
Abstract
Huntington’s disease (HD) is an autosomal dominant progressive neurodegenerative disorder which affects medium spiny GABAergic neurons mainly in the striatum. Oxidative damage, neuro-inflammation, apoptosis, protein aggregation, and signaling of neurotrophic factors are some of the common cellular pathways involved in HD. Quinolinic acid (QA) causes excitotoxicity by stimulating N-methyl-d-aspartate receptors via calcium overload leading to neurodegeneration. Neuroprotective potential of peroxisome proliferator activated receptor-γ (PPARγ) agonists and histone deacetylase (HDAC) inhibitors have been well documented in experimental models of neurodegenerative disorders; however, their exact mechanisms are not clear. Therefore, present study has been designed to explore possible neuroprotective mechanism of valproic acid (VPA) and its interaction with rosiglitazone against QA induced HD-like symptoms in rats. Single bilateral intrastriatal QA (200 nmol/2 μl saline) administration significantly caused motor incoordination, memory impairment, oxidative damage, mitochondrial dysfunction (complex I, II, II and IV), cellular alterations [tumor necrosis factor-alpha (TNF-α), caspase-3, brain derived neurotrophic factor, acetylcholinesterase], and striatal neurodegeneration as compared to sham group. Treatment with rosiglitazone (5, 10 mg/kg) and VPA (100, 200 mg/kg) for 21 days significantly attenuated these behavioral, biochemical, and cellular alterations as compared to control (QA 200 nmol) group. However, VPA (100 mg/kg) treatment in combination with rosiglitazone (5 mg/kg) for 21 days synergized their neuroprotective effect, which was significant as compared to their effects per se in QA-treated animals. The present study provides an evidence of possible interplay of PPARγ agonists and HDAC inhibitors as a novel therapeutic strategy in the management of HD.
Similar content being viewed by others
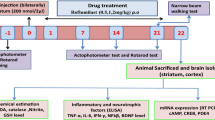

Avoid common mistakes on your manuscript.
Introduction
Huntington’s disease (HD) is an autosomal dominantly inherited neurodegenerative disorder manifested by motor dysfunction, cognitive, and psychiatric complications in middle-aged people. HD is caused due to expansion of the cytosine–adenine–guanine (CAG) repeats in the gene coding for the N-terminal region of the huntingtin protein, leading to the formation of a polyglutamine stretch physiological HD alleles which comprise 6–35 CAGs, whereas above 39 CAGs, there is obligatory disease development; greater the expansion of CAGs, earlier is the disease manifestation (Gardian and Vecsei 2004; Zadori et al. 2009). Although HD usually involves chorea and other abnormal movements, the progressive cognitive impairment and behavioral problems are perhaps even more disabling and remains as a challenge.
Quinolinic acid (QA) is one of the reliable experimental models used to reproduce HD-like symptoms as observed clinically in HD patients (Schwarcz et al. 1983, 2010; Ferrante et al. 1993; Kalonia et al. 2012). The first step in QA-induced neuronal damage involves hyper-stimulation of N-methyl-d-aspartate (NMDA) receptors (Choi 1992) leading to a massive Ca2+ influx and activation of Ca2+ dependent phospholipases A2 activity, which later converts into prostaglandin G2 (PGG2) by cyclo-oxygenases and subsequently reduces to PGH2 with the production of free radical intermediates (Kukreja et al. 1986). In addition, QA-induced neurotoxicity involves an increased expression of the pro-inflammatory cytokines such as interleukins, tumor necrosis factors (Kalonia et al. 2012).
Increasing evidences suggest that an altered balance between histone acetyl transferases and histone deacetylase (HDAC) activity may accompany the development of chronic neurodegenerative disorders, including HD (Langley et al. 2005; Sadri-Vakili and Cha 2006). The HDACs remove acetyl groups which lead to reduction in transcription processes. Valproic acid (VPA), a HDAC inhibitor has been reported to exert its neuroprotective effects in several neurodegenerative conditions (Jeong et al. 2003; Mark et al. 1995; Hashimoto et al. 2002). Besides, different authors suggest different neuroprotective mechanisms of VPA, which include inhibition of glycogen synthase kinase-3β (GSK-3β), attenuation of glutamate-induced excitotoxicity, blockade of sodium and calcium channels, elevating GABA and reducing aspartate levels within the brain, prevention of mitochondrial dysfunction, reduction of apoptosis, and promoting neuronal survival (Valvassori et al. 2010; Phiel et al. 2001; Kwan and Brodie 2001). However, its exact mechanism of action is not clear yet.
Rosiglitazone, selective peroxisome proliferator activated receptor-γ (PPARγ) agonist, has been demonstrated to regulate lipid and carbohydrate homeostasis, and exert anti-inflammatory actions in nervous system (Heneka et al. 2007; Bernardo and Minghetti 2008). Moreover, PPARγ has been detected in both neuronal and glial cells (Cristiano et al. 2001), which regulates the expression of COX, iNOS, and inflammatory cytokines (Bernardo and Minghetti 2008; Cristiano et al. 2001). Above all, neuroprotective effects of various PPARγ agonists in animal models of neurodegenerative disorders are mainly attributed to their anti-oxidant and anti-inflammatory properties (Kiaei et al. 2005; Drew et al. 2006; Kalonia et al. 2010a). However, exact neuroprotective mechanism of PPARγ agonists in HD and related conditions are yet to be explored.
Till date the neuroprotective potential of PPARγ agonists (rosiglitazone) has been attributed to their anti-oxidant, anti-inflammatory (Lee et al. 2012; Kalonia et al. 2010a; Quintanilla et al. 2008; Park et al. 2007), and microglial modulation (McGeer and McGeer 2008) properties. HDAC inhibition also has been reported to ameliorate motor deficits in several movement disorder conditions including HD, and the role of VPA has been well documented in these conditions (Sadri-Vakili and Cha 2006; Langley et al. 2005; Ferrante et al. 2003). However, none of the hypotheses have adequately explained the exact role of HDAC and PPARγ in the amelioration of this multifactorial disease (HD). Hence, the present study has been undertaken to investigate the possible interplay between VPA and rosiglitazone against QA-induced neurobehavioral, biochemical, mitochondrial, and cellular alterations in rats.
Materials and Methods
Animals
Male Wistar rats (250–300 g), bred in Central Animal House (CAH) of Panjab University, Chandigarh were used in the study. The animals were kept under standard laboratory conditions, maintained on 12-h light/dark cycle, with food and water ad libitum. Experiment was carried out between 9.00 and 17.00 h. The experimental protocol was approved by the Institutional Animal Ethics Committee, Panjab University, Chandigarh (IAEC/170-175/UIPS-17) and conducted according to the Indian National Science Academy Guidelines for the use and care of experimental animals.
Intrastriatal Administration of QA
Animals were anesthetized with intraperitoneal administration of thiopental sodium (45 mg/kg), and surface of the skull was exposed by making an incision on the scalp. Small holes (l–2 mm diameter) were made in the skull at anterior +1.7 mm; lateral ±2.7 mm; ventral −4.8 mm on both sides of bregma and dura as described by Paxinos and Watson (2007). QA (200 nmol/2 μl saline) (Sigma-Aldrich, St Louis, MO, USA) was administered bilaterally over a period of 2 min in the striatum by means of a 28-gauge stainless steel needle attached to a Hamilton syringe. The injection needle was left in place for another 2 min to prevent back diffusion of the injected drug solution.
Drug and Treatment Schedule
The present study protocol (Fig. 1) includes fifteen treatment groups (n = 12) (Table 1). Rosiglitazone (Ranbaxy Laboratories, Gurgaon, India) suspended in 0.25 % w/v sodium carboxy methyl cellulose (CMC) and VPA (Sigma Chemicals, St. Louis, MO, USA) dissolved in normal saline were administered intraperitoneally (i.p.) as per body weight (not exceeding 5 ml/kg b.w.) to all the treatment groups daily in the morning 10: 00 h, for 21 days starting from the day after QA administration. Doses were selected on the basis of our preliminary laboratory results as well as published reports (Zadori et al. 2009; Shear et al. 1998; Jin et al. 2013; Carta et al. 2011; Escribano et al. 2010; Verma et al. 2011).

Experimental protocol
Behavioral Assessments
Body Weight
The body weight of animals was recorded just before QA administration and on the last day of the study (21st day). The change in body weight was calculated according to the following formula:
Assessment of Total Locomotor Activity
The locomotor activity was assessed using actophotometer (IMCORP, Ambala, India) on weekly intervals. Animals were placed individually in the activity chamber for 3 min as a habituation period before recording actual motor activity for next 5 min. The instrument consisted of a closed arena equipped with 12 infrared light-sensitive photocells in two rows (six in each row), at a distance of 3 and 9 cm, respectively, and values expressed as counts per 5 min (Kumar et al. 2012b).
Rotarod Activity
The rotarod activity task was performed in the animals individually on weekly intervals for the evaluation of motor coordination and grip strength after QA administration. Animals were given a prior training session before rotarod performance task. Animals were placed on the rotating rod (speed 25 rpm) with a diameter of 7 cm (IMCORP, Ambala, India) individually for a fixed cut-off time (90 s), and three separate trials were performed at 5 min interval for each rat as described by Gaur and Kumar (2011).
Morris Water Maze Task
Morris water-maze (MWM) test is most commonly used to test memory (Morris 1984; Tuzcu and Baydas 2006; Kumar et al. 2012a, 2013) consisting of a large circular pool (150 cm in diameter, 45 cm in height, filled to a depth of 30 cm with water at 28 ± 1 °C). The tank was divided into four equal quadrants and a submerged platform (10 × 10 cm2), placed 1 cm below the surface of water in the middle of the target quadrant. The position of platform was kept unaltered throughout the training session. The tank was located in a large room where there were several brightly colored cues external to the maze, which were visible from the pool and could be used by the rats for spatial orientation and kept unaltered throughout the study. The water maze task was carried out for four consecutive days (seventeenth to twentieth day) where animals received four consecutive daily training trials, each at an interval of 30 min approximately. For each trial, individual rat was gently put into the water at one of four starting positions (the sequence of which being selected randomly) to locate the submerged platform (maintained at Q4) and allowed to stay on the platform for 20 s. If animal failed to find the platform within 90 s, same was gently guided onto the platform and allowed to remain there for 20 s. Escape latency time (ELT) to locate the hidden platform in water maze was noted as index of acquisition or learning using a computer tracking system with EthoVision software (Noldus Information Technology, Wageningen, The Netherlands).
Memory Consolidation Test
A probe trial was performed (Tuzcu and Baydas 2006) 24 h after the last training session (on 21st day) to assess the extent of memory consolidation wherein the animal was placed into the pool for a total duration of 90 s as in the training trial, except that the hidden platform was removed from the pool. Various parameters such as time spent in target quadrant, frequency of appearance in target quadrant, and percentage of total path length traversed in target quadrant were measured using computer tracking system with EthoVision software (Noldus Information Technology, Wageningen, The Netherlands). All these parameters indicate the degree of memory consolidation that has taken place after learning.
Biochemical Assessments
Dissection and Homogenization
Following behavioral assessments, the animals were divided into three groups; first group was used for the biochemical assays, second one used for hematoxylin and eosin staining, and the third one was used for the estimation striatal lesion volume. For biochemical analysis, animals were sacrificed by decapitation, and the brains were divided into two equal halves. One half was used for anti-oxidant enzyme activity estimation and the second one was used for estimation of mitochondrial complex analysis. Striatum, cortex, and hippocampus were dissected out on ice and weighed. For anti-oxidant enzyme activity estimation, a 10 % (w/v) tissue homogenates were prepared in 0.1 M phosphate buffer (pH 7.4). The homogenates were centrifuged at 10,000×g at 4 °C for 15 min. Aliquots of supernatants were separated and used for biochemical estimations. Double beam UV–Vis Spectrophotometer [UV-Pharmaspec 1700 Shimadzu (Japan)] was used for the estimation of biochemical and mitochondrial parameters throughout the study.
Measurement of Lipid Peroxidation
The quantitative measurement of lipid peroxidation (LPO) was carried out according to the method as described by Wills (1966). The amount of malondialdehyde (MDA), a measure of LPO was measured by the reaction with thiobarbituric acid at 532 nm using UV–Vis spectrophotometer [UV-Pharmaspec 1700 Shimadzu (Japan)]. The values were calculated using molar extinction coefficient of chromophore (1.56 × 105 M−1/cm).
Estimation of Nitrite
Accumulation of nitrite, an indicator of the production of nitric oxide (NO), was determined with a colorimetric assay with Greiss reagent as described by Green et al. (1982). Equal volumes of supernatant and Greiss reagent were mixed, and the mixture was incubated in dark for 10 min at room temperature. Absorbance was recorded at 540 nm with a UV–Vis spectrophotometer [UV-Pharmaspec 1700 Shimadzu (Japan)]. The concentration of nitrite in the supernatant was determined from a sodium nitrite standard curve.
Catalase Estimation
Catalase activity was assayed by the method of Luck, where in the breakdown of hydrogen peroxides (H2O2) is measured at 240 nm. In brief, assay mixture consisted of 3 ml of H2O2 phosphate buffer and 0.05 ml of supernatant of tissue homogenate (10 %), and change in absorbance was recorded at 240 nm with a UV–Vis spectrophotometer [UV-Pharmaspec 1700 Shimadzu (Japan)]. The results were obtained as micromole H2O2 decomposed per milligram of protein/min (Luck 1965).
Superoxide Dismutase Activity
Superoxide dismutase (SOD) activity was assayed according to the method of Kono, wherein the reduction of nitro blue tetrazolium (NBT) was inhibited by the SOD and measured at 560 nm using a UV–Vis spectrophotometer [UV-Pharmaspec 1700 Shimadzu (Japan)]. In brief, the reaction was initiated by the addition of the hydroxylamine hydrochloride to the assay mixture containing NBT and sample (Kono 1978). The results were expressed as unit/mg protein where one unit of enzyme is defined as the amount of enzyme inhibiting the rate of reaction by 100 %.
Glutathione (GSH) Assay
Different glutathione contents in striatum were estimated according to the different methods. Total glutathione analysis was done by the method of Zahler and Cleland (1968) which is based on the reduction with dithioerythritol and determination of the resulting mono thiols with DTNB in the presence of arsenite. Reduced glutathione in the brain was estimated according to the method described by Ellman (1959). A 1.0 ml of homogenate was precipitated with 1.0 ml of 4 % sulfosalicylic acid by keeping the mixture at 4 °C for 1 h, and the samples were immediately centrifuged at 1,200×g for 15 min at 4 °C. The assay mixture contains 0.1 ml of supernatant, 2.7 ml of phosphate buffer of pH 8.0, and 0.2 ml of 0.01 M dithiobisnitro benzoic acid (DTNB). The yellow color developed was read immediately at 412 nm using a UV–Vis spectrophotometer [UV-Pharmaspec 1700 Shimadzu (Japan)]. Results were calculated using molar extinction coefficient of chromophore (1.36 × 104 M−1/cm) nano mole GSH per milligram protein. Oxidized glutathione (GSSG) was quantified by subtracting the value of glutathione reduced from total glutathione. Redox ratio (reduced glutathione/oxidized glutathione) was determined for all the groups by taking the ratio of reduced glutathione/oxidized glutathione.
Protein Estimation
Protein estimation was done by biuret method using bovine serum albumin as standard (Gornall et al. 1949).
Estimation of Acetyl Cholinesterase (AChE) Activity
AChE activity (marker of cholinergic neurons in the brain) was assessed in the hippocampal region by the method of Ellman et al. (1961). The assay mixture contained 0.05 ml of supernatant, 3 ml of sodium phosphate buffer (pH 8), 0.1 ml of acetylthiocholine iodide and 0.1 ml of DTNB (Ellman reagent). The change in absorbance was measured for 2 min at 30 s interval at 412 nm using a UV–Vis spectrophotometer [UV-Pharmaspec 1700 Shimadzu (Japan)]. Results were expressed as micromoles of acetylthiocholine iodide hydrolyzed per min per mg of protein (percentage of sham).
Mitochondrial Complex Enzyme Estimation
Isolation of Rat Brain Mitochondria
Rat brain mitochondria were isolated by the method as described by Berman and Hastings. The striatal regions were homogenized in isolation buffer with EGTA (215 mM Mannitol, 75 mM sucrose, 0.1 % BSA, 20 mM HEPES, 1 mM EGTA, and pH 7.2). Homogenates were centrifuged at 13,000×g for 5 min at 4 °C. Pellets were resuspended in isolation buffer with EGTA and spun again at 13,000×g for 5 min. The resulting supernatants were transferred and topped off with isolation buffer with EGTA and again spun at 13,000×g for 10 min. Pellets containing pure mitochondria were resuspended in isolation buffer without EGTA (Berman and Hastings 1999).
Complex I (NADH Dehydrogenase) Activity
NADH dehydrogenase activity was measured spectrophotometrically according to the method of King and Howard, which involves catalytic oxidation of NADH to NAD+ with subsequent reduction of cytochrome C. In brief, the reaction mixture contained 0.2 M glycyl glycine buffer pH 8.5, 6 mM NADH in 2 mM glycyl glycine buffer, and 10.5 mM cytochrome C. The reaction was initiated by the addition of requisite amount of solubilized mitochondrial sample and the absorbance change at 550 nm was recorded over the period of 2 min, and the results were expressed as percentage of sham (King and Howard 1967).
Complex II (Succinate Dehydrogenase) Activity
Succinate dehydrogenase (SDH) activity was measured spectrophotometrically according to the previously reported method of King which involves oxidation of succinate by an artificial electron acceptor (potassium ferricyanide). The reaction mixture consists of 0.2 M phosphate buffer pH 7.8, 1 % BSA, 0.6 M succinic acid, and 0.03 M potassium ferricyanide and the reaction was initiated by the addition of mitochondrial sample (King 1967). The absorbance change was followed at 420 nm for 2 min, and the results were expressed as percentage of sham.
Complex IV (Cytochrome Oxidase Activity)
Cytochrome oxidase activity was assayed in striatal mitochondria according to the method of Sottocasa et al. The assay mixture contained 0.3 mM reduced cytochrome C in 75 mM phosphate buffer, and the reaction was started by the addition of solubilized mitochondrial sample. The absorbance change was recorded at 550 nm over the period of 2 min, and the results were expressed as percentage of sham (Sottocasa et al. 1967).
Mitochondrial Redox Activity (Complex III) Assay
The method (MTT assay) employed is based on the in vitro studies to evaluate mitochondrial redox activity through the conversion of MTT tetrazolium salt to formazan crystals by mitochondrial respiratory chain reactions in isolated mitochondria by the method as described by Liu et al. The absorbance of the resulting medium was measured by an ELISA reader (Biorad-Xmerk) at a wavelength of 580 nm, and the results were expressed as percentage of sham (Liu et al. 1997).
Estimation of Pro-inflammatory Cytokines [Tumor Necrosis Factor-Alpha (TNF-α)]
The quantifications of TNF-α were performed by rat TNF-α immunoassay kit (R&D Systems, Minneapolis, MN, USA) in different brain regions (striatum, cortex and hippocampus) with the help of an ELISA reader (Biorad, X-merk). The assay employs the sandwich enzyme immunoassay technique. Final concentrations of TNF-α were calculated from plotted standard curves, and results were expressed as percentage of sham.
Estimation of Caspase-3
The quantitative measurement of caspase-3 was carried out in different brain regions (striatum, cortex, and hippocampus) using caspase-3 colorimetric kit (R&D Systems, Minneapolis, MN, USA) according to the manufacturer’s instructions. The cleavage of the peptide by the caspase releases the chromophore pNA, which was assessed spectrophotometrically at a wavelength of 405 nm.
Estimation of Brain Derived Neurotrophic Factor (BDNF)
BDNF acts on certain neurons of the central nervous system and the peripheral nervous system, helping to support the survival of existing neurons and encourage the growth and differentiation of new neurons and synapses. With the ChemiKine BDNF assay system, mouse monoclonal antibodies generated against human BDNF are coated onto a microplate and are used to capture BDNF from a sample. BDNF-specific, biotin-conjugated, mouse monoclonal antibodies detect the captured BDNF. The BDNF assay was carried out for different brain regions (striatum, cortex, and hippocampus) using ChemiKine™ BDNF Sandwich ELISA Kit (Millipore, USA) according to the supplied manufacturer’s instruction with the kit.
Striatal Lesion Volume Measurement (TTC Staining)
On the 21st day, the animals were sacrificed for 2,3,5-triphenyltetrazolium chloride (TTC) staining. Brains were quickly removed, placed in ice-cold saline, and sectioned at 2 mm intervals using rat brain matrix and stained in 2 % TTC solution in the dark at 37 °C for 30 min. After 30 min, the sections were removed and placed in 4 % phosphate buffered formalin for photography. Complete lack of TTC staining was defined as lesioned area. The lesioned regions of the hemispheres were selected, and lesion volumes were measured using computer-based image analysis software (Image J 1.42q, NIH, USA) as described by Kim et al. (2005).
Histopathological Analysis (Hematoxylin and Eosin Staining)
Animals were sacrificed by decapitation immediately after the last behavioral test. The brains were removed and transferred to formalin (10 %, v/v). The brain tissues were cut into 3 mm thickness, and its blocks were embedded in paraffin. The brain sections (5–10 μm) thick were prepared and stained with hematoxylin and eosin.
Statistical Analysis
Graph Pad Prism (Graph Pad Software, San Diego, CA) was used for all statistical analysis. All the values were expressed as mean ± SEM. The data were analyzed using two-way analysis of variance (ANOVA) followed by Bonferroni’s test. In all the tests, criterion for statistical significance was considered to be P < 0.05.
Results
Effect of VPA and Rosiglitazone on Body Weight
Single bilateral intrastriatal QA (200 nmol) administration significantly reduced body weight on day 21 as compared to sham group. Rosiglitazone (5, 10 mg/kg) and VPA (100, 200 mg/kg) treatment for 21 days significantly attenuated loss in body weight as compared to control (QA 200 nmol) group (Fig. 2). However, treatment of VPA (100 mg/kg) in combination with rosiglitazone (5 mg/kg) for 21 days further synergized their protective effect which was significant as compared to their effect per se in QA-treated animals (F = 12.45; P < 0.0001). Meanwhile, treatment of VPA (200 mg/kg) in combination with rosiglitazone (10 mg/kg) for 21 days did not display any significant improvement in body weight as compared to their effect per se in QA-treated animals which could be possibly due to the ceiling effect. Sham group did not show any significant difference on body weight as compared to naive group. Similarly, per se treatments of VPA, rosiglitazone, and their combinations were found to be non-significant as compared to sham group.

Effect of valproic acid, rosiglitazone, and their combination on body weight in QA-treated rats. Data expressed as mean ± SEM. (a) P < 0.05 compared to sham group, (b) P < 0.05 compared to control (QA-treated group), (c) P < 0.05 compared to Rosi (5), (d) P < 0.05 compared to VPA (100); (two-way ANOVA followed by Bonferroni’s test). QA Quinolinic acid, Rosi rosiglitazone, VPA valproic acid
Effect of VPA and Rosiglitazone on Locomotor Activity
Locomotor activity in all treatment groups was invariable prior to surgery. Single intrastriatal QA (200 nmol) administration significantly increased locomotor activity on day 7; however, with disease progression, a substantial decrease in locomotor activity was observed on day 14 and 21 as compared to sham group. No significant difference in locomotor activity was observed between sham and naive group. Further, rosiglitazone (5, 10 mg/kg) and VPA (100, 200 mg/kg) treatment for 21 days showed significant improvement in locomotor activity at the end of first, second, and third week as compared to control (QA treated) group (Fig. 3). Moreover, VPA (100 mg/kg) in combination with rosiglitazone (5 mg/kg) for 21 days further synergized their protective effect, which was significant only on day 21 as compared to their effects per se in QA-treated animals (F = 6.86; P < 0.0001). Meanwhile, treatment of VPA (200 mg/kg) in combination with rosiglitazone (10 mg/kg) for 21 days did not display any marked improvement in locomotor activity as compared to their effect per se in QA-treated animals which might be attributed to their ceiling effect. However, per se treatments of VPA, rosiglitazone, and their combinations did not produce any significant effect on locomotor activity as compared to sham group.

Effect of valproic acid, rosiglitazone, and their combination on locomotor activity in QA-treated rats. Data expressed as mean ± SEM. (a) P < 0.05 compared to sham group, (b) P < 0.05 compared to control (QA-treated group), (c) P < 0.05 compared to Rosi (5), (d) P < 0.05 compared to VPA (100); (two-way ANOVA followed by Bonferroni’s test). QA Quinolinic acid, Rosi rosiglitazone, VPA valproic acid
Effect of VPA and Rosiglitazone on Rotarod Test in QA-Treated Rats
Single intrastriatal QA (200 nmol) administration significantly impaired rotarod performance and shortened fall off time of the animals as compared to sham group in rotarod test on day 7, 14, and 21, whereas sham group did not produce any significant effect as compared to naive animals. However, rosiglitazone (5, 10 mg/kg) and VPA (100, 200 mg/kg) treatment significantly attenuated fall off time on day 7, day 14, and day 21 (Fig. 4) demonstrating improved grip strength in QA-treated animals. Moreover, combination treatment of VPA (100 mg/kg) with rosiglitazone (5 mg/kg) improved fall off time which was significant as compared to their effects per se in QA-treated animals (F = 10.68; P < 0.0001). Meanwhile, treatment of VPA (200 mg/kg) in combination with rosiglitazone (10 mg/kg) for 21 days did not display any significant improvement in the grip strength performance as compared to their effect per se in QA-treated animals which might be attributed to their ceiling effect. However, per se treatments of VPA, rosiglitazone, and their combinations were found to be non-significant as compared to sham group.

Effect of valproic acid, rosiglitazone, and their combination on rotarod test in QA-treated rats. Data expressed as mean ± SEM. (a) P < 0.05 compared to sham group, (b) P < 0.05 compared to control (QA-treated group), (c) P < 0.05 compared to Rosi (5), (d) P < 0.05 compared to VPA (100); (two-way ANOVA followed by Bonferroni’s test). QA Quinolinic acid, Rosi rosiglitazone, VPA valproic acid
Effect of VPA, Rosiglitazone, and Their Combination on Escape Latency of QA-Treated Rats in Morris Water Maze Task
The cognitive function was assessed in the Morris water maze test. The mean escape latency was found to be invariable between all the treatment groups on the first day of training (day 17) but from second day (day 18) onwards, significant difference in transfer latency was observed. Single intrastriatal QA (200 nmol) administration demonstrated a poor ability to find the platform and learn its location till the fourth day (twentieth day) of training (Fig. 5) which was significant as compared to the sham group. This poor performance was significantly attenuated by rosiglitazone (5, 10 mg/kg) and VPA (100, 200 mg/kg) treatment and demonstrated a decrease in latency to find the platform from second day of training (eighteenth day) onwards as compared to the control (QA 200 nmol) group. However, treatment of VPA (100 mg/kg) in combination with rosiglitazone (5 mg/kg) demonstrated recovery in memory performance (transfer latency) in this period (eighteenth to twentieth day), which was significant as compared to their effects per se in QA-treated animals (F = 4.88; P < 0.0001). Meanwhile, treatment of VPA (200 mg/kg) in combination with rosiglitazone (10 mg/kg) for 21 days did not display any synergistic effect as compared to their effect per se in QA-treated animals which might be attributed to their ceiling effect. However, sham treatment did not offer any significant result as compared to naive group animals. Similarly, per se treatments of VPA, rosiglitazone, and their combinations were found to be non-significant as compared to sham group.

Effect of valproic acid, rosiglitazone, and their combination on escape latency of QA-treated rats in Morris water maze task. Data expressed as mean ± SEM. (a) P < 0.05 compared to sham group, (b) P < 0.05 compared to control (QA-treated group), (c) P < 0.05 compared to Rosi (5), (d) P < 0.05 compared to VPA (100); (two-way ANOVA followed by Bonferroni’s test). QA Quinolinic acid, Rosi rosiglitazone, VPA valproic acid
Effect of VPA, Rosiglitazone and Their Combination on Probe Trial/Memory Consolidation Test (Time Spent, Frequency of Appearance and Percentage of Total Path Length Traversed in Target Quadrant) in QA-Treated Rats
The probe trial reveals how well the animals were learned and consolidated the platform location during the training in Morris water maze task. All the animals showed a significant difference in the probe trial (time spent, frequency of appearance, and percentage of total path length traversed in target quadrant) on 21st day which was significantly decreased in the QA (200 nmol)-treated rats as compared to the sham group. However, treatment of VPA (200 mg/kg) and rosiglitazone (10 mg/kg) significantly improved these probe trial parameters (time spent, frequency of appearance, and percentage of total path length traversed in target quadrant) as compared to control (QA 200 nmol) group. Further, VPA (100 mg/kg) and rosiglitazone (5 mg/kg) treatment significantly improved time spent and percentage of total path length traversed in target quadrant in QA-treated animals. Meanwhile, treatment of VPA (100 mg/kg) and rosiglitazone (5 mg/kg) combination significantly augmented these parameters in QA administered animals which were significant as compared to their effects per se in QA-treated animals [time spent in target quadrant (F = 41.7); frequency in target quadrant (F = 5.06); percentage of path length traversed in target quadrant (F = 41.78); P < 0.0001]. Meanwhile, treatment of VPA (200 mg/kg) in combination with rosiglitazone (10 mg/kg) for 21 days did not display any marked improvement in memory performance as compared to their effect per se in QA-treated animals which might be attributed to their ceiling effect. However, sham treatment did not offer any significant effect on these parameters as compared to naive group animals (Fig. 6a–c). Further, per se treatments of VPA, rosiglitazone, and their combinations were found to be non-significant as compared to sham group.

Effect of valproic acid, rosiglitazone, and their combination on probe trial/memory consolidation test (time spent, frequency of appearance, and percentage of total path length traversed in target quadrant) in QA-treated rats. Data expressed as mean ± SEM. (a) P < 0.05 compared to sham group, (b) P < 0.05 compared to control (QA-treated group), (c) P < 0.05 compared to Rosi (5), (d) P < 0.05 compared to VPA (100); (two-way ANOVA followed by Bonferroni’s test). QA Quinolinic acid, Rosi rosiglitazone, VPA valproic acid
Effect of VPA, Rosiglitazone and Their Combination on Hippocampal Acetylcholinesterase (AChE) Level in QA-Treated Rats
Sham group animals did not illustrate any significant effect on the hippocampal acetylcholinesterase enzyme (AChE) activity as compared to the naive animals, whereas single bilateral intrastriatal QA (200 nmol) administration significantly increased AChE levels as compared to the sham group. Unlike the lower dose of VPA (100 mg/kg) and rosiglitazone (5 mg/kg), 21 days pre-treatment with VPA (200 mg/kg) and rosiglitazone (10 mg/kg) significantly attenuated AChE activity as compared to control (QA 200 nmol) animals. However, treatment of VPA (100 mg/kg) in combination with rosiglitazone (5 mg/kg) for 21 days significantly attenuated AChE activity in the hippocampus which was significant as compared to control (QA 200 nmol) as well as their effects per se in QA-treated animals (F = 87.28; P < 0.001)(Fig. 7). Meanwhile, treatment of VPA (200 mg/kg) in combination with rosiglitazone (10 mg/kg) for 21 days did not display any marked synergizm in the AChE activity as compared to their effect per se in QA-treated animals. Similarly, per se treatments of VPA, rosiglitazone, and their combinations were found to be non-significant as compared to sham group.

Effect of valproic acid, rosiglitazone, and their combination on hippocampal acetylcholinesterase (AChE) level in QA-treated rats. Data expressed as mean ± SEM [(percentage of sham (0.03 ± 0.007)]. (a) P < 0.05 compared to sham group, (b) P < 0.05 compared to control (QA-treated group), (c) P < 0.05 compared to Rosi (5), (d) P < 0.05 compared to VPA (100); (two-way ANOVA followed by Bonferroni’s test). QA Quinolinic acid, Rosi rosiglitazone, VPA valproic acid
Effect of VPA, Rosiglitazone and Their Combination on Brain Oxidative Damage (LPO, Nitrite, SOD, Catalase and Glutathione Levels) in QA-Treated Rats
Single intrastriatal QA administration significantly increased LPO and nitrite levels, and depleted endogenous anti-oxidant enzymes [SOD, catalase, and glutathione (GSH)] levels in the striatum as compared to the sham group. However, sham treatment did not show any significant effect as compared to naive group animals. Moreover, 21 days treatment with rosiglitazone (5, 10 mg/kg) and VPA (100, 200 mg/kg) significantly attenuated oxidative stress (decreased LPO, nitrite concentration and restored the levels of SOD, catalase, glutathione, and redox ratio) as compared to control (QA 200 nmol) group (Tables 2, 3). Meanwhile, treatment of VPA (100 mg/kg) in combination with rosiglitazone (5 mg/kg) for 21 days significantly synergized their protective effects which were significant as compared to their effects per se in QA-treated animals [LPO (F = 13.21); nitrite (F = 21.21); SOD (F = 10.23); GSH (F = 16.36); P < 0.001)]. Meanwhile, treatment of VPA (200 mg/kg) in combination with rosiglitazone (10 mg/kg) for 21 days did not display any improvement in the oxidative biomarkers as compared to their effect per se in QA-treated animals probably due to their ceiling effect. However, per se treatments of VPA, rosiglitazone, and their combinations were found to be non-significant as compared to sham group.
Effect of VPA, Rosiglitazone, and Their Combination on Mitochondrial Enzyme Complex (I, II, III and IV) Activities in QA-Treated Animals
Single intrastriatal QA administration significantly impaired mitochondrial enzyme complex activities (I, II, and IV) as well as mitochondrial redox activity (complex III) in the striatum as compared to the sham group, whereas the sham treatment did not substantiate any significant effect on the mitochondrial enzyme complex activities (I, II, and IV) and mitochondrial redox activity as compared to naive animals. However, rosiglitazone (5, 10 mg/kg) and VPA (100, 200 mg/kg) treatment significantly restored mitochondrial enzyme complex activities (I and II) as well as mitochondrial redox activity as compared to control (QA 200 nmol) group. Unlike the lower dose of VPA (100 mg/kg) and rosiglitazone (5 mg/kg), treatment of VPA (200 mg/kg), and rosiglitazone (10 mg/kg) for 21 days significantly restored mitochondrial enzyme complex IV activity as compared to control (QA 200 nmol) group, whereas combination of VPA (100 mg/kg) with rosiglitazone (5 mg/kg) treatment for 21 days significantly restored these QA-induced mitochondrial alterations which were significant as compared to their effects per se in QA-treated animals [I (F = 23.8); II (F = 25.5); III (F = 28.28); IV (F = 33.68); P < 0.001)] (Fig. 8a–d). Meanwhile, treatment of VPA (200 mg/kg) in combination with rosiglitazone (10 mg/kg) for 21 days did not display any significant improvement in the mitochondrial complex enzyme activities as compared to their effect per se in QA-treated animals which might be attributed to their ceiling effect. However, per se treatments of VPA, rosiglitazone, and their combinations were found to be non-significant as compared to sham group.

Effect of valproic acid, rosiglitazone, and their combination on mitochondrial enzyme complex (I, II, III, and IV) activities in QA-treated animals. Data expressed as mean ± SEM (percentage of sham). (a) P < 0.05 compared to sham group, (b) P < 0.05 compared to control (QA-treated group), (c) P < 0.05 compared to Rosi (5), (d) P < 0.05 compared to VPA (100); (two-way ANOVA followed by Bonferroni’s test). QA Quinolinic acid, Rosi rosiglitazone, VPA valproic acid. [Mean ± SEM (sham) Complex I 207 ± 12.57, Complex II 182 ± 8.93, Complex III 169 ± 8.35, and Complex IV 194 ± 7.68]
Effect of VPA, Rosiglitazone, and Their Combination on TNF-α, Caspase-3, and BDNF Levels in QA-Treated Animals
Single intrastriatal QA administration (200 nmol/2 μl) resulted in significant increase in the levels of apoptotic marker (caspase-3) and pro-inflammatory cytokine (TNF-α) but conversely reduced BDNF levels in the striatum, cortex, and hippocampus as compared to sham group. However, sham treatment did not produce any significant effect on these parameters (TNF-α, caspase-3 and BDNF levels) as compared to naive group. Further, treatment of VPA (100, 200 mg/kg) and rosiglitazone (5, 10 mg/kg) for 21 days significantly attenuated the increase in levels of TNF-α (Fig. 9) and capase-3 (Fig. 10), and restored BDNF levels (Fig. 11) as compared to QA-treated group. Meanwhile, treatment of VPA (100 mg/kg) in combination with rosiglitazone (5 mg/kg) for 21 days synergized their protective effects which were significant as compared to their effects per se in QA-treated animals [TNF-α (F = 10.87); caspase-3 (F = 14.59); BDNF (F = 12.98); P < 0.001]. Meanwhile, treatment of VPA (200 mg/kg) in combination with rosiglitazone (10 mg/kg) for 21 days did not display any marked improvement in their protective effect as compared to their effect per se in QA-treated animals which might be attributed to their ceiling effect. However, per se treatments of VPA, rosiglitazone, and their combinations were found to be non-significant as compared to sham group.

Effect of valproic acid, rosiglitazone, and their combination on TNF-α level in QA-treated animals. Data expressed as mean ± SEM. [percentage of sham (57.62 ± 2.3)]. (a) P < 0.05 compared to sham group, (b) P < 0.05 compared to control (QA-treated group), (c) P < 0.05 compared to Rosi (5), (d) P < 0.05 compared to VPA (100); (two-way ANOVA followed by Bonferroni’s test). QA Quinolinic acid, Rosi rosiglitazone, VPA valproic acid

Effect of valproic acid, rosiglitazone, and their combination on caspase-3 level in QA-treated animals. Data expressed as mean ± SEM [percentage of sham (percentage above control; 137.62 ± 7.3)]. (a) P < 0.05 compared to sham group, (b) P < 0.05 compared to control (QA-treated group), (c) P < 0.05 compared to Rosi (5), (d) P < 0.05 compared to VPA (100); (two-way ANOVA followed by Bonferroni’s test). QA Quinolinic acid, Rosi rosiglitazone, VPA valproic acid

Effect of valproic acid, rosiglitazone, and their combination on BDNF level in QA-treated animals. Data expressed as mean ± SEM [percentage of sham (153.58 ± 7.3)]. (a) P < 0.05 compared to sham group, (b) P < 0.05 compared to control (QA-treated group), (c) P < 0.05 compared to Rosi (5), (d) P < 0.05 compared to VPA (100); (two-way ANOVA followed by Bonferroni’s test). QA Quinolinic acid, Rosi rosiglitazone, VPA valproic acid
Effect of VPA, Rosiglitazone, and Their Combination on Striatal Lesion Volume in QA-Treated Animals
Intra-striatal QA (200 nmol) injection significantly increased striatal lesion volume as compared to the sham group, whereas treatment with of VPA (200 mg/kg) and rosiglitazone (10 mg/kg) for 21 days significantly reduced the QA-induced striatal lesion volume (Fig. 12). Furthermore, VPA (100 mg/kg) and rosiglitazone (5 mg/kg) treatment for 21 days could not produce any significant effect as compared to control (QA 200 nmol) group, whereas their combination treatment for 21 days, significantly attenuated striatal lesion size/volume which was found to be significant as compared to control (QA 200 nmol) as well as their effects per se in QA-treated animals (F = 9.52; P < 0.0001). Meanwhile, treatment of VPA (200 mg/kg) in combination with rosiglitazone (10 mg/kg) for 21 days did not display any marked improvement in the striatal lesion volume as compared to their effect per se in QA-treated animals which might be attributed to their ceiling effect. There was no marked difference observed in sham group as compared to naive animals. Similarly, per se treatments of VPA, rosiglitazone, and their combinations were found to be non-significant as compared to sham group.

Effect of valproic acid, rosiglitazone, and their combination on striatal lesion volume in QA-treated animals. Data expressed as mean ± SEM. (a) P < 0.05 compared to sham group, (b) P < 0.05 compared to control (QA-treated group), (c) P < 0.05 compared to Rosi (5), (d) P < 0.05 compared to VPA (100); (two-way ANOVA followed by Bonferroni’s test). QA Quinolinic acid, Rosi rosiglitazone, VPA valproic acid
Effect of VPA, Rosiglitazone, and Their Combination on Histopathological Analysis in QA-Treated Animals
Histopathological study was carried out on striatal and hippocampal sections of sham and drug-treated rat brains. Sham-treated striatal sections showed more number of optimum sized, undamaged, neuronal cells, and cell layers (Fig. 13a). However, neuronal inflammation and increased number of apoptotic cells are found in the striatal sections of QA-injected rats (Fig. 13b). However, VPA (100, 200 mg/kg) and rosiglitazone (5, 10 mg/kg) treatments attenuated these parameters as compared to QA-injected rats (Fig. 13c–f). Furthermore, treatment of VPA (100 mg/kg) in combination with rosiglitazone (5 mg/kg) showed further improvement in the histopathological characteristics (neuronal inflammation and increased number of apoptotic cells) as compared to their effects per se (Fig. 13g). Meanwhile, treatment of VPA (200 mg/kg) in combination with rosiglitazone (10 mg/kg) for 21 days did not display any marked modification in the histopathological characteristics as compared to their effect per se in QA-treated animals. However, the hippocampal sections did not show any visible alterations in the histopathological parameters (data not shown).

Effect of valproic acid, rosiglitazone, and their combination on histopathological analysis (H&E staining) in QA-treated animals representative photomicrographs of striatum of rat brain sections. Sections were stained with hematoxylin and eosin. Short arrows indicate neuro-inflammation and long arrows indicate apoptotic cells. a sham: neurons are intact, b QA (200 nmol): infiltration of inflammatory cells with large number of apoptotic cells, c Rosi (5): moderate inflammation of neurons with less apoptotic cells, d Rosi (10): mild inflammation of neurons with very less apoptotic cells, e VPA (100): moderate inflammation of neurons with less apoptotic cells, f VPA (200): mild inflammation of neurons with very less apoptotic cells, g Rosi (5) + VPA (100): mild inflammation of neurons with very less apoptotic cells, and h Rosi (10) + VPA (200): mild inflammation of neurons with very less apoptotic cells
Discussion
Single bilateral intrastriatal QA (200 nmol) administration produced HD-like symptoms in rats characterized by significant loss in body weight, motor incoordination, memory impairment, oxidative damage, mitochondrial (complex I, II, III, IV) dysfunction, alteration in the levels TNF-α, caspase-3, BDNF, acetylcholinesterase as well as striatal degeneration and histopathological alterations (Kalonia et al. 2010a, 2012; Rossato et al. 2002; Pemberton et al. 1997; Sanberg et al. 1989). Moreover, treatment with VPA, rosiglitazone, and their combination significantly attenuated these behavioral, biochemical, and cellular alterations suggesting possible synergistic interaction between HDAC and PPARγ pathways in HD-like conditions.
The use of QA in animal models of HD has been well established in our laboratory and has been shown to mimic HD-like symptoms as hyperactivity (Sanberg et al. 1989; Kalonia et al. 2010b, 2012), learning deficits (Block et al. 1993; Curtis et al. 1992; Furtado and Mazurek 1996), and loss of neurons when injected into the striatum of rats. In consistent with these reports, present study demonstrated marked motor impairment (decrease in locomotor activity and grip strength in rotarod test) following intrastriatal QA administration which are further supported by several findings of our own laboratory as well (Kalonia et al. 2010a; Shear et al. 1998). However, Jin and its co-workers reported that chronic administration of rosiglitazone (10 mg/kg/day, i.p.) significantly improved motor function in N171-82Q HD mice demonstrating the neuroprotective efficacy of rosiglitazone against motor abnormality in HD-like conditions (Jin et al. 2013). Consistent with these reports, marked attenuation in these motor impairments in QA-treated animals following VPA and rosiglitazone treatment suggest their neuroprotective potential in HD-like conditions. Furthermore, the combination of VPA and rosiglitazone at their lower doses revealed further improvement in their neuroprotective potential demonstrating possible synergizm, which was further supported by several study reports (Dash et al. 2010; Lee et al. 2012).
In Morris water maze paradigm, escape latency to reach the hidden platform was significantly increased, whereas in probe trial, the time spent in target quadrant, frequency of appearance in target quadrant and percentage of total path length traversed in target quadrant was significantly decreased in QA (200 nmol) administered animals as compared to sham group suggesting cognitive impairment. Several reports demonstrated that altered AChE activity in hippocampus modulates memory performance of the animals (Sudha et al. 1995; Baitharu et al. 2013). Similarly, in the present study, memory impairment following QA administration could be attributed to the increased hippocampal AChE levels, which were significantly ameliorated by rosiglitazone and VPA treatment demonstrating marked improvement in memory performance of the QA-treated animals. Although the lower doses were unable to demonstrate significant effect on the AChE levels, still they were able to demonstrate significant improvement in memory performance of the QA-treated animals which could be attributed to their beneficial effect on oxidative stress, neuro-inflammation, apoptosis, and BDNF levels. Our results are in accordance with the previous findings (Block et al. 1993), who found significant deficits in acquisition and probe trial performance in Morris water maze task in QA administered rats. These results are further supported by several earlier study reports demonstrating improvement in memory performance following VPA and rosiglitazone administration in several neurological conditions possibly via HDAC and PPARγ pathways, respectively, (Pedersen et al. 2006; Escribano et al. 2009). However, treatment of VPA in combination with rosiglitazone, showed further improvement in memory of the QA administered animals suggesting possible synergizm.
Several studies in our laboratory as well as other laboratories indicated that QA induces oxidative stress and support the possible relationship between excitotoxicity and oxidative process (Rossato et al. 2002; Ganzella et al. 2006; Kalonia et al. 2012). Studies have confirmed that QA induces LPO (Santamaria and Rios 1993) and neuronal damage that can be inhibited by neuroprotectants and anti-oxidants. Further, PPARγ agonists and HDAC inhibitors have been reported to enhance the activities of several anti-oxidant enzymes like superoxide dismutase-1 and -2 (SOD-1 and -2), catalase (Hwang et al. 2005; Ding et al. 2007). Further, the behavioral abnormalities in HD have been attributed to the death of dopaminergic and GABAergic neurons due to QA-induced excitotoxicity (Poeggeler et al. 2007) and other related cellular cascades such as oxidative stress (Perez-De La Cruz et al. 2012), mitochondrial dysfunction (Kalonia et al. 2010a), and neuro-inflammation. Recent reports suggest that rosiglitazone protected a neuroblastoma cell line (N2A) from mHtt-evoked mitochondrial deficiency as well as normalized the reactive oxygen species levels (Chiang et al. 2012). In consistent with these reports, the results of the present study demonstrated impairment in anti-oxidant defense (increase in LPO, nitrite concentration and decrease in SOD, catalase as well as glutathione enzyme activities) as well as mitochondrial complex (I, II, III, and IV) activities following QA administration, suggesting increased oxidative damage. However, treatment with rosiglitazone and VPA showed increased endogenous anti-oxidant defense, suggesting their protective effects against increased oxidative and nitrosative stress. Meanwhile, combined treatment with VPA, rosiglitazone at their lower doses showed a marked protection against QA-induced oxidative and nitrosative dysfunction which is further supported by the several reports (Valvassori et al. 2010; Kwan and Brodie 2001; Kalonia et al. 2012). As HDAC inhibition and PPARγ activation by VPA and rosiglitazone, respectively, ameliorated the QA-induced oxido-nitrosative stress and mitochondrial dysfunction, the PPARγ-HDAC interaction may be considered as a rational therapeutic target in the treatment HD-like symptoms (Quintanilla et al. 2008).
The role of HDAC in the pathogenesis of HD has recently been addressed showing a stark correlation between abnormal HDAC activity and disease progression (Langley et al. 2005; Sadri-Vakili and Cha 2006). Further, VPA treatment has been reported to decrease brain infarct volume and inhibit other inflammatory markers in the brain (Kim et al. 2007). In addition to this, VPA has also been reported to attenuate the secretion of TNF-α and interferon-γ (Chen et al. 2007; Peng et al. 2005). Recently, rosiglitazone has also been reported to attenuate dopaminergic cell loss in mouse model of Parkinson’s disease (PD), an effect attributed to its anti-inflammatory properties (Lee et al. 2012). Supplementary to this, several reports from our laboratory suggest that intrastriatal QA administration results in increased level of pro-inflammatory cytokines (TNF-α and IL-6) (Kalonia et al. 2012). In consistent with the above-mentioned study report, present study demonstrated a significant rise in the TNF-α levels following QA administration. However, treatment with VPA and rosiglitazone for 21 days restored TNF-α levels in different brain regions in QA-treated animals suggesting their anti-inflammatory potential against neuroinflammatory conditions. Further, treatment of VPA in combination with rosiglitazone for 21 days restored TNF-α level providing evidence toward possible synergizm between HDAC and PPARγ activities.
It has been well reported that striatal neuronal apoptosis is preferentially enhanced by NMDA receptor activation following excitotoxin injection (Shehadeh et al. 2006). Several other reports suggest that destruction of striatal GABAergic and dopaminergic neurons following excitotoxin injection are principally mediated through the apoptotic mechanisms (Simonian et al. 1996; Kalonia et al. 2011). In consistent with the above reports, intrastriatal QA administration in the present study resulted in the increased caspase-3 enzyme levels in striatum, cortex, and hippocampus. However, treatment with VPA and rosiglitazone showed a marked protection against QA-induced apoptosis as evident from the significantly restored caspase-3 level. Moreover, treatment of VPA in combination with rosiglitazone further attenuated the caspase-3 levels in the different brain areas suggesting that the PPARγ-HDAC interaction may play a crucial role in the management of HD-like conditions.
Neurotrophic factors are signaling molecules that mediate important physiological processes in the nervous system. BDNF deficiency is a major contributor to striatal degeneration and many phenotypes in HD. Maintenance and plasticity of striatal neurons are dependent on BDNF, which are depleted in HD striatum due to reduced expression and disrupted cortico-striatal transportation (Zuccato and Cattaneo 2007; Baquet et al. 2004). Several reports suggest that levels of BDNF are decreased especially in striatum, cortex, and hippocampus of as observed in post-mortem HD brains as well as in number of mouse and cellular models for HD (Zuccato and Cattaneo 2007; Gauthier et al. 2004; Ferrer et al. 2000; Jiang et al. 2013; Baquet et al. 2004). Jin et al. (2013) demonstrated that rosiglitazone (10 mg/kg i.p.) administration rescued BDNF deficiency in the cerebral cortex and prevented loss of orexin-A-immunopositive neurons in the hypothalamus of N171-82Q HD mice. However, BDNF improves pathology and delayed neuronal dysfunction in HD mice, and overexpression of BDNF in the striatum or administration of compounds increasing BDNF levels delayed the onset of motor dysfunction (Jin et al. 2013). In agreement with the above study reports, our results demonstrated considerably reduced BDNF levels following QA administration, which confirms cortical BDNF-dependent striatal vulnerability in rats. However, treatment with rosiglitazone, VPA and their combination resulted in substantial rescue in BDNF levels of QA-treated animals, which may be correlated with the positive impact on the behavior (motor function and memory performance) of the animals. Our study results are further supported by several reports which suggest that superior BDNF levels result in reversal of motor and cognitive dysfunction in neurodegenerative disorders (Samadi et al. 2013; Reiner et al. 2012; Wang et al. 2012).
Further, intrastriatal QA administration, caused a significant increase in the striatal lesion volume, which was subsequently reduced by the 21 days treatment with rosiglitazone, VPA and their combination (Kalonia et al. 2011). Finally, the histopathological analysis confirmed neuronal inflammation and increased number of apoptotic cells in the striatal sections of QA administered animals, which are remarkably altered by the treatment of VPA, rosiglitazone suggesting neuroprotection. These findings are further supported by the previous of our laboratory suggesting striatal histological alterations following QA administration (Kalonia et al. 2009). Further, the treatment of VPA in combination with rosiglitazone additionally improved these histological alterations showing remarkable neuroprotection.
Interestingly, reports suggest that VPA interacts with the PPARs (Lampen et al. 2001). In the non-neuronal tissue, valproate treatment has been found to increase the activity of peroxisomal enzymes (Van den Branden and Roels 1985) and alter the ultrastructure of the peroxisomes (Ponchaut et al. 1991), indicating possible modulation of PPAR signaling. It is an established fact that HDAC redistributes under oxidative stress, moving into the nucleus, and co-localizing with PPARγ in neuronal nuclei (Yang et al. 2011). Yang et al. (2011) demonstrated that transient transfection of cortical neurons with HDAC significantly reduced endogenous PPARγ transcriptional activity. Thus, HDAC inhibition results in activation of PPARγ activity in the neurons. In agreement with the published report, in the present study, the possible synergistic interaction between VPA and rosiglitazone could be attributed to the increased PPARγ activity following HDAC inhibition by VPA. However, further detailed studies are necessary before moving toward any final conclusion.
Taken altogether, the neuroprotective effect produced by VPA and rosiglitazone treatment might be attributed to the interplay of HDAC inhibition and PPARγ activation in QA-mediated oxidative stress, mitochondrial dysfunction, and neuroinflammatory-apoptotic signaling cascades. To our knowledge, this study is the first report showing the beneficial effects of the PPARγ agonist rosiglitazone, VPA, and their combination in QA-induced HD-like symptoms; the novelty of the present results is that we found synergizm between PPARγ agonist and HDAC inhibitors normalizing the behavioral, biochemical, cellular, and histopathological abnormality in QA-induced HD model.
Conclusion
The present study suggests the possible involvement of PPARγ–HDAC interaction in the neuroprotective effect of VPA, rosiglitazone. and their combination against QA-induced behavioral, biochemical, cellular. and histological alterations in rats, and further recommends that targeting HDAC and PPARγ pathways could be a useful approach in the management of HD-like symptoms. However, detailed investigations on interplay of these pathways are warranted before approaching toward any final implications.
References
Baitharu I, Jain V, Deep SN, Hota KB, Hota SK, Prasad D, Ilavazhagan G (2013) Withania somnifera root extract ameliorates hypobaric hypoxia induced memory impairment in rats. J Ethnopharmacol 145(2):431–441
Baquet ZC, Gorski JA, Jones KR (2004) Early striatal dendrite deficits followed by neuron loss with advanced age in the absence of anterograde cortical brain-derived neurotrophic factor. J Neurosci 24(17):4250–4258
Berman SB, Hastings TG (1999) Dopamine oxidation alters mitochondrial respiration and induces permeability transition in brain mitochondria: implications for Parkinson’s disease. J Neurochem 73(3):1127–1137
Bernardo A, Minghetti L (2008) Regulation of glial cell functions by PPAR-gamma natural and synthetic agonists. PPAR Res 2008:864140
Block F, Kunkel M, Schwarz M (1993) Quinolinic acid lesion of the striatum induces impairment in spatial learning and motor performance in rats. Neurosci Lett 149(2):126–128
Carta AR, Pisanu A, Carboni E (2011) Do PPAR-gamma agonists have a future in Parkinson’s disease therapy? Parkinsons Dis 2011:689181
Chen PS, Wang CC, Bortner CD, Peng GS, Wu X, Pang H, Lu RB, Gean PW, Chuang DM, Hong JS (2007) Valproic acid and other histone deacetylase inhibitors induce microglial apoptosis and attenuate lipopolysaccharide-induced dopaminergic neurotoxicity. Neuroscience 149(1):203–212
Chiang MC, Chern Y, Huang RN (2012) PPARgamma rescue of the mitochondrial dysfunction in Huntington’s disease. Neurobiol Dis 45(1):322–328
Choi DW (1992) Excitotoxic cell death. J Neurobiol 23(9):1261–1276
Cristiano L, Bernardo A, Ceru MP (2001) Peroxisome proliferator-activated receptors (PPARs) and peroxisomes in rat cortical and cerebellar astrocytes. J Neurocytol 30(8):671–683
Curtis CL, Ross GJ III, Hyde EA, Szymanski RE, Hull JS, Dunbar GL (1992) Intrastriatal injections of quinolinic acid cause spatial learning deficits in rats. Soc Neurosci Abstr 18:1602
Dash PK, Orsi SA, Zhang M, Grill RJ, Pati S, Zhao J, Moore AN (2010) Valproate administered after traumatic brain injury provides neuroprotection and improves cognitive function in rats. PLoS ONE 5(6):e11383
Ding G, Fu M, Qin Q, Lewis W, Kim HW, Fukai T, Bacanamwo M, Chen YE, Schneider MD, Mangelsdorf DJ, Evans RM, Yang Q (2007) Cardiac peroxisome proliferator-activated receptor gamma is essential in protecting cardiomyocytes from oxidative damage. Cardiovasc Res 76(2):269–279
Drew PD, Xu J, Storer PD, Chavis JA, Racke MK (2006) Peroxisome proliferator-activated receptor agonist regulation of glial activation: relevance to CNS inflammatory disorders. Neurochem Int 49(2):183–189
Ellman GL (1959) Tissue sulfhydryl groups. Arch Biochem Biophys 82(1):70–77
Ellman GL, Courtney KD, Andres V Jr, Feather-Stone RM (1961) A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol 7:88–95
Escribano L, Simon AM, Perez-Mediavilla A, Salazar-Colocho P, Del Rio J, Frechilla D (2009) Rosiglitazone reverses memory decline and hippocampal glucocorticoid receptor down-regulation in an Alzheimer’s disease mouse model. Biochem Biophys Res Commun 379(2):406–410
Escribano L, Simon AM, Gimeno E, Cuadrado-Tejedor M, de Maturana RL, Garcia-Osta A, Ricobaraza A, Perez-Mediavilla A, Del Rio J, Frechilla D (2010) Rosiglitazone rescues memory impairment in Alzheimer’s transgenic mice: mechanisms involving a reduced amyloid and tau pathology. Neuropsychopharmacology 35(7):1593–1604
Ferrante RJ, Kowall NW, Cipolloni PB, Storey E, Beal MF (1993) Excitotoxin lesions in primates as a model for Huntington’s disease: histopathologic and neurochemical characterization. Exp Neurol 119(1):46–71
Ferrante RJ, Kubilus JK, Lee J, Ryu H, Beesen A, Zucker B, Smith K, Kowall NW, Ratan RR, Luthi-Carter R, Hersch SM (2003) Histone deacetylase inhibition by sodium butyrate chemotherapy ameliorates the neurodegenerative phenotype in Huntington’s disease mice. J Neurosci Methods 23(28):9418–9427
Ferrer I, Goutan E, Marin C, Rey MJ, Ribalta T (2000) Brain-derived neurotrophic factor in Huntington disease. Brain Res 866(1–2):257–261
Furtado JC, Mazurek MF (1996) Behavioral characterization of quinolinate-induced lesions of the medial striatum: relevance for Huntington’s disease. Exp Neurol 138(1):158–168
Ganzella M, Jardim FM, Boeck CR, Vendite D (2006) Time course of oxidative events in the hippocampus following intracerebroventricular infusion of quinolinic acid in mice. Neurosci Res 55(4):397–402
Gardian G, Vecsei L (2004) Huntington’s disease: pathomechanism and therapeutic perspectives. J Neural Transm 111(10–11):1485–1494
Gaur V, Kumar A (2011) Neuroprotective potentials of candesartan, atorvastatin and their combination against stroke induced motor dysfunction. Inflammopharmacology 19(4):205–214
Gauthier LR, Charrin BC, Borrell-Pages M, Dompierre JP, Rangone H, Cordelieres FP, De Mey J, MacDonald ME, Lessmann V, Humbert S, Saudou F (2004) Huntingtin controls neurotrophic support and survival of neurons by enhancing BDNF vesicular transport along microtubules. Cell 118(1):127–138
Gornall AG, Bardawill CJ, David MM (1949) Determination of serum proteins by means of the biuret reaction. J Biol Chem 177(2):751–766
Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS, Tannenbaum SR (1982) Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal Biochem 126(1):131–138
Hashimoto R, Hough C, Nakazawa T, Yamamoto T, Chuang DM (2002) Lithium protection against glutamate excitotoxicity in rat cerebral cortical neurons: involvement of NMDA receptor inhibition possibly by decreasing NR2B tyrosine phosphorylation. J Neurochem 80(4):589–597
Heneka MT, Landreth GE, Hull M (2007) Drug insight: effects mediated by peroxisome proliferator-activated receptor-gamma in CNS disorders. Nat Clin Pract Neurol 3(9):496–504
Hwang J, Kleinhenz DJ, Lassegue B, Griendling KK, Dikalov S, Hart CM (2005) Peroxisome proliferator-activated receptor-gamma ligands regulate endothelial membrane superoxide production. Am J Physiol Cell Physiol 288(4):C899–C905
Jeong MR, Hashimoto R, Senatorov VV, Fujimaki K, Ren M, Lee MS, Chuang DM (2003) Valproic acid, a mood stabilizer and anticonvulsant, protects rat cerebral cortical neurons from spontaneous cell death: a role of histone deacetylase inhibition. FEBS Lett 542(1–3):74–78
Jiang M, Peng Q, Liu X, Jin J, Hou Z, Zhang J, Mori S, Ross CA, Ye K, Duan W (2013) Small-molecule TrkB receptor agonists improve motor function and extend survival in a mouse model of Huntington’s disease. Hum Mol Genet 22(12):2462–2470
Jin J, Albertz J, Guo Z, Peng Q, Rudow G, Troncoso JC, Ross CA, Duan W (2013) Neuroprotective effects of PPAR-γ agonist rosiglitazone in N171-82Q mouse model of Huntington’s disease. J Neurochem 125(3):410–419
Kalonia H, Kumar P, Kumar A, Nehru B (2009) Effect of caffeic acid and rofecoxib and their combination against intrastriatal quinolinic acid induced oxidative damage, mitochondrial and histological alterations in rats. Inflammopharmacology 17(4):211–219
Kalonia H, Kumar P, Kumar A (2010a) Pioglitazone ameliorates behavioral, biochemical and cellular alterations in quinolinic acid induced neurotoxicity: possible role of peroxisome proliferator activated receptor-γ (PPARγ) in Huntington’s disease. Pharmacol Biochem Behav 96(2):115–124
Kalonia H, Kumar P, Kumar A, Nehru B (2010b) Protective effect of montelukast against quinolinic acid/malonic acid induced neurotoxicity: possible behavioral, biochemical, mitochondrial and tumor necrosis factor-α level alterations in rats. Neuroscience 171(1):284–299
Kalonia H, Kumar P, Kumar A (2011) Attenuation of proinflammatory cytokines and apoptotic process by verapamil and diltiazem against quinolinic acid induced Huntington like alterations in rats. Brain Res 1372:115–126
Kalonia H, Mishra J, Kumar A (2012) Targeting neuro-inflammatory cytokines and oxidative stress by minocycline attenuates quinolinic-acid-induced Huntington’s disease-like symptoms in rats. Neurotox Res 22(4):310–320
Kiaei M, Kipiani K, Chen J, Calingasan NY, Beal MF (2005) Peroxisome proliferator–activated receptor-gamma agonist extends survival in transgenic mouse model of amyotrophic lateral sclerosis. Exp Neurol 191(2):331–336
Kim J-H, Kim S, Yoon I-S, Lee J-H, Jang B-J, Jeong SM, Lee J-H, Lee B-H, Han J-S, Oh S, Kim H-C, Park TK, Rhim H, Nah S-Y (2005) Protective effects of ginseng saponins on 3-nitropropionic acid-induced striatal degeneration in rats. Neuropharmacology 48(5):743–756
Kim HJ, Rowe M, Ren M, Hong JS, Chen PS, Chuang DM (2007) Histone deacetylase inhibitors exhibit anti-inflammatory and neuroprotective effects in a rat permanent ischemic model of stroke: multiple mechanisms of action. J Pharmacol Exp Ther 321(3):892–901
King TE (1967) Preparation of succinate dehydrogenase and reconstitution of succinate oxidase. In: Estabrook RW, Pullman ME (eds) Methods in enzymology, vol 10. Academic Press, New York, pp 322–331
King TE, Howard RL (1967) Preparations and properties of soluble NADH dehydrogenases from cardiac muscle. In: Estabrook RW, Pullman ME (eds) Methods in enzymololgy, vol 10. Academic Press, New York, pp 275–294
Kono Y (1978) Generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase. Archiv Biochem Biophy 186(1):189–195
Kukreja RC, Kontos HA, Hess ML, Ellis EF (1986) PGH synthase and lipoxygenase generate superoxide in the presence of NADH or NADPH. Circ Res 59(6):612–619
Kumar A, Prakash A, Pahwa D, Mishra J (2012a) Montelukast potentiates the protective effect of rofecoxib against kainic acid-induced cognitive dysfunction in rats. Pharmacol Biochem Behav 103(1):43–52
Kumar A, Sharma N, Gupta A, Kalonia H, Mishra J (2012b) Neuroprotective potential of atorvastatin and simvastatin (HMG-CoA reductase inhibitors) against 6-hydroxydopamine (6-OHDA) induced Parkinson-like symptoms. Brain Res 1471:13–22
Kumar A, Sharma N, Mishra J, Kalonia H (2013) Synergistical neuroprotection of rofecoxib and statins against malonic acid induced Huntington’s disease like symptoms and related cognitive dysfunction in rats. Eur J Pharmacol 709(1–3):1–12
Kwan P, Brodie MJ (2001) Neuropsychological effects of epilepsy and antiepileptic drugs. Lancet 357(9251):216–222
Lampen A, Carlberg C, Nau H (2001) Peroxisome proliferator-activated receptor delta is a specific sensor for teratogenic valproic acid derivatives. Eur J Pharmacol 431(1):25–33
Langley B, Gensert JM, Beal MF, Ratan RR (2005) Remodeling chromatin and stress resistance in the central nervous system: histone deacetylase inhibitors as novel and broadly effective neuroprotective agents. Curr Drug Targets CNS Neurol Disord 4(1):41–50
Lee EY, Lee JE, Park JH, Shin IC, Koh HC (2012) Rosiglitazone, a PPAR-gamma agonist, protects against striatal dopaminergic neurodegeneration induced by 6-OHDA lesions in the substantia nigra of rats. Toxicol Lett 213(3):332–344
Liu Y, Peterson DA, Kimura H, Schubert D (1997) Mechanism of cellular 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) reduction. J Neurochem 69(2):581–593
Luck H (1965) Catalase. In: Hans UB (ed) Methods of enzymatic analysis, 2nd edn. Academic Press, New York, pp 885–894
Mark RJ, Ashford JW, Goodman Y, Mattson MP (1995) Anticonvulsants attenuate amyloid beta-peptide neurotoxicity, Ca2+ deregulation, and cytoskeletal pathology. Neurobiol Aging 16:187–198
McGeer PL, McGeer EG (2008) Glial reactions in Parkinson’s disease. Mov Disord 23(4):474–483
Morris R (1984) Developments of a water-maze procedure for studying spatial learning in the rat. J Neurosci Methods 11(1):47–60
Park SW, Yi JH, Miranpuri G, Satriotomo I, Bowen K, Resnick DK, Vemuganti R (2007) Thiazolidinedione class of peroxisome proliferator-activated receptor gamma agonists prevents neuronal damage, motor dysfunction, myelin loss, neuropathic pain, and inflammation after spinal cord injury in adult rats. J Pharmacol Exp Ther 320:1002–1012
Paxinos G, Watson C (2007) The rat brain in stereotaxic coordinates, 6th edn. Academic Press, San Diego
Pedersen WA, McMillan PJ, Kulstad JJ, Leverenz JB, Craft S, Haynatzki GR (2006) Rosiglitazone attenuates learning and memory deficits in Tg2576 Alzheimer mice. Exp Neurol 199:265–273
Pemberton LA, Kerr SJ, Smythe G, Brew BJ (1997) Quinolinic acid production by macrophages stimulated with IFN-gamma, TNF-alpha, and IFN-alpha. J Interf Cytokine Res 17:589–595
Peng GS, Li G, Tzeng NS, Chen PS, Chuang DM, Hsu YD, Yang S, Hong JS (2005) Valproate pretreatment protects dopaminergic neurons from LPS-induced neurotoxicity in rat primary midbrain cultures: role of microglia. Brain Res 134(1):162–169
Perez-De La Cruz V, Carrillo-Mora P, Santamaria A (2012) Quinolinic acid, an endogenous molecule combining excitotoxicity, oxidative stress and other toxic mechanisms. Int J Tryptophan Res 5:1–8
Phiel CJ, Zhang F, Huang EY, Guenther MG, Lazar MA, Klein PS (2001) Histone deacetylase is a direct target of valproic acid, a potent anticonvulsant, mood stabilizer, and teratogen. J Biol Chem 276:36734–36741
Poeggeler B, Rassoulpour A, Wu HQ, Guidetti P, Roberts RC, Schwarcz R (2007) Dopamine receptor activation reveals a novel, kynurenate-sensitive component of striatal N-methyl-d-aspartate neurotoxicity. Neuroscience 148:188–197
Ponchaut S, Draye JP, Veitch K, Van Hoof F (1991) Influence of chronic administration of valproate on ultrastructure and enzyme content of peroxisomes in rat liver and kidney. Oxidation of valproate by liver peroxisomes. Biochem Pharmacol 41:1419–1428
Quintanilla RA, Jin YN, Fuenzalida K, Bronfman M, Johnson GV (2008) Rosiglitazone treatment prevents mitochondrial dysfunction in mutant huntingtin-expressing cells: possible role of peroxisome proliferator-activated receptor-gamma (PPARgamma) in the pathogenesis of Huntington disease. J Biol Chem 283:25628–25637
Reiner A, Wang HB, Del Mar N, Sakata K, Yoo W, Deng YP (2012) BDNF may play a differential role in the protective effect of the mGluR2/3 agonist LY379268 on striatal projection neurons in R6/2 Huntington’s disease mice. Brain Res 1473:161–172
Rossato JI, Zeni G, Mello CF, Rubin MA, Rocha JB (2002) Ebselen blocks the quinolinic acid-induced production of thiobarbituric acid reactive species but does not prevent the behavioral alterations produced by intra-striatal quinolinic acid administration in the rat. Neurosci Lett 318:137–140
Sadri-Vakili G, Cha JH (2006) Histone deacetylase inhibitors: a novel therapeutic approach to Huntington’s disease (complex mechanism of neuronal death). Curr Alzheimer Res 3(4):403–408
Samadi P, Boutet A, Rymar VV, Rawal K, Maheux J, Kvann JC, Tomaszewski M, Beaubien F, Cloutier JF, Levesque D, Sadikot AF (2013) Relationship between BDNF expression in major striatal afferents, striatum morphology and motor behavior in the R6/2 mouse model of Huntington’s disease. Genes Brain Behav 12(1):108–124
Sanberg PR, Calderon SF, Giordano M, Tew JM, Norman AB (1989) The quinolinic acid model of Huntington’s disease: locomotor abnormalities. Exp Neurol 105(1):45–53
Santamaria A, Rios C (1993) MK-801, an N-methyl-d-aspartate receptor antagonist, blocks quinolinic acid-induced lipid peroxidation in rat corpus striatum. Neurosci Lett 159(1–2):51–54
Schwarcz R, Whetsell WO Jr, Mangano RM (1983) Quinolinic acid: an endogenous metabolite that produces axon-sparing lesions in rat brain. Science 219(4582):316–318
Schwarcz R, Guidetti P, Sathyasaikumar KV, Muchowski PJ (2010) Of mice, rats and men: revisiting the quinolinic acid hypothesis of Huntington’s disease. Prog Neurobiol 90(2):230–245
Shear DA, Dong J, Gundy CD, Haik-Creguer KL, Dunbar GL (1998) Comparison of intrastriatal injections of quinolinic acid and 3-nitropropionic acid for use in animal models of Huntington’s disease. Prog Neuro-psychopharmacol Biol Psychiatry 22(7):1217–1240
Shehadeh J, Fernandes HB, Mullins MMZ, Graham RK, Leavitt BR, Hayden MR, Raymond LA (2006) Striatal neuronal apoptosis is preferentially enhanced by NMDA receptor activation in YAC transgenic mouse model of Huntington disease. Neurobiol Dis 21(2):392–403
Simonian NA, Getz RL, Leveque JC, Konradi C, Coyle JT (1996) Kainic acid induces apoptosis in neurons. Neuroscience 75:1047–1055
Sottocasa GL, Kuylenstierna B, Ernster L, Bergstrand A (1967) An electron-transport system associated with the outer membrane of liver mitochondria. A biochemical and morphological study. J Cell Biol 32:415–438
Sudha S, Lakshmana MK, Pradhan N (1995) Chronic phenytoin induced impairment of learning and memory with associated changes in brain acetylcholine esterase activity and monoamine levels. Pharmacol Biochem Behav 52(1):119–124
Tuzcu M, Baydas G (2006) Effect of melatonin and vitamin E on diabetes-induced learning and memory impairment in rats. Eur J Pharmacol 537(1–3):106–110
Valvassori SS, Rezin GT, Ferreira CL, Moretti M, Goncalves CL, Cardoso MR, Streck EL, Kapczinski F, Quevedo J (2010) Effects of mood stabilizers on mitochondrial respiratory chain activity in brain of rats treated with d-amphetamine. J Psychiatr Res 44(14):903–909
Van den Branden C, Roels F (1985) Peroxisomal beta-oxidation and sodium valproate. Biochem Pharmacol 34(12):2147–2149
Verma R, Mishra V, Gupta K, Sasmal D, Raghubir R (2011) Neuroprotection by rosiglitazone in transient focal cerebral ischemia might not be mediated by glutamate transporter-1. J Neurosci Res 89:1849–1858
Wang L, Ashley-Koch A, Steffens DC, Krishnan KR, Taylor WD (2012) Impact of BDNF Val66Met and 5-HTTLPR polymorphism variants on neural substrates related to sadness and executive function. Genes Brain Behav 11(3):352–359
Wills ED (1966) Mechanisms of lipid peroxide formation in animal tissues. Biochem J 99(3):667–676
Yang Y, Qin X, Liu S, Li J, Zhu X, Gao T, Wang X (2011) Peroxisome proliferator-activated receptor γ is inhibited by histone deacetylase 4 in cortical neurons under oxidative stress. J Neurochem 118(3):429–439
Zadori D, Geisz A, Vamos E, Vecsei L, Klivenyi P (2009) Valproate ameliorates the survival and the motor performance in a transgenic mouse model of Huntington’s disease. Pharmacol Biochem Behav 94(1):148–153
Zahler WL, Cleland WW (1968) A specific and sensitive assay for disulfides. J Biol Chem 243(4):716–719
Zuccato C, Cattaneo E (2007) Role of brain-derived neurotrophic factor in Huntington’s disease. Prog Neurobiol 81(5–6):294–330
Acknowledgments
The Major Research Project sanctioned to Professor Anil Kumar by University Grants Commission (UGC), New Delhi is gratefully acknowledged. Mr. Jitendriya Mishra is the project fellow in this project.
Conflict of interest
The authors have no competing financial interests to declare. There is no conflict of interest between any of the authors.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mishra, J., Chaudhary, T. & Kumar, A. Rosiglitazone Synergizes the Neuroprotective Effects of Valproic Acid Against Quinolinic Acid-Induced Neurotoxicity in Rats: Targeting PPARγ and HDAC Pathways. Neurotox Res 26, 130–151 (2014). https://doi.org/10.1007/s12640-014-9458-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12640-014-9458-z