
Abstract
Cymothoidae, Leach, 1818 (Crustacea: Isopoda) are ectoparasites of marine, brackish and freshwater fishes that are reported to induce deleterious tissue impacts on the infested fish hosts. The present study aimed for the first-time screening of Mothocya melanosticta collected from the red sea fish Nemipterus randalli in Egypt. Surveillance study was conducted for isopod infestation among the Red Sea fish N. randalli revealing a total infestation rate of 40.96% with the species identified as M. melanosticta. The parasite species was isolated from the branchial cavity and gills. Morphometric description was estimated using dissecting stereo-microscope, and scanning electron microscope (SEM) provided new additional features for the isolated species including the microtrich sensillum in the body cuticle and the fine structure of the mouth parts and body appendages. Mitochondrial COI gene of M. melanosticta female isolated in the present study from N. randalli was detected for the first time in Egypt and recorded in the GenBank (MK168807). The study showed that the detected isopod species represents one monophyletic group closely affiliated to the genospecies of M. melanosticta, and can be distinguished clearly from other isopods genospecies. Based on the genetic distance values, lower level of genetic divergence was indicated within the genospecies of the present M. melanosticta isolated from Egypt and the same species isolated from India. The present investigation recorded N. randalli fish as a new host for the isopod M. melanosticta in Egypt and provided additional morphological features through SEM as well as molecular characterization of this isopod species for the first time.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Parasitism is an important factor of negative impacts on the productivity and the economical values of fish production and industry (Başusta et al. 2017). Cymothoidae, Leach, 1818 (Crustacea: Isopoda) are ectoparasites of marine and freshwater fishes and are mostly attached to the buccal or branchial cavity, gills and body surface of their hosts. They are characterized by having various adaptation behaviors to their parasitic lifestyles with great diversity in tropical marine region. All cymothoids are protandric hermaphrodites, and their sexual inversion depends on neurohormonal and androgenic regulation (Raibaut and Trilles 1993). Adult Cymothoids are mostly host and site specific and reported inducing deleterious tissue impacts of variable degrees depending on the species and location of their host (Bunkley-Williams and Williams 1998; Rameshkumar and Ravichandran 2013; Mahmoud et al. 2016). The impact of branchial isopods was reflected on the fish heart and respiratory metabolism (Trilles 1994). The threadfin bream Nemipterus randalli (Perciformes: Nemipteridae) is a widespread fish species in the Red Sea, Gulf of Aqaba (Baranes and Golani 1993), Arabian Gulf, the East African Coast and in the western Indian Ocean. Lelli et al. (2008) and Otero et al. (2013) concluded that previous description of Nemipterus japonicus from the Mediterranean Sea by Ben-Tuvia and Grofit (1973) and Eggleston (1972) and also from the Red sea by Golani and Sonin (2006) was considered to be misidentification and could be referred to as N. randalli. This species was recorded to have established communities in the eastern side of Mediterranean coast as a migrant from the Red sea (Otero et al. 2013). Despite the damage caused by Cymothoid isopods to the fish host, knowledge about their infestation among the Egyptian Red Sea fishes is still lacking and did not receive enough research attention. Although there have been several molecular phylogenetic studies on the family Cymothoidae, the phylogenetic analysis of the cymothoid isopods is seldom strictly confined to the recovery of a pattern of phylogenetic relationships among asset of studied taxa (Avise 2004). Moreover, the previous reference frameworks are unclear as obscure sequences were included in previous phylogenies based on 16S rRNA and COI sequences (Ketmaier et al. 2008). The phenotypic adaptations of cymothoids were restricted by the type of parasitic strategy and may interfere with the choice of accurate morphological features suitable for phylogenic reconstruction (Horton 2000). Some previous studies have successfully focused on the location of phenotypically measurable adaptive features on main chief branches of the phylogenetic tree of several groups of organisms. The reconstruction of the evolutionary history of several high-profile taxa was provided for reinterpretations of the adaptive transformations that led to ecology diversity (Milinkovitch et al. 1994; Springer et al. 2001; Teeling et al. 2002). The present study was conducted to investigate the isopod species parasitizing N. randalli fish, morphometric identification of the isolated isopod species using SEM as well as phylogenetic analysis of the detected species from coastal region of Red Sea, Egypt.
Materials and methods
Sampling and laboratory examination
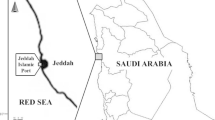
During a survey on the isopod infestation, 415 freshly dead samples of N. randalli of total body length 12–18 (mean 15) cm were seasonally collected from fishing centers at Hurghada, Safaga and Quseir along the Red Sea (Fig. 1). The period of sampling was from December 2017 to November 2018. The specimens were brought to the laboratory of parasitology, Faculty of Veterinary Medicine, Cairo University, to be examined. The body surface, fins, gills, inner wall of operculum, branchial and buccal cavities of each specimen were examined for parasitic isopods. The recovered isopods were measured to the nearest millimeter (mm), counted, washed with saline solution then preserved in 70% ethanol (Hadfield et al. 2014) and identified according to the key of Bruce (1986) and Hadfield et al. (2015). The prevalence, seasonal dynamics and intensity were calculated following Margolis et al. (1982). The examined fish species N. randalli was characterized by the silver pinkish colored body with darker dorsal part and four pale yellow stripes below the lateral line. Identification of the examined fish species was in accordance with Lelli et al. (2008) and Otero et al. (2013).

Red Sea map showing sites of fish sampling
Dissecting stereo-microscope examination
The isolated parasites were examined and photographed under the dissecting stereo-microscope (Olympus Japan SZ40) using digital camera (Canon 12 mega pixel).
Scanning electron microscopy (SEM)
Specimens of the isolated isopods were fixed in 2.5% glutaraldehyde (Colwell et al. 2007) after serial washing in physiological saline solution, then dehydrated by using ethanol series (95% and 100%) for 10 min, followed by the critical point drying in Co2 drier (Autosamdri-815, Germany) according to (Lee 1992). The specimens were mounted over stubs, coated with gold (Spi-Module Sputter Coater, UK) and observed and photographed with scanning electron microscope (JSM 5200, Electron probe microanalyzer, Jeol, Japan) at Faculty of Agriculture, Cairo University.
Molecular analysis
DNA extraction
DNA from the isolated adult female isopods was extracted using DNeasy Tissue Kit (Qiagen, Germany) according to the manufacturer’s instructions.
The cytochrome oxidase subunit (COI) gene of isopods was amplified using the primers, Fish-F1 (5′-TCAACCAACCACAAAGACATTGGCAC-3′) and Fish-R2 (5′-TAGACTTCTGGGTGGCCAAAGAATCA-3′) to get a 776 bp fragment according to Thangaraj et al. (2014). The extracted DNAs from isopods were stored at − 20 °C till use.
DNA amplification polymerase chain reaction (PCR)
The reaction volume was 25 µl, and the mixture contained 12.5 µl of 2 × PCR master mix (Qiagen, Germany), 0.1 μM of each primer, 1 µl of DNA template and completed up to 25 µl with nuclease-free water. The reaction was performed in T100™ Thermal Cycler (Bio-Rad, USA). The reaction was performed under the following PCR conditions: The thermal cycling included 35 cycles of initial denaturation at 94 °C for 2 min, denaturation for 30 s at 95 °C, annealing at 54 °C for 30 s, an extension at 72 °C for 1 min and then a final extension step for 10 min at 72 °C. The negative control was used a nuclease-free water.
Sequence alignment and phylogenetic analysis
After purification of PCR products of positive samples using a QIA quick purification kit (Qiagen, Germany), the purified products were sequenced using Big Dye Terminator V3.1 kit (Applied Biosystems) in ABI 3500 Genetic Analyzer (Applied Biosystems, USA). The results of sequences were matched with those data available in the GenBank using a BLAST server on the NCBI website. Sequences were aligned against other sequences of the COI gene recorded worldwide. The analysis was carried out using the Clustal W, BioEdit software (ver. 7.0.9). A neighbor-joining phylogenetic tree was constructed using MEGA v.6.0 software. A similarity matrix was also constructed using the DNASTAR program (Lasergene, version 8.0). The genetic distance values of inter- and intra-species variations of isopods were also analyzed by MegAlign project of DNSTAR software.
Nucleotide sequence accession number
The partial sequences of M. melanosticta female mitochondrial COI gene were submitted to GenBank, which assigned it the following accession number: MK168807.
Results
The isolated isopod species from N. randalli was identified as M. melanosticta.
Prevalence and intensity
Out of the examined 410 fish samples of N. randalli, 170 were found infested with isopod parasites with an overall prevalence of 40.96% and intensity being 1.27. All the infested fish have one parasite/only one branchial cavity except 46 (27.06%) which was found to be infested with 2 parasites (one/each branchial cavity). The total number of the collected isopods was 216 (214 females and 2 males).
Regarding the seasonal dynamics, isopods showed the highest rate of infestation during spring (57%), while the lowest was in autumn (26.96%) (Table 1).
Site of infestation
Both males and females were in the host branchial cavity with their dorsal aspect toward the gill arch and attached to the inner wall of the operculum cover with their periopods (Fig. 2a, b). Gills of the infested fishes appeared pale in color, and thickening of the gill arches and gill filaments was observed.

a, b The position of M. melanosticta in the branchial cavity of N. randalli; c ventral aspect of the female M. melanosticta showing (marsupium); d the dorsal aspect of the female M. melanosticta showing 7 pereonites (the pereon); e the anterior ventral aspect of the ovigerous female M. melanosticta; f cephalon immersed in the first pereonite; g posterior of dorsal aspect showing pleopods and the uropod; h posterior of the dorsal aspect showing 5 pleonites (The Pleon) and pleotelson; i posterior of the dorsal aspect of male M. melanosticta showing penis on pereonites 7
Description of the isolated M. melanosticta (based on light dissecting stereo-microscope and SEM examination)
Size
The isolated non-ovigerous females M. melanosticta are of 15.9–18.6 (mean 16.5) mm; ovigerous females are 16.3–23.2 (mean 19.2) mm, and males are 9.8–11.6 (mean 10.5) mm long.
Color
The color is light creamy when fresh and of tan color when preserved in alcohol.
Female (based on 10 parasites)
The body is elliptical to ovoid in shape with its greatest width at pereonites 4 and 5. It is slightly bent to one side according to the position of the parasite either in the right or left branchial cavity of the fish host (Fig. 2c–e). The cephalon (Figs. 2f, 3a) is immersed deeply in the first pereonite, and its posterior margin appears slightly curved from the dorsal view. The rostrum is anteriorly truncated (Fig. 3c). The cuticle of the cephalon has multiple irregularly scattered microtrich sensillum (MS) with circular-to-ovoid cuticular collar enclosing the shaft base which can be clearly seen only by SEM at high magnifications (Fig. 3b).The two eyes are located anterolaterally, of oval shape with marked margins and about 0.40–0.55 times the width of the cephalon. Antennules are long, of 8 segments and articulate on the anterior aspect of the cephalon. Antennae are shorter and of 9 segments (Fig. 3b). The mouth parts (Fig. 3d, e, f) are in the anterior portion of the cephalon forming a truncated cone which is confined anteriorly by the labrum (lb), posteriorly by maxillipeds (mb) and laterally by the maxillae (max) and paragnaths (pg). The labrum arises anteriorly from a sclerotized plate forming a triangular rounded arch that covers the anterior mouth parts. The paragnath forms a backward pointing triangular protuberance, and its edge anteriorly forms a rail for the mandible on each side. The mandible (md) is extended with its prolonged proximal region which is elongated into a blade-like edge incisor (inc) and partly surrounding the paragnaths. The incisors of the two mandibles are situated opposing each other. The maxilla median lobe is with 2 spines. The maxilliped is of 3 articles, the apical one is with 4 recurved large setae, and the maxillule is with 4 terminal robust spines. The pereon is of 7 segments of different widths with the maximum at pereonite 5. The anterior margin of pereonite 1 and the posterior margin of pereonite 7 are indented to enclose the cephalon and the first pleonite segment, respectively. Coxae of pereonites 2–4 are fairly visible in dorsal aspect, while coxae of pereonites 5–7 are produced being longer than the segments. Pereopods are large (Fig. 4e–g). Pereopods 1–3 are morphologically similar, directed interiorly and shorter than pereopods 4–7 which are posteriorly directed. The base of pereopod 1 is longer than the width (1.6 times), ischium is longer than the lengths (1.5 times) of carpus and merus, and propodus is longer than its width and the palm is straight, while the dactylus is strongly curved. The brood pouch (marsupium) of gravid female consists of 7 pairs of overlapped oostegites (Fig. 2c). The pleon (Figs. 2h and 4a) consists of 5 segments; the lateral margins of the first are totally overlapped by pereonite 7, and those of pleonites 2 and 3 are partially overlapped, while the pleonites 4 and 5 are uncovered. All pleopods (Figs. 2g and 4b) are with rounded ramai. The lateral lobe of pleopods 1 and 2 is ill developed, while that of the pleopods 3, 4 and 5 is more developed. The pleotelson (Fig. 2h) is short, about 1.1–1.3 times wider than its length and with an evenly rounded posterior margin. The peduncle of the uropod (Fig. 4c) is short with convex lateral margin. The rami are short with tapered narrow apex and slightly reach the posterior margin of the pleotelson. The exopod is longer than the endopod. The microtrich sensilla are scattered on the dorsal and lateral aspect of the body of all the examined isopod specimens and decrease in number and size toward the posterior of the body (Fig. 4h).

Electron microscopy illustrated a dorsal view of the cephalon immersed in the first pereonite; b lateral view of the cephalon showed the antennulae (1), antenna (2) and the microtrich sensillum (arrow); c anterolateral view showed the truncate rostrum and the cone of mouth parts; d–f showed the structure of the mouth parts (lb: labrum, mp: maxillipeds, max: maxilla, mxu: maxillule, pg: paragnaths, md: mandible, inc: incisor)

a Posterior dorsal aspect showed pereonite (pe) 7 and the 5 segments of the pleon (pl); b the rami of the pleopods (plp); c posterior dorsal aspect showed the pleotelson (pt) and the left uropod (up); ped: peduncle, en: endopod, ex: exopod; d ventrolateral margin of the pleon segments and the uropod; e, f, pereopod (pep) 1 and 4, respectively. Co: coxae, ba: base, is: ischium, c: corpus, ps: propodus, d: dactylus; g pereopod (pep) 6 and 7; h the microtrich sensilla scattered on the cuticle of the body dorsal aspect (arrows)
Male (based on 2 parasites)
Morphologically it is similar to female but differ in that the body is not twisted, the eyes are smaller. Penis is on pereonites 7 and appears with two wide than long papillae (Fig. 2i). The pleonites width is nearly subequal. The peduncles on the first and second pleopods and also the uropods are longer with the rami that are of rounded apex.
Molecular finding
Genetic and phylogenetic analysis
To clarify the genetic identity of M. melanosticta, we employed the mitochondrial COI gene sequence. In this study, the PCR assays reveal an amplified DNA fragment of approximately 776 bp (Fig. 5). The phylogenetic analysis revealed that there was a relationship between isopods species and other Mothocya species. From our knowledge, M. melanosticta was recorded the first time in Egypt. Moreover, this species was recorded in the GenBank on the NCBI with accession number: MK168807. Phylogenetic analysis based on mitochondrial COI genes also reveal that this detected isopoda represents one monophyletic group closely affiliated to the genospecies of M. melanosticta, and can be distinguished clearly from other isopods genospecies by neighbor-joining methods (Fig. 6). Based on the COI gene data, there were no clear clades that get separated, the NJ tree having 12 small clades with less than 50% bootstrap value (Fig. 6). The comparison between inter- and intra-species analysis of genetic distance among 12 isopods species revealed that the genetic identity of M. melanosticta isolated from the branchial cavity and gills of N. randalli fish of Red Sea in Egypt was verified with a high sequence homology of 97.5% similarity with M. plagulophor and M. karobran, and of 97.4% similarity with M. melanosticta isolated from India (Fig. 7). On the other hand, low level of genetic divergence (GD) of the isolated M melanosticta from Egypt with the genospecies of M. melanosticta isolated from India (37.5%) and with that of the isopoda species (37.8%) was detected. High genetic divergence was recorded (GD.42.7) among the isolated M. melanosticta from Egypt and Anilocra spp. (Fig. 7). Furthermore, the cymothoid species studied in NJ tree can be distinguished through their site of attachment and sampling location (Table 2).

Agarose gel showing PCR product from DNA genomic of Mothocya melanosticta infested Nemipterus randalli, lane (M): 100 bp DNA ladder, lane (S): positive samples with size marker 77 6 bp, Lane (N): negative control

Neighbor-joining method (NJ) used to estimate phylogenetic relationships among Cymothoidae obtained based on mitochondrial COI sequence

Similarity and genetic divergence of mitochondrial COI sequences of Mothocya melanosticta from Red Sea with the most similar references sequences from the GenBank database
Discussion
The present investigation revealed an overall prevalence of 40.96% for the isopod M. melanosticta among the examined Red Sea N. randalli. This result was nearly similar to that mentioned by Ghobashy (2000) (42.8%) from different Red Sea fish species and El-Halfnawy et al. (2011) (41.6%) for the isopod Irona nanodies isolated from N. japonicum species. On the other hand, lower prevalence of M. melanosticta was reported by Elshahawy and Desouky (2012) from the same locality among the Egyptian pinecone soldierfish and by Vigneshwaran et al. (2019) from the Indian blue flying and Sardinella sindensis fish (31.3% and 22%, respectively). The highest rate of M. melanosticta infestation recorded during the present investigation period was in spring (57%). This record supported the report of Rijin et al. (2018) and Aneesh et al. (2013) for other isopod species from Malabar Coast of India. On the other hand, Elshahawy and Desouky (2012), El-Halfnawy et al. (2011) and Ghobashy (2000) recorded the highest rate of isopod infestation during summer (42.5%, 52.9% and 70%, respectively). These variations in prevalence and seasonal dynamics may be due to the difference in the timing of the investigation period with the subsequent environmental changes and the differences in the types and number of the fish samples examined as well as to the behavior difference of the detected parasitic isopod species. The species M. melanosticta was previously reported from the family Exocoetidae (Bruce 1986; Hadfield et al. 2015), family Holocentridae (Elshahawy and Desouky 2012) and family Clupeidae (Vigneshwaran et al. 2019). Isolation of M. melanosticta in the present investigation is considered the second report after Elshahawy and Desouky (2012) of this isopod species in Egypt, and N. randalli (family: Nemipteridae) is suggested to be identified as a new host for this Cymothoid species.
Detection of M. melanosticta from its host branchial cavity came in agreement with the report of Bruce (1986), in that all the recorded Mothocya species are branchial attaching parasites except Mothocya ihi which was described from its host mouth (Stephenson 1969; Bruce 1986). M. melanosticta seem to be site-specific parasite, the character of which is determined by the needs of the parasite and the adaptation exerted by the morphology and habits of their host (Morton 1974). The body bending of the detected female M. melanosticta was denoted as a remarked adaptation supporting the permanent set of this isopod species in the branchial cavity of their fish host (Panakkool-Thamban et al. 2016). The noticed damages and pale color of the gills of infested fish indicated anemia, which could be due to the homophagous nature of the branchial cymothoids and the obstruction of branchial circulation that resulted from the pressure attachment of the large-sized Mothocya parasite (Bunkley-Williams and Williams 1998).
Genus Mothocya can be differentiated from some other Cymothoid genera in having antennulae that are longer than the antenna, pereopods are with long dactyli, maxilliped article 3 is with 3–5 robust setae and the shape of laminal pleopods (Hadfield et al. 2014). The isolated female and male M. melanosticta from N. randalli was generically identical and morphologically similar to that recorded by Bruce (1986), Elshahawy and Desouky (2012), Vigneshwaran et al. (2019) except minor differences in some measurements, the position of the parasite inside the branchial cavity and the absence of the small rounded protrusion on the lateral side of pereonite II that was described by Elshahawy and Desouky (2012) as a new criterion for M. melanosticta. The smaller size of the recovered M. melanosticta male than that of females is the characteristic which was previously reported for the branchial and buccal parasitic genera of family Cymothoidae (Smit et al. 2014). According to the available studies, the present investigation provided for the first time the detailed morphological characters as well as additional features of M. melanosticta (the microtrich sensillum in the body cuticle, the fine structure of the mouth parts and the body appendages) using stereo-microscope and SEM examination.
The morphological studies of the family Cymothoidae are still facing a challenge to differentiate among species (Smit et al. 2014; Hata et al. 2017). So, molecular markers are of great importance in taxonomic terms specially for bivalves parasitic on various host species (Goto et al. 2012). The present study was based on the mitochondrial COI sequence data of M. melanosticta which was recorded in the GenBank with accession number: MK168807. This gene of M. melanosticta is the first time to be recorded in Red Sea of Egypt. Furthermore, the phylogenetic analysis revealed a relation between M. melanosticta in Egypt with other Mothocya species and Cymothoidae. On the other hand, the cross-relation among COI gene of M. melanosticta and COI gene of hosted fish (N. randalli) was observed, the data which came in agreement with Hata et al. (2017) who reported a molecular similarity among Cymothoid species and host fish. Moreover, the phylogeny of Cymothoids and that of attachment site on host fishes are not concordant, in which each species of isopoda lives in a different site of fish and different country. This result may be related to coevolution among isopods, site of attachment and host fishes, in which Hata et al. (2017) mentioned that Cymothoids living in the buccal cavity and on external body surfaces of fishes have wide ranges of host fish species, suggesting frequent host shift in the coevolution between Cymothoids and host fishes. Finally, the taxonomy of Cymothoidae is important to be detected by molecular besides morphological studies, and the cross-relation among Cymothoidae, host fish, attachment site and environment will need further study.
Conclusion
Mothocya melanosticta is considered a new fauna for the Egyptian Nemipterus randalli. New morphological characters of M. melanosticta were added through dissecting stereo-microscope and SEM examination, depending on phylogenetic analysis of COI gene cross-relation between M. melanosticta in Egypt with other Mothocya species and Cymothoidae. Moreover, the cross-relation among Cymothoidae, host fish, attachment site and environment will need further study.
References
Aneesh PT, Sudha K, Arshad K, Anilkumar G, Trilles JP (2013) Seasonal fluctuation of the prevalence of cymothoids representing the genus Nerocila (Crustacea, Isopoda), parasitizing commercially exploited marine fishes from the Malabar Coast, India. Acta Parasitol 58(1):80–90
Avise JC (2004) Molecular markers, natural history, and evolution, 2nd edn. Sinauer Inc., Sunderland, pp 401–418
Baranes A, Golani D (1993) An annotated list of deep-sea fishes collected in the northern Red Sea, Gulf of Aqaba. Isr J Ecol Evol 39(4):299–336
Başusta N, Mutlu E, Deval MC (2017) Parasitic isopods (Anilocra frontalis H. Milne Edwards, 1830 and Ceratothoa capri (Trilles, 1964)) from the Antalya Bay (Turkey) with new host records. Fırat Univ Turk J Sci Technol 12(1):11–15
Ben-Tuvia A, Grofit E (1973) Exploratory trawling in the Gulf of Suez, November 1972. Fish Fish Breed Isr 8:8–16
Bruce NL (1986) Revision of the isopod crustacean genus Mothocya Costa, in Hope, 1851 (Cymothoidae: Flabellifera), parasitic on marine fishes. J Nat Hist 20(5):1089–1192
Bunkley-Williams L, Williams EH (1998) Isopods associated with fishes: a synopsis and corrections. J Parasitol 84:893–896. https://doi.org/10.2307/3284615
Colwell MA, Hurley SJ, Hall JN, Dinsmore SJ (2007) Age-related survival and behavior of Snowy Plover chicks. Condor 109(3):638–647
Eggleston D (1972) Patterns of biology in the Nemipteridae. J Mar Biol Assoc India 14:357–364
El-Halfnawy MM, Abdel-Mawla HI, Ramadan AM (2011) Isopoda infestation and its effects on the reproductive maturation of Nemipterus japonicus in Red Sea. Egypt J Aquat Res 37(3):275–282
Elshahawy IS, Desouky ARY (2012) First record of Mothocya melanosticta Schioedte and Meinert, 1884 (Isopoda: Cymothoidae) from Egyptian pinecone soldierfish with special reference to its infestation status. Turk J Vet Anim Sci 36(6):577–584
Ghobashy MAA (2000) Morphological and Epidemiological studies on marine fishes ectoparasitic crustacean from the Red Sea. Ph.D. Thesis, Fac. of Science, Suez Canal University
Golani D, Sonin O (2006) The Japanese threadfin bream Nemipterus japonicus, a new Indo Pacific fish in the Mediterranean Sea. J Fish Biol 68(3):940–943
Goto R, Kawakita A, Ishikawa H, Hamamura Y, Kato M (2012) Molecular phylogeny of the bivalve superfamily Galeommatoidea (Heterodonta, Veneroida) reveals dynamic evolution of symbiotic lifestyle and interphylum host switching. BMC Evol Biol 12(1):172. https://doi.org/10.1186/1471-2148-12-172
Hadfield KA, Sikkel PC, Smit NJ (2014) New records of fish parasitic isopods of the gill-attaching genus Mothocya Costa, in Hope, 1851 from the Virgin Islands, Caribbean, with description of a new species. ZooKeys 439:109–125
Hadfield KA, Bruce NL, Smit NJ (2015) Review of Mothocya Costa, in Hope, 1851 (Crustacea: Isopoda: Cymothoidae) from southern Africa, with the description of a new species. Afr Zool 50(2):147–163
Hata H, Sogabe A, Tada S, Nishimoto R, Nakano R, Kohya N, Kawanishi R (2017) Molecular phylogeny of obligate fish parasites of the family Cymothoidae (Isopoda, Crustacea): evolution of the attachment mode to host fish and the habitat shift from saline water to freshwater. Mar Biol 164(5):105. https://doi.org/10.1007/s00227-017-3138-5
Horton T (2000) Ceratothoa steindachneri (Isopoda: Cymothoidae) new to British waters with a key to north-east Atlantic and Mediterranean Ceratothoa. J Mar Biol Assoc UK 80(6):1041–1052
Ketmaier V, Joyce DA, Horton T, Mariani S (2008) A molecular phylogenetic framework for the evolution of parasitic strategies in cymothoid isopods (Crustacea). J Zool Syst Evol Res 46:19–23. https://doi.org/10.1111/j.1439-0469.2007.00423.x
Lee PE (1992) Scanning electron microscopy and X ray microanalysis. Prentice Hall, Englewood Cliffs, p 458
Lelli S, Colloca F, Carpentieri P, Russell BC (2008) The threadfin bream Nemipterus randalli (Perciformes: Nemipteridae) in the eastern Mediterranean Sea. J Fish Biol 73(3):740–745
Mahmoud NE, Fahmy MM, Abuowarda MM, Khattab MS (2016) Parasitic cymothoid isopods and their impacts in commercially important fishes from Lake Qarun, Egypt. Int J Chem Tech Res Mumbai 9(12):221–229
Margolis L, Esch GW, Holmes JC, Kuris AM, Schad G (1982) The use of ecological terms in parasitology (report of an ad hoc committee of the American Society of Parasitologists). J Parasitol 68(1):131–133
Milinkovitch MC, Meyer A, Powell JR (1994) Phylogeny of all major groups of cetaceans based on DNA sequences from three mitochondrial genes. Mol Biol Evol 11(6):939–948
Morton B (1974) Host specificity and position on the host in Nerocila phaeopleura Bleeker (Isopoda, Cymothoidae). Crustaceana 26:143–148
Otero M, Cebrian E, Francour P, Galil B, Savini D (2013) Monitoring marine invasive species in Mediterranean marine protected areas (MPAs): a strategy and practical guide for managers. IUCN, Malaga, p 136
Panakkool-Thamban A, Kottarathil HA, Kappalli S (2016) Branchial cymothoids infesting the marine food fishes of Malabar coast. J Parasit Dis 40(4):1270–1277
Raibaut A, Trilles JP (1993) The sexuality of parasitic crustaceans. In: Advances in parasitology, vol 32. Academic Press, pp 367–444
Rameshkumar G, Ravichandran S (2013) Histopathological changes in the skins and gills of some marine fishes due to parasitic isopod infestation. J Coast Sci Med 1(1):74–80
Rijin K, Sudha K, Vineesh PJ, Anilkumar G (2018) Seasonal variation in the occurrence of parasitic isopods and copepods (crustacea) infecting the clupeidaen fishes of Malabar Coast, India. Turk J Fish Aquat Sci 19(3):241–249
Smit NJ, Bruce NL, Hadfield KA (2014) Global diversity of fish parasitic isopod crustaceans of the family Cymothoidae. Int J Parasitol Parasit Wildl 3(2):188–197. https://doi.org/10.1016/j.ijppaw.2014.03.004
Springer MS, Teeling EC, Madsen O, Stanhope MJ, de Jong WW (2001) Integrated fossil and molecular data reconstruct bat echolocation. Proc Natl Acad Sci 98(11):6241–6246
Stephenson AB (1969) Irona Melanosticta (Isopoda: Cymothoidae): a new record for New Zealand waters, with descriptions of male, female and larval states. Rec Auckl Inst Mus 6:427–434
Teeling EC, Madsen O, Van Den Bussche RA, de Jong WW, Stanhope MJ, Springer MS (2002) Microbat paraphyly and the convergent evolution of a key innovation in Old World rhinolophoid microbats. Proc Natl Acad Sci 99(3):1431–1436
Thangaraj M, Saranya S, Divya S, Ramanadevi V, Subburaj J (2014) Molecular phylogenetic status of some marine Cymothoid isopods in southeast coast of India. Indian J Geo Mar Sci 43(2):271–276
Trilles JP (1994) Les Cymothoidae (Crustacea, Isopoda) du monde (Prodrome pour une faune). Stud Mar 21(22):1–288
Vigneshwaran P, Ravichandran S, Martin MB, Rajan DK (2019) First record of parasitic isopod Mothocya melanosticta (Schiödte and Meinert, 1884) (Isopoda: Cymothoidae) on blue flying fish Exocoetus volitans and Indian sardine Sardinella sindensis from the southeast coast of India. Thalassas Int J Mar Sci 35(1):99–107
Acknowledgements
We are grateful to the fisher men, Hurghada, Safaga and Quseir along the Red Sea, for helping to collect the specimen.
Author information
Authors and Affiliations
Contributions
NM contributed to the study design, surveillance, and morphological isopod species identification; MF contributed to the study design, reviewing the paper, and species identification; MA contributed to the study design, molecular characterization, data analysis, and reviewing the paper.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there is no conflict of interest.
Informed consent
Consent was obtained from fisher men included in this study.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Mahmoud, N.E., Fahmy, M.M. & Abuowarda, M.M. Additional morphometric and phylogenetic studies on Mothocya melanosticta (Isopoda: Cymothoidae) parasitizing the Red Sea Nemipterus randalli fish in Egypt. J Parasit Dis 44, 289–298 (2020). https://doi.org/10.1007/s12639-020-01194-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12639-020-01194-9