
Abstract
Coccidiosis is the most important protozoan disease affecting the poultry industry worldwide. Control of poultry coccidiosis is presently based on managerial skills and the use of prophylactic coccidiostatic drugs. With the emergence of drug resistant Eimeria strains, emphasis has been laid on development and use of safer vaccines; some of them have been commercialized successfully. The present review deals with the various factors responsible for the development of clinical coccidiosis in poultry as well as an overview of the currently available inducers and boosters of immunity against coccidiosis. There are three groups of vaccines currently available against coccidiosis which can be distinguished on the basis of characteristics of the Eimeria species included in the respective products, viz. vaccines based on live virulent strains, vaccines based on live attenuated strains, and vaccines based on live strains that are relatively tolerant to the ionophore compounds. The latter vaccine combines the early chemotherapeutic effect of ionophores with the late prophylactic effect of vaccination. Although in the near future more varieties of oocyst based live vaccines are expected, identification of selective coccidian-specific immunoprotective molecules is likely to get more attention to facilitate the sustainable control of poultry coccidiosis.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
During the last two decades the growth of poultry industry has been remarkable and India presently holding the third position in the world in egg production and poultry meat production has reached about 2.2 million tons per year. It is evident that the poultry industry is not only playing a great role for the nutritional security of the country, but also providing working opportunity to a sizeable population of the world. About three million people in India are directly linked to the poultry industry which in turn contributing to the national economy to the extent of US $5.7 billion annually. Although most of the diseases of infectious origin affecting the poultry industry have been controlled successfully using protective vaccines, coccidiosis caused by several Eimera spp. is still considered as the most challenging deterrent for poultry development. Intensive system of poultry rearing provides excellent opportunities for accumulation and transmission of the coccidian oocysts and in the absence of effective control measures clinical disease is the inevitable outcome resulting in extensive mortality of the young stocks. The reduced production efficiency and the costs of veterinary and prophylactic interventions resulting from poultry coccidiosis costs the UK poultry industry alone in excess of £38M per annum and the global loss due to poultry coccidiosis is estimated to exceed £500M per annum (Shirley et al. 2007) which may be considered as the singular reason for devising improved control strategies.
Eimerian reproduction
The lifecycle of the organisms includes development both inside and outside of the host. Within the host the parasite undergoes both asexual and sexual stages of development. In general, oocysts voided through feces get sporulated within about 24 h in the external environment in presence of oxygen. The excystation of the oocysts within the intestinal lumen is facilitated by trypsin, bile, and CO2 and the released sporozoites penetrate the villous epithelial cells. Sporozoites of some species (E. brunetti and E. praecox) develop within cells at the site of penetration. Sporozoites of other species (E. acervulina, E. maxima, E. necatrix, and E. tenella) are transported to other sites, viz. the crypt epithelium, where they develop as a trophozoites (Lawn and Rose 1982; Trout and Lillehoj 1993). Within the host cells, trophozoites undergo asexual reproduction (schizogony or merogony) to produce merozoites which penetrate healthy enteric cells. A few cycles of merogony take place which is followed by sexual reproduction or gametogeny. Merozoites enter host cells and differentiate into either male (microgamonts) or female (macrogamonts) forms. The microgamonts divide to form microgametes, which fertilize the macrogamonts leading to the development of oocysts which are voided through the feces. The prepatent periods generally range from 4 to 5 days following oral infection and maximum oocyst output ranges from day 6 to 9 postinfection. The precise biochemical and genetic mechanisms which control the development of Eimeria spp. within host cells is yet to be fully understood. Two linkage groups associated with intracellular coccidian development have been identified on chromosome No.1 and 2 of E. tenella using precocious and drug resistant lines, (Shirley and Harvey 1996). This information may help identifying other genetic loci involved in regulating the life cycle of E. tenella. Recently a developmentally regulated gene (ets3a) (Ouarzane et al. 1998) has been identified.
Diagnosis and identification at species level
Each of the seven valid species of chicken coccidia is developmentally location-specific within the chick digestive tract. It is common to find at least six species (e.g., E. acervulina, E. maxima, E. tenella, E. brunetti, E. mitis, and E. praecox) in litter samples from a single flock during its first 6 weeks (Williams 1995). Identification of the five species, viz. E. acervulina, E. brunetti, E. maxima, E. necatrix, and E. tenella, is comparatively easy based on characteristic gross lesions. The pathogenicity ranges from moderate to severe. Although E. praecox and E. mitis are considered non pathogenic, experimental infections with these species may produce pathogenesis resulting in enteritis, diarrhoea, and reduced feed efficiencies (Williams 1998), which warrants their inclusion in the list of coccidia to be controlled. Generally, the five most pathogenic species can be differentiated in the host on the basis of clinical signs, characteristic tissue lesions, prepatent period, size of oocysts, and morphology of intracellular stages.
Several molecular biological approaches have been successfully employed, viz. differentiation of species on the basis of isoenzyme patterns of oocysts by starch block electrophoresis (Shirley 1975), use of the rRNA and rDNA probes for identification of individual species through characteristic restriction fragment patterns (Ellis and Bumstead 1990), a randomly amplified polymorphic DNA assay to differentiate E. acervulina and E. tenella and their strains (Procunier et al. 1993). Molecular tools and markers have been developed for differentiation of different strains of E. tenella as well as precocious and drug-resistant strains (Shirley 1994; Shirley and Harvey 1996). Sensitive detection and differentiation of the major coccidian species have been possible applying PCR. PCR amplification of internal transcribed spacer region 1 from genomic DNA has been used to detect and differentiate six Eimeria species (Schnitzler et al. 1997). A two-step PCR technique was used for differentiation of eight coccidian species. A PCR RFLP technique was used for characterisation of six species of coccidia in Australia (Woods et al. 2000). However, conventional PCR has limitations in quantifying the number of Eimeria oocysts present in samples as well as detection of mixed-species infections which can be overcome by applications of real-time PCR. A TaqMan® probe based qPCR targeting the nuclear ribosomal DNA (ITS-2) has been used for detection and quantification of seven species of Eimeria infecting chickens opening new vistas for complex epidemiological studies(Morgan et al. 2009).
Sustainable control of coccidiosis
Management of poultry house
Oocysts of coccidia with high reproduction potential can easily disseminate in the poultry house environment and it is a challenging task to keep chickens coccidia free, especially under intensive rearing conditions. Oocysts sporulate readily in the litter. Nevertheless, the sporulation of oocysts is affected by degradation, bacterial damage as well as ammonia generated from the litter which may affect their viability (Williams 1995). In the USA, removal of caked litter and aeration of the poultry houses at the interval of 2–3 weeks is practised and top dressing with fresh litter before placing a new flock is a rule. On the other hand, a thorough cleanout between flocks is a common practice in most European countries and Canada. Strict biocontrol measures adopted by the caretakers of poultry houses can play a great role in restricting the spread of infective oocysts (Allen and Fetterer 2002).
Prophylactic application of anticoccidials
The use of anti coccidial feed additives during past 50 years has played a major role in the growth of the poultry industry and has facilitated increased availability of high quality, affordable poultry products to the consumers. On the basis of specific modes of action, the anticoccidials may be classified as (i) chemicals affecting parasite metabolism, such as halofuginone, clopidol, decoquinate, amprolium, or (ii) polyether ionophores such as lasalocid, monensin, narasin, maduramycin, and salinomycin, which alter ion transport and disrupt the osmotoic balance. These latter compounds are now the mainstay of coccidiosis control (Jeffers 1997). It is quite an alarming fact that some degree of resistance to all anticoccidial drugs, including ionophores, has developed (Chapman 1994, 1998). To overcome the problem of resistance, poultry producers are practising rotational use of various anticoccidials with successive flocks or combination of chemical and ionophore treatments. Identification of the enzymes of sporozoite mannitol cycle (Allocco et al. 1999) and histone deacetylase from the trophozoites (Schmatz 1997) might be helpful in the development of new anticoccidial.
Immunoprophylaxis
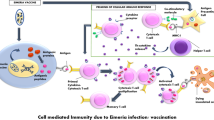
The immunity induced following infection with avian Eimeria is species specific. Parasitic stages that are thought to be important in generating protective immune responses include the initial asexual developmental stages where trophozoites multiply within the epithelial cell of intestinal villi (Jenkins et al. 1991). T cell and cytokines play an important role in protective immunity to Eimeria. While Th1 type cytokines play an important role in limiting parasite multiplication in the early stages of infection, regulatory cytokines and other immune cells may play a vital role in limiting the immunopathology associated with proinflammatory cytokines (Roberts et al. 1996).
Protective immunity in chickens can be induced by deliberate infection with low numbers of Eimeria parasites and it is established that following two to three consecutive infections the birds develop solid immunity. All commercially available coccidiosis vaccines are based on this principle. Depending on the characteristics of the vaccine strains used, these vaccines may be divided into three groups.
Live, virulent strain based vaccine
These vaccines incorporate a variable number of wild type strains (Lee 1987). For broiler-breeders up to eight Eimeria species are included in these products (Coccivac® D, Immucox® C2), where as for use in the broiler industry the number is restricted up to four species (Coccivac® B, Immucox® C1). The mode of application is with drinking water for all the four live vaccines. Besides drinking water Immucox C1 can also be applied by incorporating in an oral gel (Vermeulen et al. 2001).
Live, attenuated strain based vaccines
The attenuated lines of Eimeria parasites can be developed through repeated selection for early maturation (pre-cociousness) or by serial passage through embryonated eggs (Long 1972; Jeffers 1975). The most important feature of these attenuated lines is their reduced proliferative capacity resulting in less damage to the intestinal epithelium after one passage through the gut. Paracox® (precocious strains; Shirley and Millard 1986) and Livacox® (precocious: TA strains, Bedrnik, 1989) are the example of such attenuated vaccine and the performance was comparable to anticoccidial drugs (Williams et al. 1999).
Establishment of a low level of infection in the early part of life through immunization of the chicks with live or attenuated strains helps in boosting the immune system of the naive birds in the litter and thereby protect from a future fulminating disease. Uses of therapeutics or feed additives that interfere with development of the oocysts are not permisible during the period of development of immunity. Therefore, the risk of development of clinical coccidiosis is highest at the early age (weeks 1–3) which decreases as immunity starts developing (from weeks 3–4 onwards) (Vermeulen et al. 2001). A live vaccine comprising an precocious attenuated line of E. tenella and a non-attenuated line of each E. maxima and E. acervulina, namely Supercox1, has been introduced with the promise of controlling clinical coccidiosis in broilers as well as achievement of production performance superior to that of using anticoccidial drugs against which resistance might have developed (Suo et al. 2006).
Live, ionophore tolerant strain based vaccine
Use of ionophore tolerant Eimeria strains has given a new dimension to the anticoccidial vaccine development. An experimental vaccine comprising a strain of E. acervulina and a strain from E. maxima, both of which were partially tolerant to salinomycin, have been developed in USA. Later a live vaccine that can be used with different ionophores has been introduced (Nobilis® COXATM) to the market (Schetters et al. 1999). The vaccine comprises strains of three different Eimeria species, namely E. acervulina, E. tenella, and E. maxima which are relatively tolerant to ionophores. The advantage of these specific vaccines is that they allow the use of ionophores during the first 3–4 weeks when immunity is not complete and the birds are still susceptible. The specific anticoccidial viz. salinomycin limits the increase of infection pressure to the field strains during the period of development of immunity (Vermeulen et al. 2001).
Recombinant vaccine
A major drawback of live vaccines is their limited shelf life and relatively high production costs associated with attenuation. Besides this, it is a herculean task to produce enough material sufficient to vaccinate billions of broilers each year. Therefore, the need for developing a protective recombinant protein based vaccines has been emphasized (Jenkins 2001; Vermeulen 1998). As a matter of fact several target apicomplexan proteins have been expressed and used for either diagnostic or prophylactic purposes (Velmurugan et al. 2008; Tewari et al. 2007, 2010). Microneme proteins are common to apicomplexan parasites and are involved in host cell adhesion and penetration. Several genes coding microneme proteins of E. tenella have been identified and cloned (Ryan et al. 2000). The microneme proteins from E.tenella, Etmic2 and Etmic4 have shown protective effect (Du and Wang 2005) and in ovo vaccination with Etmic2 gene reportedly stimulated intestinal protective immunity against E. tenella and E. acervulina (Ding et al. 2005). Administration of short oligodeoxy nucleotide containing unmethylated CpG motifs (CpG ODNs), which are known to enhance both innate and adaptative immune responses (Krieg 1995), along with the MIC2 antigen in ovo has shown promising results (Dalloul et al. 2005). Enhancement of host immune response was also achieved by co-administration of the chicken interleukin-2 (IL-2) gene (Ding et al.2005). EtMIC1 and EtMIC2 could induce high humoral and CMI response and vaccination along with interleukin (IL)-8, IL-16, transforming growth factor-β4, or lymphotactin resulted in considerably reduction of oocyst output and increased bodyweight gain, following challenge, (Subramanian et al. 2008; Lillehoj et al. 2005).
Several proteins associated with the sexual stage of Eimeria maxima including14, 30, 56, 82 and 230 kDa antigens have been identified as potential vaccine targets for inducing transmission-blocking immunity. Immunization with the 56 and 230 kDa antigens has shown cell mediated immunity against experimental coccidiosis and reduced fecal oocyst shedding (Wallach 1997). Interestingly, the 56 and 230 kDa antigens also induced the production of IgY antibodies which are transferrable to the embryo via egg yolk (Belli et al. 2002) and can provide protective immunity against coccidiosis following challenge infection (Lee et al. 2009). An 82 kDa recombinant protein associated with gametocyte of E. maxima has could induce protective intestinal immunity resulting in decreased oocysts shedding and reduced gut pathology (Jang et al. 2010) associated with increase in the body weights of chickens (Anwar et al. 2008).
Use of viral vector delivery systems currently considered as a promising step forward for producing sustainable cost effective Eimeria vaccines suitable for mass application. Fowl pox and Herpes virus of turkeys are considered lucrative candidates for their ability to harbor the insert size needed for expression of multiple Eimeria genes necessary to control the various species of parasite (Cornenberg et al. 1999; Boyle and Heine 1993).
Factors influencing vaccine efficacy
Methods of vaccination
Even distribution of the vaccine oocysts is crucial for the induction of protective immunity using live vaccines. As a practice the oral vaccines are administered through drinking water or by a variety of feed-based administrations. The limited available data about the use of oral vaccine in broiler flocks indicate that induction of protective immunity is not complete (Williams et al. 1999) which may be ascribed to the fact that success of immunization is dependent upon the chance of the individual chicken to receive a full dose of vaccine. Therefore, it is a common phenomenon that some of the birds receiving more than one dose while others receiving no vaccine at all following application through feed. This situation could be avoided by spraying the vaccine on the birds. Spray-cabinets are widely adapted to vaccinate broilers against coccidiosis (Coccivac-B®). A red coloured dye is incorporated in the vaccine which stains the feathers of approximately 90–95% of the birds following spray. Part of the vaccine sprayed may be taken up through the eyes and part through the beak when the birds arrange their feathers (Chapman et al. 2002). Spray vaccination, therefore, recommended as a more efficient mode of prophylactic approach and it has been observed that over 94% of birds vaccinated by spray had indeed taken up the oocysts (Schetters et al. 1999).
Incorporation of sporozoite antigens of Eimeria tenella into Iscoms could protect broiler chicks following a vaccination through the intra-nasal route. These results suggest that this route of vaccination could also be effective in inducing protection against avian coccidiosis (Garcia et al. 2008).
Combination of ionophores with vaccine
Following proper administration, all available vaccines require a 3–4 weeks time to induce protective immunity in the chickens. Danforth (1998) observed that most broiler breeds exhibited a transient drop in weight gain following vaccination, but recovered quickly and compensated for the loss within 3 or 4 weeks. In another series of field experiments it was shown that vaccinated flocks had higher feed conversion rates than non-vaccinated but medicated flocks. The improvement of feed conversion by 4% in the absence of coccidial challenge has been widely recognized for drugs such as salinomycin or maduramycin. One of the most interesting observations was that the combination of such ionophore treatment with vaccination could further improve the effect of vaccination or treatment alone (Li et al. 2005).
Application of cytokines and ISCOM as adjuvants
Since cytokines are responsible for determining the outcome of immune response, some of them are often considered as potent immunomodulators. Interferon gamma, the important cytokine of Th1 pathway is considered to have adjuvant effect with an anti-coccidial vaccine. Chicken IFNγ has been cloned and expressed (Digby and Lowenthal 1995), and treatment with recombinant IFNγ alone has been shown to help in preventing reductions in body weight gain as well as oocysts output with E. acervulina and E. tenella infections. However, application of cytokines is not free from limitation and the cytokines are subjected to rapid degradation and clearance in vivo. To overcome the problem, the cytokines may be administered in the form of its coding DNA sequence (gene) and may be packaged in a suitable viral vector to facilitate its application in large flocks. Simultaneous inoculation of cDNAs encoding chicken IFN-γ along with 3-1E cDNA of E. acervulina, a stimulator of interferon gamma, could further enhance immunity (Lillehoj et al. 2000). Following inoculation of chickens with a fowl adenovurus chicken IFN-γ construct resulted in less reduction in weight following challenge with E. acervulina.
Immunostimulating complexes (ISCOMs) are unique multimolecular structures formed by encapsulating antigens, lipids and triterpene saponins and are one of the most successful antigen delivery systems for microbial antigens. In a recent communication, significant immunostimulation and protection to challenge have been reported following immunization of chickens with ISCOMs containing purified saponins and native E. tenella antigens. The observation is significant in a way that ISCOMs could be used to develop a safe and effective vaccine because of their ability to stimulate humoral immunity and simultaneously protecting chickens against a challenge with E. tenella infection (Berezin et al. 2010).
Alternative control strategies
Dietary modulators of immunity
A number of natural products or feedstuffs have been tested as anticoccidial dietary additives including mushroom and herb extracts incorporation of which resulted in enhancement of both cellular and humoral immune responses in E. tenella infected chickens (Guo et al. 2004). Poultry feeds containing fish oils, flaxseed oil etc. are rich in n-3 fatty acids, viz. decosahexaenoic acid, linoleic acid, eicosa pentaenoic acid could reduce the lesion caused by E. tenella when incorporated in the starter ration (Allen et al. 1997a, b). The fish oil and flaxseed oil diets could not only significantly reduce the degree of infection but also development of E. tenella and caused ultrastructural degradation of both asexual and sexual stages by inflicting oxidative stress (Danforth et al. 1997).
Dietary plum shown to promote protective immunity against coccidiosis as assessed by reduced body weight loss, decreased oocyst shedding, enhanced splenocyte proliferation, and elevated expression of transcripts encoding IFN-g and IL-15 (Lee et al. 2008).
Artemisinin, a Chinese herb isolated from Artemisia annua works as a naturally occurring endoperoxide with antimalarial properties. It has also been found to be effective in reducing oocyst output from both E. acervulina and E. tenella infections when incorporated in starter diets (Allen et al. 1997a, 1997b). Although the mode of action is not known, it may be presumed that the oxidative stress induced could have the lethal effects on the parasites (Allen and Fetterer 2002).
The commercially available probiotic Mitomax® containing Pedococcus acidilactici and Saccharomyces boulardii, was evaluated as a prophylactic drug for controlling poultry coccidiosis caused by E.acervulina and E.tenella. It has shown to enhance resistance of birds against coccidiosis when incorporated in the broiler diet at ≥0.1% which could be demonstrated by increased coccidian specific humoral immunity and less faecal oocyst production (Lee et al. 2007). Incorporation of immunobiotics, particularly lactic acid bacteria could be useful as immunomodulators to stimulate the gut-associated immune system in neonatal chicks, and thereby protect them from disease without decreasing growth performance as a possible substitution of antibiotics (Sato et al. 2009).
Antioxidants, viz. gamma tocopherol, abaundantly present in wheat, corn and soybeen, as well as curcumin, present in turmeric, could reduce the severity of infection of upper and middle part of small intestine caused by E. acervulina and E. maxima (Allen et al. 1998).
The sugar beet containing betaine, an osmoprotectant, has long been known to have growth promoting effect on livestock. In an experimental tissue culture, betaine and salinomycin could significantly reduce cell invasion by E. acervulina by directly affecting the development of the parasite (Augustine 1997; Augustine and Danforth 1999; Augustine et al. 1997).
A number of nonspecific immunomodulatory agents have been used to enhance immune response against various pathogens in the livestock and poultry industry. A recent study showed that Mycobacterium phlei can be used as an immunotherapeutic agent in augmenting gut mucosal immune response in broiler birds to prevent enteric pathogens during a critical phase in their lifespan (Bera et al. 2010).
Future research
Over the past several decades, plethora of information on the genetic makeup of avian Eimeria spp. and their immunobiological interaction with host has been generated. Increasing reports of development of resistance of the parasites to traditional anticoccidials is one of the important reasons for the impetus given to find new and effective ways for controlling coccidiosis. The success of search of molecules taken place both on the parasites and the host may be attributed to the application of techniques based on molecular biology and which has further driven the search for effective vaccines. The key for successful control of coccidiosis is expected to reveal from a better understanding of parasite biology. The key signal molecules associated with the host and site specificity of avian coccidia need to be identified. Although the host cell invasion process by the parasite is largely understood, the limitations of defense mechanisms that promote the survival of the parasite within the host need to be explored. The genetic basis of survival of the parasite and in that process the key molecules associated need to be identified. The application of the knowledge of bioinformatics may help in the understanding of the complicated process. The regulatory mechanisms involved in the host for expression of key molecules is equally important for better understanding the pathogenesis process as well as may help in devising strategies for their amelirioration.
References
Allen PC, Fetterer RH (2002) Recent advances in biology and immunobiology of Eimeria species and in diagnosis and control of infection with these coccidian parasites of poultry. Clin Microbiol Rev 15(1):58–65
Allen PC, Danforth HD, Levander OA (1997a) Interaction of dietary flaxseed with coccidia infections in chickens. Poult Sci 76:822–827
Allen PC, Lydon J, Danforth HD (1997b) Effects of components of Artemisia annua on coccidia infections in chickens. Poult Sci 76:1156–1163
Allen PC, Danforth HD, Augustine PC (1998) Dietary modulation of avian coccidiosis. Int Parasitol 28:1131–1140
Allocco JJ, Profous-Juchelka H, Myers RW, Nare B, Schmatz DM (1999) Biosynthesis and catabolism of mannitol is developmentally regulated in the protozoan parasite Eimeria tenella. J Parasitol 85:167–173
Anwar MI, Akhtar M, Hussain I, Muhammad F, Haq AU (2008) Effects of local gametocyte and livacox vaccines on live body weight gain and lymphoid organs in chickens. Pak Vet J 28(3):136–138
Augustine PC (1997) Effect of betaine on invasion and development of avian coccidia and growth performance in coccidia-infected chickens. Aust Poult Sci Symp 9:39–45
Augustine PC, Danforth HD (1999) Influence of betaine and salinomycin on intestinal absorption of methionine and glucose and on the ultrastructure of intestinal cells and parasite development stages in chicks infected with Eimeria acervulina. Avian Dis 43:89–97
Augustine PC, McNaughton JL, Virtanen E, Rosi L (1997) Effect of betaine on the growth performance of chicks inoculated with mixed cultures of avian Eimeria species and on invasion and development of Eimeria tenella and Eimeria acervulina in vivo and in vitro. Poult Sci 76:802–809
Bedrnik P (1989) The role of different Eimeria species in a prospective coccidiosis vaccine. In: Yvore P (ed) Coccidian and intestinal coccidiomorphs, INRA. Tours, France, pp 667–670
Belli SI, Lee M, Thebo P, Wallach MG, Schwartsburd B, Smith NC (2002) Biochemical characterisation of the 56 and 82 kDa immunodominant gametocyte antigens from Eimeria maxima. Int J Parasitol 15(32):805–816
Bera AK, Bhattacharya D, Pan D, Manna B, Bandyopadhyay S, Das SK (2010) Effect of heat killed Mycobacterium phlei on body weight gain and management of caecal coccidiosis in broiler chickens. Res Vet Sci 89:196–199
Berezin VE, Bogoyavlenskyi AP, Khudiakova SS, Alexuk PG, Omirtaeva ES, Zaitceva IA, Tustikbaeva GB, Barfield RC, Fetterer RH (2010) Immunostimulatory complexes containing Eimeria tenella antigens and low toxicity plant saponins induce antibody response and provide protection from challenge in broiler chickens. Vet Parasitol 167(1):28–35
Boyle DB, Heine HG (1993) Recombinant fowl pox virus vaccine for poultry. J Immunol cell Biol 71:391–397
Chapman HD (1994) Sensitivity of field isolates of Eimeria to monensin following the use of a coccidiosis vaccine in broiler chickens. Poult Sci 73:476–478
Chapman HD (1998) Evaluation of the efficacy of anticoccidial drugs against Eimeria species in the fowl. Int J Parasitol 28:1141–1144
Chapman HD, Cherry TE, Danforth HD, Richard G, Shirley MW, Williams RB (2002) Sustainable coccidiosis control in poultry production: the role of live vaccines. Int J Parasitol 32:617–629
Cornenberg AM, Van Geffen CE, Dorrestein J, Vermeulen AN, Sondermeijer PJ (1999) Vacination of virus with HVT expressing an Eimeria acervulina antigen improves performance after challenge with Eimeria. Acta Virol 43:192–197
Dalloul RA, Lilleho HS, Klinman DM, Ding X, Min W, Heckert RA, lillehoj EP (2005) In ovo administration of CpG oligodeoxynucleotide and the recombinant microneme protein MIC2 protects against Eimeria infections. Vaccine 23:3108–3113
Danforth HD (1998) Use of live oocyst vaccines in the control of avian coccidiosis: experimental studies and field trials. Int J Parasitol 28:1099–1109
Danforth HD, Allen PC, Levander OA (1997) The effect of high n-3 fatty acid diets on the ultrastructural development of E. tenella. Parasitol Res 83:440–444
Digby MR, Lowenthal JW (1995) Cloning and expression of the chicken interferon-gamma gene. J Interf Cytokine Res 15:939–945
Ding X, Lillehoj HS, Dalloul RA, Min W, Sato T, Yasuda A, Lillehoj EP (2005) In ovo vaccination with the Eimeria tenella EtMIC2 gene induces protective immunity against coccidiosis. Vaccine 23:3733–3740
Du A, Wang S (2005) Efficacy of a DNA vaccine delivered in attenuated Salmonella typhinurium against Eimeria tenella infection in chickens. Int J Parasitol 35:777–785
Ellis J, Bumstead J (1990) Eimeria species: studies using rRNA and rDNA probes. Parasitol 101:1–6
Garcia JL, Guimarães Jda S, Jr Headley SA, Bogado AL, Bugni FM, Ramalho DC, de Souza LM (2008) Eimeria tenella: utilization of a nasal vaccine with sporozoite antigens incorporated into Iscom as protection for broiler breeders against a homologous challenge. Exp Parasitol 120(2):185–190
Guo FC, Kwakkel RP, Williams BA, Parmentier HK, Li WK, Yang ZQ, Verstegen MW (2004) Effects of mushroom and herb polysaccharides on cellular and humoral immune responses of Eimeria tenella infected chickens. Poult Sci 83(7):1124–1132
Jang SI, Lillehoj HS, Lee SH, Lee KW, Park MS, Cha SR, Lillehoj EP, Subramanian BM, Sriraman R, Srinivasan VA (2010) Eimeria maxima recombinant Gam82 gametocyte antigen vaccine protects against coccidiosis and augments humoral and cell-mediated immunity. Vaccine 28:2980–2985
Jeffers TK (1975) Attenuation of Eimeria tenella through selection for pre-cociousness. J Parasitol 61:1083–1090
Jeffers T (1997) Tyzzer to tomorrow: control of avian coccidiosis into the next millenium. In: Shirley MW, Tomley FM, Freeman BM (eds) Control of coccidiosis into the next millennium. Proceedings of 7th international coccidiosis conference, p 16
Jenkins MC (2001) Advances and prospects for subunit vaccine against protozoa of veterinary importance. Vet Parasitol 101:291–310
Jenkins MC, Augustine PC, Danforth HD, Barta JR (1991) X-irradiation of Eimeria tenella oocysts provides direct evidence that sporozoites invasion and early schizont development induce protective immune response(s). Infect Immun 59:4042–4048
Krieg AM (1995) CpG DNA:a pathogenic factor in systemic lupus erythematosus? J Clin Immunol 15:284–292
Lawn AM, Rose ME (1982) Mucosal transport of Eimeria tenella in the cecum of the chicken. J Parasitol 68:1117–1123
Lee EH (1987) Vaccination against coccidiosis in commercial roaster chickens. Can Vet J 28:434–436
Lee S, Lillehoj HS, Park DW, Hong YH, Lin JJ (2007) Effects of Pediococcus- and Saccharomyces-based probiotic (MitoMaxs)® on coccidiosis in broiler chickens. Comp Immunol Microbiol Infect Dis 30:261–268
Lee SH, Lillehoj HS, Lillehoj EP, Cho SM, Park DW, Hong YH, Chun HK, Park HJ (2008) Immunomodulatory properties of dietary plum on coccidiosis. Comp Immunol Microbiol Infect Dis 31:389–402
Lee SH, Lillehoj HS, Park DW, Jang SI, Morales A, García D, Lucio E, Larios R, Victoria G, Marrufo D, Lillehoj EP (2009) Induction of passive immunity in broiler chickens against Eimeria acervulina by hyper immune egg yolk immunoglobulin Y. Poult Sci 88:562–566
Li GQ, Kanu S, Xiao SM, Xiang FY (2005) Responses of chickens vaccinated with a live attenuated multi-valent ionophore-tolerant Eimeria vaccine. Vet Parasitol 129(3–4):179–186
Lillehoj HS, Choi KD, Jenkins MC, Vakharia VN, Song KD, Han JY, Lillehoj EP (2000) A recombinant Eimeria protein inducing interferon gamma production: comparison of different gene expression systems and immunization strategies for vaccination against coccidiosis. Avian Dis 44:379–389
Lillehoj HS, Ding X, Dalloul RA, Sato T, Yasuda A, Lillehoj EP (2005) Embryo vaccination against Eimeria tenella and E. acervulina infections using recombinant proteins and cytokine adjuvants. J Parasitol 91(3):666–673
Long PL (1972) Eimeria tenella: reproduction, pathogenecity and immunogenicity of a strain maintained in chick embryos by serial passage. J Comp Pathol 82:429–437
Morgan JAT, Morris GM, Wlodek BM, Byrnes R, Jenner M, Constantinoi CC, Anderson GR, Lew-Tabor AE, Molloy JB, Gasser RB, Jorgensen WK (2009) Real-time polymerase chain reaction (PCR) assays for the specific detection and quantification of seven Eimeria species that cause coccidiosis in chickens. Mol Cell Probes 23:83–89
Ouarzane M, Labbe M, Pery P (1998) Eimeria tenella:cloning and characterization of cDNA encoding a s3a ribosomal protein. Gene 28:125–130
Procunier JD, Fernando MA, Barta JR (1993) Species and strain differentiation of Eimeria spp. of the domestic fowl using DNA polymorphisms amplified by arbitrary primers. Parasitol Res 79:98–102
Roberts SJ, Smith AL, West AB, Wen L, Findly RC, Owen MJ, Hayday AC (1996) T-cell alpha + and gamma delta + deficient mice display abnormal but distinct phenotypes toward a natural, widespread infection of the intestinal epithelium. Proc Nat Acad Sci USA 93:11774–11779
Ryan R, Shirley M, Tomely F (2000) Mapping and expression of microneme genes in Eimeria tenella. Int J Parasitol 30:1493–1499
Sato K, Takahashi K, Tohno M, Miura Y, Kamada T, Ikegami S, Kitazawa H (2009) Immunomodulation in gut-associated lymphoid tissue of neonatal chicks by immunobiotic diets. Poult Sci 88:2532–2538
Schetters TPM, Janssen HAJM, Vermeulen AN (1999) A new vaccination concept against coccidiosis in poultry. In: van der Sluis W (ed) World poultry. Elsevier, Amsterdam, pp 23–24
Schmatz DM (1997) Anticoccidial drug discovery and design. In: Shirley MW, Tomley FM, Freeman BM (eds) Control of coccidiosis into the next millennium. Proceedings of 7th international coccidiosis conference. Oxford University, England, pp 20–21
Schnitzler BE, Thebo P, Tomley FM, Shirley MW, Uggla A (1997) Identification of Eimeria sp. in poultry by in vitro amplification of the internal transcribed spacer 1. In: Shirley MW, Tomley FM, Freeman BM (eds) Control of coccidiosis into the next millennium. Proceedings of the 7th international coccidiosis conference, p 64
Shirley MW (1975) Enzyme variation in Eimeria species of the chicken. Parasitol 71:369–376
Shirley MW (1994) Coccidial parasites from the chicken: discrimination of different populations of Eimeria tenella by DNA hybridisation. Res Vet Sci 57:10–14
Shirley MW, Harvey DA (1996) Eimeria tenella: genetic recombination of markers for precoccious development and arprinocid resistance. Appl Parasitol 37:293–299
Shirley MW, Millard BJ (1986) Studies on the immunogenicity of seven attenuated lines of Eimeria given as a mixture to chickens. Avian Pathol 15:629–638
Shirley MW, Smith AL, Blake DP (2007) Challenges in the successful control of avian coccidian. Vaccine 25:5540–5547
Subramanian BM, Sriraman R, Rao NH, Raghul J, Thiagarajan D, Srinivasan VA (2008) Cloning, expression and evaluation of the efficacy of a recombinant Eimeria tenella sporozoite antigen in birds. Vaccine 26:3489–3496
Suo X, Zhang JX, Li ZG, Yang CT, Min QR, Xu LT, Liu Q, Zhu XQ (2006) The efficacy and economic benefits of Supercox1, a live anticoccidial vaccine in a commercial trial in broiler chickens in China. Vet Parasitol 142:63–70
Tewari AK, Velmurugan GV, Chandramukhi A, Rao JR, Nagarathna S, Natarajan A (2007) Recombinant SAG1 for detection of Toxoplasma gondii specific antibodies in human patients. In: Proceedings of XVIII national conference of veterinary parasitology, p 166
Tewari AK, Singh H, Sudan V, Rao JR (2010) Recombinant surface antigen 2 (SAG 2) based serodetection of toxoplasmosis in cattle. In: Proceedings of XX national congress of veterinary parasitology, p 42
Trout JM, Lillehoj HS (1993) Transport of Eimeria acervulina sporozoites, evidence of a role for intestinal CD8 T lymphocytes and macrophages. J Parasitol 79:790–792
Velmurugan GV, Tewari AK, Rao JR, Baidya S, Udaya KM, Mishra AK (2008) High level expression of SAG1 and GRA7 gene of Toxoplasma gondii (Izatnagar isolate) and their application in serodiagnosis of goat toxoplasmosis. Vet Parasitol 154:185–192
Vermeulen AN (1998) Progress in recombinant vaccine development against coccidiosis. A review and prospects into the next millennium. Int J Parasitol 28:1121–1130
Vermeulen AN, Schaap DC, Schetters TPM (2001) Control of coccidiosis in chicken by vaccination. Vet Par 100:13–20
Wallach M (1997) The importance of transmission-blocking immunity in the control of infections by apicomplexan parasites. Int J Parasitol 27:1159–1167
Williams RB (1995) Epidemiological studies of coccidiosis in the domestic fowl (Gallus gallus). II. Physical condition and survival of Eimeria acervulina oocysts in poultry house liter. Appl Parasitol 36:90–96
Williams RB (1998) Epidemiological aspects of the use of live anticoccidial vaccines for chickens. Int J Parasitol 28:1089–1098
Williams RB, Carlyle WW, Bond DR, Brown IA (1999) The efficacy and economic benefits of Paracox, a live attenuated anticoccidial vaccine, in commercial trials with standard broiler chickens in the United Kingdom. Int J Parasitol 29(2):341–355
Woods WG, Whithear KG, Richards DG, Anderson GR, Jorgensen WK, Gasser RB (2000) Single-strand restriction fragment length polymorphism analysis of the second internal transcribed spacer (ribosomal DNA) for six species of Eimeria from chickens. Aust Int J Parasitol 30:1019–1023
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Tewari, A.K., Maharana, B.R. Control of poultry coccidiosis: changing trends. J Parasit Dis 35, 10–17 (2011). https://doi.org/10.1007/s12639-011-0034-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12639-011-0034-7