride was 0.06 mg/L. Samples for trace elements analysis were filtered on site (0.45 μm) and collected in acid-washed plastic bottles with nitric acid added in the field for preservation. Total dissolved B analyses were done by ICP-MS, with detection limit of 0.005 mg/L. Copper, As, Pb, and Zn analyses were obtained by flame atomic absorption spectroscopy (Black and Craw 2001). Water pH was determined in the field using a portable Oakton meter calibrated with standard pH solutions.
Samples of substrates were taken mainly from the upper 30 cm, and there are negligible chemical differences between surface samples and deeper (>50 cm) samples in the same substrate type. Paste pH of substrates over the site (Fig. 1b) was obtained from distilled water slurries of substrates over the disturbed part of the site (Fig. 1b). Slurries and measurements were made in the field by the method of Sobek et al. (1978). Total sulfur (S) contents of coals were determined using a Carlo Erba Elemental Analyser EA 1108, and total B contents were determined by Eschka fusion followed by ICP-MS analysis (Craw et al. 2006). Total N, Olsen P, major cations, CEC, organic matter, and pH were determined by standard soil methods (Blakemore et al. 1987). Substrate moisture content was estimated from fresh samples that were weighed, dried at 105°C for 16 h, and then re-weighed to calculate a moisture factor.
Invertebrate diversity on vegetated, unvegetated and rehabilitated surfaces was determined from random collections of substrate cores (7 cm width; 0–5 cm depth), which included leaf litter. Invertebrates were extracted from sampling cores over 48 h under a 60 W lamp bulb positioned 20 cm above the core (Rufaut et al. 2006). Extracted invertebrates were collected live on plastic discs floating on top of water in collection jars. Substrate invertebrates were separated into different orders and counted. Beetles were examined in more detail on naturally revegetated hillslopes to obtain an idea of the types of species that may colonise adjacent areas of rehabilitated waste rock (Fig. 1c). Beetle species diversity and abundance were determined on litter samples collected from beneath vegetation dominated by native trees, mostly manuka (Leptospermum scoparium) and/or kanuka (Kunzea ericoides), in differing sized patches that remained after mining and rehabilitation earthworks were completed (Fig. 1c). The beetles were removed from the litter in heat extraction funnels over 48 h, using 150 W light bulbs positioned 40 cm above the litter.
Foliage samples from a range of plants introduced during the anthropogenic rehabilitation in 2003 were sampled in 2004 after 1 year of growth. These samples were washed to remove dust, oven dried, powdered with a steel grinder, digested with nitric and hydrochloric acids, and analysed for P, B, Zn and Cu by ICP-MS (Craw et al. 2006). Growth success of a selection of introduced plants was quantified by observations of mortality, and measurement of heights of surviving plants in 2003 and again in 2006 (Todd et al. 2009).
Results
Acid rock drainage
Most substrates exposed during the site rehabilitation have moderately acid paste pH over the site (pH 3.6–5.4; Fig. 1b). This range is more acid than background soils developed on Taratu Formation rocks elsewhere in the region, which are typically weakly acid (pH 5–5.5). In addition, some substrates have more acid paste pH, ranging from 3.6 down to 1 (Fig. 1b). Coal-bearing waste rock dumps are typically strongly acidic, with pH near 3. The most acid substrates developed after the 2003 paste pH survey (Fig. 1b), on a slope immediately north of the Upper wetland (Fig. 1a). Recontouring of this slope as part of the rehabilitation operation exposed quartz-rich waste rock with a cement of pyrite (Fig. 2b). Sulfuric acid from oxidation of that pyrite acidified surrounding substrate on the cm scale (Fig. 2b), and rainwater runoff spread this acid downslope so that within a year, the whole lower part of the slope had pH near 1 (Begbie et al. 2007).
The sulfur-bearing substrates on the site cause acidification of the surface and ground waters over the whole site, although acidification is less intense than for the acid slope described above. This water–rock interaction shows up in major ion concentrations of waters, particularly with the proportion of dissolved sulfate (Fig. 4). Background waters are dominated by chloride (Figs. 4a, 5a) from marine aerosols in rain (Craw and Beckett 2004). Most of these waters are dilute (Cl− < 50 mg/L), but some incoming groundwater has Cl− near 100 mg/L (Fig. 5a). This background chloride is progressively overwhelmed by dissolved sulfate when the waters come in contact with the various substrates on the mine site, and dissolved sulfate ranges up to 700 mg/L (Figs. 4b, 5b). These waters eventually make their way to lakes on the site (Fig. 1a), and lake waters are dominated by dissolved sulfate (Fig. 4c). However, the lake waters have only moderate concentrations of sulfate and chloride because these waters are a mixture of all site waters (Figs. 4c, 5a, b). The substrates contain negligible carbonate minerals, so the water–rock interaction does not yield significant dissolved carbonate in any site waters (Fig. 4b, c).

Ternary diagrams showing relative proportions of principal dissolved ions in surface and ground waters and lakes at the Wangaloa site. Incoming water is dominated by chloride, and most site waters show varying degrees of sulfate dissolution from substrate acidification (Fig. 1b)

Geoecological features of water chemistry at the Wangaloa site. a Variations in dissolved chloride and pH. b Variations in dissolved sulfate and dissolved boron
Despite the dominance of dissolved sulfate, not all the site waters are acid, and there is a wide range of surface and ground water pH, from 3 to 7 (Fig. 5a). Main lake water had pH near 4 at the start of the rehabilitation programme, and that rose erratically to near 6 by 2006 (Begbie et al. 2007). The rise in lake pH was assisted by addition of hydrated lime in 2002, and the higher pH had stabilized by 2004 even though water residence time in the lake is only 2 years (Begbie et al. 2007).
Elevated boron concentrations
The coal extracted from the main seam in the Wangaloa mine has elevated boron content (up to 450 mg/kg; Craw et al. 2008) although B concentrations are variable (Fig. 6a). Likewise, coal fragments in waste rock piles at the site also have elevated B contents (Fig. 6a). These elevated B contents are typical of coals formed in a marginal marine setting (Goodarzi and Swaine 1994; Craw et al. 2006), and accompany the high sulfur contents that also arise in a marginal marine setting (Fig. 6a). Weathering of coal fragments in the waste rock piles liberates both sulfur and boron, and these elements are locally concentrated by evaporation to even higher levels as gypsum-rich efflorescences on coal fragments and adjacent waste rock particles. Boron occurs as boric acid and/or colemanite (Ca2B6O11·5H2O) in these efflorescences (Craw et al. 2006).

Geoecologcal features of solids and plants at Wangaloa site. a Concentrations of sulfur and boron in the coal seam (Fig. 1a) and in coal-rich waste rock substrates in which plants are being established. b Phosphorus and boron contents of plants growing on the three principal substrate types
Boron in coal and associated efflorescences on waste rock piles is readily mobilised into solution during rain events, and ephemeral ponds have 3–6 mg/L dissolved boron. The locally high levels of boron at the surface of waste rock piles are diluted in streams and groundwater (Fig. 5b). In addition, adsorption of boron to iron oxyhydroxide precipitates formed from oxidation of pyrite in the waste rock results in further attenuation of boron (Craw et al. 2006). Consequently, lakes on the site which receive surface and ground waters have dissolved B near 1 mg/L (Fig. 5b).
The elevated levels of boron in the surficial zone of the coal-bearing waste rock are transferred to the plants growing on that waste rock. These plants have B contents in their foliage up to 200 mg/kg dry weight, distinctly higher than the same species growing on loess and quartz-rich substrate (Fig. 6b). The distinction between quartz-rich and coal-rich substrates is not clear-cut, as coal fragments occur in both types, to some degree. Hence, there are boron sources in some quartz-rich waste rock, and elevated boron contents arise in some plants on quartz-rich waste rock as well (Fig. 6b).
Nutrients
The waste rock and adjacent mine site substrates are essentially variably disaggregated rocks that have been highly disturbed by mining activity (to 1989) and subsequent recontouring (2002), so negligible soil development has occurred in these substrates. Nutrient availability is therefore limited to those directly released from the rocks. However, leaf litter accumulation has occurred beneath natural revegetation patches, which facilitates additional ecosystem development (Rufaut et al. 2006; Craw et al. 2007). Apart from this litter, plants are becoming established in essentially barren rock material, and traditional soil nutrient parameters are not strictly applicable to this material. Nevertheless, a general indication of nutrient status of the substrates can be obtained with these traditional soil parameters, and some data are presented here in comparison to primitive soils in the fully vegetated large patch of regenerating native forest on the north side of the site (Fig. 1a). Available nutrient levels in all substrates on the site are generally low but adequate for growth of vegetation. The loess and quartz-rich substrates contain negligible organic matter. Coal, by definition, has a high organic content, but the availability of this organic matter for plant growth is poorly known (Vetterlein et al. 1994).
Loess contains the most nitrogen, and although these levels are variable, they commonly exceed the total N levels in the regenerating forest (Fig. 7a). In contrast, most quartz-rich and coal-rich substrates have low total N, although a few coal samples have anomalously high total N (Fig. 7a). The N in loess and quartz-rich substrates is located predominantly in muscovite (Pitcairn et al. 2006), whereas the N in coal-rich substrates is contained in the organic component of coal (up to 0.8 wt% N; Suggate 1959). The availability of these geological N sources to plants is poorly known, but the natural revegetation processes occurring at the site demonstrate that some of this N is being released.

Nutrient status of the three principal substrate types at Wangaloa mine site, compared to a fully regenerated forest patch on the margin of the site. a Total nitrogen and Olsen phosphorus. b Cation exchange capacity, and percentage of moisture present in substrate samples
Plant-available phosphorus, measured as Olsen P, is similar in all substrates, and similar to that of the regenerating native forest (Fig. 7a). Quartz-rich waste rock has the lowest Olsen P (Fig. 7a), but this is largely a result of dilution of sample material by inert quartz. The matrix material in the quartz-rich substrates, in which plants extend their roots for nutrient uptake, has similar Olsen P to the other substrates when analysed without quartz pebble dilution (Craw et al. 2007). Accessory apatite is the main source of P in the quartz-rich and loess substrates, and some P is dispersed through the organic matter in coal, to locally high levels (up to 120 mg/kg; Fig. 7a). There is negligible difference in P contents of plants, introduced during rehabilitation of the site, on the three different substrates (Fig. 6b). These plants were introduced with P-bearing fertilizer in 2003 and the effects of minor differences in P contents of the substrates may not have been fully apparent a year later, although differences in substrate boron contents were apparent in plants at that time (Fig. 6b).
Cation exchange capacity (CEC) of the substrates is similar to that of the developing soil in the regenerating native forest, although the waste rocks display considerable variability (Fig. 7b). Potassium and sodium from weathering of muscovite and albite are the most abundant cations available in the loess and quartz-rich substrates. The apparently low CEC of many quartz-rich substrate samples (Fig. 7b) arises because of dilution by quartz pebbles (see above), and the matrix of this material commonly has high CEC. Likewise, some high-ash coal fragments in the coal-rich waste rock piles have abundant muscovite and clays, and high CEC (Fig. 7b). Dissolution of gypsum, formed during weathering of coal, releases Ca from coal-rich substrates.
The high degree of anthropogenic disturbance of the mine site resulted in open textures and high porosities and permeabilities in all substrates. These physical effects, combined with the negligible organic matter in the soil and low clay contents, has resulted in substrates with poor water retention properties (Fig. 7b). This has been exacerbated by the lack of vegetation cover, so that the substrate surfaces dry rapidly after rain events, and this drying process extends to at least a metre in low-rainfall seasons. Loess has higher clay content than the other two waste rocks, and consequently has the highest measured moisture contents of the site substrates (Fig. 7b). The measured moisture contents of all three substrate types is distinctly lower than those of soils in the regenerating native forest (Fig. 7b).
Revegetation
Natural regeneration of vegetation on the site has been most effective on loess substrate. The first waste rock piles constructed during initial mine development in the 1940s were dominated by the loess that capped part of the original topography. These waste rock piles have been almost fully revegetated by natural colonisation, mainly with native species (Rufaut et al. 2006; Craw et al. 2007). Likewise, disturbed parts of the site that have not been mined and still retain their loess cap have undergone natural revegetation, albeit mainly with exotic weed species. In contrast, natural revegetation of quartz-rich and coal-rich waste rock has been less successful, and introduced native plants have grown more poorly than those on loess substrate (Fig. 8). Natural revegetation of the different waste rock types by native plants is dominated by colonisation by manuka (L. scoparium), and the following descriptions focus on this species. Other species that accompany, and locally succeed, manuka are indicated in Table 1.

Comparison of height increase in individuals of planted shrubs on different substrates, with data extracted from Todd et al. (2009). Shrubs were measured in 2003 and again in 2006. Line indicates where no growth has occurred. Dead plants are excluded
Quartz-rich waste rock piles are more readily eroded than loess, and manuka colonisation has been poor on steep slopes (Fig. 2a). Erosion of this type of waste rock leads to preferential removal of fine matrix material, leaving a substrate with an armouring layer of quartz pebbles up to 5-cm thick. This high porosity, low nutrient layer prevents establishment of seedlings. Erosional rills in the waste rock piles locally accumulate finer grained material with more available nutrients, and manuka can establish in these rills (Fig. 2a). However, this establishment can be temporary, as large rain events cause erosion and/or burial of the colonising plants. The flat tops of quartz-rich waste rock piles, from which the waste was dumped, have been less prone to erosional processes, and these have become more effectively colonised where ongoing human disturbance (e.g., roads) was minor.
All the above colonisation-inhibiting processes can be mitigated by the presence of admixed loess in the surficial zone of quartz-rich waste rock. Lumps of loess-rich material as small as 50 cm across can act as a nucleus for manuka colonisation on even the steepest slopes in quartz-rich waste rock. Litter from shrubs growing on a patch of loess-rich substrate helps to bind the surface zone of adjacent quartz-rich substrate. This litter also facilitates new colonisation by manuka, or colonisation by moss in the shade of established plants, followed by new manuka establishment. These mechanisms have facilitated manuka colonisation of sloping surfaces in the Eastern waste rock piles (Fig. 1a; Craw et al. 2007) and on the western edge of the site (Fig. 3).
Most naturally regenerated manuka was avoided during site recontouring in 2002, and this vegetation now form patches of shrubs of varying height (Fig. 3). These patches are surrounded by, or adjacent to, areas of waste rock that have lesser degrees of natural regeneration or were completely devegetated during the rehabilitation process (Fig. 3). Pine trees planted in the initial stages of mine closure (1980s) also inhibited natural regeneration and growth of colonising manuka. With removal of these pine trees, manuka that was present grew rapidly to enhance the integrity of native shrub patches (Fig. 3).
Despite the widespread natural colonisation by manuka on the site in general, large areas of bare waste rock persisted to, and beyond, the time of site rehabilitation (2002). These areas are underlain by quartz-rich and coal-rich waste rock (e.g. Fig. 2c). Most areas that have been bare for several decades have some scattered manuka shrubs (Fig. 3), commonly as small individuals or patches centred on fine grained material washed into shallow rills. Even after rehabilitation and the extensive native plantings, many of these areas remain bare, a high proportion of introduced native plants have died, and the surviving introduced native plants have shown poor growth (Fig. 8). It is notable that >70% of introduced manuka plants have died since 2003 (Todd et al. 2009). However, manuka seedlings are actively colonising these bare substrates around, and up to 100 m from, the patches of naturally regenerating manuka that have been developing since the 1940s (Figs. 2c, 3). This colonisation began immediately after site recontouring in 2002, but accelerated after 2005. In places, this natural colonisation of manuka is crowding, and locally out-competing, the introduced native plantings.
Invertebrate colonisation
Invertebrate assemblages on 40- to 50-year-old naturally revegetated loess were more diverse than on loess-rich and coal-rich waste rock, respectively. Invertebrates were less diverse on naturally revegetated quartz-rich waste rock compared to loess-rich waste rock (Fig. 9) because of a sparse plant cover (see above). Unvegetated quartz-rich waste rock areas, sampled up to 4 m from the nearest vegetation, had few invertebrates (Fig. 9), and some samples contained no invertebrates. By comparison numbers on unvegetated loess-rich waste rock were more than ten times higher (Fig. 9), but no sampling sites were more than 50 cm from vegetation. Invertebrate diversity was low on quartz-rich waste rock recently rehabilitated with plantings. However, numbers of individuals were comparable to naturally revegetated substrates, and showed rapid colonisation by Collembola specifically. Although the abundance of Collembola spp (springtails) and Acari spp (mites) strongly dominated all sites at which invertebrates were present, other groups were more common on loess-rich waste rock type, such as Coleoptera (beetles) and Hymenoptera (ants), than either naturally revegetated or rehabilitated quartz-rich waste rock. Some groups, such as earthworms, amphipods, and centipedes were present only in loess-rich substrate. Details of invertebrates present along a gradient of waste rock types is listed by Rufaut et al. (2006).

Comparison of invertebrate numbers (means from four plots) on quartz-rich waste rock and loess-rich waste rock in the Eastern waste rock pile, for a range of groups (indicated and see text). Black dots are for invertebrates on vegetated plots on the different waste rock substrates, and the grey box is for unvegetated plots. Dashed line indicates expected pattern if there is no difference between substrate types. Note the logarithmic scales
Beetles appeared much more abundant in and near relict bush patches on loess (Fig. 1c), in contrast to the variably revegetated waste rock sample sites described above. These bush patches have well-established trees and most have a range of understorey vegetation (Table 1), and abundant leaf litter compared to a negligible or absent litter layer on waste rock. A total of 81 beetle species from 24 families were identified from the studied bush patches (C. Rufaut, unpublished data). Beetle species richness and abundance was greatest for large (>1,000 m2) bush patches (Table 1), but even single relict trees had more than 20 beetle species established beneath them (Fig. 10a). The overwhelming majority of beetles were native (90% of species and 98% of individuals collected), but adventive beetle counts were locally elevated in small patches and single trees (C. Rufaut, unpublished data),

Discussion
Principal issues hindering ecosystem recovery
Ecosystem recovery at the Wangaloa site is largely dependent on establishment of vegetation on the waste rock piles. Once some vegetation is established, plant litter and moss ensure that the substrate becomes covered, and colonisation by invertebrates can occur from the nearest bush patches. Hence, the first plants to become established on a substrate are the key to subsequent ecosystem recovery. This first plant establishment can be by natural colonisation or by deliberate planting. However, these first plants must survive, and there are several issues which may affect this plant survival, as outlined below.
Substrate acidification and acid mine drainage was perceived to be one of the main environmental issues at the site when rehabilitation commenced in 2000. Many areas of waste rock that had remained unvegetated or poorly vegetated before that time had distinctly low pH substrates (Fig. 1b). Acid mine substrates can inhibit nutrient uptake and enhance toxic metal uptake by plants, resulting in poor germination and death of seedlings, and death of larger plants (Bradshaw et al. 1982; Bullock and Bell 1997; Wong 2003). Kaolinite is abundant in the matrix of quartz-rich waste rock, and accompanying coal-rich waste rock, so acid-mediated dissolution of this mineral may contribute to uptake of toxic levels of Al by plants (Bradshaw et al. 1982; Kochian 1995; Bullock and Bell 1997; Turnau et al. 2007). However, Monterroso et al. (1998) suggest that Al toxicity at coal mines is limited by detoxifying effects of dissolved ions such as Mg2+, Ca2+, and SO4 2−. The latter two ions are abundant in the coal-rich substrates at Wangaloa, as indicated by ephemeral gypsum efflorescences on coal fragments, and these are readily redissolved in rain events to yield high sulfate in mine waters (Figs. 4c, 5b).
Unstable steep slopes of some substrates, particularly quartz-rich waste rock, are a major inhibitor of natural plant colonisation (Fig. 2a). Surface erosion of these steep slopes develops an armour zone of quartz pebbles that inhibits seedling establishment (Craw et al. 2007). Recontouring of some of these quartz-rich waste piles in 2002 helped to ameliorate this issue over large areas of the site, and there has been widespread natural revegetation since then (Fig. 3). However, this substrate disturbance event locally exposed pyrite to oxidation at the surface, and exacerbated acidification of the substrate. The acid slope above the Upper wetland (Fig. 1a) is an extreme example of this, as all introduced vegetation died when pH 1 substrates developed (Fig. 2b).
Elevated boron concentrations in coal fragments in waste rock, particularly the coal-rich substrate areas (Fig. 6a) translate into elevated boron concentrations in plants growing on those substrates (Fig. 6b). Boron reached toxic levels in some plants, including pine trees planted in the 1980s (Craw et al. 2006). There is no physical evidence of boron toxicity in plants at the site (Craw et al. 2006). However, there has been high mortality and poor growth of plants introduced in 2003, and boron toxicity cannot be discounted. In particular, introduced plantings of the native shrub Griselinia littoralis (broadleaf) have had high mortality on coal-rich substrate, possibly because of boron toxicity (Slack et al. 2008; Todd et al. 2009). Conversely, Stass et al. (2007) suggest that moderate amounts of available boron helps to limit Al toxicity and that boron deficiency contributes to Al toxicity effects in soybeans, so some degree of elevated boron in coal-rich substrates may not necessarily be a negative feature for revegetation.
Key factors facilitating natural ecosystem recovery
One of the more spectacular features of this rehabilitation programme is the large amount of natural colonisation of all waste rock types by manuka (L. scoparium) (Figs. 2c, 3). This colonisation is particularly noticeable on recontoured substrates where introduced plants did not thrive, or which died (Fig. 2c). However, this colonisation has occurred throughout the history of mining and subsequent languishment of several parts of the site (Fig. 3; Craw et al. 2007). Natural colonisation by manuka has occurred despite the apparent hindrances to vegetation establishment by some substrate types (outlined in the previous section).
Once the natural colonisation occurred, natural ecosystem recovery ensued, as indicated by invertebrate numbers and diversity data (Figs. 9, 10). This natural ecosystem recovery occurred on all colonised waste rock, and invertebrate data are essentially independent of substrate type. Since the majority of observed invertebrate groups feed either on plants or in the decomposer cycle, their populations are structured around the condition of the flora, not the substrate. Therefore, the invertebrate data patterns observed are linked to plant assemblage spatial patterns, rather than the underlying waste rock type per se (see also Pižl 2001; Wanner and Dunger 2002; Davis et al. 1997). Rapid invertebrate recolonisation of vegetated mine sites, as illustrated at Wangaloa by the invasion of Collembola on planted surfaces <1 year old, has been noted elsewhere as well, involving similar invertebrate groups (Greenslade and Majer 1993; Dunger et al. 2001; Wanner and Dunger 2002).
Natural colonisation by manuka is primarily limited by the available seed sources. Hence, the most effective colonisation has occurred adjacent to, and within 50 m of, forest on the margins of the site and islands of already-established manuka (Figs. 1c; 3). The islands consist of bush patches that became established at various times in the history of the mine, and have survived mine-related earth-moving and subsequent rehabilitation activity. The importance of nearby seed sources for plant colonisation and ecosystem recovery at mine sites has been emphasized by others (e.g. Prach and Pyšek 2001), and McClanahan (1986) showed that numerous small islands are better than a small number of large islands for seed dispersal and colonisation. Habitat islands are also important sites of species richness and sources for colonisation by other ecosystem components such as invertebrates, as shown in this study (Figs. 9, 10) and elsewhere (Holt et al. 1995; Cook et al. 2002; Lovei et al. 2006). Importantly, the beetle data from manuka/kanuka islands show dominant sources of native rather than exotic invertebrates, and that bigger islands favour lower counts of exotic invertebrates. Direct transfer of small islands of established vegetation and associated ecosystem, including soil, from newly disturbed ground to waste rock piles has been trial run successfully (Ross et al. 2000).
Natural colonisation by manuka at the Wangaloa site has been more successful for ecosystem recovery than deliberately introducing seedlings of the same species, as less than 30% of manuka seedlings introduced to disturbed ground had died by 2006 (Todd et al. 2009). The local manuka at Wangaloa has had several million years of history of establishment and re-establishment on the site after natural fires and erosion affected the area repeatedly (above). For example, fire has been suggested to be an important agent in developing local genetic variations in manuka (Bond et al. 2004). In addition, the local acidification, high boron, and low-nutrient quartz-rich substrates have been issues to contend with throughout the evolutionary history of manuka on the site. Hence, use of local genetic stock is clearly an important factor in vegetation re-establishment on sites with significant negative chemical and physical issues. It is undoubtedly more cost-effective to facilitate revegetation with local genetic stock than to attempt extensive modification of substrate chemistry at a site.
Provided water discharge criteria are met, there is no advantage to the ecosystem recovery process in acid neutralisation, substrate nutrient additions, or boron suppression at the Wangaloa site, and same principle applies to many other mine site rehabilitation projects around the world. In particular, local genetic stock of L. scoparium is extremely tolerant of all the above issues, and is well known as a coloniser of disturbed land (Stephens et al. 2005). Essentially identical L. scoparium occurs in Australia and Tasmania (Thompson 1989), and the Leptospermum genus is widespread elsewhere around the world, with at least 83 species recognized (Dawson 1997). Hence, colonisation of local genetic stock of this shrub genus should be facilitated wherever mine site rehabilitation is planned, particularly via shrub islands. Ecosystem recovery will follow rapidly and naturally after this Leptospermum colonisation.
Conclusions
The Wangaloa coal mine site is undergoing effective revegetation via natural and anthropogenic processes. The site was extensively recontoured in 2003 to limit erosion of waste rocks of varying types, and over 70,000 native plants have been introduced. There have been geoecological controls on site recovery, as different substrates, formed from different rock types disturbed in the mining operation, have been revegetated at different rates. Plant establishment on loess siltstone substrate has been most successful, and these parts of the site are now largely revegetated, mainly by anthropogenic plantings and adventive weed incursion. Substrates rich in quartz pebbles and coal fragments have been more difficult to revegetate because of localised acidification, elevated boron, desiccation, low nutrient status, evaporative salt encrustations, or some combination of these issues. Some areas of these substrates are still completely barren, and all or most introduced plants have died.
Natural colonisation of chemically and physically challenging substrate areas has been occurring locally throughout the mine life. Most of this natural colonisation has been done by manuka, a widespread shrubby species (Leptospermum scoparium). Natural colonisation of recontoured waste rock occurred most rapidly around islands of pre-existing manuka that have developed on the site during and after mining. The local genetic stock of L. scoparium has been extremely successful at colonisation, whereas introduced plants of the same species have had more than 70% mortality on all compositions of disturbed substrates.
There has been rapid buildup of both numbers and diversity of invertebrates in and around manuka patches on the site. Invertebrate numbers and diversity are unrelated to substrate composition. Hence, establishment of vegetation is the key to full ecosystem recovery, irrespective of the chemical and physical makeup of the substrate. Since manuka from local genetic stock can apparently colonise all substrates irrespective of acidity, boron content and nutrient status, expensive substrate modification is unnecessary for ecosystem recovery at this mine site. The same principle applies for all mine sites where suitable tolerant local shrubs are available, and facilitation of colonisation by, for example, formation of vegetation islands, will be a more cost-effective and successful mine site rehabilitation strategy leading to ecosystem recovery.
References
Begbie M, Craw D, Rufaut C, Martin C (2007) Temporal and spatial variability of acid rock drainage in a rehabilitated coal mine, Wangaloa, South Otago, New Zealand. NZ J Geol Geophys 50:227–238
Bell LC (2001) Establishment of native ecosystems after mining—Australian experience across diverse biogeographic zones. Ecol Eng 17:179–186
Black A, Craw D (2001) Arsenic, copper and zinc at Wangaloa coal mine, southeast Otago, New Zealand. Int J Coal Geol 45:181–193
Blakemore LC, Searle PL, Daly PK (1987) Methods for chemical analysis of soils. NZ Soil Bureau Scientific Report 80. Dept Sci Ind Res, NZ
Bond WJ, Dickinson KJM, Mark AF (2004) What limits the spread of fire-dependent vegetation? Evidence from geographic variation of serotiny in a New Zealand shrub. Global Ecol Biogeogr 13:115–127
Bradshaw AD (1983) The reconstruction of ecosystems. J Appl Ecol 20:1–17
Bradshaw AD, Williamson MS, Johnson MS (1982) Mine wastes reclamation. Min J 299:75–80
Bruce JG (1973) Loessial deposits in southern South Island, with a definition of Stewarts Claim Formation. NZ J Geol Geophys 16:533–548
Bullock SET, Bell FG (1997) Some problems associated with past mining at a mine in the Witbank coalfield, South Africa. Environ Geol 33:61–71
Carroll C, Merton L, Burger P (2000) Impact of vegetative cover and slope on runoff, erosion, and water quality for field plots on a range of soil and spoil materials on central Queensland coal mines. Aust J Soil Res 38:313–327
Cook WM, Lane KT, Foster BL, Holt RD (2002) Island theory, matrix effects and species richness patterns in habitat fragments. Ecol Lett 5:619–623
Craw D, Beckett S (2004) Water and sediment chemistry of Sutton Salt Lake, east Otago, New Zealand. NZ J Mar Freshw Res 38:315–328
Craw D, Rufaut CG, Haffert L, Todd A (2006) Mobilisation and attenuation of boron during coal mine rehabilitation, Wangaloa, New Zealand. Sci Total Environ 368:444–455
Craw D, Rufaut CG, Hammitt S, Clearwater S, Smith CM (2007) Geological controls on natural ecosystem recovery on mine waste in southern New Zealand. Environ Geol 51:1389–1400
Craw D, Mulliner T, Haffert L, Paulsen H–K, Peake B, Pope J (2008) Stratigraphic controls on water quality at coal mines in southern New Zealand. NZ J Geol Geophys 51:59–72
Davis MR, Langer ER, Ross CW (1997) Rehabilitation of native forest species after mining. NZ J For Sci 27:51–68
Dawson M (1997) A history of Leptospermum scoparium in cultivation: discoveries from the wild. New Plantsman 4:67–78
Dunger W, Wanner M, Hauser H, Hohberg K, Schulz H-J, Schwalbe T, Sefert B, Vogel J, Voigtländer K, Zimdars B, Zulka KP (2001) Development of soil fauna at mine sites during 46 years after afforestation. Pedobiologia 45:243–271
Goodarzi F, Swaine DJ (1994) The influence of geological factors on the concentration of boron in Australian and Canadian coals. Chem Geol 118:301–318
Greenslade P, Majer JD (1993) Recolonization by Collembola of rehabilitated bauxite mines in Western Australia. Aust J Ecol 18:385–394
Harrington HJ 1958 Geology of the Kaitangata coalfield. NZ Geol Surv Bull 59, DSIR, Wellington, NZ
Holmes PM (2001) Shrubland restoration following woody invasion and mining: effects of topsoil depth, seed source, and fertilizer addition. Restor Ecol 9:71–84
Holt RD, Robinson GR, Gaines MS (1995) Vegetation dynamics in an experimentally fragmented landscape. Ecology 76:1610–1624
Kochian LV (1995) Cellular mechanisms of Al toxicity and resistance in plants. Ann Rev Plant Physiol Plant Mol Biol 46:237–260
Lovei GL, Magura T, Tothmeresz B, Kodobocz V (2006) The influence of matrix and edges on species richness patterns of ground beetles (Coleoptera: Carabidae) in habitat islands. Global Ecol Biogeogr 15:283–289
Mains D, Craw D, Rufaut CG, Smith CMS (2006) Phytostabilisation of gold mine tailings, New Zealand. Part 2: experimental evaluation of arsenic mobilization during revegetation. Int J Phytoremediation 8:163–183
McClanahan TR (1986) Seed dispersal from vegetation islands. Ecol Model 32:301–309
Mench M, Bussière S, Boisson J, Castaing E, Vangronsveld J, Ruttens A, De Koe T, Bleeker P, Assunção A, Manceau A (2003) Progress in remediation and revegetation of the barren Jales gold mine spoil after in situ treatment. Plant Soil 249:187–202
Monterroso C, Alvarez E, Marcos MLF, Macias F (1998) Aluminium speciation and phytotoxicity in minesoils of a lignite mine in NW Spain. Fresnius Environ Bull 7:65–73
Munshower FF (1994) Disturbed land revegetation. Lewis, CRC, FL
Neel C, Bril H, Courtin-Normade A, Dutreuil JP (2003) Factors affecting natural development of soil on 35-year-old sulphide-rich mine tailings. Geoderma 111:1–20
Pensa M, Sellin A, Luud A, Valgma I (2004) An analysis of vegetation restoration on opencast soil shale mines in Estonia. Restor Ecol 12:200–206
Pitcairn IK, Teagle DAH, Craw D, Olivo GR, Kerrich R, Brewer TS (2006) Sources of metals and fluids in orogenic gold deposits: insights from the Otago and Alpine Schists, New Zealand. Econ Geol 101:1525–1546
Pižl V (2001) Earthworm succession in afforested colliery spoil heaps in the Sokolov Region, Czech Republic. Restor Ecol 9:359–364
Prach K, Pyšek P (2001) Using spontaneous succession for restoration of human-disturbed habitats: experience from Central Europe. Ecol Eng 17:55–62
Roberts RD, Marrs RH, Skeefington RA, Bradshaw AD (1981) Ecosystem development on naturally colonized china clay wastes. J Ecol 69:153–161
Ross C, Simcock R, Williams P, Toft R, Flynn S, Birchfield R, Comeskey P (2000) Salvage and direct transfer for accelerating restoration of native ecosystems on mine sites in New Zealand. NZ Min Mining Conf Proc, 6
Rufaut CG, Hammitt S, Craw D, Clearwater SG (2006) Plant and invertebrate assemblages on waste rock at Wangaloa coal mine, Otago, New Zealand. NZ J Ecol 30:311–319
Russell WB, La Roi GH (1986) Natural vegetation and ecology of abandoned coal-mined land, Rocky Mountain Foothills, Alberta, Canada. Can J Bot 64:1286–1298
Slack G, Morgan R, Craw D 2008 Chemical controls on establishment success of plants on coal mine waste rocks, Wangaloa, South Otago. Australasian Inst Min Metall NZ Branch Conf, Wellington, Proc, pp 525-534
Sobek AA, Schuller WA, Freeman J, Smith RM 1978. Field and laboratory methods applicable to overburdens and minesoils. EPA-600/2-78-054, Environ Protection Agency, Washington, DC
Stass A, Kotur Z, Horst WJ (2007) Effect of boron on the expression of aluminium toxicity in Phaseolus vulgaris. Physiol Plant 131:283–290
Stephens JMC, Molan PC, Clarkson BD (2005) A review of Leptospermum scoparium (Myrtacae) in New Zealand. NZ J Bot 43:431–449
Suggate RP 1959 New Zealand coals: their geological setting and its influence on their properties. DSIR Bull 134, Wellington, NZ
Thompson J (1989) A revision of the genus Leptospermum. Teleopea 3:301–448
Todd A, Rufaut CG, Craw D, Begbie MA (2009) Indigenous plant species establishment during rehabilitation of an opencast coal mine, southeast Otago, New Zealand. NZ J Forestry Sci (in press)
Turnau K, Henriques FS, Anielska T, Renker C, Buscot F (2007) Metal uptake and detoxification mechanisms in Erica andevalensis growing in a pyrite mine tailing. Environ Exp Biol 61:117–123
Vetterlein D, Waschkies C, Weber E (1994) Nutrient availability in the initial stages of surface mine spoil reclamation—impact on plant growth. J Plant Nutr Soil Sci 162:315–321
Wanner M, Dunger W (2002) Primary immigration and succession of soil organisms on reclaimed opencast coal mining areas in eastern Germany. Eur J Soil Biol 38:137–143
Widerlund A, Ebena G, Landin J (2004) Potential biogeochemical and ecological development of a flooded tailings impoundment at the Kristineberg Zn-Cu mine, northern Sweden. Sci Total Environ 333:249–266
Wong MH (2003) Ecological restoration of mine degraded soils, with emphasis on metal contaminated soils. Chemosphere 50:775–780
Acknowledgments
This study was supported financially by Solid Energy (NZ) Ltd and University of Otago. Background research by, and subsequent discussions with, Amanda Black, Michelle Begbie, Simon Clearwater, Laura Haffert, and Andrea Todd contributed substantially to various aspects of the data set discussed herein. John Nunn and Simon Clearwater laboriously identified many invertebrates. Discussions with Craig Evans, Tim Preston, Dave Thomas, and Arne Cleland helped elucidate many aspects of the site rehabilitation process.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Rufaut, C.G., Craw, D. Geoecology of ecosystem recovery at an inactive coal mine site, New Zealand. Environ Earth Sci 60, 1425–1437 (2010). https://doi.org/10.1007/s12665-009-0278-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12665-009-0278-z