
Abstract
Three locally isolated strains of entomopathogenic nematodes (EPNs), viz. Heterorhabditis indica, Steinernema thermophilum and Steinernema glaseri, from Meghalaya, India were characterized in terms of storage temperature and survival and infectivity of their infective juveniles (IJs). The survival and infectivity of nematode IJs was studied at, 5 ± 2 and 25 ± 2 °C, for a period of 120 days, using deionized water as storage medium. The viability of nematode IJs was checked by mobility criterion at different storage periods, while the infectivity of nematode IJs was ascertained on the basis of establishment of IJs, using Galleria mellonella larva mortality tests in petridishes. The results of this study revealed that storage temperature markedly affects the survival as well as the establishment of nematode IJs of the three EPN species. At 5 °C, comparatively higher rate of IJ’s survival (i.e. 74–86 %) was observed for 15 days of storage, but the same reduced drastically to 28–32 % after 30 days of storage for H. indica and S. thermophilum. On the other hand, at 25 °C, the survival of nematode IJs was observed till 120 days for all the three studied EPNs. In case of S. thermophilum and S. glaseri, higher rate of IJs survival (>75 %) was observed respectively at 15 and 30 days of observation. The study also showed that the establishment of IJs of the three EPN species declines with increase in storage periods, at both the test temperatures. In general, the nematodes stored at 25 °C showed comparatively better establishment than those stored at 5 °C. Among the three EPN studied, the establishment of S. glaseri was comparatively better than the rest of the species at both the temperatures and for different storage durations. In conclusion, our study adds further valuable information about the effect of storage temperature on survival and infectivity of three indigenous EPN species of Meghalaya, India which appears to be promising biocontrol agents of local insect pests.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Entomopathogenic nematodes (EPNs) of the families Heterorhabditidae and Steinernematidae are obligate parasites of insects and are frequently used as biological control agents of many soil-inhabiting insect pests (Georgis et al. 2006). At present, there are about a dozen of EPN species that are available commercially as biocontrol agents for various insect pests. By and large, most of these commercial EPNs target the high-value crop pests, either in North America or European countries, and a few such products are also being used in Korea (Kaya et al. 2006). Many studies have demonstrated that the native species/isolates of EPNs, that are adapted to local environmental conditions, are especially good biocontrol agents for local insect pests (Koppenhöfer and Kaya 1999). Several previous studies have also reported that the isolates of EPNs from different geographical regions exhibit considerable variations in terms of their host range, infectivity potentials, reproduction, tolerance of various environmental factors, etc (Bedding 1990). Therefore, to a great extent, the biocontrol potentials of different EPN species/strains are greatly influenced by a number of specific abiotic and biotic factors (Kaya 1990; Koppenhöfer et al. 1995).
In India, prior to any scientific report about the occurrence of indigenous EPNs, initially attempts were made to utilize a few exotic species/strains of EPNs, like Steinernema carpocapsae, Steinernema feltiae, Heterorhabditis bacteriophora, etc that were imported by the researchers from other countries, as biocontrol agents (Rao et al. 1971; Hussaini 2003). However, in none of these attempts the exotic EPN isolates were able to establish successfully in the fields due to their poor adaptability to the prevailing agro-climatic conditions of the country. As a result, in the later years, attention was diverted to search the native strains of EPNs, assuming that perhaps they may adapt well to the local climatic conditions of the country (Nayak et al. 1977; Singh 1977). The surveys made in different regions of India have resulted into isolation of a number of indigenous species/isolates of EPNs (Rahman et al. 2000; Hussaini 2003). Ever since, efforts are being made to understand the basic biology and ecology of the native EPN isolates, so as to explore their possible use against the local insect pests of the country (Ganguly and Singh 2001; Ganguly and Gavas 2004a, b). However, at present, none of the indigenous EPN species/strain is available as commercial product for possible use against any insect pest in India.
In a recent study, three indigenous isolates of EPNs, viz. Heterorhabditis indica—Poinar, Karunakar and David, Steinernema thermophilum—Ganguly and Singh and Steinernema glaseri (Steiner) were isolated from the forest soils in Meghalaya, India (Lalramliana 2007). In laboratory trials, the isolates of these nematodes showed promising control against two important local insect pests, viz. the mustard sawfly, Athalia lugens proxima (Klug) and against a chrysomelid beetle, Aplosonyx chalybaeus (Hope), a pest of taro (Yadav and Lalramliana 2012a, b). It is widely recognized now that the selection of a suitable EPN species as a biocontrol agent in a given geographical area requires adequate knowledge about the biology and ecology of the nematode. Information relating to the above three EPN isolates, such as the effects of soil moisture on the establishment and the effects of relative humidity on the emergence and reproduction were studied by Lalramliana and Yadav (2009) and Yadav and Lalramliana (2012c). Among other abiotic factors, the temperature is also considered as an important factor in determining the suitability of EPN as an appropriate biocontrol agent. For example, the temperature may affect the mobility, development, persistence, survival, reproduction, etc of nematodes in several ways (Griffin 1993; Grewal et al. 1994; Koppenhöfer and Kaya 1999; Ganguly and Singh 2001; Cagnolo and Campos 2008). The effects of temperature on survival and infectivity of nematodes infective juveniles (IJs) have also been studied for some EPNs. In these studies, the different isolates of the same species have also been found to exhibit differential responses to storage temperatures (Cagnolo and Campos 2008). In view of this, it is necessary to characterize the indigenous nematodes to be used for the biocontrol of local insect pests in terms of temperature requirements as well. Hence, in the present study, attempts have been made to find out the influence of storage temperature on survival and infectivity of isolates of three indigenous EPNs, i.e. H. indica, S. thermophilum and S. glaseri.
Materials and methods
The isolates of nematodes used in this study, viz. H. indica, S. glaseri and S. thermophilum, were collected using Galleria mellonella traps from the forest soils of Ri-Bhoi District (91′40′16″ latitude and 25′40′–26′20′N longitude) in Meghalaya, India. These EPNs have been duly identified using light and scanning electron microscope and also by morphometric criteria (Lalramliana 2007). For the present study, the nematodes were reared in the laboratory on late instar larvae of G. mellonella at 25 °C, as described by Woodring and Kaya (1988). The freshly emerged IJs (3–5 days after first day of emergence) of each nematode species were then collected from wax moth larval cadavers, using modified white traps (Woodring and Kaya 1988), and used for various experiments.
To determine the effect of storage temperature on survival and infectivity of EPNs, the population densities of IJs were adjusted to 1,000 IJs/ml in 20 ml of de-ionized water in 50 ml conical flasks. The flasks were then plugged with non-absorbent cotton and kept inside BOD incubators at two different temperatures, i.e. 5 ± 2 and 25 ± 2 °C, for 120 days. The selection of test temperatures was done in accordance with the prevailing average minimum and maximum monthly temperatures of the area from where the nematode isolates were collected. Before setting-up the experiment, the viabilities of nematode IJs were checked by observing their movements under a stereomicroscope. Accordingly, the nematode populations were assumed to be 100 % viable on the first day of experiment. The survival of IJs in the suspensions was monitored at 15, 30, 60, 90 and 120 days of storage periods for each test temperature. For this, about 1 ml of IJs suspension was withdrawn from each conical flask and the viability of IJs was determined under a stereomicroscope by mobility criterion. To ascertain the mobility status of IJs, the immobile IJs were first located in the suspension and later they were touched with a fine-wire probe, those that did not react by touching were considered as dead IJs. To determine the establishment percentage of nematodes, the infectivity tests of IJs were done by petridish assay, using eight numbers of G. mellonella larvae at the concentration of 100 IJs/larva and the number of IJs established/larva were recorded (Lalramliana 2007). For control group, the establishments of IJs were also checked on the day the experiments were set-up (shown as day 0), wherein the same numbers of moth larvae were exposed to 100 IJs from the original stock suspension of IJs to record the number of IJs established/larva. There was three replicates for each test temperature and IJs observation for survival and infectivity tests.
Statistical analysis
The data were analyzed statistically and are represented as mean ± standard error of the mean (SEM). The significance of the difference was determined by the one-way analysis of variance (ANOVA) and student’s t test. P values <0.05 were accepted as statistically significant. Correlation between the parameters was determined by regression analysis.
Results
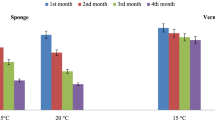
The data regarding the survival and establishment of EPN IJs after storage at 5 °C for 120 days are shown in Fig. 1. The storage temperature showed a marked effect on both the survival and establishment of the IJs of three EPN species. At 5 °C, the survival of IJs was observed only till 60 days for H. indica and S. thermophilum. At this storage condition, comparatively higher rates of IJ’s survival (i.e. 74–86 %) were observed for 15 days of storage for both these EPN species. The IJs survival, however, reduced drastically to 28–32 % after 30 days of storage and no survival of nematode IJs was observed beyond 60 days storage for both these EPNs. In contrast to H. indica and S. thermophilum, the IJs of S. glaseri survived till 120 days and the highest rate of survival (99.37 %) was observed for 15 days storage period. At the same temperature, the infectivity of EPN IJs (as measured by rate of IJs establishment in G. mellonella) was also significantly (p < 0.05) affected by storage durations. The infectivity of H. indica and S. thermophilum IJs was maximum at 15 days storage period. However, the IJs of S. glaseri showed the maximum infectivity rate at 30 days storage time (Fig. 1).

Percentage survival and establishment of infective juveniles (IJs) of entomopathogenic nematode species at 5 °C. a Survival of IJs and b establishment of IJs
The data pertaining to the survival and establishment of EPN IJs after storage at 25 °C are presented in Fig. 2. At this temperature, the survival of nematode IJs was observed till 120 days for all the three test EPN species. The rate of IJ survival was moderately high till 90 days observation time, but reduced significantly thereafter. As far as the infectivity of IJs is concerned, significant differences (p < 0.05) were observed in the infectivity of IJs of H. indica and S. thermophilum from 90 days to 120 days storage periods with the IJs from control. However, no such differences were observed in the infectivity of IJs of S. glaseri when compared to the infectivity of IJs from control at 120 days storage period (Fig. 2).

Percentage survival and establishment of infective juveniles of (IJs) of entomopathogenic nematode species at 25 °C. a Survival of IJs and b establishment of IJs
Discussion
One of the important requirements in utilizing EPNs as biocontrol agent for insect pests is the proper storage of EPN IJs, in a given population and also at optimum temperature, so that maximum IJs can survive for long periods before they are used for field applications. The aim of this study was to characterize the three locally isolated strains of EPNs from Meghalaya (H. indica, S. thermophilum and S. glaseri) in terms of storage temperature on survival and infectivity of IJs. It was observed in this study that, at the lower temperature (5 °C), the survival rate of H. indica and S. thermophilum IJs drastically reduced beyond 60 days and no survival of IJs was observed after 90 days storage. On the other hand, at the same temperature, the survival of IJs was observed even up to 120 days for S. glaseri. At the higher temperature (25 °C), survival of nematode IJs was, however, observed till 120 days for all the three studied nematode species. Nevertheless, at this temperature, the survival rate of H. indica and S. thermophilum was comparatively low (<25 %) as compared to S. glaseri which revealed >60 % survival. It is thus obvious from these results that the IJs of these EPN species can remain viable for at least 60 days. Poinar (1979) reported the favorable temperature for the storage of Steinernema sp. varies between 5 and 9 °C. Likewise, Bedding (1981) also reported that 12 °C is the best storage temperature for Heterorhabditis spp. Our present observation on EPN IJs survivals at 25 °C is in disagreement with the findings of Poinar (1979) and Bedding (1981). A possible reason behind these observed differences may be that in the present study the isolates of both H. indica and S. thermophilum species were isolated from the areas where the mean monthly temperature varies between 20 and 25 °C and this may perhaps be a possible reason as to why at lower temperature of 5 °C the nematode IJs showed relatively less survival beyond 30 days storage periods. In a related study, Katti et al. (2006) reported 100 % survival of S. thermophilum after storage up to 50 days at 28 °C, but thereafter the survival declined to 10 % after 150 days of storage. The differences in survival of two S. thermophilum isolates could be due to differences in their natural origins. While our isolates were collected from forest soils in Meghalaya, the EPNs in Katti et al. studies were collected from soils in Hyderabad. In another study, Strauch et al. (2000) reported that a maximum survival of H. indica can be achieved at 15 °C and the highest mortality of IJs occurs at 5 °C. The increased mortality at low temperature observed in this study agrees with the findings of Cagnolo and Campos (2008) where the IJs of Steinernema rarum stored at 5 ± 1 °C showed less survival whereas those stored at 23 ± 2 °C showed above 95 % survival rate. The observed survival of IJs of the three EPN species at 25 °C for 120 days duration also reflects the longevity of IJs without the host at the prevailing temperature, from where these isolates were collected. Furthermore, a comparatively high rate of survival of S. glaseri at 25 °C may be due to the slower utilization of lipid reserves in this species as compared to H. indica and S. thermophilum. It is reported that large nematode species, like S. glaseri, survive starvation longer than small species with the same percentage lipid content (Atkinson 1980). Grewal (2000) also opined that the differences in longevity of IJs between species of Steinernema correlates very well with the initial lipid content and with the rate of lipid utilization by nematode.
Apart from these results, in the present study, though the IJs establishment showed a decreasing trend following the storage period, the insect mortality was observed to be still high wherever IJs established. In a related study, Cagnolo and Campos (2008), while comparing the effect of storage temperature (5 ± 1 and 23 ± 2 °C) on survival and infectivity of S. rarum, reported that both infectivity and survival of IJs would be higher when stored at 23 ± 2 °C. Furthermore, Fan and Hominick (1991) also reported that IJs of Steinernema sp. and S. feltiae, after being stored in sand at 5 °C, lose their ability to parasitize a host, however, effects of storage temperature on infectivity were not reflected in the data for mortality of the insects, which generally remained high in all the tests. In contrast, O’Leary et al. (1998) found that the host finding ability of IJs of H. megidis improved with increased duration of storage.
Taken together, the results of our study indicate that the three EPN species show variations in their survival and infectivity with regard to storage periods. A storage temperature of 25 °C appears comparatively better for the storage of all the three EPNs, as at this temperature they can remain viable up to 2 months period. However, from infectivity point of view, at 25 °C these EPNs isolates can be best stored for about a month, as their infectivity lowers afterwards. It is expected that these ecological data on effects of storage temperature on isolates of indigenous strains of EPNs from Meghalaya may be useful in exploring the role of these species as the biocontrol agents of local insect pest species.
References
Atkinson HJ (1980) Respiration in nematodes. In: Zuckerman BM (ed) Nematodes as biological models, vol 2. Academic Press, London, pp 101–142
Bedding RA (1981) Low cost in vitro mass production of Neoaplectana and Heterorhabditis species (Nematoda) for field control of insect pests. Nematologica 27:109–114
Bedding RA (1990) Logistics and strategies for introducing entomopathogenic nematode technology in developing countries. In: Gaugler R, Kaya HK (eds) Entomopathogenic nematodes in biological control. CRC Press, Boca Raton, pp 233–248
Cagnolo S, Campos V (2008) Effect of storage temperature on survival and infectivity of Steinernema rarum (OLI strain) (Rhabditida: Steinernematidae). J Invertebr Pathol 98:114–115
Fan X, Hominick WM (1991) Effects of low storage temperature on survival and infectivity of two Steinernema species (Nematoda: Steinernematidae). Rev Nématol 14:407–412
Ganguly S, Gavas R (2004a) Effect of soil moisture on the infectivity of entomopathogenic nematode, Steinernema thermophilum Ganguly & Singh. Int J Nematol 14:78–80
Ganguly S, Gavas R (2004b) Host range of entomopathogenic nematode, Steinernema thermophilum Ganguly and Singh (Steinernematidae: Rhabditida). Int J Nematol 14:221–228
Ganguly S, Singh LK (2001) Optimum thermal requirements for infectivity and development of an indigenous entomopathogenic nematode, Steinernema thermophilum Ganguly & Singh 2000. Indian J Nematol 31:148–152
Georgis R, Koppenhöfer AM, Lacey LA, Bélair G, Duncan LW, Grewal PS, Samish M, Tan L, Torr P, van Tol RWHM (2006) Successes and failures in the use of parasitic nematodes for pest control. Biol Control 38:103–123
Grewal PS (2000) Anhydrobiotic potential and long-term storage of entomopathogenic nematodes (Rhabditida: Steinernematidae). Int J Parasitol 30:995–1000
Grewal PS, Selvan S, Gaugler R (1994) Thermal adaptation of entomopathogenic nematodes: niche breadth for infection, establishment, and reproduction. J Therm Biol 19:245–253
Griffin CT (1993) Temperature responses of entomopathogenic nematodes for the success of biological control programs. In: Bedding RA, Akhurst RJ, Kaya HK (eds) Nematodes and the biological control of insect pests. CSIRO Publications, East Melbourne, pp 101–111
Hussaini SS (2003) Progress of research work on entomopathogenic nematodes in India. In: Hussaini SS, Rabindra RJ, Nagesh M (eds) Current status of research on entomopathogenic nematodes in India. Project directorate of biological control, Bangalore, pp 27–68
Katti G, Prasad JS, Padmakumari AP, Sankar M (2006) Effect of storage periods on survival and infectivity of indigenous entomopathogenic nematodes of insect pests of rice. Nematol Medit 34:37–41
Kaya HK (1990) Soil ecology. In: Gaugler R, Kaya HK (eds) Entomopathogenic nematodes in biological control. CRC Press, Boca Raton, pp 93–116
Kaya HK, Aguillera MM, Alumai A, Choo HY, de la Torre M, Fodor A, Ehlers RU (2006) Status of entomopathogenic nematodes and their symbiotic bacteria from selected countries of regions of the world. Biol Control 38:134–155
Koppenhöfer AM, Kaya HK (1999) Ecological characterization of Steinernema rarum. J Invertebr Pathol 73:120–128
Koppenhöfer AM, Kaya HK, Taormino SP (1995) Infectivity of entomopathogenic nematodes (Rhabditida: Steinernematidae) at different soil depths and moistures. J Invertebr Pathol 65:193–199
Lalramliana (2007) Faunal composition and distribution of entomopathogenic nematodes and their bioefficacy against major insect pests in Ri-bhoi district of Meghalaya. PhD Thesis, North-Eastern Hill University, Shillong, India
Lalramliana, Yadav AK (2009) Effect of relative humidity on the emergence and reproduction of three entomopathogenic nematode species (Rhabditida: Steinernematidae and Heterorhabditidae) from Meghalaya, India. Sci Vis 9:92–96
Nayak P, Rao YRVJ, Yadava CP, Rao YS (1977) Occurrence of a new entomophilic nematode on rice stem borer, Sesamia inferens. Oryza 14:51–54
O’Leary SA, Stack CS, Chubb MA, Burnell AM (1998) The effect of day of emergence from the insect cadaver on the behavior and environmental tolerances of infective juveniles of the entomopathogenic nematode Heterorhabditis megidis (strain UK211). J Parasitol 84:665–672
Poinar GO Jr (1979) Nematodes for biological control of insects. CRC Press, Boca Raton
Rahman PF, Sharma SB, Wightman JA (2000) A review of insect parasitic nematodes research in India: 1927-1997. Int J Pest Manag 46:19–28
Rao YR, PrakasaRao PS, Verma A, Israel P (1971) Tests with an insect parasitic nematode DD-136 (Nematoda: Steinemematidae) against rice stem borer, Tryporyza incertulas walk. Indian J Entomol 33:215–217
Singh SP (1977) Isolation of entomophilic nematode from potato cutworms. Curr Sci 46:454–455
Strauch O, Niemann I, Neumann A, Schmidt AJ, Peters A, Ehlers RU (2000) Storage and formulation of the entomopathogenic nematodes Heterorhabditis indica and H. bacteriophora. Biol control 45:483–500
Woodring JL, Kaya HK (1988) Steinernematid and heterorhabditid nematodes: a handbook of techniques southern cooperative series bulletin 331. Arkansas Agricultural Experiment Station, Fayatteville
Yadav AK, Lalramliana (2012a) Evaluation of the efficacy of three indigenous strains of entomopathogenic nematodes from Meghalaya, India against mustard sawfly, Athalia lugens proxima Klug (Hymenoptera: Tenthredinidae). J Parasit Dis 36:175–180
Yadav AK, Lalramliana (2012b) Efficacy of indigenous entomopathogenic nematodes from Meghalaya, India against the larvae of taro leaf beetle, Aplosonyx chalybaeus (Hope). J Parasit Dis 36:149–154
Yadav AK, Lalramliana (2012c) Soil moisture effects on the activity of three entomopathogenic nematodes (Steinernematidae and Heterorhabditidae) isolated from Meghalaya, India. J Parasit Dis 36:94–98
Acknowledgments
Authors thank the Head, Department of Zoology, NEHU, Shillong for providing the necessary research facilities. Lalramliana was recipient of UGC-CSIR, NET Research fellowship.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Lalramliana, Yadav, A.K. Effects of storage temperature on survival and infectivity of three indigenous entomopathogenic nematodes strains (Steinernematidae and Heterorhabditidae) from Meghalaya, India. J Parasit Dis 40, 1150–1154 (2016). https://doi.org/10.1007/s12639-014-0639-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12639-014-0639-8