
Abstract
Eribulin mesylate (E7389, INN:eribulin mesilate Halaven®) is a non-taxane microtubule dynamics inhibitor currently in clinical use for advanced breast cancer. Other microtubule-targeting agents for breast cancer, including paclitaxel and ixabepilone, display a common treatment dose-limiting toxicity of peripheral neuropathy (PN). In an earlier study, we found eribulin mesylate had a lower propensity to induce PN in mice than either paclitaxel or ixabepilone. In the current study, we compared additional PN induced by paclitaxel versus eribulin mesylate when administered to mice with preexisting paclitaxel-induced PN. Initially, paclitaxel at 0.75 × its maximum tolerated dose (MTD; 22.5 mg/kg) was given on a Q2Dx3 regimen for 2 weeks. The second chemotherapy was 0.5 MTD eribulin mesylate (0.875 mg/kg) or paclitaxel (15 mg/kg) on a similar regimen, starting 2 weeks after the first. Initial paclitaxel treatment produced significant decreases in caudal nerve conduction velocity (NCV; averaging 19.5 ± 1 and 22.2 ± 1.3 %, p < 0.001) and amplitude (averaging 53.2 ± 2.6 and 72.4 ± 2.1 %, p < 0.001) versus vehicle when measured 24 h or 2 weeks after dosing cessation, respectively. Additional 0.5 MTD paclitaxel further reduced caudal NCV and amplitude relative to immediately before initiation of the second regimen (by 11 ± 2.1 and 59.2 ± 5 %, p < 0.01, respectively). In contrast, 0.5 MTD eribulin mesylate caused no further decrease in caudal NCV. In conclusion, unlike additional paclitaxel treatment, eribulin mesylate administered to mice with preexisting paclitaxel-induced PN had limited additional deleterious effects at 6 weeks. These preclinical data suggest that eribulin mesylate may have reduced tendency to exacerbate preexisting paclitaxel-induced PN in clinical settings.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Paclitaxel is a microtubule dynamics inhibitor widely used in cancer chemotherapy. It was originally isolated from the bark of the Pacific yew tree, Taxus brevifolia, and was subsequently developed commercially by Bristol-Myer Squibb and sold under the trademark Taxol. Paclitaxel stabilizes microtubules, and as a result, interferes with normal microtubule breakdown during cell division, resulting in suppression of microtubule dynamics (Wilson and Jordan 2004). Paclitaxel is approved as a treatment for patients with lung, ovarian, breast, head and neck cancer, and advanced forms of Kaposis sarcoma (Rowinsky 1993; Rowinsky et al. 1993a, b), however, its effectiveness is limited by the development of severe peripheral neuropathy (PN) that appears to be dose-dependent (Cavaletti et al. 1995a, b; Mimura et al. 2000). The incidence of neuropathy in patients receiving paclitaxel is estimated to be 50–90 % and is characterized by dysesthesia (numbness, tingling and burning pain) of the hands and feet (Cavaletti et al. 1995b; Rowinsky 1993; Rowinsky et al. 1993a; Wiernik et al. 1987). A number of rodent models of paclitaxel-induced neuropathy have also been developed (Apfel et al. 1991; Authier et al. 2000; Boyle et al. 1999; Cavaletti et al. 1995b; Cliffer et al. 1998; Dina et al. 2001; Polomano et al. 2001; Wozniak et al. 2011), which may also have clinical relevance.
Eribulin mesylate (E7389) is a non-taxane, structurally simplified, synthetic analogue of the naturally occurring anticancer agent, halichondrin B (Mani and Swami 2010). Eribulin inhibits microtubule dynamics via a novel mechanism of action (Dabydeen et al. 2006; Jordan et al. 2005; Okouneva et al. 2008), thought to involve binding to a unique tubulin binding site (Dabydeen et al. 2006) resulting in the suppression of microtubule polymerization together with sequestration of tubulin into non-functional aggregates, causing irreversible mitotic block and apoptosis (Dabydeen et al. 2006; Jordan et al. 2005; Okouneva et al. 2008). Its encouraging preclinical activity prompted its initial entry into clinical trial in 2002 and its ultimate FDA approval in 2010 for metastatic breast cancer patients who have previously received at two lines of chemotherapy for advanced disease and have been treated with anthracycline and a taxane in either the adjuvant or metastatic setting (Jain and Vahdat 2011).
In the clinical setting, there have been reports of eribulin being associated with a relatively low incidence of neuropathy as compared to paclitaxel and ixabepilone, especially the more severe grades (Jain and Vahdat 2011; Swami et al. 2012; Vahdat et al. 2009). In support of this, we also showed that eribulin mesylate had a lesser tendency to induce nerve conduction deficits in naïve mice than did paclitaxel or ixabepilone (Wozniak et al. 2011).
Neuropathy from a chemotherapy treatment can be serious, cumulative and long-lasting and is often the dose-limiting toxicity that forces patients to lower doses of chemotherapy drugs or even discontinue further treatment, with possible long-term effects on patient survival. This is further confounded because patients can often receive multiple regimens consisting of different chemotherapies. In an attempt to replicate this situation preclinically, we describe a novel model utilizing two consecutive chemotherapy regimens administered to mice. Initially, we induced a paclitaxel neuropathy, as evidenced by significant and maintained reduction in caudal nerve conduction and amplitude measures. Thereafter, a second chemotherapy regimen was dosed consisting of further paclitaxel or eribulin mesylate, after which caudal nerve conduction velocity (NCV) and amplitude were monitored and compared. We report that a second chemotherapy regimen of eribulin mesylate produced less neuropathy (as assessed by nerve conduction and amplitude changes) than did paclitaxel in mice with a residual paclitaxel-induced neuropathy. This may have implications for the clinical setting.
Methods
Drugs and Formulations
Eribulin mesylate (synthesized at Eisai Research Institute and stored at −80 °C in the dark) was dissolved in 100 % anhydrous DMSO (Sigma-Aldrich, St. Louis, MO) to produce a 10 mg/ml stock solution, which was separated into aliquots and stored at −80 °C until day of administration. Each administration day the stock solution was thawed and diluted with saline to a final concentration of 0.25 mg/mL in 2.5 % DMSO/97.5 % saline yielding dosing solutions in a 10 mL/kg volume.
Paclitaxel, purchased from LC Laboratories, Woburn, MA and stored at −20 °C, in the dark, was dissolved in ethanol (100 %) at 10 % of final volume. An equal volume of cremophor (10 % of final volume) was then added and the mixture re-vortexed for about 10 min. Immediately prior to injection, ice cold saline was added to final volume (as 80 % of final), and the solution was maintained on ice during dosing. Dosing solutions were made fresh daily and dosed in a volume of 10 mL/kg.
Animals
Female BALB/c mice (approximately 7–8 weeks old at onset of dosing) were obtained from Harlan Laboratories Inc. (Indianapolis, IN) and maintained with free access to water and a standardized synthetic diet (Harlan Teklab). Animal housing and procedure room temperature and humidity were maintained at 20 ± 2 °C and 55 ± 10 %, respectively. Artificial lighting provided a 12 h light/12 h dark cycle (light 7 a.m.–7 p.m.).
All experimental protocols were approved by the Institutional Animal Care and Use Committee of Sobran Inc and adhered to all of the applicable institutional and governmental guidelines for the humane treatment of laboratory animals.
Nerve Conduction Velocity and Amplitude Measurement
Baseline NCV was measured 1 week prior to initiation of dosing in all mice. Mice were anesthetized with 2 % isoflurane (by inhalation, for induction and maintenance) and placed on a heating pad with rectal temperature monitored and maintained between 37.0 and 40.0 °C. Platinum subdermal needle electrodes (Grass Technologies, West Warwick, RI) were used. Each nerve segment stimulation was repeated a total of at least 3 times, up to a maximum of 6 times, with increasing voltage until the maximal response had been achieved, using AcqKnowledge software, version 3.7.3 (BIOPAC Systems Inc.). Mice were assigned into a vehicle (10 mice) or paclitaxel treatment group (20 mice). Paclitaxel was administered on a Q2Dx3 for 2 weeks schedule at what was earlier determined to be 0.75 MTD (Wozniak et al. 2011). After completion of this treatment, mice were again tested for nerve conduction (24 h after last dosing, Week 2) and left for 2 weeks before retesting (Week 4). At this time, mice having received initial paclitaxel were re-assigned to treatment groups of 10 mice each, to receive either additional paclitaxel (at 0.5 MTD) or eribulin mesylate (at 0.5 MTD). All mice were again tested 24 h after the last dosing of this second regimen (Week 6).
A separate group of mice which received a single 0.75 MTD paclitaxel regimen followed by vehicle only were studied for 10 weeks.
Differences in measurements across groups were determined by direct statistical comparisons (ANOVA, followed by Tukey’s post hoc comparisons; using Prism Graphpad Software, Version: 4.03). Statistical significance was defined as p < 0.05.
Sciatic Nerve Histology
Immediately after recording of final nerve conduction parameters, five randomly selected mice from each group were chosen for nerve morphological analysis. Mice were deeply anesthetized with 10 % chloral hydrate and euthanized by transcardial perfusion with 1× PBS followed by 2 % paraformaldehyde in 0.1 mM phosphate buffer, pH 7.4, for 10–15 min. Sciatic nerves were dissected out at mid-thigh level and postfixed in osmium tetroxide and embedded in Epon. Cross-sections, 1 μm thickness, were stained with toluidine blue and examined under light microscopy. Images covering the whole cross-section stained for each nerve were taken at 40× and tiled. Total degenerating profiles of myelinated axons were counted in tiled montages of each nerve (n = 3–5 per treatment).
Results
Paclitaxel at 0.75 MTD Causes Neuropathy in Mice
Paclitaxel administered at 0.75 MTD (22.5 mg/kg) on a Q2Dx3 for 2 weeks regimen produced a significant deficit in caudal NCV and amplitude [17.4 ± 1.4 and 17.9 ± 2.5 % deficit in NCV and 50.2 ± 4.2 and 44.2 ± 5.8 % deficit in amplitude for groups 1 and 2, respectively, compared to vehicle administration (a 2.3 ± 1.2 % deficit); Fig. 1, week 2 vs. baseline]. This effect persisted for 2 weeks after dosing, as evidenced by maintained NCV and amplitude deficits at week 4 (19.6 ± 1.9 and 17.9 ± 2.7 and 73.9 ± 2.1 and 65.9 ± 5.4 % vs. baseline for groups 1 and 2, respectively. Figure 1, week 4).

Effect of two chemotherapy regimens on mouse caudal nerve conduction velocity (a) and amplitude (b). Paclitaxel (0.75 MTD) administered to mice on a Q2Dx3 for 2 weeks regimen produced a significant decrease in caudal velocity and amplitude (Week 2). This effect persisted throughout an additional 2 week period, despite no additional dosing (Week 4). Thereafter, mice were treated with additional paclitaxel or eribulin mesylate, each administered at 0.5 MTD. Paclitaxel induced additional deficits in nerve conduction velocity and amplitude, with a significantly greater velocity deficit in the paclitaxel/paclitaxel group than in the paclitaxel/eribulin group at 6 weeks (p < 0.05), while additional eribulin mesylate had no significant additional effect (A and B; week 6 vs. week 4)
Additional 0.5 MTD Paclitaxel, but not Eribulin Mesylate, Causes Further Deleterious Effects When Dosed in Mice with Preexisting Neuropathy
Additional 0.5 MTD paclitaxel given to animals with preexisting neuropathy induced further adverse effects, as indicated by additional deficits in NCV and amplitude of 9.5 ± 2.1 and 54.0 ± 5 % respectively, (p < 0.01; Fig. 1, week 6 vs. week 4).
In contrast, eribulin mesylate at 0.5 MTD given to mice with preexisting neuropathy caused no additional significant deficits in caudal NCV or caudal amplitude (p > 0.05; Figs. 1, week 6 vs. week 4). At 6 weeks, the deficit in conduction velocity caused by additional paclitaxel in paclitaxel pretreated mice was significantly more severe than that induced by additional eribulin (p < 0.05) in mice with a preexisting paclitaxel neuropathy. Similar findings, although not significant, were apparent for amplitude (Fig. 1, week 6).
Paclitaxel at 0.75 MTD Induces NCV Deficits Which are Reversible, While Amplitude Deficits are Maintained
To ensure that the week 6 and 10 deficits in NCV and amplitude observed were impacted by the second chemotherapy regimen and not just a consequence of the initial paclitaxel treatment, a separate study was conducted in which animals were treated with 0.75 MTD paclitaxel regimen followed by a second regimen of vehicle. In these mice, the NCV deficits seen at week 4 completely recovered by week 6 (Fig. 2a). However, the amplitude deficit seen at 4 weeks stayed constant through week 10 (Fig. 2b). These data suggest that the amplitude deficits induced by the initial paclitaxel regimen are long-lasting and may have partially impacted the recordings at week 6 obtained in mice receiving the dual chemotherapy regimens.

Long-term effects of a single paclitaxel regimen on caudal nerve velocity (a) and amplitude (b). The deficit in conduction velocity induced by a single regimen of paclitaxel (0.75 MTD) is shown in (a) and appeared recovered by week 6, while the amplitude deficit was maintained through week 10 (b)
Paclitaxel and Eribulin Mesylate Cause Similar Morphological Changes When Administered to Mice with Preexisting Neuropathy
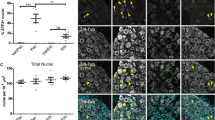
We previously showed that paclitaxel causes degeneration of myelinated axons in the sciatic nerve (Wozniak et al. 2011). To compare the effect of paclitaxel and eribulin in mice with preexisting neuropathy, we examined signs of degeneration in the sciatic nerves. When a myelinated axon degenerates, the axoplasm disintegrates quickly, within the first 48 h, and the myelin sheath breakdowns to smaller segments termed “ovoids” (Griffin et al. 1993), thus leaving behind a “footprint” of a degenerated axon. In 1 μm nerve cross-sections, these degenerated axons are interspersed with intact axons in nerves treated with a single regimen of paclitaxel (Wozniak et al. 2011). In this report, we counted the profiles of myelin ovoids in whole cross-sectional areas of the sciatic nerves to determine whether a second treatment of paclitaxel or eribulin mesylate exacerbates nerve degeneration. The number of degenerated profiles was not statistically different between paclitaxel and eribulin mesylate treated mice with preexisting neuropathy (Fig. 3).

Morphological analysis of mice with preexisting neuropathy (a–e). After perfusion, sciatic nerves at mid-thigh were harvested, processed for toluidine blue staining, and analyzed for axonal degeneration. (a–d) show cross-sectioned semi-thin (1 μm) plastic sections from nerve. (a) shows a representative nerve section from mouse receiving veh/veh treatment showing no axonal degeneration whereas nerve from paclitaxel/vehicle treated mouse show degeneration of myelinated axons (b). In (c), nerves from mice treated with paclitaxel plus paclitaxel show more degeneration of myelinated axons, of similar magnitude to that seen in nerves from mice treated with paclitaxel followed by eribulin (d). (e) shows the actual quantification numbers of degenerated myelinated axons in the whole cross-sectional area of the nerves. Values are ± SEM. N = 5 for each treatment, except vehicle/vehicle group, where n = 3
Discussion
Taxanes are among the most effective and commonly used antineoplastic drugs for solid tumor treatment, but their use is limited by peripheral neurotoxicity (Guastalla and Dieras 2003; Kuroi and Shimozuma 2004). The exact mechanism of the neurotoxicity is unknown (Hagiwara and Sunada 2004) and to date there are no acceptable therapies that prevent or minimize the nerve damage (Gordon et al. 1997; Rowinsky et al. 1993b; Wasserheit et al. 1996). Paclitaxel is known to cause decreased nerve conduction velocities in sensory nerves (Lipton et al. 1989) and elevations in vibratory and thermal thresholds (Wiernik et al. 1987), suggesting axonal degeneration and demyelination in the sensory nervous system with effects appearing to be dose and schedule dependent (Mimura et al. 2000). Previous models of paclitaxel-induced neuropathy in rodents have also demonstrated sensory neuropathy (Apfel et al. 1991; Cavaletti et al. 1997; Cliffer et al. 1998). The use of animal models is a well-established method for investigating various aspects of chemotherapy-induced neuropathy (Cavaletti et al. 1995b; Cliffer et al. 1998; Pisano et al. 2003; Schmidt et al. 1995; Wang et al. 2004). We previously implemented a model to compare the neuropathy-inducing propensities of three microtubule-targeting chemotherapies, paclitaxel, ixabepilone, and eribulin mesylate, each administered on a similar schedule (Q2Dx3 for 2 weeks) using an MTD basis (Wozniak et al. 2011). Ixabepilone and paclitaxel were found to produce significant deficits in conduction parameters of caudal and digital nerves at 0.5 MTD doses and above. In contrast, eribulin mesylate induced no deficits in any monitored nerve conduction parameter at any dose tested, including MTD (1.75 mg/kg). Similarly, although all compounds produced morphological changes in dorsal root ganglion (DRG) and sciatic nerves at MTD, the changes induced by eribulin mesylate were noticeably fewer and less severe than those seen with paclitaxel or ixabepilone at their respective MTDs. The reason for this difference is unclear although differences in microtubule-binding properties may have significant effects on the toxicity profile of each agent (Authier et al. 2009; Perez 2009). This possibility is currently under further investigation.
In the current investigation, we extended our initial studies to investigate the effects of an additional chemotherapy regimen in mice with a preexisting paclitaxel-induced neuropathy. To our knowledge, this is the first preclinical study undertaking this complex and fairly common clinical scenario. We report that eribulin mesylate had minimal deleterious effects on caudal nerve conduction in mice with preexisting paclitaxel neuropathy, whereas a second regimen of paclitaxel caused additional nerve conduction and amplitude deficits. Despite the functional differences seen in the two experimental groups, there were no apparent morphology differences. When paclitaxel and eribulin were dosed to animals with preexisting paclitaxel neuropathy, both groups showed similar numbers of degenerating fibers and unmyelinated axons. This could be because the initial paclitaxel regimen that was common to both treatment groups produced permanent or long-lasting changes which persisted through the second chemotherapy regimen. This is supported by the evidence of persistent amplitude changes following the single 0.75 MTD paclitaxel regimen, determined in a separate group of mice.
It is also possible that eribulin causes similar neuropathy, but with a slower development than does additional paclitaxel in mice with preexisting neuropathy. In order to ascertain this, the neuropathy needs to be followed out for a longer period of time.
Clinically, the scenario of a chemotherapy regimen being administered to patients with a residual neuropathy induced by an earlier regimen is relatively common. If our preclinical findings in mice translate into the human arena, we predict that a patient treated with paclitaxel may suffer less additional or slower onset, neuropathy-related side effects if treated with a subsequent regimen of the antineoplastic eribulin, as opposed to paclitaxel.
References
Apfel SC, Lipton RB, Arezzo JC, Kessler JA (1991) Nerve growth factor prevents toxic neuropathy in mice. Ann Neurol 29:87–90
Authier N, Gillet JP, Fialip J, Eschalier A, Coudore F (2000) Description of a short-term taxol-induced nociceptive neuropathy in rats. Brain Res 887:239–249
Authier N, Balayssac D, Marchand F, Ling B, Zangarelli A, Descoeur J, Coudore F, Bourinet E, Eschalier A (2009) Animal models of chemotherapy-evoked painful peripheral neuropathies. Neurotherapeutics 6:620–629
Boyle FM, Wheeler HR, Shenfield GM (1999) Amelioration of experimental cisplatin and paclitaxel neuropathy with glutamate. J Neurooncol 41:107–116
Cavaletti G, Bogliun G, Marzorati L, Zincone A, Marzola M, Colombo N, Tredici G (1995a) Peripheral neurotoxicity of taxol in patients previously treated with cisplatin. Cancer 75:1141–1150
Cavaletti G, Tredici G, Braga M, Tazzari S (1995b) Experimental peripheral neuropathy induced in adult rats by repeated intraperitoneal administration of taxol. Exp Neurol 133:64–72
Cavaletti G, Cavalletti E, Montaguti P, Oggioni N, De Negri O, Tredici G (1997) Effect on the peripheral nervous system of the short-term intravenous administration of paclitaxel in the rat. Neurotoxicology 18:137–145
Cliffer KD, Siuciak JA, Carson SR, Radley HE, Park JS, Lewis DR, Zlotchenko E, Nguyen T, Garcia K, Tonra JR, Stambler N, Cedarbaum JM, Bodine SC, Lindsay RM, DiStefano PS (1998) Physiological characterization of Taxol-induced large-fiber sensory neuropathy in the rat. Ann Neurol 43:46–55
Dabydeen DA, Burnett JC, Bai R, Verdier-Pinard P, Hickford SJ, Pettit GR, Blunt JW, Munro MH, Gussio R, Hamel E (2006) Comparison of the activities of the truncated halichondrin B analog NSC 707389 (E7389) with those of the parent compound and a proposed binding site on tubulin. Mol Pharmacol 70:1866–1875
Dina OA, Chen X, Reichling D, Levine JD (2001) Role of protein kinase Cepsilon and protein kinase A in a model of paclitaxel-induced painful peripheral neuropathy in the rat. Neuroscience 108:507–515
Gordon AN, Stringer CA, Matthews CM, Willis DL, Nemunaitis J (1997) Phase I dose escalation of paclitaxel in patients with advanced ovarian cancer receiving cisplatin: rapid development of neurotoxicity is dose-limiting. J Clin Oncol 15:1965–1973
Griffin JW, George R, Ho T (1993) Macrophage systems in peripheral nerves. A review. J Neuropathol Exp Neurol 52:553–560
Guastalla JP 3rd, Dieras V (2003) The taxanes: toxicity and quality of life considerations in advanced ovarian cancer. Br J Cancer 89(Suppl 3):S16–S22
Hagiwara H, Sunada Y (2004) Mechanism of taxane neurotoxicity. Breast Cancer 11:82–85
Jain S, Vahdat LT (2011) Eribulin mesylate. Clin Cancer Res 17:6615–6622
Jordan MA, Kamath K, Manna T, Okouneva T, Miller HP, Davis C, Littlefield BA, Wilson L (2005) The primary antimitotic mechanism of action of the synthetic halichondrin E7389 is suppression of microtubule growth. Mol Cancer Ther 4:1086–1095
Kuroi K, Shimozuma K (2004) Neurotoxicity of taxanes: symptoms and quality of life assessment. Breast Cancer 11:92–99
Lipton RB, Apfel SC, Dutcher JP, Rosenberg R, Kaplan J, Berger A, Einzig AI, Wiernik P, Schaumburg HH (1989) Taxol produces a predominantly sensory neuropathy. Neurology 39:368–373
Mani S, Swami U (2010) Eribulin mesilate, a halichondrin B analogue, in the treatment of breast cancer. Drugs Today (Barc) 46:641–653
Mimura Y, Kato H, Eguchi K, Ogawa T (2000) Schedule dependency of paclitaxel-induced neuropathy in mice: a morphological study. Neurotoxicology 21:513–520
Okouneva T, Azarenko O, Wilson L, Littlefield BA, Jordan MA (2008) Inhibition of centromere dynamics by eribulin (E7389) during mitotic metaphase. Mol Cancer Ther 7:2003–2011
Perez EA (2009) Microtubule inhibitors: differentiating tubulin-inhibiting agents based on mechanisms of action, clinical activity, and resistance. Mol Cancer Ther 8:2086–2095
Pisano C, Pratesi G, Laccabue D, Zunino F, Lo Giudice P, Bellucci A, Pacifici L, Camerini B, Vesci L, Castorina M, Cicuzza S, Tredici G, Marmiroli P, Nicolini G, Galbiati S, Calvani M, Carminati P, Cavaletti G (2003) Paclitaxel and Cisplatin-induced neurotoxicity: a protective role of acetyl-L-carnitine. Clin Cancer Res 9:5756–5767
Polomano RC, Mannes AJ, Clark US, Bennett GJ (2001) A painful peripheral neuropathy in the rat produced by the chemotherapeutic drug, paclitaxel. Pain 94:293–304
Rowinsky EK (1993) Clinical pharmacology of Taxol. J Natl Cancer Inst Monogr 15:25–37
Rowinsky EK, Chaudhry V, Cornblath DR, Donehower RC (1993a) Neurotoxicity of Taxol. J Natl Cancer Inst Monogr 15:107–115
Rowinsky EK, Eisenhauer EA, Chaudhry V, Arbuck SG, Donehower RC (1993b) Clinical toxicities encountered with paclitaxel (Taxol). Semin Oncol 20:1–15
Schmidt Y, Unger JW, Bartke I, Reiter R (1995) Effect of nerve growth factor on peptide neurons in dorsal root ganglia after taxol or cisplatin treatment and in diabetic (db/db) mice. Exp Neurol 132:16–23
Swami U, Chaudhary I, Ghalib MH, Goel S (2012) Eribulin—a review of preclinical and clinical studies. Crit Rev Oncol Hematol 81:163–184
Vahdat LT, Pruitt B, Fabian CJ, Rivera RR, Smith DA, Tan-Chiu E, Wright J, Tan AR, Dacosta NA, Chuang E, Smith J, O’Shaughnessy J, Shuster DE, Meneses NL, Chandrawansa K, Fang F, Cole PE, Ashworth S, Blum JL (2009) Phase II study of eribulin mesylate, a halichondrin B analog, in patients with metastatic breast cancer previously treated with an anthracycline and a taxane. J Clin Oncol 27:2954–2961
Wang MS, Davis AA, Culver DG, Wang Q, Powers JC, Glass JD (2004) Calpain inhibition protects against Taxol-induced sensory neuropathy. Brain 127:671–679
Wasserheit C, Frazein A, Oratz R, Sorich J, Downey A, Hochster H, Chachoua A, Wernz J, Zeleniuch-Jacquotte A, Blum R, Speyer J (1996) Phase II trial of paclitaxel and cisplatin in women with advanced breast cancer: an active regimen with limiting neurotoxicity. J Clin Oncol 14:1993–1999
Wiernik PH, Schwartz EL, Einzig A, Strauman JJ, Lipton RB, Dutcher JP (1987) Phase I trial of taxol given as a 24-hour infusion every 21 days: responses observed in metastatic melanoma. J Clin Oncol 5:1232–1239
Wilson L, Jordan MA (2004) New microtubule/tubulin-targeted anticancer drugs and novel chemotherapeutic strategies. J Chemother 16(Suppl 4):83–85
Wozniak KM, Nomoto K, Lapidus RG, Wu Y, Carozzi V, Cavaletti G, Hayakawa K, Hosokawa S, Towle MJ, Littlefield BA, Slusher BS (2011) Comparison of neuropathy-inducing effects of eribulin mesylate, paclitaxel, and ixabepilone in mice. Cancer Res 71:3952–3962
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Wozniak, K.M., Wu, Y., Farah, M.H. et al. Neuropathy-Inducing Effects of Eribulin Mesylate Versus Paclitaxel in Mice with Preexisting Neuropathy. Neurotox Res 24, 338–344 (2013). https://doi.org/10.1007/s12640-013-9394-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12640-013-9394-3