
Abstract
Breast carcinogenesis is a multistep process involving both genetic and epigenetic changes. Epigenetics is defined as reversible changes in gene expression, not accompanied by alteration in gene sequence. DNA methylation, histone modification, and nucleosome remodeling are the major epigenetic changes that are dysregulated in breast cancer. Several genes involved in proliferation, anti-apoptosis, invasion, and metastasis have been shown to undergo epigenetic changes in breast cancer. Because epigenetic changes are potentially reversible processes, much effort has been directed toward understanding this mechanism with the goal of finding effective therapies that target these changes. Both demethylating agents and the histone deacetylase inhibitors (HDACi) are under investigation as single agents or in combination with other systemic therapies in the treatment of breast cancer. In this review, we discuss the role of epigenetic regulation in breast cancer, in particular focusing on the clinical trials using therapies that modulate epigenetic mechanisms.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
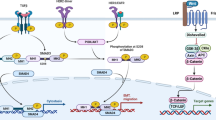
The modern definition of epigenetics is heritable or transient changes in gene expression without a specific change in the DNA sequence [1]. Over the past decade, a tremendous explosion of research in the area of epigenetics and cancer has resulted in a much better understanding of the various mechanisms and effects underlying these non-structural changes to the genome. The exciting aspect of discovering various epigenetic alterations is the potential opportunity to reverse the effects with therapies that can result in gene re-expression. DNA methylation and histone modification resulting in nucleosome remodeling lead to transcriptional silencing of critical tumor-suppressor and growth-regulatory genes in breast cancer. Much of the preclinical work has focused upon using epigenetic therapies to re-express the silenced gene in cell lines. The more challenging aspect has been the clinical application of what we see in the laboratory and clinical effectiveness of such a strategy. For example, preclinical work clearly demonstrates that the maspin gene can be re-expressed in breast cancer cell lines using demethylating agents and histone deacetylase inhibitors (HDACi), but the crucial question is whether even if such a re-expression was achieved in a patient’s tumor, would it have any impact on clinical outcomes [2]. Despite these challenging aspects of the clinical application of epigenetic therapy, advances have been made in exploring and identifying the potential role of such therapies in breast cancer. At present though, all of the epigenetic therapies are in investigational stages in solid tumors and not yet considered standard of care. The areas where we are gaining more knowledge about the potential use of these agents in breast cancer are as combination with targeted therapies, chemotherapy, and radiation to overcome resistance and improve sensitivity to treatment (Fig. 1). Another exciting area in early development is the role of epigenetic modulation in prevention of breast cancer. In this review, we summarize the key epigenetic targets and give an overview of the completed and ongoing preclinical and clinical studies with epigenetic therapies in breast cancer.

Application of epigenetic therapies in breast cancer
Principles of Epigenetics
DNA Methylation
The role of DNA methylation in the epigenetic regulation of human breast cancer has been well characterized. Hypermethylation of CpG islands, which are CpG dinucleotide-rich regions located in the 5′ end of region of genes, commonly occurs in cancer cells [3]. Poorly differentiated breast tumors have more methylated CpG islands than moderately or well-differentiated tumors in breast cancer [4]. Hypermethylated CpG dinucleotides within promoter regions play an important role in regulation of individual genes that promote carcinogenesis. In breast cancer, promoter hypermethylation and subsequent gene silencing occurs in estrogen response genes (ESR1, PR), proapoptotic genes (HOXA5, TMS1), cell cycle–inhibitor genes (p16, RASSF1A), DNA repair genes (BRCA1), and many others [5]. Advances in technologies that assess genome-wide DNA methylation patterns are an important step toward understanding the complex role of this process in malignancy [6].
DNA methylation is mediated by the action of DNA methyltransferases (DNMTs). There are three active DNA methyltransferases: DNMT1, DNMT3a, and DNMT3b. DNMT1 and 3B show increased expression in breast cancer [7]. DNMTs directly interact with histone deacetylases (HDACs) and the methyl-CpG-binding domain (MBD) family of proteins at the promoter regions to form a repressive transcription complex [8]. This complex is important in the conversion of acetylated histones to the deacetylated inactive form. As discussed in subsequent sections, DNA methylation, histone modification, and subsequent nucleosome remodeling are coordinated processes that can be targeted in breast cancer therapeutics [9].
Post-translational Histone Modification
Epigenetic regulation of gene transcription relies on the cooperative mechanisms of DNA methylation and histone acetylation. Histone modification and overall chromatin structure are governed by balanced counteractivity of histone acetyltransferases (HAT) and HDAC. Increased acetylation of histones, which promotes a more open chromatin, is associated with gene activation. In addition, histone methyltransferases (HMTs) mediate histone methylation. Recently, distinct patterns in post-translational histone modification were observed in breast cancer–specific subtypes. Higher global histone acetylation and histone methylation have been observed in the more favorable luminal A subtype, and decreased levels have been seen in the basal subtype [10].
Post-translational histone modifications include the loss of acetylation in lysine 16 and trimethylation of lysine 20 in histone H4 [11]. In addition, two major histone methylation patterns have been identified: 1) methylation of lysine residue 9 on the core histone protein H3, resulting in gene silencing; and 2) methylation of lysine 4 and recently lysine 27 on H3, resulting in activated genes [12]. In addition, the loss of trimethylation of lysine 27 of H3 (H3K27) has been shown to be a negative prognostic indicator in several cancers, including breast cancer [12]. Histone and nucleosome conformational changes are also regulated by the chromatin remodeling complex [13]. These intricate mechanisms that regulate histones are potentially modifiable by epigenetic therapy, including with HDACi therapy.
Known Epigenetic Changes in Breast Cancer
A number of epigenetic changes have been described in breast cancer. Epigenetic changes are not limited to CpG islands and histone post-translational modifications but also include microRNA (miRNAs) [14•]. Aberrant hypermethylation of mir-9-1, mir-124a3, mir-148, mir-152, and mir-663 is present in primary breast tumor specimens [14•]. Additional epigenetic changes in breast cancer include genetic instability with aneuploidy and telomere erosion, which may be associated with epigenetic modifications [15]. The tumor microenvironment may also undergo epigenetic changes that regulate the transition of in situ to invasive breast cancer [16]. Although it is beyond the focus of this review to summarize all of the known epigenetic changes in breast cancer, we can focus on the epigenetic changes in some the key genes involved and their clinical implications.
Epigenetic Regulation of ER
More than two thirds of breast cancers are positive for estrogen receptor (ER) expression at the time of diagnosis and have the potential to benefit from endocrine therapy. However, both de novo and acquired resistance to endocrine therapy pose significant clinical challenges. Response to endocrine therapy is proportional to the degree of ER expression, and loss of ER expression over time is one of the major mechanisms underlying endocrine resistance. A better understanding of the epigenetic regulation of ER presents opportunities for devising rational epigenetic therapies to reverse endocrine resistance to tamoxifen as well as other endocrine therapies, such as aromatase inhibitors. This also provides novel opportunities to treat ER-negative tumors by epigenetic modulation (leading to ER re-expression) and hormonal therapy.
The role of epigenetic changes in the regulation of ER is well established. In brief, the epigenetic regulation of ER is mediated though the recruitment of multimolecular complexes containing HDAC1 and DNMT1 that are recruited to the promoter region as well as other members of the corepressor complex [17]. DNMTl is highly expressed in ER-negative cells throughout the cell cycle. In addition to DNMTs, the interaction of deacetylated histones with the methylated ER gene promoter is a prerequisite for its inactivation. Based on this biologic rationale, there is a synergistic interaction between DNMT inhibitors and HDACi in the induction of ER expression in ER-negative cell lines [18].
Interestingly, HDACi alone can also restore ER expression via chromatin remodeling [19]. Further evidence for the role of histone post-translational modification in ER regulation comes from studies showing that SET7, an HMT that is known to monomethylate H3K4, stabilizes ER, which increases sensitivity to estrogens and enhances transcription of estrogen-response elements [20]. This suggests that post-translational modification of ER may play a role in hormone responsiveness, and hence presents a target for epigenetic therapy with HDACi.
The epigenetic silencing of ER target genes is crucial to the development of ER- independent growth. As a consequence of ER inactivation, the downstream ER targets are silenced by the recruitment of repressor complexes to the promoters of the target genes [21]. A number of preclinical studies have shown that expression of ER can be impacted by epigenetic therapy. Collectively, these data are the basis for the biological rationale for combining hormone therapy with epigenetic-targeted therapies. These trials are discussed in subsequent sections.
Epigenetic Regulation of PR
Similarly to ER, progesterone receptor (PR) expression levels are regulated by epigenetic mechanisms that may impact endocrine therapy responsiveness. A CpG island is located in the first exon of the PR gene, and PR-negative tumors have higher rates of PR promoter hypermethylation [22]. Demethylation of the CpG island is required for the ER binding to this regulatory sequence [23]. In ER-negative tumors, silencing of the ER gene also affects downstream estrogen-regulated gene expression, including PR expression [23]. In vitro, long-term exposure to tamoxifen inhibits PR expression via epigenetic mechanisms, including chromatin remodeling [24].
Epigenetic Regulation of HER2 Neu
Human epidermal growth factor receptor 2 (HER2) neu–overexpressing tumors have higher numbers of genes with methylated CpG islands [25]. In addition, methylation of specific genes, including those for H-cadherin (CDH-13), PR, and 17-B-hydroxysteroid dehydrogenase (important in ER metabolism) correlate with HER2 amplification [26]. Synergistic interaction of the HDACi vorinostat with trastuzumab has been shown in Her2-positive cell lines [27]. Similarly, the HDACi LAQ824 sensitizes human breast cancer cells to trastuzumab via downregulation of HER2 [28].
Biologic Rationale of DNMT Inhibitor Therapy
Preclinical Data with DNMT Inhibitors
The two most commonly investigated DNMT inhibitors are nucleoside analogues 5-azacytidine (5-aza) and 5-aza-2′-deoxycytidine (decitabine). Treatment of ER-negative human breast cancer cell lines with 5-aza promotes the partial demethylation of the ER CpG island, re-expression of ER mRNA, and synthesis of functional ER protein [29]. Treatment with 5-aza results in the increased expression of mir-9-1 [14•]. Inhibition of DNMT1 by antisense, oligo, or siRNA may increase estrogen responsiveness in ER-negative breast cells and increase responsiveness to 5-aza [30]. As described earlier, the addition of HDACi to DNMT inhibitors is synergistic [18].
Clinical Studies with Single-agent DNMT Inhibitors
Currently, no data are available on the clinical activity of 5-aza or decitabine as single agent in breast cancer. A phase I multicenter trial of decitabine in patients with advanced solid tumor malignancies including breast cancer who had failed standard therapy was recently completed (NCT00030615) (http://clinicaltrials.gov). Results of the trial are not yet available. Other nucleoside analogue DNMT inhibitors such as zebularine and 5-fluoro-2′-deoxycitine are in clinical development [31]. An anti-sense oligonucleotide that specifically targets DNMT1 called MG98 is also in clinical development.
Clinical Studies of DNMT Inhibitors with HDACi
A phase I study of the combination of decitabine and HDACi vorinostat given sequentially in advanced solid tumors showed disease stabilization in seven of 22 evaluable patients, including two patients with breast cancer. Dose-limiting toxicities (DLT) include myelosuppression and gastrointestinal toxicities [32•]. A proof-of-principle phase I study was conducted to study the effects of addition of hydralazine and magnesium valproate to neoadjuvant doxorubicin and cyclophosphamide for locally advanced breast cancer [33]. Eligible patients received oral hydralazine and valproate starting day 7 through completion of four cycles of chemotherapy. Core biopsies performed on day 8 of this combination epigenetic therapy showed significant reduction of the global methylation, and HDAC inhibition was observed in peripheral blood mononuclear cells (PBMCs). The combined chemotherapy and epigenetic therapy was generally well tolerated, and increased incidence of drowsiness was attributed to the addition of valproate to the regimen. Although most patients had a clinical response, only one (6.6%) patient had a complete pathologic response. This proof-of-concept study led to a phase II single-arm study of hydralazine and magnesium valproate added to the same chemotherapy on which patients (with advanced solid tumors) were progressing to overcome chemotherapy resistance [34]. Three patients with breast cancer were treated on this study. Two patients who had progressed on paclitaxel were continued on the same along with hydralazine and magnesium valproate and both progressed. Another patient was continued on the same failed hormone therapy along with the combined epigenetic therapies and had stable disease for 4.5 months. Although responses were seen in a few patients with genitourinary tumors, this approach has not been developed further in breast cancer.
Biologic Rationale of HDACi Therapy
Mechanisms of Action of HDACi
HDACi are a class of drugs with broad anti-proliferative and pro-apoptotic effects in a variety of malignant cell types [3]. The only HDACi that is currently approved by the US Food and Drug Administration is vorinostat (suberoylanilide hydroxamic acid) for the treatment of cutaneous T-cell lymphoma. HDACi promote hyperacetylation of histones, which leads to the transcriptional upregulation of pro-apoptotic genes and cyclin-dependent kinase inhibitors such as p21 and p27. In addition, HDACi increase acetylation of the Hsp90 chaperone complex, leading to the destabilization [35]. As a result, proteosome targeting and degradation of multiple client proteins, including HER2, ER, AKT and c-Raf, are increased. Downstream effects of client protein degradation affect a myriad of pathways, including dose-dependent increase of pro-apoptotic proteins Bak and Bim and attenuation of anti-apoptic proteins Bcl-2 and Bcl-xL [36]. In preclinical models, ER is one of the Hsp90 client proteins that are most sensitive to Hsp90 inhibition [37]. By attenuating these downstream signaling pathways, HDACi sensitize breast cancer cells to endocrine, HER2-directed, and cytotoxic therapy.
Preclinical Data with HDACi
Vorinostat is part of the hydroxamic acid family of HDACi and inhibits the proliferation of both ER-positive and ER-negative human breast cancer cell lines [38]. Exposure to vorinostat and another HDACi, LAQ824, downregulates p-Akt, Akt, and c-Raf, sensitizing ER-positive breast cancer cells to tamoxifen [39]. Selective inhibition of HDAC2 by siRNA downregulates both ER and PR expression and potentiates the effects of tamoxifen in ER-positive breast tumor cells [40]. Furthermore, the combination of vorinostat with trastuzumab or docetaxel is synergistic in breast cancer cells with attenuated levels of c-Raf and AKT [36]. Prolonged exposure to vorinostat also sensitizes breast cancer cells to topoisomerase inhibitors [41]. Based on the promising preclinical data, there are multiple clinical trials in breast cancer investigating the effects of vorinostat in combination with endocrine therapy and cytotoxic agents.
Clinical Studies with Single-agent HDACi Therapy
A phase I trial of vorinostat administered intravenously in patients with advanced solid tumor and hematologic malignancies was well tolerated. Post-therapy tumor biopsies showed the accumulation of acetylated histones [42]. No partial responses (PR) or complete responses (CR) were seen, but there was radiographic evidence of tumor regression in two patients with lymphoma and two patients with bladder cancer. A subsequent phase I trial using oral vorinostat was completed showing one (1%) CR and three (4%) PR [43]. Histone acetylation was present in PBMCs, which persisted beyond the half-life of vorinostat. In the phase II study conducted in advanced breast cancer with single-agent oral vorinostat (200 mg twice daily for 14/21 days), 4 of 14 patients (29%) achieved stabilization of disease (SD) (range, 4–14 months) but no patients had objective responses. The most common toxicities included fatigue, nausea, diarrhea, and lymphopenia. Due to the lack of objective response, the study was not continued beyond the first stage [44].
Combination Studies of HDACi and Endocrine Therapy
Based on promising preclinical data of combining HDACi with endocrine therapy, a phase II study of vorinostat in combination with tamoxifen was conducted in metastatic breast cancer patients who had progressed on previous lines of hormone therapy [45•]. Seven patients (21%) had a confirmed PR and one with bone-only disease had an objective response by PET/CT, whereas another four (12%) patients had SD of >6 months in duration. The toxicity profile of the combination was acceptable with pulmonary emboli (5%) in two patients. The median response duration was 8 months. Correlative studies revealed histone H3 and H4 acetylation at day 8, suggesting adequate vorinostat plasma levels in the majority of the patients.
Preliminary results from a phase II study of a benzamide HDACi, entinostat, suggest a potential role for reversal of aromatase inhibitor (AI) endocrine therapy resistance [46]. Eligible postmenopausal women who had progressed on an AI were continued on the same AI they were receiving along with entinostat at 5 mg weekly on a 28-day cycle. One patient had a confirmed PR and one patient had SD > 6 months. Preliminary biomarker analysis indicated that with the addition of HDACi therapy there was increased lysine acetylation and increased apoptosis in PBMCS. A phase II, randomized, double-blind, multicenter study of exemestane with and without entinostat in postmenopausal women with locally recurrent or metastatic ER-positive breast cancer progressing on treatment with a nonsteroidal AI is currently ongoing (http://clinicaltrials.gov).
Combination Studies of HDACi and Chemotherapy
In breast cancer, several trials are ongoing, including multicenter phase I/II trials with the addition of vorinostat to paclitaxel and bevacizumab in metastatic breast cancer [47•]. Interim analysis reported that among 45 evaluable patients, 24 (53%) patients had an objective response and 11 patients had SD, with 5 patients with SD ≥ 24 weeks. The median time to progression was 13.8 months. Two patients had tumor biopsies, and PBMCs collected before and 4 h after the third dose of vorinostat showed increased acetylation of K69 lysine residue of the chaperone protein Hsp 90, upregulation of Hsp70, and downregulation of AKT by Western Blot, providing evidence of Hsp90 inhibition in tumors and PBMCs. Final analysis of an eight-patient cohort with mandatory before and after treatment biopsies is in process. Additional ongoing studies with vorinostat and other cytotoxic agents are noted in Tables 1 and 2.
Other combination studies of HDACi with chemotherapy include a phase I trial of valproic acid followed by epirubicin in advanced, heavily pretreated solid tumors. This combination was well tolerated and had anti-tumor activity [48]. In the dose-expansion part of this study, 15 patients with breast cancer received valproic acid with FEC 100 (5-fluorouracil, epirubicin, and cyclophosphamide). Objective responses were seen in nine of 14 (64%) evaluable patients. Based on promising results of the phase I study, currently a phase II neoadjuvant trial of valproic acid with FEC 100 in locally advanced breast cancer is ongoing (NCT 00437801) (http://clinicaltrials.gov).
Combination Studies of HDACi and HER2-Targeted Therapy
Based on the preclinical rationale of downregulation of HER2 by HDACi, a phase I/II study of vorinostat in combination with trastuzumab in patients with Her2-positive advanced breast cancer resistant to trastuzumab-containing regimens was conducted [49]. Eligible patients were those with measurable disease who had failed prior trastuzumab and could have received prior lapatinib. Sixteen patients were enrolled, of which only 10 were centrally confirmed to have Her2-positive disease (5 were Her2 negative and 1 had insufficient tissue for marker analysis). None of the 11 patients (Her2 positive or unknown) had an objective response. However, one patient with centrally confirmed HER2-negative disease had an objective response. The preliminary data indicate that the addition of HDACi to trastuzumab in heavily pretreated patients was not associated with a response to support a reversal of trastuzumab resistance. As shown in Table 1, additional studies with vorinostat are planned with combination of chemotherapy and trastuzumab and with lapatinib.
A phase I/II multicenter study of pan-HDAC inhibitor, panobinostat, with trastuzumab in metastatic breast cancer patients who have failed trastuzumab is currently ongoing (NCT00567879) and preliminary results have been reported [50]. In the first cohort, two patients with liver metastasis experienced tumor reduction of 29%, and four patients received over eight cycles of the combination. These preliminary results demonstrate that the combination of panobinostat and trastuzumab shows promising activity. Another study with the addition of paclitaxel to this regimen is currently ongoing (NCT00788931) (http://clinicaltrials.gov). Preliminary results from a phase I study of panobinostat with lapatinib with or without capecitabine were recently presented showing tolerability and promising activity in the combination regimen [51]. Additional ongoing studies with HDACi and HER2-targeted therapies are summarized in Tables 2 and 3.
Epigenetic Therapy as a Chemopreventive Strategy
Sulforaphane is an isothiocyanate present in abundance in broccoli sprouts and has been shown to exert anti-cancer effects through histone acetylation, induction of P21 and Bax, and induction of cell cycle arrest and apoptosis. Ingestion of 1 cup of broccoli inhibited HDAC activity in PBMCs 3–6 h after consumption, resulting in induction of H3 and H4 acetylation, providing rationale to use this dietary ingredient as a chemopreventive agent. Based on this, a phase II randomized, placebo-controlled, double-blind trial of broccoli sprout extract in women with newly diagnosed ductal carcinoma in situ and/or atypical ductal hyperplasia is currently ongoing (NCT00843167) (http://clinicaltrials.gov). Results from this study may establish a role for epigenetic therapy in prevention of breast cancer.
Epigenetic Prognostic and Predictive Factors in Therapy
Epigenetic changes have been investigated as both prognostic and predictive factors in breast cancer. A number of epigenetic changes have been investigated as prognostic factors. For example, DNA hypermethylation of PITX2 is a marker of poor prognosis in node-negative, hormone receptor–positive breast cancer after adjuvant tamoxifen therapy [52]. Promoter methylation of the NEUROD1, a member of the stem cell, polycom group oftarget genes, has been investigated as a marker for chemosensitivity in the neoadjuvant setting [53]. Most recently, investigators from Europe reported on the first large-scale epigenotyping study of the peripheral blood DNA and demonstrated that DNA methylation provides a good prediction of breast cancer risk that is not completely independent of traditional risk factors for breast cancer [54•].
A variety of methods have been used by different studies to evaluate methylation patterns, including methyl-specific polymerase chain reaction, MethyLight, and bisulfite pyrosequencing with lack of standardization across groups, which has made it challenging to compare results across studies. With advances in high-throughput technology for assessment of genome-wide methylation patterns such the MALDI-TOF mass array, additional hypermethylated CpG islands have been identified in breast cancer [55•].
The challenge remains as to how to best define optimal biological endpoints with epigenetic therapy. Because the epigenetic changes induced by DNMTs and HDACi are transient and reversible, it particularly important to have tissue-specific measures of whether epigenetic targets are effectively impacted by therapy. As discussed in the previous section, several trials have utilized PBMCs for evaluation of H3 and H4 acetylation levels as well as the AKT pathway. Pretreatment and post-treatment tissue biopsies remain the best way to assess pharmacodynamics’ epigenetic effects. The phase I/II study with addition of vorinostat to paclitaxel and bevacizumab has an expanded cohort of eight patients with required tumor biopsies before and after treatment. Results are pending from this portion of the study. However, serial biopsies are not always readily possible in many women with breast cancer due to inaccessibility of tumor sites and patient preference [47•]. Furthermore, even if we do establish pharmacodynamic endpoints that can be easily assessed, no studies have established the clinical relevance of meeting these endpoints. The search is also ongoing for validated predictive biomarkers that can help to identify tumors that are particularly sensitive to epigenetic modulation.
Conclusions
Targeting epigenetic changes in breast cancer is an exciting and evolving arena that has the potential to change our current standard-of-care therapies. In this review, we have summarized the complex epigenetic regulatory pathways that can impact breast cancer prognostication and therapeutics. Particularly promising is the combination of epigenetic therapy with our existing cytotoxic and endocrine therapies in breast cancer. However, many uncertainties still remain as to how to best translate these findings in the clinical arena. Given that the epigenetic changes induced by DNMTs and HDACi are transient and reversible, a number of studies are ongoing to help define the optimal doses and treatment schedules for these agents.
References
Papers of particular interest, published recently, have been highlighted as: • Of importance
Bird A: Perceptions of epigenetics. Nature 2007, 447(7143):396–398.
Maass N, Biallek M, Rosel F, Schem C, Ohike N, Zhang M, Jonat W, Nagasaki K: Hypermethylation and histone deacetylation lead to silencing of the maspin gene in human breast cancer. Biochem Biophys Res Commun 2002, 297(1):125–128.
Stearns V ZQ, Davison NE.: Epigenetic Regulation as a New Target for Breast Cancer Therapy. Cancer Investigation 2007, 25:659–665.
Yan P PM, Laux D, Asare A, Caldwell C, Huang T: CpG Island Arrays: An application toward Deciphering epigenetic signatures of breast cancer. Clinical Cancer Research 2000, 6:1432–1438.
Portela A, Esteller M: Epigenetic modifications and human disease. Nat Biotechnol, 28(10):1057-1068.
Esteller M: Cancer epigenomics: DNA methylomes and histone-modification maps. Nat Rev Genet 2007, 8(4):286–298.
Giacinti L, Claudio PP, Lopez M, Giordano A: Epigenetic information and estrogen receptor alpha expression in breast cancer. Oncologist 2006, 11(1):1–8.
Lopez-Serra L, Ballestar E, Ropero S, Setien F, Billard LM, Fraga MF, Lopez-Nieva P, Alaminos M, Guerrero D, Dante R et al.: Unmasking of epigenetically silenced candidate tumor suppressor genes by removal of methyl-CpG-binding domain proteins. Oncogene 2008.
Esteller m: Epigenetics in Cancer. The New England Journal of Medicine 2008, 358:1148–1159.
Elsheikh SE, Green AR, Rakha EA, Powe DG, Ahmed RA, Collins HM, Soria D, Garibaldi JM, Paish CE, Ammar AA et al.: Global histone modifications in breast cancer correlate with tumor phenotypes, prognostic factors, and patient outcome. Cancer Res 2009, 69(9):3802–3809.
Fraga MF, Ballestar E, Villar-Garea A, Boix-Chornet M, Espada J, Schotta G, Bonaldi T, Haydon C, Ropero S, Petrie K et al.: Loss of acetylation at Lys16 and trimethylation at Lys20 of histone H4 is a common hallmark of human cancer. Nat Genet 2005, 37(4):391–400.
Wei Y, Xia W, Zhang Z, Liu J, Wang H, Adsay NV, Albarracin C, Yu D, Abbruzzese JL, Mills GB et al.: Loss of trimethylation at lysine 27 of histone H3 is a predictor of poor outcome in breast, ovarian, and pancreatic cancers. Mol Carcinog 2008, 47(9):701–706.
Harikrishnan KN, Chow MZ, Baker EK, Pal S, Bassal S, Brasacchio D, Wang L, Craig JM, Jones PL, Sif S et al.: Brahma links the SWI/SNF chromatin-remodeling complex with MeCP2-dependent transcriptional silencing. Nat Genet 2005, 37(3):254–264.
• Lehmann U, Hasemeier B, Christgen M, Muller M, Romermann D, Langer F, Kreipe H: Epigenetic inactivation of microRNA gene hsa-mir-9-1 in human breast cancer. J Pathol 2008, 214(1):17–24. This is a study of epigenetic regulation of miRs in breast cancer.
Matzke MA, Mette MF, Kanno T, Matzke AJ: Does the intrinsic instability of aneuploid genomes have a causal role in cancer? Trends Genet 2003, 19(5):253–256.
Polyak K, Haviv I, Campbell IG: Co-evolution of tumor cells and their microenvironment. Trends Genet 2009, 25(1):30–38.
Peterson TJ, Karmakar S, Pace MC, Gao T, Smith CL: The silencing mediator of retinoic acid and thyroid hormone receptor (SMRT) corepressor is required for full estrogen receptor alpha transcriptional activity. Mol Cell Biol 2007, 27(17):5933–5948.
Yang X, Phillps DL, Fergusson AT et al.: Synergistic activation of functional estrogen receptor (ER)-alpha by DNA methyltransferase and histone deacetylase inhibition in human ER-alpha-negative breast cancer cells. Cancer Res 2001, 16:7025–7029.
Zhou Q, Atadja P, Davidson NE: Histone deacetylase inhibitor LBH589 reactivates silenced estrogen receptor alpha (ER) gene expression without loss of DNA hypermethylation. Cancer Biol Ther 2007, 6(1):64–69.
Subramanian K, Jia D, Kapoor-Vazirani P, Powell DR, Collins RE, Sharma D, Peng J, Cheng X, Vertino PM: Regulation of estrogen receptor alpha by the SET7 lysine methyltransferase. Mol Cell 2008, 30(3):336–347.
Leu YW, Yan PS, Fan M, Jin VX, Liu JC, Curran EM, Welshons WV, Wei SH, Davuluri RV, Plass C et al.: Loss of estrogen receptor signaling triggers epigenetic silencing of downstream targets in breast cancer. Cancer Res 2004, 64(22):8184–8192.
Lapidus RG, Ferguson AT, Ottaviano YL, Parl FF, Smith HS, Weitzman SA, Baylin SB, Issa JP, Davidson NE: Methylation of estrogen and progesterone receptor gene 5′ CpG islands correlates with lack of estrogen and progesterone receptor gene expression in breast tumors. Clin Cancer Res 1996, 2(5):805–810.
Fleury L, Gerus M, Lavigne AC, Richard-Foy H, Bystricky K: Eliminating epigenetic barriers induces transient hormone-regulated gene expression in estrogen receptor negative breast cancer cells. Oncogene 2008.
Badia E, Duchesne MJ, Semlali A, Fuentes M, Giamarchi C, Richard-Foy H, Nicolas JC, Pons M: Long-term hydroxytamoxifen treatment of an MCF-7-derived breast cancer cell line irreversibly inhibits the expression of estrogenic genes through chromatin remodeling. Cancer Res 2000, 60(15):4130–4138.
Terada K, Okochi-Takada E, Akashi-Tanaka S, Miyamoto K, Taniyama K, Tsuda H, Asada K, Kaminishi M, Ushijima T: Association between frequent CpG island methylation and HER2 amplification in human breast cancers. Carcinogenesis 2009, 30(3):466–471.
Fiegl H, Millinger S, Goebel G, Muller-Holzner E, Marth C, Laird PW, Widschwendter M: Breast cancer DNA methylation profiles in cancer cells and tumor stroma: association with HER-2/neu status in primary breast cancer. Cancer Res 2006, 66(1):29–33.
Bali P PM, Swaby R, Fiskus W, Yamaguchi H, Balasis M, Rocha K, Wang H, Richon V, Bhalla K: Activity of Suberoylanilide Hydroxamic acid against human breast cancer cells with amplificatoin of Her 2. Clinical Cancer Research 2005, 11(17):6382–6389.
Fuino L BP, Wittman S et al.: Histone deacetylse inhibitor LAQ824 down regulated Her-2 and sensitizes human breast cancer cells to trastuzumab, taxotere, gemcitabine, and epothilone. B Mol Cancer Ther 2003, 2:971–984.
Yang X FA, Nass SJ: Transcriptional activation of estrogen receptor alpha in human breast cancer cells by histone decetylase inhibition. Cancer Res 2000, 60:6890–6894.
Robert MF, Morin S, Beaulieu N et al.: DNMT1 is requiered to maintain CpG methylation and aberrant gene silencing in human cancer cells. Nat Genet 2003, 33:61–65.
Cheng JC, Yoo CB, Weisenberger DJ, Chuang J, Wozniak C, Liang G, Marquez VE, Greer S, Orntoft TF, Thykjaer T et al.: Preferential response of cancer cells to zebularine. Cancer Cell 2004, 6(2):151–158.
• Stathis A, Hottie S, Hirte, H et al.: Phase I study of intravenous decitabine in combination with oral vorinostat in patients with advanced solid tumors and non-Hodgkin’s lymphomas (NHL). J Clin Oncol 2009, 27:15s, (suppl; abstr 3528). This is a phase I study of DNMT inhibitor in combination with HDACi in solid tumors.
Arce C, Perez-Plasencia C, Gonzalez-Fierro A, de la Cruz-Hernandez E, Revilla-Vazquez A, Chavez-Blanco A, Trejo-Becerril C, Perez-Cardenas E, Taja-Chayeb L, Bargallo E et al.: A proof-of-principle study of epigenetic therapy added to neoadjuvant doxorubicin cyclophosphamide for locally advanced breast cancer. PLoS ONE [Electronic Resource] 2006, 1:e98.
Candelaria M, Gallardo-Rincon D, Arce C, Cetina L, Aguilar-Ponce JL, Arrieta O, Gonzalez-Fierro A, Chavez-Blanco A, de la Cruz-Hernandez E, Camargo MF et al.: A phase II study of epigenetic therapy with hydralazine and magnesium valproate to overcome chemotherapy resistance in refractory solid tumors. Annals of Oncology 2007, 18(9):1529–1538.
Isaacs JS, Xu W, Neckers L: Heat shock protein 90 as a molecular target for cancer therapeutics. Cancer Cell 2003, 3(3):213–217.
Bali P, Pranpat M, Swaby R, Fiskus W, Yamaguchi H, Balasis M, Rocha K, Wang HG, Richon V, Bhalla K: Activity of suberoylanilide hydroxamic Acid against human breast cancer cells with amplification of her-2. Clinical Cancer Research 2005, 11(17):6382–6389.
Thomas S, Munster PN: Histone deacetylase inhibitor induced modulation of anti-estrogen therapy. Cancer Lett 2009, 280(2):184–191.
Munster PN, Troso-Sandoval T, Rosen N, Rifkind R, Marks PA, Richon VM: The histone deacetylase inhibitor suberoylanilide hydroxamic acid induces differentiation of human breast cancer cells. Cancer Res 2001, 61(23):8492–8497.
Fiskus W, Ren Y, Mohapatra A, Bali P, Mandawat A, Rao R, Herger B, Yang Y, Atadja P, Wu J et al.: Hydroxamic acid analogue histone deacetylase inhibitors attenuate estrogen receptor-alpha levels and transcriptional activity: a result of hyperacetylation and inhibition of chaperone function of heat shock protein 90. Clin Cancer Res 2007, 13(16):4882–4890.
Bicaku E, Marchion DC, Schmitt ML, Munster PN: Selective inhibition of histone deacetylase 2 silences progesterone receptor-mediated signaling. Cancer Res 2008, 68(5):1513–1519.
Marchion DC, Bicaku E, Daud AI, Richon V, Sullivan DM, Munster PN: Sequence-specific potentiation of topoisomerase II inhibitors by the histone deacetylase inhibitor suberoylanilide hydroxamic acid. J Cell Biochem 2004, 92(2):223–237.
Kelly WK, Richon VM; O’Connor O, et al.: Phase I clinical trial of histone deacetylase inhibitor suberoylanilide hydroxamic acid administered intravenously. Clin Cancer Res 2003, 9:3578–3588.
Kelly WK, O’Connor OA, Krug LM. et al.: Phase I study of an oral histone deacetylase inhibitor, suberoylanilide hydroxamic acid in patients with advanced cancer. J Clin Oncol 2005, 23:3923–3931.
Luu TH, Morgan RJ, Leong L, et al.: A phase II trial of vorinostat (suberoylanilide hydroxamic acid) in metastatic breast cancer: a California Cancer Consortium Study. Clinical Cancer Research 2008, 14:7138–7142.
• Munster PN, Lacevic M, Thomas S , Christian C, Ismail-Khan R, Melisko M, Rugo HE and Minton SE. Phase II trial of the histone deacetylase inhibitor, vorinostat, to restore hormone sensitivity to the antiestrogen tamoxifen in patients with advanced breast cancer who progressed on prior hormone therapy. J Clin Oncol 2009;Vol 27, No 15S (May 20 Supplement): 1075. This is a proof-of-principle study showing the effects of vorinostat in reverting tamoxifen resistance.
A. M. Wardley RS, J. McCaffrey, J. Crown, Z. Malik, D. Rea, P. J. Barrett-Lee, G. T. Lee: Phase II data for entinostat, a class 1 selective histone deacetylase inhibitor, in patients whose breast cancer is progressing on aromatase inhibitor therapy J Clin Oncol 2010, J Clin Oncol 28:15s, (suppl; abstr 1052).
• Ramaswamy B BK, Cohen B, et al.: Phase II trial of the histone deacetylase inhibitor (HDACi) vorinostat plus paclitaxel and bevacizumab in metastatic breast cancer (MBC): New York Cancer Consortium Trial 7703. AACR 100th Annual Meeting April 18–22 2009, 09-AB4116-AACR Denver, CO. This is an important study demonstrating the benefits of combining chemotherapy and vorinostat in breast cancer.
Munster P, Marchion D, Bicaku E, Schmitt M, Lee JH, DeConti R, Simon G, Fishman M, Minton S, Garrett C et al.: Phase I trial of histone deacetylase inhibition by valproic acid followed by the topoisomerase II inhibitor epirubicin in advanced solid tumors: a clinical and translational study. J Clin Oncol 2007, 25(15):1979–1985.
R. Swaby MW, J. Sparano, K. Bhalla, N. Meropol, C. Falkson, C. Pellegrino, P. Klein, L. Goldstein and G. Sledge, Jr: A Phase II Study of the Histone Deacetylase Inhibitor, Vorinostat, in Combination with Trastuzumab in Patients with Advanced Metastatic and/or Local Chest Wall Recurrent HER-2 Amplified Breast Cancer Resistant to Transtuzumab-Containing Therapy: (E1104) a Trial of the Eastern Cooperative Oncology Group. Cancer Res 2009, 69(24 Suppl):Abstract nr 5084.
Conte P, Campone, M, Pronzato, P, D. Amadori, R. Frank, F. Schuetz, D. Rea, A. Wardley, C. Britten and A. Elias Phase I trial of panobinostat (LBH589) in combination with trastuzumab in pretreated HER2-positive metastatic breast cancer (mBC): Preliminary safety and tolerability results. Journal of Clinical Oncology, 2009 ASCO Annual Meeting Proceedings (Post-Meeting Edition)2009, Vol 27, No 15S (May 20 Supplement), 2009: 1081
N. W. Peacock, SF Jones, D. A. Yardley, J. C. Bendell, J. R. Infante, P. B. Murphy, H. A. Burris III: A phase I study of panobinostat (LBH589) with capecitabine with or without lapatinib. J Clin Oncol 2010, 28:15s, (suppl; abstr 1115)
Nimmrich I, Sieuwerts AM, Meijer-van Gelder ME, Schwope I, Bolt-de Vries J, Harbeck N, Koenig T, Hartmann O, Kluth A, Dietrich D et al.: DNA hypermethylation of PITX2 is a marker of poor prognosis in untreated lymph node-negative hormone receptor-positive breast cancer patients. Breast Cancer Res Treat 2007.
Fiegl H, Jones A, Hauser-Kronberger C, Hutarew G, Reitsamer R, Jones RL, Dowsett M, Mueller-Holzner E, Windbichler G, Daxenbichler G et al.: Methylated NEUROD1 promoter is a marker for chemosensitivity in breast cancer. Clin Cancer Res 2008, 14(11):3494–3502.
• Widschwendter M, Apostolidou S, Raum E, Rothenbacher D, Fiegl H, Menon U, Stegmaier C, Jacobs IJ, Brenner H: Epigenotyping in peripheral blood cell DNA and breast cancer risk: a proof of principle study. PLoS ONE 2008, 3(7):e2656. This article presents the first blood-based study of methylation profiles in breast cancer.
• Radpour R, Kohler C, Haghighi MM, Fan AX, Holzgreve W, Zhong XY: Methylation profiles of 22 candidate genes in breast cancer using high-throughput MALDI-TOF mass array. Oncogene 2009, 28(33):2969–2978. This article reviews a new method for genome-wide methylation studies.
Disclosure
Maryam B. Lustberg reports no potential conflict of interest relevant to this article. Bhuvaneswari Ramaswamy reports no potential conflict of interest relevant to this article.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lustberg, M.B., Ramaswamy, B. Epigenetic Therapy in Breast Cancer. Curr Breast Cancer Rep 3, 34–43 (2011). https://doi.org/10.1007/s12609-010-0034-0
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12609-010-0034-0