
Abstract
Chemotherapy is the most common method to treat cancer. The use of certain antineoplastic drugs, however, is associated with the development of peripheral neuropathy that can be dose-limiting. Excitotoxic glutamate release, leading to excessive glutamatergic neurotransmission and activation of N-methyl-d-aspartate (NMDA) receptors, is associated with neuronal damage and death in several nervous system disorders. N-Acetyl-aspartyl-glutamate (NAAG) is an abundant neuropeptide widely distributed in the central and peripheral nervous system which is physiologically hydrolyzed by the enzyme glutamate carboxypeptidase into N-Acetyl-aspartyl (NAA) and glutamate. Pharmacological inhibition of glutamate carboxypeptidase results in decreased glutamate and increased endogenous NAAG and has been shown to provide neuroprotection in several preclinical models. Here, we report the neuroprotective effect of an orally available glutamate carboxypeptidase inhibitor on three well-established animal models of chemotherapy (cisplatin, paclitaxel, bortezomib)-induced peripheral neuropathy. In all cases, glutamate carboxypeptidase inhibition significantly improved the chemotherapy-induced nerve conduction velocity deficits. In addition, morphological and morphometrical alterations induced by cisplatin and bortezomib in dorsal root ganglia (DRG) were improved by glutamate carboxypeptidase inhibition. Our data support a novel approach for the treatment of chemotherapy-induced peripheral neuropathy.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Glutamate is one of the major neurotransmitters of the human nervous system and, acting on its neuronal and glial ionotropic and metabotropic receptors, plays a crucial role in several physiological functions such as learning, memory, and plasticity (Conn 2003). Excitotoxic glutamate release, leading to excessive glutamatergic transmission and activation of N-methyl-d-aspartate (NMDA) receptors, is associated with neuronal damage and death in several neurological disorders of both the central nervous system, such as ischemia and stroke (for reviews see Aarts et al. 2003; Kwak and Weiss 2006), and the peripheral nervous system (Vincent et al. 1997; Sagara and Schubert 1998; Cai et al. 1999; Leker and Shohami 2002; Aarts and Tymianski 2003; Blaabjerg et al. 2003; Bacich et al. 2005; Berent-Spillson and Russell 2007; for a review see Carozzi et al. 2008a). N-Acetyl-aspartyl-glutamate (NAAG) is an abundant neuropeptide widely distributed in the central and peripheral nervous systems. NAAG has been shown to be a presynaptic agonist at the metabotropic glutamate receptor 3 (mGluR3) on neurons and glia. It acts by reducing cAMP levels via an inhibitory G protein (Wroblewska et al. 1993, 1997, 1998), affecting a large spectrum of neuronal and glial functions including inhibiting transmitter release (Cartmell and Schoepp 2000) and increasing production of neuroprotective trophic factors (Jackson et al. 1996; Bruno et al. 1998; Slusher et al. 1999; Thomas et al. 2001). Westbrook et al. (1986) showed that high concentrations of NAAG activate NMDA receptors in mouse spinal cord neuron cell culture. In contrast, however, recent data demonstrated that NAAG had no activity at human recombinant mGluR2 and mGluR3 in a cellular G protein-activated K+ channel electrophysiology assay (Fricker et al. 2009) or at NMDA receptors in rat hippocampal neuronal cultures (Fricker et al. 2009; Losi et al. 2004). The reasons for these discrepancies are not currently understood. NAAG is physiologically hydrolyzed by a group of neuropeptidases, termed glutamate carboxypeptidases, also known as NAAG peptidases and NAALADase, into N-Acetyl-aspartyl (NAA) and glutamate (Riveros and Orrego 1984; Robinson et al. 1987; Carter et al. 1996; Bzdega et al. 1997; Bzdega et al. 2004). Two specific glutamate carboxypeptidases, glutamate carboxypeptidase II and III, have been described with a similar kinetic and pharmacological profiles (Bzdega et al. 2004). The enzymes have been localized in the proximal gastrointestinal system, renal tubules, as well as in rat brain astrocytes (Berger et al. 1999). In the peripheral nervous system, glutamate carboxypeptidase is present in Schwann cells surrounding the peripheral nerve fibers and NAAG is found in dorsal root ganglia (DRG) and in axons (Lieberman et al. 1994; Cangro et al. 1987; Berger et al. 1995; Berger and Schwab 1996; Carozzi et al. 2008b). Moreover, glutamate carboxypeptidase and NAAG have been shown to play a key role in the glutamatergic mechanisms of axon-glia signalling (Lieberman 1991; Lieberman et al. 1994; Urazaev et al. 2001).
Glutamate carboxypeptidase inhibition results in decreased glutamate and increased endogenous NAAG (Slusher et al. 1999; Zhong et al. 2006). The inhibition of glutamate carboxypeptidase not only reduces extracellular glutamate directly by decreasing the rate of glutamate originated by NAAG hydrolysis but also by the inhibition of the synaptic release of glutamate through NAAG activation of group II metabotropic receptors (Zhao et al. 2001; Sanabria et al. 2004).
Glutamate carboxypeptidase pharmacological inhibition has previously been demonstrated to be neuroprotective in experimental models of cerebral ischemia (Slusher et al. 1999; Bacich et al. 2005), chronic pain (Carpenter et al. 2003), amyotrophic lateral sclerosis (ALS) (Ghadge et al. 2003) as well as in diabetic neuropathy (Zhang et al. 2002, 2006). Several other drugs that attenuate glutamate excitotoxicity through postsynaptic receptor blockade have been shown to relieve neuropathic pain (Kawamata and Omote 1996), ischemia (Nishizawa 2001) and ALS (Kidd and Isaac 2000). However, debilitating side effects have limited their clinical use. In contrast, pharmacological inhibition of glutamate carboxypeptidase does not seem to affect glutamate function under normal levels of nervous system activity in animals (Slusher et al. 1999) or in initial clinical studies (van der Post et al. 2005). This may be because glutamate carboxypeptidase inhibition enhances a naturally ongoing regulatory process rather than chronically activating or inhibiting receptors in a manner that is unrelated to basal chemical neurotransmission (Zhou et al. 2005). In addition, Urazaev et al. (2001) reported that in crayfish nerve fibers, glutamate carboxypeptidase activity was limited under basal condition, and only selectively activated under stimulated conditions. This could explain why glutamate carboxypeptidase inhibition does not affect basal glutamatergic transmission, although this finding has not been replicated in mammalian system.
Chemotherapy represents one of the most effective ways to treat cancer. Nevertheless, the use of certain antineoplastic drugs, such as proteasome inhibitors (bortezomib), anti-tubulins (paclitaxel), thalidomide, platinum-derived compounds (cisplatin, oxaliplatin) and vinca alkaloids, alone or in combination, frequently results in the development of a dose-limiting peripheral neuropathy which preferentially damages neurons of DRG and/or peripheral nerve fibers (Cavaletti et al. 1991, 1992, 1995, 2007; Authier and Gillet 2003; Carozzi et al. 2009).
Here, we evaluated the neuroprotective effect of a glutamate carboxypeptidase inhibitor (E2072, EISAI Corp, Baltimore, USA) on neurophysiological impairment, morphological and morphometrical alterations of DRG and peripheral nerve fiber morphological changes using three different well-established animal models of chemotherapy-induced peripheral neuropathies (including cisplatin, paclitaxel and bortezomib).
Materials and Methods
Animals
Young adult female Wistar rats (175–200 g at the housing room arrival, Harlan Italy, Correzzana) were used for the study. The care and husbandry of animals were in conformity with the institutional guidelines in compliance with national (D.L. n. 116, Gazzetta Ufficiale della Repubblica Italiana, suppl. 40, Feb. 18, 1992) and international laws and policies (EEC Council Directive 86/609, OJ L 358, 1, Dec. 12, 1987; Guide for the Care and Use of Laboratory Animals, U.S. National Research Council, 1996). The experimental plan was preliminarily examined and approved by the ad hoc committee of the University of Milan Bicocca. Animals were housed in a limited access animal facility where animal room temperature and relative humidity were set at 22 ± 2°C and 55 ± 10%, respectively. Artificial lighting provided a 24-h cycle of 12 h light/12 h dark (light 7 a.m.–7 p.m.).
At the end of pharmacological treatments, animals were sacrificed by cervical dislocation under deep CO2 anaesthesia.
Drugs
Cisplatin (Platamine, Pfizer, Nerviano, Italy), paclitaxel, bortezomib (LC Laboratories, Woburn, MA) and E2072 (EISAI Corp, Baltimore, USA) solutions were prepared immediately before each administration.
Cisplatin was dissolved in sterile saline and intraperitoneally (IP) administered, while paclitaxel and bortezomib were dissolved in absolute ethanol/Tween 80/saline solution (5%/5%/90%) and intravenously (IV) administered.
E2072 was dissolved in Hepes buffered saline (50 mM Hepes in 0.95% w/v NaCl) and orally (per os, PO) administered daily.
Pharmacokinetic Studies
Daily oral doses of 10 mg/kg E2072 and lower were chosen for the neuropathy efficacy experiments based upon data from pharmacokinetic and tolerability studies. Sprague–Dawley rats were dosed with 10 mg/kg of E2072 in 50 mM Hepes buffered saline via oral gavage for either 1 or 5 days. At 0.25, 0.5, 1, 2, 4, 6, and 8 h following dosing, the rats (n = 4–6/group) were sacrificed and terminal plasma and sciatic nerve samples were collected and analyzed for E2072 via liquid chromatography with tandem mass spectrometry detection (LC/MS/MS).
Schedules of Drug Administration
As shown in Table 1, three experiments were set up to test the neuroprotective effects of E2072 against peripheral neurotoxicity induced by three different chemotherapies: cisplatin, paclitaxel, and bortezomib. All antineoplastic drugs were administered as a 1 ml/kg solution. In each experiment, 40 animals were randomly divided into five groups of 8 animals: one group of healthy (control) rats, one group of rats treated with the chemotherapy alone and three groups of rats that received both the chemotherapy treatment and daily doses of oral E2072 (either 10, 1, or 0.1 mg/kg). The final E2072 dosing schedule was selected based on pilot experiments performed in our laboratory (data not shown). E2072 was delivered as 5 ml/kg solution. On days with concomitant administrations, chemotherapy drugs were administered 1 h prior to E2072.
Since the solvents used in these experiments were already extensively investigated in previous studies and no general or neurological toxicity was evidenced (Cavaletti et al. 1992, 1995; Persohn et al. 2005), control rats were left untreated.
Clinical Signs, Mortality, and Body Weight
The general clinical condition of the animals was assessed daily. Body weight was recorded twice weekly for the assessment of general toxicity of pharmacological treatments and for dose adjustment.
Neurophysiological Assessment
In each experiment, before starting the pharmacological treatment and 4 days after the last administration, nerve conduction velocity (NCV) was determined in the caudal nerve of each animal as previously described in several experimental paradigms (Cavaletti et al. 1995, 2000; Pisano et al. 2003). Briefly, the antidromic NCV in the tail nerve was assessed by placing recording ring electrodes distally on the tail, while the stimulating ring electrodes were placed 5 and 10 cm proximally with respect to the recording point. The latency of the potentials recorded at the two sites after nerve stimulation was determined (peak-to-peak) and NCV was calculated accordingly. All the neurophysiological determinations were performed under standard conditions in a temperature-controlled room.
Prior to the initiation of dosing in each experiment, NCV was evaluated for each experimental animal and then rats were divided into five treatment groups so that baseline NCV measures were homogeneous and ranged between 28 and 32 m/s among the groups (data not shown).
Neuropathological Examinations
In each experiment, 5 animals/group were sacrificed at the end of the 4-week treatment period and used for biological sampling. The left sciatic nerves and the L4–L5 DRG were obtained from the sacrificed animals and processed according to previously reported protocols, resin embedded and used for light microscope observations (Cavaletti et al. 1992; Persohn et al. 2005). Briefly, 1-μm-semithin sections were prepared from at least three tissue blocks for each animal. The sections were stained with toluidine blue and examined with a Nikon Eclipse E200 light microscope.
Morphometric Examinations
DRG of control and treated rats were used for morphometric examinations. On toluidine blue stained 1-μm-thick semithin sections, DRG from cisplatin and bortezomib experiments were analyzed with a computer-assisted image analyzer (ImageJ NIH software). In the cisplatin experiment, the somatic, nuclear, and nucleolar sizes of DRG sensory neurons were measured in randomly selected sections according to previously reported methods on at least 300 DRG neurons/rat (Cavaletti et al. 1992).
On the basis of the previously observed bortezomib-induced toxicity on DRG satellite cells (Cavaletti et al. 2007), the incidence of damaged satellite cells was determined in randomly selected 1-μm-thick semithin DRG sections of bortezomib experiment. The results are expressed as a rate (%) of vacuolated cells on a total of 500 cells/rat.
Statistical Evaluations
The differences between all experimental groups within each experiment, in body weight, NCV, and morphometric data, were statistically evaluated using the analysis of variance (one-way ANOVA) and the Tukey–Kramer post- hoc test (significance level set at P < 0.05).
Results
E2072 Pharmacokinetic and Tolerability Studies
The levels of E2072 in plasma and sciatic nerve were comparable following single and multiple doses, indicating no compound accumulation. Plasma E2072 reached maximal levels of 5000 ng/ml at 1 h after administration. The plasma half-life (t1/2) was approximately 4 h and the total area under the curve (AUC) was 19000 ng*h/ml. In the rat sciatic nerve, E2072 reached levels of 200 ng/g (approximately 700 nM) at 1 h and remained above 70 ng/g (approximately 250 nM) at the last measured 8 h post-administration time point. These data show that E2072 peripheral nerve levels exceed those necessary to inhibit the glutamate carboxypeptidase enzymatic activity (Ki = 10 nM) for at least 8 h following a 10 mg/kg oral dose. In addition, tolerability studies were conducted in rats using 28 daily oral doses of 10 mg/kg E2072. Body weights were unchanged relative to vehicle-treated animals and no obvious clinical signs were observed. Furthermore, neurophysiological analysis and neuropathological examinations of DRG and peripheral nerve fibers did not show any neurotoxic effect of 28 daily oral doses of 10 mg/kg E2072 on the rat peripheral nervous system (data not shown). Based on these data, doses of 10 mg/kg E2072 and lower were chosen for the neuropathy studies.
Prevention of Cisplatin-Induced Peripheral Neuropathy
General Toxicity for Cisplatin-Treated Rats
Although cisplatin induced mild piloerection, hypokinesia, and kyphosis in rats, the selected schedules of treatment were generally well tolerated and no mortality was observed during the study. At the end of the experiment, as shown in Table 2, the cisplatin regimen induced a significant reduction in body weight compared to controls (−26.4%). The co-administration of 10, 1, and 0.1 mg/kg E2072 caused a less marked reduction in body weight compared to controls (−14.1, −9.6, and −13.6%, respectively). However, due to the variability and small sample size per treatment group, the only E2072 treatment group in which cisplatin-induced weight loss was significantly prevented was the group receiving co-administration of 1 mg/kg E2072.
Neurophysiological Assessment for Cisplatin-Treated Rats
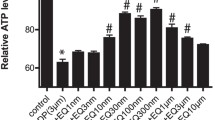
As shown in Fig. 1, cisplatin treatment alone caused a significant reduction of nerve conduction velocity (NCV) compared to controls (−43.9%). E2072 (at 1 and 10 mg/kg), when co-administered with cisplatin, significantly prevented the NCV reduction. Co-administration of 0.1 mg/kg E2072 conveyed no significant protection.

Nerve conduction velocity mean values (+SD) at the end of the cisplatin experiment. Statistical analysis: one-way ANOVA, Tukey–Kramer post-test. CTRL controls, C cisplatin, NCV nerve conduction velocity
Neuropathological Examinations for Cisplatin-Treated Rats
Sciatic Nerves
Using light microscope examination, performed at the end of the experiments, the sciatic nerves of animals treated with cisplatin alone showed only mild morphological axonal changes, represented by fibers undergoing wallerian-like degeneration in the absence of primary demyelinating aspects. These results were consistent with our previous data (Cavaletti et al. 1991). Given the limited severity of cisplatin-induced alterations, a protective effect of E2072 could not be observed (data not shown).
Dorsal Root Ganglia
As previously described (Cavaletti et al. 1991), DRG from cisplatin-treated rats showed morphologic alterations (Fig. 2). Although multinucleolated neurons and nucleolar eccentricity were sporadically seen in control rats, in cisplatin-treated rats their incidence was increased. DRG of rats co-treated with cisplatin and E2072 (10, 1, and 0.1 mg/kg) showed milder alterations compared to cisplatin-treatment alone.

Light microscopy analysis of DRG morphology in cisplatin experiment. Multinucleolated sensory neurons and nucleolar eccentricity in cisplatin-treated animals are shown by the arrows and arrowheads, respectively. Similar but milder alterations (arrows and arrowheads) were observed in DRG of animals co-treated with cisplatin and E2072; bar: 50 μm. CTRL control, C cisplatin
Morphometrical Analysis of DRG Sensory Neurons
Based on the documented changes induced by cisplatin administration in DRG neurons (Cavaletti et al. 1992), we performed morphometrical analysis of sensory neurons from cisplatin-treated rats and cisplatin/E2072 treated rats.
As shown in Table 3, cisplatin treatment alone induced a statistically significant decrease in somatic and nucleolar, but not nuclear, size compared to controls. By contrast, the co-administration of 10 mg/kg E2072 with cisplatin prevented this reduction both at the somatic and nucleolar levels. Co-administration of both 0.1 and 1 mg/kg E2072 with cisplatin had no significant protective effect.
Prevention of Paclitaxel-Induced Peripheral Neuropathy
General Toxicity for Paclitaxel-Treated Rats
The selected schedule for paclitaxel was generally well tolerated with no mortality observed in any treatment group during the study. At the end of the experiment, as shown in Table 2, paclitaxel in presence and absence of E2072 had no significant alteration on body weight compared to controls (−4.8% for group P, −3% for group P + E2072 10, −1.3% for group P + E2072 1, and −1.9% for group P + E2072 0.1).
Neurophysiological Assessment for Paclitaxel-Treated Rats
As shown in Fig. 3, paclitaxel treatment caused significant reductions in NCV compared to controls (−33.1%). When 10 mg/kg E2072 was co-administered with paclitaxel, it significantly prevented the NCV impairment (−10%).

Nerve conduction velocity mean values (+SD) at the end of the paclitaxel experiment. Statistical analysis: one-way ANOVA, Tukey–Kramer post-test. CTRL controls, P paclitaxel, NCV nerve conduction velocity
Neuropathological Examinations
Using light microscope examination performed at the end of the experiments, sciatic nerves of animals treated with paclitaxel alone showed mild to moderate morphological axonal changes (data not shown). DRG morphology was normal which is consistent with previous observations from our laboratory (Cavaletti et al. 2000). Given the lack of significant pathological changes with paclitaxel itself, a protective effect of E2072 could not be observed (data not shown).
Prevention of Bortezomib-Induced Peripheral Neuropathy
General Toxicity of Bortezomib-Treated Rats
Although one bortezomib-treated animal died during the experiment, the pharmacological treatments were generally well tolerated by the rats. At the end of the experiment, as shown in Table 2, all schedules of treatment induced a significant reduction in body weight compared to controls (−5.7% for group B, −6.3% for group B + E2072 10, −7.7% for group B + E2072 1, and −5.4% for group B + E2072 0.1). Treatment with E2072 did not prevent the bortezomib-induced body weight loss.
Neurophysiological Evaluation of Bortezomib-Treated Rats
As shown in Fig. 4, bortezomib treatment caused a significant reduction in NCV compared to controls (−39.3%). When 10 mg/kg E2072 was co-administered with bortezomib, it significantly prevented the NCV reduction caused by bortezomib (−15%), while 1 and 0.1 mg/kg E2072 co-administration conveyed no significant protection (−33.3 and −33.4%, respectively).

Nerve conduction velocity mean values (+SD) at the end of the bortezomib experiment. Statistical analysis: one-way ANOVA, Tukey–Kramer post-test). CTRL controls, B bortezomib, NCV nerve conduction velocity
Neuropathological Examinations of Bortezomib-Treated Rats
Sciatic Nerves
At the light microscope examination level, performed at the end of the experiment, sciatic nerves of animals treated with bortezomib showed only mild to moderate morphological axonal changes (data not shown). Consequently, these results did not permit any clear-cut effect of E2072 co-administration on a morphological basis.
Dorsal Root Ganglia
As previously described (Cavaletti et al. 2007), DRG from bortezomib-treated rats showed a diffuse intracytoplasmatic vacuolization in satellite cells (Fig. 5). Only rarely, DRG sensitive neurons had a pathological appearance represented by cytoplasm having clear vacuoles. Similar, but milder, alterations were observed in DRG of rats co-treated with bortezomib and E2072.

Light microscopy analysis of DRG morphology in the bortezomib experiment. Diffuse intracytoplasmatic vacuolization in satellite cells in bortezomib-treated rats are shown by the arrows in B while only rare pathological neurons, shown in B(a), are characterized by small clear cytoplasmic vacuoles. Similar but milder alterations (arrows) were observed in DRG of animals co-treated with bortezomib and E2072; bar: 50 μm. CTRL control, B bortezomib
Morphometrical Analysis of DRG Satellite Cells
Based on our observation in satellite cells, we performed the morphometrical analysis of the incidence of altered satellite cells in bortezomib-treated versus bortezomib/E2072-treated rats. As shown in Table 4, the incidence of vacuolated satellite cells was significantly increased in all treated groups compared to controls. The co-administration of 10 mg/kg E2072 with bortezomib was able to partially, but significantly, prevent this increase. Co-administration of both 0.1 and 1 mg/kg E2072 with bortezomib showed only a mild protective effect which did not reach a statistical significance.
Discussion
Glutamate carboxypeptidase pharmacological inhibitors have been demonstrated to exert neuroprotective effects both in in vitro (Thomas et al. 2000) and in vivo models of disorders of the central nervous system (for reviews see Neale et al. 2005; Thomas et al. 2006). Zhang et al. (2002, 2006) showed that glutamate carboxypeptidase inhibition prevents nerve conduction velocity (NCV) alterations, hyperalgesia, and the related pathology of C and myelinated fibers in short-term and chronic diabetic painful peripheral neuropathies. Glutamate carboxypeptidase inhibitors have also been shown to reduce neuropathic pain and ectopic discharges from injured nerve and minimize central sensitization in animal models of inflammatory and injury-induced neuropathies (Yamamoto et al. 2001; Carpenter et al. 2003). When locally injected, glutamate carboxypeptidase inhibitors have also been shown to induce a reduction in the perception of inflammatory pain, demonstrating a potential effect at both peripheral and central sites (Yamamoto et al. 2007).
The effects of glutamate carboxypeptidase inhibition are thought to be directly exerted by a reduction in glutamate release and by an increase in NAAG which may act as an agonist at mGluR3 (Wroblewska et al. 1997; Bruno et al. 1998) and, although there are some controversies in the literature, this may interfere with downstream NMDA receptor sensitization and transmission (Westbrook et al. 1986; Losi et al. 2004; Fricker et al. 2009).
The use of several antineoplastic drugs such as platinum-derived compounds, antitubulins, and inhibitors of proteasome is frequently associated with the development of a peripheral neuropathy which preferentially damages DRG and/or peripheral nerve fibers (Cavaletti et al. 1991, 1995, 2007; Authier and Gillet 2003) and often represents a dose-limiting side effect (Windebank 1999; Quasthoff and Hartung 2002; for a review see Ocean and Vahdat 2004). Since glutamate carboxypeptidase inhibitors do not alter chemotherapy efficacy (Tang et al. 2005), here we examined the neuroprotective effects of a glutamate carboxypeptidase inhibitor (E2072) on experimental peripheral neuropathy triggered by three different chemotherapy treatments. Chemotherapy-induced peripheral neuropathy is characterized by neurophysiological changes, morphological alterations in DRG, and myelinated nerve fibers (Cavaletti et al. 1991, 1995, 2007) and, sometimes, as in the case of platinum-derived compounds, also by morphometrical changes in cellular sizes of DRG sensory neurons (Cavaletti et al. 1992). Based on the known mechanisms of antineoplastic action, the pathological examination of peripheral nervous system specimens enables one to obtain important information on the mechanisms and severity of the neurotoxic action of the compounds and any prevention of these changes, as seen with E2072 in this study. In the case of cisplatin, it is now widely accepted that all these alterations are secondary to the formation of platinum-DNA adducts in the DRG neurons (Meijer et al. 1999), leading to nucleolar components segregation and nucleolar disruption with cell body shrinkage, while nerve fiber changes are secondary to DRG injury. Moreover, these pathological changes are not particular to cisplatin and they have been reported also in animal models of carboplatin or oxaliplatin peripheral neurotoxicity, thus indicating a common mechanism of action for all platinum-derived drugs (Cavaletti et al. 2008). In the case of bortezomib, the intracytoplasmatic vacuolation observed in DRG satellite cells (and more rarely in neurons) is due to mitochondrial and endoplasmic reticulum enlargement (Cavaletti et al. 2007) and these changes can be related to bortezomib’s ability to induce the activation of the mitochondrial-based (“intrinsic”) apoptotic pathway and the expression of proteins associated with the endoplasmic reticulum secretory pathway, thus activating caspase 12, calcium homeostasis dysregulation and, finally, cell death (Landowski et al. 2005). Regarding taxanes, such as paclitaxel, obvious DRG changes have never been reported at the morphological level, while axonal microtubule changes have been described only after extensive morphometric analysis at the ultrastructural level (Persohn et al. 2005), in agreement with their well-established tubulin hyperpolymerating activity.
In our study, the neuroprotective effect of the glutamate carboxypeptidase inhibition on NCV deficits and on cisplatin-induced alterations in DRG neurons morphometry was strongly exerted by the highest dose tested (10 mg/kg) and, to a lesser extent, by the intermediate dose (1 mg/kg). In these models, the lowest dose of E2072 (0.1 mg/kg) did not show any significant neuroprotective action. On the basis of NCV measurements, bortezomib- and paclitaxel-induced peripheral neuropathy was prevented only by the highest dose of E2072, while in cisplatin neuropathy the intermediate dose (1 mg/kg) was also effective. At the morphometric examination level, only the higher dose of E2072 protected DRG neurons against the cisplatin-induced atrophy. This may be due to the fact that the DRG neuronal insults are one of the first events that occur in the early phases of the pathogenesis of cisplatin-induced peripheral neuropathy (Cavaletti et al. 1992) and only later the degeneration of nerve fibers with the subsequent NCV impairment becomes evident. At the morphological level, all tested doses of E2072 were able to partially prevent the cisplatin-and bortezomib-induced alterations in DRG. However, since all chemotherapy regimens caused only mild morphological axonal changes in the peripheral nerves, no clear-cut protective effect of E2072 co-administration on sciatic nerve morphology was evident. Importantly, however, E2072 did not negatively influence the pathology at any dose tested.
In co-administration regimens, E2072 was able to prevent the cisplatin-induced weight loss but, due to variability and the relatively small sample size, this effect reached statistical significance only at one dose group. The mechanism by which glutamate carboxypeptidase inhibition would prevent weight loss is unknown, unless it is secondary to the peripheral nerve protection providing the animals better mobility and improved sense of well being. In contrast, however, no weight loss prevention was observed in the bortezomib rats. Importantly, the E2072-induced weight loss protection cannot explain the E2072-induced improvement in neurotoxicity. This is based on literature data showing minimal effects of malnutrition on peripheral nerve function (Cornblath and Brown 1988), on our previous peripheral nerve observations with animals of different weights treated with cisplatin (Cavaletti et al. 1991, 1992; Carozzi et al. 2009) and on the fact that no weight change occurred in this study in paclitaxel-treated rats, despite significant NCV changes. Finally, the extent of weight change was clearly more evident in the cisplatin-treated rats than in the bortezomib-treated group, although the extent of NCV reduction was similar.
Doses of E2072 (0.1–10 mg/kg) were chosen for the neurotoxicity studies based upon data from drug tolerance and pharmacokinetics studies. Dosing with 10 mg/kg E2072 in the pharmacokinetic study provided concentrations of inhibitor in sciatic nerves that exceeded the Ki for inhibition of glutamate carboxypeptidase. In the tolerance study, we demonstrated that the administration of E2072 10 mg/kg alone was neither systemically toxic nor neurotoxic on the basis of body weight monitoring, neurophysiological analysis, and DRG morphological and morphometrical observations.
Glutamate carboxypeptidase inhibition can trigger both a reduction in glutamate and an increase in NAAG (Slusher et al. 1999; Zhong et al. 2006). In the peripheral nervous system, the neuroprotective effects of glutamate carboxypeptidase inhibition are most likely mediated via inhibition of glutamate release from Schwann cells, by decreased excitability of damaged peripheral afferents (Berger and Schwab 1996), by inhibiting glutamate release and promoting endogenous NAAG in the spinal dorsal horn (Kawamata and Omote 1996; Carpenter et al. 2003). Interestingly, the co-administration of E2072 provided similar results in models where three different pathogenic mechanisms have been postulated. According to the commonly accepted hypothesis, cisplatin damages the primary sensory neurons, paclitaxel targets axonal tubulin, while bortezomib (at least in animal models) is toxic on DRG satellite cells and nerve Schwann cells, rather than axons. However, it is possible that other less obvious targets exist for these antineoplastic compounds that have not yet been identified. Until a common mechanism of action is recognized, our experimental results can be explained by the fact that E2072 may have a broad mechanism of action affecting any situation where there is neuronal-glial stress and excessive glutamatergic transmission. Further support to this hypothesis is provided by the fact that E2072 is active in experimental diabetic neuropathy models. Zhang et al. (2006) suggested that the neuroprotective effect of glutamate carboxypeptidase inhibition against neurophysiological deficits in an experimental chronic diabetic neuropathy was due to the prevention of the impairment of neural Na+/K+-ATPase activity. In diabetic peripheral neuropathy, endoneurial hypoxemia is an early and major pathogenetic component of the pathology that is associated with injuries to the Na+/K+-ATPase activity contributing to the nerve conduction defect (Cameron and Cotter 1994; Cotter et al. 1995; Sima 2003). Since Na+/K+-ATPase had also been shown to be involved in the transport of some chemotherapy drugs into cells and to be inhibited by chemotherapy treatments (Sakakibara et al. 1999), it is conceivable that NCV deficits in chemotherapy-induced peripheral neurotoxicity and its protection by glutamate carboxypeptidase inhibition could be explained, at least in part, by effects on the Na+/K+-ATPase system. However, this mechanism alone cannot explain the protective activity of E2072 on DRG neuronal cells, demonstrated at the morphometric level in the cisplatin model.
Earlier studies (Jackson et al. 1996; Bruno et al. 1998; Slusher et al. 1999; Thomas et al. 2001) demonstrated that increased levels of NAAG, related to the inhibition of glutamate carboxypeptidase and the subsequent activation of mGluR2/3, may increase the production of neuroprotective growth factors, including trophic factors such as the transforming growth factor β (TGF β), from glial cells. It is possible that the increase of endogenous TGF β (or other neurotrophic factors), triggered by glutamate carboxypeptidase inhibition, could mediate neurotrophic activity also in damaged peripheral nerve fibers, re-establishing their ability in conducting electrical stimuli and improving neurophysiological parameters. This might be particularly important in chemotherapy-induced peripheral neurotoxicity where decreased levels of nerve growth factor (NGF) have been reported in clinical practice, as well as in experimental models (Tredici et al. 1999).
In conclusion, this study presents evidence for a neuroprotective role of glutamate carboxypeptidase inhibition in several chronic models of chemotherapy-induced peripheral neuropathy and claims an approach for the clinical treatment of this pathology. These results also support manipulation of the glutamatergic system in the management of toxic peripheral neuropathy.
References
Aarts MM, Tymianski M (2003) Novel treatment of excitotoxicity: targeted disruption of intracellular signalling from glutamate receptors. Biochem Pharmacol 66:877–886
Aarts MM, Arundine M, Tymianski M (2003) Novel concepts in excitotoxic neurodegeneration after stroke. Expert Rev Mol Med 5(30):1–22
Authier N, Gillet JP (2003) An animal model of nociceptive peripheral neuropathy following repeated cisplatin injections. Exp Neurol 182:12–20
Bacich DJ, Wozniak KM, May Lu XC, O’Keefe DS, Callizot N, Heston WD, Slusher BS (2005) Mice lacking glutamate carboxypeptidase II are protected from peripheral neuropathy and ischemic brain injury. J Neurochem 95:314–323
Berent-Spillson A, Russell JW (2007) Metabotropic glutamate receptor 3 protects neurons from glucose-induced oxidative injury by increasing intracellular glutamate concentration. J Neurochem 101:342–354
Berger UV, Schwab ME (1996) N-Acetylated alpha-linked acidic dipeptidase may be involved in axon Schwann cell signaling. J Neurocytol 25:499–512
Berger UV, Carter RE, McKee M, Coyle JT (1995) N-Acetylated alpha-linked acidic dipeptidase is expressed by non-myelinating Schwann cells in the peripheral nervous system. J Neurocytol 2(2):99–109
Berger UV, Luthi-Carter R, Passani LA, Elkabes S, Black I, Konradi C, Coyle JT (1999) Glutamate carboxypeptidase II is expressed by astrocytes in the adult rat nervous system. J Comp Neurol 415(1):52–64
Blaabjerg M, Fang L, Zimmer J, Baskys A (2003) Neuroprotection against NMDA excitotoxicity by group I metabotropic glutamate receptors is associated with reduction of NMDA stimulated currents. Exp Neurol 183:573–580
Bruno V, Wroblewska B, Wroblewska JT, Fiori L, Nicoletti F (1998) Neuroprotective activity of N-acetyl-aspartyl-glutamate in cultured cortical cells. Neuroscience 85:751–757
Bzdega T, Turi T, Wroblewska B, She D, Chung HS, Kim H, Neale JH (1997) Molecular cloning of a peptidase against N-acetyl-aspartyl-glutamate from a rat hippocampal cDNA library. J Neurochem 69(6):2270–2277
Bzdega T, Crowe SL, Ramadan ER, Sciarretta KH, Olszewski RT, Ojeifo OA, Rafalski VA, Wroblewska B, Neale JH (2004) The cloning and characterization of a second brain enzyme with NAAG peptidase activity. J Neurochem 89(3):627–635
Cai Z, Xiao F, Fratkin JD, Rhodes PG (1999) Protection of neonatal rat brain from hypoxic-ischemic injury by LY379268, a Group II metabotropic glutamate receptor agonist. Neuroreport 10:3927–3931
Cameron NE, Cotter MA (1994) The relationship of vascular changes to metabolic factors in diabetes mellitus and their role in the development of peripheral nerve complications. Diab Metab Rev 10:189–224
Cangro CB, Namboodiri MA, Sklar LA, Corigliano-Murphy A, Neale JH (1987) Immunohistochemistry and biosynthesis of N-acetyl-aspartyl-glutamate in spinal sensory ganglia. J Neurochem 49(5):1579–1588
Carozzi VA, Marmiroli P, Cavaletti G (2008a) Focus on the role of glutamate in the pathology of peripheral nervous system. CNS Neurol Disord Drug Targets 7(4):348–360
Carozzi VA, Canta A, Oggioni N, Ceresa C, Marmiroli P, Konvalinka J, Zoia C, Bossi M, Ferrarese C, Tredici G, Cavaletti G (2008b) Expression and distribution of “High affinity” glutamate transporters and GCPII in the peripheral nervous system. J Anat 213(5):539–546
Carozzi V, Chiorazzi A, Canta A, Oggioni N, Gilardini A, Rodriguez-Menendez V, Avezza F, Crippa L, Ceresa C, Nicolini G, Bossi M, Cavaletti G (2009) Effect of the chronic combined administration of cisplatin and paclitaxel in a rat model of peripheral neurotoxicity. Eur J Cancer 45(4):656–665
Carpenter KJ, Sen S, Matthews WA, Flatters SL, Wozniak KM, Slusher BS, Dickenson AH (2003) Effects of GCP II inhibition on responses of dorsal horn neurones after inflammation and neuropathy: an electrophysiological study in the rat. Neuropeptides 37:2998–3006
Carter RE, Feldman AR, Coyle JT (1996) Prostate-specific membrane antigen is a hydrolase with substrate and pharmacologic characteristics of a neuropeptidase. Proc Natl Acad Sci USA 93(2):749–753
Cartmell J, Schoepp DD (2000) Regulation of neurotransmitter release by metabotropic glutamate receptors. J Neurochem 75(3):889–907
Cavaletti G, Petruccioli MG, Tredici G, Marmiroli P, Barajon I, Fabbrica D, Di Francesco A (1991) Effects of repeated administration of low doses of cisplatin on the rat nervous system. Int J Tissue React 13(3):151–157
Cavaletti G, Tredici G, Marmiroli P, Petruccioli MG, Barajon I, Fabbrica D (1992) Morphometric study of the sensory neuron and peripheral nerve changes induced by chronic cisplatin (DDP) administration in rats. Acta Neuropathol 84(4):364–371
Cavaletti G, Tredici G, Braga M, Tazzari S (1995) Experimental peripheral neuropathy induced in adult rats by repeated intraperitoneal administration of taxol. Exp Neurol 133(1):64–72
Cavaletti G, Cavalletti E, Oggioni N, Sottani C, Minoia C, D’Incalci M, Zucchetti M, Marmiroli P, Tredici G (2000) Distribution of paclitaxel within the nervous system of the rat after repeated intravenous administration. Neurotoxicology 21:389–393
Cavaletti G, Gilardini A, Canta A, Rigamonti L, Rodriguez-Menendez V, Ceresa C, Marmiroli P, Bossi M, Oggioni N, D’Incalci M, De Coster R (2007) Bortezomib-induced peripheral neurotoxicity: a neurophysiological and pathological study in the rat. Exp Neurol 204(1):317–325
Cavaletti G, Nicolini G, Marmiroli P (2008) Neurotoxic effects of antineoplastic drugs: the lesson of pre-clinical studies. Front Biosci 13:3506–3524
Conn PJ (2003) Physiological roles and therapeutic potential of metabotropic glutamate receptors. Ann NY Acad Sci 1003:12–21
Cornblath DR, Brown MJ (1988) Influence of malnutrition on developing rat peripheral nerves. Exp Neurol 99(2):403–411
Cotter MA, Love A, Watt MJ, Cameron NE, Dines KC (1995) Effects of natural free radical scavengers on peripheral nerve and neurovascular function in diabetic rats. Diabetologia 38:1285–1294
Fricker AC, Mok MH, de la Flor R, Shah AJ, Woolley M, Dawson LA, Kew JN (2009) Effects of N-acetyl-aspartyl-glutamate (NAAG) at group II mGluRs and NMDAR. Neuropharmacology 56(6–7):1060–1067
Ghadge GD, Slusher BS, Bodner A, Canto MD, Wozniak K, Thomas AG, Rojas C, Tsukamoto T, Majer P, Miller RJ, Monti AL, Roos RP (2003) Glutamate carboxypeptidase II inhibition protects motor neurons from death in familiar amyotrophic lateral sclerosis models. Proc Natl Acad Sci USA 100(16):9554–9559
Jackson PF, Cole DC, Slusher BS, Stetz SL, Ross LE, Donzanti BA, Trainor DA (1996) Design, synthesis and biological activity of a potent inhibitor of the neuropeptidase N-acetylated-α-linked acidic dipeptidase. J Med Chem 39:619–622
Kawamata M, Omote K (1996) Involvement of increased excitatory amino acids and intracellular Ca2+ concentration in the spinal dorsal horn in an animal model of neuropathic pain. Pain 68:85–96
Kidd FL, Isaac JT (2000) Glutamate transport blockade has a differential effect on AMPA and NMDA receptor-mediated synaptic transmission in developing barrel cortex. Neuropharmacology 39(5):725–732
Kwak S, Weiss JH (2006) Calcium-permeable AMPA channels in neurodegenerative disease and ischemia. Curr Opin Neurobiol 16(3):281–287
Landowski TH, Megli CJ, Nullmeyer KD, Lynch RM, Dorr RT (2005) Mitochondrial-mediated dysregulation of Ca2+ is a critical determinant of Velcade (PS-341/bortezomib) cytotoxicity in myeloma cell lines. Cancer Res 65:3828–3838
Leker RR, Shohami E (2002) Cerebral ischemia and trauma-different aetiologies yet similar mechanisms: neuroprotective opportunities. Brain Res Rev 39:55–73
Lieberman EM (1991) Role of glutamate in axon-Schwann cell signaling in the squid. Ann NY Acad Sci 633:448–457
Lieberman EM, Hargittai PT, Grossfeld RM (1994) Electrophysiological and metabolic interactions between axons and glia in crayfish and squid. Prog Neurobiol 44(4):333–376
Losi G, Vicini S, Neale J (2004) NAAG fails to antagonize synaptic and extrasynaptic NMDA receptors in cerebellar granule neurons. J Neurosci 6:3385–3392
Meijer C, de Vries EG, Marmiroli P, Tredici G, Frattola L, Cavaletti G (1999) Cisplatin-induced DNA-platination in experimental dorsal root ganglia neuronopathy. Neurotoxicology 20(6):883–887
Neale JH, Olszewski RT, Gehl LM, Wroblewska B, Bzdega T (2005) The neurotransmitter N-acetylaspartylglutamate in models of pain, ALS, diabetic neuropathy, CNS injury and schizophrenia. Trends Pharmacol Sci 26(9):477–484
Nishizawa Y (2001) Glutamate release and neuronal damage in ischemia. Life Sci 69(4):369–381
Ocean AJ, Vahdat LT (2004) Chemotherapy-induced peripheral neuropathy: pathogenesis and emerging therapies. Support Care Cancer 12(9):619–625
Persohn E, Canta A, Schoepfer S, Traebert M, Mueller L, Gilardini A, Galbiati S, Nicolini G, Scuteri A, Lanzani F, Giussani G, Cavaletti G (2005) Morphological and morphometric analysis of paclitaxel and docetaxel-induced peripheral neuropathy in rats. Eur J Cancer 41(10):1460–1466
Pisano G, Pratesi D, Laccabue F, Zunino P, Lo Giudice A, Bellucci L, Pacifici B, Camerini L, Vesci M, Castorina A, Cocuzza M, Tredici G, Marmiroli P, Nicolini G, Galbiati S, Calvani M, Carminati P, Cavaletti G (2003) Paclitaxel and cisplatin-induced neurotoxicity: a protective role of acetyl-l-carnitine. Clin Cancer Res 9:5756–5767
Quasthoff S, Hartung HP (2002) Chemotherapy-induced peripheral neuropathy. J Neurol 249(1):9–17
Riveros N, Orrego F (1984) A study of possible excitatory effects of N-acetyl-aspartyl-glutamate in different in vivo and in vitro brain preparations. Brain Res 299(2):393–395
Robinson MB, Blakely RD, Couto R, Coyle JT (1987) Hydrolysis of the brain dipeptide N-acetyl-l-aspartyl-l-glutamate. Identification and characterization of a novel N-acetylated alpha-linked acidic dipeptidase activity from rat brain. J Biol Chem 262(30):14498–14506
Sagara Y, Schubert D (1998) The activation of metabotropic glutamate receptors protects nerve cells from oxidative stress. J Neurosci 18:6662–6671
Sakakibara N, Suzuki K, Kaneta H, Yoshimura Y, Deyama Y, Matsumoto A, Fukuda H (1999) Inhibition of Na+, K+-ATPase by cisplatin and its recovery by 2-mercaptoethanol in human squamous cell carcinoma cells. Anticancer Drugs 10(2):203–211
Sanabria ER, Wozniak KM, Slusher BS, Keller A (2004) GCP II (NAALADase) inhibition suppresses mossy fiber-CA3 synaptic neurotransmission by a presynaptic mechanism. J Neurophysiol 91:182–193
Sima AAF (2003) New insights into the metabolic and molecular basis for diabetic neuropathy. Cell Mol Life Sci 60:2445–2464
Slusher BS, Vornov JJ, Thomas AG, Hurn PD, Harukuni I, Bhardwaj A (1999) Selective inhibition of NAALADase, which converts NAAG to glutamate, reduces ischemic brain injury. Nat Med 5:1396–1402
Tang Z, Lapidus RG, Wozniak KM, Scher HI, Slusher BS (2005) 2-MPPA, a selective inhibitor of Prostate Specific Membrane Antigen (PSMA), attenuates taxol-induced neuropathy in mice. 2005 American Association for Cancer Research (AACR) meeting, Anaheim, CA, USA
Thomas AG, Vornov JJ, Olkowski JL, Merion AT, Slusher BS (2000) N-Acetylated alpha-linked acidic dipeptidase converts N-acetyl-aspartyl-glutamate from a neuroprotectant to a neurotoxin. J Pharmacol Exp Ther 295:16–22
Thomas AG, Liu W, Olkowski JL, Tang Z, Lin Q, Lu XC, Slusher BS (2001) Neuroprotection mediated by glutamate carboxypeptidase II (NAALADase) inhibition requires TGF-β. Eur J Pharmacol 430:33–40
Thomas AG, Wozniak KM, Tsukamoto T, Calvin D, Wu Y, Rojas C, Vornov J, Slusher BS (2006) Glutamate carboxypeptidase II (NAALADase) inhibition as a novel therapeutic strategy. Adv Exp Med Biol 576:327–337
Tredici G, Braga M, Nicolini G, Miloso M, Marmiroli P, Schenone A, Nobbio L, Frattola L, Cavaletti G (1999) Effect of recombinant human nerve growth factor on cisplatin neurotoxicity in rats. Exp Neurol 159:551–558
Urazaev AK, Grossfeld RM, Fletcher PS, Speno H, Gafurov BS, Buttram JG (2001) Synthesis and release of N-acetyl-aspartyl-glutamate (NAAG) by crayfish nerve fibers: implications for axon-glia signaling. Neuroscience 107:697–703
van der Post SJ, de Visser ML, de Kam M, Woelfler DC, Hilt C, Vornov J, Burak ES, Bortey E, Slusher BS, Limsakun T, Cohen AF, van Gerven JM (2005) The central nervous system effects, pharmacokinetics and safety of the NAALADase-inhibitor GPI 5693. Br J Clin Pharmacol 60(2):128–136
Vincent AM, Mohammad Y, Ahmad I, Greenberg R, Maiese K (1997) Metabotropic glutamate receptors prevent nitric oxide-induced programmed cell death. J Neurosci Res 50:549–564
Westbrook GL, Mayer ML, Manboodiri MA, Neale JH (1986) High concentrations of N-acetyl-aspartyl-glutamate (NAAG) selectively activates NMDA receptors on mouse spinal cord neurons in cell culture. J Neurosci 6:3385–3392
Windebank AJ (1999) Chemotherapeutic neuropathy. Curr Opin Neurol 12(5):565–571
Wroblewska B, Wroblewska JT, Saab OH, Neale JH (1993) N-Acetyl-aspartyl-glutamate inhibits forskolin-stimulated cyclic AMP levels via a metabotropic glutamate receptor in cultured cerebellar granule cells. J Neurochem 61(3):943–948
Wroblewska B, Wroblewska JT, Pshenichkia S, Surin A, Sullivan SH, Neale JH (1997) N-Acetyl-aspartyl-glutamate selectively activates mGluR3 receptors in transfected cells. J Neurochem 69:174–181
Wroblewska B, Santi MR, Neale JH (1998) N-Acetyl-aspartyl-glutamate activates cyclic AMP-coupled metabotropic glutamate receptors in cerebellar astrocytes. Glia 24(2):172–179
Yamamoto T, Nozalis-Tagushi N, Sakashita Y (2001) Inhibition of spinal N-acetyl-alpha-linked acidic dipeptidase produces an antinociceptive effect in the rat formalin test. Neuroscience 102:473–479
Yamamoto T, Saito O, Aoe T, Bartolozzi A, Sarva J, Zhou J, Kozikowski A, Wroblewska B, Bzdega T, Neale JH (2007) Local administration of N-acetyl-aspartyl-glutamate (NAAG) peptidase inhibitors is analgesic in peripheral pain in rats. Eur J Neurosci 25(1):147–158
Zhang W, Slusher B, Murakawa Y, Wozniak KM, Tsukamoto T, Jackson PF, Sima AA (2002) GCPII (NAALADase) inhibition prevents long-term diabetic neuropathy in type 1 diabetic BB/Wor rats. J Neurol Sci 194:21–28
Zhang W, Murakawa Y, Wozniak KM, Slusher BS, Sima AA (2006) The preventive and therapeutic effects of GCPII (NAALADase) inhibition on painful and sensory diabetic neuropathy. J Neurol Sci 247:217–223
Zhao J, Ramadan E, Cappiello M, Wroblewska B, Bzdega T, Neale JH (2001) NAAG inhibits [3H]-GABA release from cortical neurons via the type 3 metabotropic glutamate receptor. Eur J Neurosci 13:340–346
Zhong C, Zhao X, Van KC, Bzdega T, Smyth A, Zhou J, Kozikowski AP, Jiang J, O’Connor WT, Berman RF, Neale JH, Lyeth BG (2006) NAAG peptidase inhibitor increases dialysate NAAG and reduces glutamate, aspartate and GABA levels in the dorsal hippocampus following fluid percussion injury in the rat. J Neurochem 97(4):1015–1025
Zhou J, Neale JH, Pomper MG, Kozikowski AP (2005) NAAG peptidase inhibitors and their potential for diagnosis and therapy. Nat Rev Drug Discov 4(12):1015–1026
Acknowledgment
This work was supported partially by an unrestricted research grant by “Fondazione Banca del Monte di Lombardia” (Guido Cavaletti).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Carozzi, V.A., Chiorazzi, A., Canta, A. et al. Glutamate Carboxypeptidase Inhibition Reduces the Severity of Chemotherapy-Induced Peripheral Neurotoxicity in Rat. Neurotox Res 17, 380–391 (2010). https://doi.org/10.1007/s12640-009-9114-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12640-009-9114-1