
Abstract
In this study, the effects of orally administrated two native probiotics (Lactobacillus plantarum and Lactobacillus delbrueckii ssp. bulguricus), isolated from the intestine of Shabot fish, Tor grypus, on some immune response parameters and immune-related genes expression against Aeromonas hydrophila in T. grypus were evaluated. Four hundred and eighty juveniles weighing 45 ± 10 g were randomly divided into four groups (with three replications) and fed with the experimental diet containing 5 × 107 cfu g−1 of L. plantarum (G1), Lactobacillus bulgaricus (G2), Lactobacillus casei (G3), and a control diet (without probiotics) for 60 continuous days. At the end of the dietary treatments, fish were challenged with a lethal concentration of A. hydrophila (5 × 108 CFU ml−1) via intra peritoneal (i.p) injection. Blood and head kidney samples were taken from six fish in each treatment before challenging and 6, 12, 24, and 48 h and also 7 days after injection. The results showed that lysozyme, complement, bactericidal, and NBT activity of probiotic-treated groups were significantly elevated (P < 0.05). The IL-8, IL-1β, and TNF-α gene expressions were significantly higher in all probiotic-treated groups (P < 0.05). Meanwhile, a high direct correlation was observed between serum immune parameters and expression of immune-related genes (P < 0.0001); furthermore, the highest correlation (R 2 = 0.634, P < 0.0001) was recorded between IL-1β expression and NBT activity. It can be concluded that not only two native probiotics strains stimulate serum immune responses parameters and immune-related gene expression in T. grypus, but also a high correlation was seen among these indices. The study suggests that gastrointestinal colonization is preferred for host specificity as the strain previously derived from shabot fish displayed better colonization than the non-indigenous bacteria strain such as L. casei. Therefore, these native probiotics bacteria can be accounted as suitable candidates to immune stimulation in fish.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In traditional aquaculture systems, bacterial disease outbreaks are typically treated with antibiotics; however, the intensive use of antibiotics is being discouraged due to the emergence of antibiotic resistance. Therefore, it is very essential to find new methods for preventing infectious diseases. In addition, there is a growing interest in the beneficial use of microorganisms to prevent or control pathogenic microorganisms as an alternative to traditional treatments [5, 18]. Probiotics are microorganisms that beneficially affect the host by selectively stimulating beneficial gastrointestinal microbial communities. The use of probiotics have also been suggested to be an alternative method to reduce pathogenic organisms in the gastrointestinal tract of fish due to their antagonistic activity in colonization site on the host’s intestine, resulting in prevention and control of diseases [4, 28]. The use of probiotics in aquaculture is rather new [39]. Various microorganisms have been evaluated as probiotics in aquatic animals [6, 7, 27]. In an autochthonous approach, many putative probiotic strains have been isolated from their immediate aquatic environment in which fish exist; the historical safety status of these isolated microbial strains may not be similar and the historical safety status of the common counterparts likewise [1]. However, lactic acid bacteria (LABs) are the most commonly used probiotics in aquaculture [2, 36, 59, 61].
Discovery of a vast number of immune relevant genes during the recent years facilitated the study of many immune processes in more detail. Hence, studies on the expression of immune-related genes of teleost, infected with various pathogens, are increasing [12]. Remarkable progress has been achieved in isolating and characterizing cytokine genes of fish in recent years [51, 55]. In addition, the identification and characterization of various immune-regulatory genes have promoted the study of gene expression during disease processes. There are a large number of studies reported the expression of immune-regulatory genes in fish infected with bacterial pathogens [8, 40, 43, 45, 47].
Shabot fish (Tor grypus) is one of the most important fish species in southwest Asian countries (i.e., Iran, Iraq, Turkey, and Syria) since it has excellent biological characteristics such as fast growth rate and high resistance against natural stressors, good marketing, and high economic value. These species, which have been artificially propagated, were introduced to cyprinid farms as a new species in cyprinid poly-culture systems during the last decade. The lack of knowledge is a major obstacle to the establishment of effective preventive measures against a wide range of infectious agents. With intensification of aquaculture production, problems with opportunistic pathogens, such as Aeromonas hydrophila, are increasing. This pathogen is an important common freshwater pathogen of fish in temperate and tropical regions, leading more disease control programs should be developed to reduce economic losses following this kind of disease outbreaks [33]. However, there is no information available on immune responses of T. grypus against A. hydrophila infection, and it is not clear how systemic immunity is modulated by probiotics after bacterial infection. Because of a lack of information on using Lactobacillus probiotic in Barbus species fish and few reports on immune response of Barbus fish, this study has focused on the effects of two lactic acid bacteria (LABs), Lactobacillus plantarum and Lactobacillus delbrueckii subsp. bulgaricus, isolated from the intestine of T. grypus and a standard Lactobacillus strain (Lactobacillus casei ATTC1608®) on some non-specific immune parameters and immune-related gene expression as well as their correlation in juvenile T. grypus infected with A. hydrophila.
Materials and Methods
Bacterial Strains
Lactobacillus plantarum subsp. Plantarum and Lactobacillus delbrukei subsp. Bulgaricus were used in food supplementation. These strains were chosen from over 30 LAB obtained from the intestine of healthy wild T. grypus, according to their high in vitro probiotic characteristics [33]. These strains were primarily identified based on colony and cell morphology, gram staining, biochemical characteristics, and 16S rRNA gene sequencing [3, 33]. The positive control strain (L. casei, PTCC 1608) was obtained from Pasteur Institute, Tehran, Iran. They were cultured in the DeMan Regosa and Sharpe (MRS) broth (Pronadisa, Madrid, Spain) at 30 °C. Bacterial strains were preserved in skim milk at − 80 °C until used. The fish pathogens A. hydrophila strain ATCC AH04 was obtained from marine laboratory, Institute of Aquaculture, University of Stirling, Scotland.
Preparation of the Experimental Feed
Three Lactobacillus bacterial strains were grown after 48 h in MRS broth in a shaking incubator at 25 °C. After incubation, the cells were harvested by 10 min centrifugation (2000g), then bacteria were washed twice with phosphate-buffered saline (PBS, 0.1 M, pH = 7.2) and re-suspended in the same bacterial solution. The concentration of bacteria was adjusted to 3 × 109 CFU ml−1 using a spectrophotometer. The proximate analysis of the basal diet according to the AOAC method was 37.1% crude protein, 8.8% crude lipid, 9.6% ash, and 390 Kcal 100 g−1 gross energy. The probiotic-enriched diets were prepared by gently spraying the required amount of bacterial suspension on the control diet (16 ml bacterial suspension per kilogram diet) and mixing, bit by bit, in a drum mixer to obtain a final probiotic concentration of 5 × 107 CFU g−1. They were packed in sterile propylene containers and stored at 4 °C for viability studies. This dose was selected based on a previously recommended probiotic concentration in food [31, 33].
Fish and Experimental Design
T. grypus fish weighing 45 ± 10 g (mean ± SD) were obtained from a commercial fish farm, Ahvaz, Khouzestan province, Iran, and then transferred to the fish room of Faculty of Veterinary Medicine, Shahid Chamran University of Ahvaz. The fish were acclimatized for 2 weeks in indoor 300 L fiberglass tanks and were fed with a standard diet. All fish were then randomly divided into four treatments, including control, L. plantarum (G1), Lactobacillus bulgaricus (G2), and L. casei PTTC1608 (G3, as a positive control) with three aquaria as replication for each treatment (totally 12 aquaria, including 40 fish/aquarium). The final concentration of each probiotics was about 5 × 107 CFU g −1 of the diet. The aquaria were supplied with filtered water at a temperature of 26 ± 1 °C. The fish were fed with probiotic diets for 60 days (twice a day). Daily feeding rate was about 2% of body weight. The study was carried out in accordance with EC Directive 86/609/EEC for animal experiments.
Resistance to A. hydrophila Infection
At the end of the experiment, on day 60, fish in each group were challenged with A. hydrophila (strain AH04). AH04 was grown for 24 h in the tryptic soy broth (TSB, FlukaBiochemika) at 37 °C in a shaking incubator at 200 rpm. Bacteria were washed twice with PBS and re-suspended in the same buffer. The concentration of bacteria was adjusted to bacterial LD50 (3.7 × 108 CFU ml−1) using a spectrophotometer and the plate counting method. The concentration of the bacterial suspension was determined using a bacterial counting chamber. The fish were anesthetized with eugenol (1:10,000) (Shanghai Reagent, China) before injection. The concentration of 3.7 × 108 CFU ml−1 live A. hydrophila was injected into fish (LD50 resulted in previous study). All fish in each group were intraperitoneally injected with 0.2 ml of A. hydrophila suspensions using 1 ml sterile syringe. The control group was divided in two subgroups, one injected with 0.2 ml of A. hydrophila suspensions and another group injected intraperitoneally with 0.2 ml PBS. Mortalities were recorded every day during 2-week post-challenge, and all of the dead T. grypus were examined bacteriologically to ensure the presence of the pathogen [43].
Sample Collection
On day 60 of the experiment, six fish from each group were bled through the caudal vein after anesthesia with Eugenol (100 ppm) in six sampling intervals: before challenge and also 6, 12, 24, 48 h, and 7 days after challenge of A. hydrophila. An aliquot of the blood was heparinized (50 IU ml−1) and the remaining part was used for collecting serum. The collected serum samples were stored at − 80 °C until further analysis for various immune parameters.
Head kidneys from thoroughly bled fish (six samples from each group in each sampling time point) were aseptically dissected and immediately stored in cold PBS, pH =7.2, and stored in 1 ml Trizol at − 80 °C.
RNA Isolation and cDNA Synthesis
Total RNA was isolated from tissues using the TriPure isolation reagent according to the manufacturer’s procedure (Roche, Canada). The concentration of extracted RNA was calculated at a wavelength of 260 nm using nanodrop spectrophotometry (Eppendorf, Germany). To detect the purity of RNA, the optical density (OD) absorption ratio at 260/280 nm was determined and samples having a ratio more than 1.8 were used for the cDNA synthesis. Possible DNA contamination was removed by the treatment of RNA (1 μg) with DNase I (2 U μl−1) for 1 h at 37 °C (Vivantis, Malaysia). Reverse transcription was carried out with the Rocket Script RT PreMix Kit using 1 μg of RNA and oligo dT based on the manufacturer’s protocol (Bioneer Corporation, South Korea).
Real-Time Quantitative RT-PCR
To evaluate the expression levels of IL1-β, IL-8, and TNF-α mRNA in head kidneys, real-time PCR was performed using qPCRTM Green Master Kit for SYBR Green I® (Jena Biosciense, Germany) on a Lightcycler® Detection System (Roche, USA). Relative expression levels of the all transcripts were compared to β-actin as a housekeeping gene. Specific sets of primers (Bioneer, South Korea) were designed based on Cyprinus carpio (Table 1). Reactions were performed in a 12.5 μl mixture containing 6.25 μl qPCRTM Green Master Mix (2X), 0.25 μl of each primer (10 μM), 3 μl (100 ng) cDNA, and 2.75 μl nuclease-free water. The PCR protocol consisted of a 5-min denaturation at 94 °C followed by 45 cycles of 94 °C for 15 s and 60 °C for 30 s. Reactions were performed in triplicate. Two separate reactions without cDNA or with RNA were performed as control groups in parallel with experimental groups. According to the comparative 2-ΔΔCt method, the relative quantification was performed using Lightcycler 96® software. Validation of assay to check that the primers for the chβ-actin and chCASQ2 had similar amplification efficiencies was carried out as described previously [44]. All qPCR analysis was performed according to the Minimum Information for Publication of Quantitative Real-Time PCR Experiments (MIQE) guideline [11].
Non-specific Immune Responses
A turbidometric assay using lyophilized Micrococcus lysodeikticus (Sigma–Aldrich) was employed to determine lysozyme activity in serum [57, 33]. One hundred thirty-five milliliters of M. lysodeikticus at a concentration of 0.2 mg ml−1 (w/v) in 0.02 M sodium phosphate buffer (SPB) (pH = 5.8) (Sigma–Aldrich) was added to 15 μl of serum sample. As a negative control, SPB was replaced instead of serum. Results were expressed in the unit of lysozyme ml−1 serum. A unit of lysozyme activity was defined as the amount of serum causing a reduction of 0.001 per minute at 450 nm at 22 °C.
Serum bactericidal activity was determined using the previously described method by Kajita et al. [26]. Sera samples were diluted three times with 0.1% gelatin-veronal buffer (GVBC2) (pH = 7.5). A. hydrophila was suspended in the same buffer to make a concentration of 105 CFU ml−1. The diluted sera and bacteria were mixed at 1:1 ratio and incubated for 90 min at 25 °C. The number of viable bacteria was then calculated by counting the colonies from the resulting mixture incubated on TSA plates for 24 h in duplicate. The bactericidal activity of test serum was expressed as a percentage of the ratio of colony forming units in the experimental group to those in the control group.
NBT reduction: Blood (0.1 ml) was placed in micro titer plate wells; an equal amount of 0.2% NBT solution was added in each well and incubated for 30 min at room temperature. A sample of NBT blood cell suspension (0.1 ml) was added to a glass tube containing 2 ml N, N-dimethyl formamide and centrifuged for 5 min at 3000g. The optical density of supernatant was measured in a spectrophotometer (Biophotometer, Eppendorf, Germany) at 620 nm [21, 60].
The complement activity was assayed using Rabbit Red Blood Cells (RaRBC) as a target. RaRBC were prepared in 1.5% agarose (pH = 7.2) containing 0.5 mM MgCl2 and 1.5 mM CaCl2. The RaRBCs in agarose were washed with PBS (0.1 M pH = 7.0) and centrifuged at 750×g for 5 min, and the cell concentration is adjusted to 1 × 108 cell ml−1. Agarose containing RaRBC was dispensed into plates, incubated at 4 °C and hole punched (3 mm in diameter). Subsequently, each hole was filled with 15 μl of serum of T. grypus and was incubated at room temperature. After 24 h of incubation, the zone of lysis was measured and expressed in AU/ml [9, 34].
Statistical Analysis
All treatment data were checked for normality (Shapiro-Wilk) and Leven’s test for homogeneity of variance. If normality and homogeneity were achieved, general linear model, Univariate ANOVA was used. When interaction effects between different independent parameters were not significant, one-way ANOVA and Tukey’s multiple comparison test were used to determine the significant variation (P < 0.05) in the immune response between the control and experimental groups. All statistics were performed using SPSS for version 19.0 (SPSS, Chicago, USA). All experimental data were presented as the mean ± SE, and the level of significance for all tests was set at P < 0.05. The Pearson correlation test was used to find any correlation between immunological parameters and the respective immune gene expression.
Result
Challenge Test
Analysis of mortalities after challenge-testing with A. hydrophila showed significant differences in the resistance to bacterial infection among probiotic-treated and control groups (P < 0.05). Although there were no significant differences among the probiotic-treated groups (P > 0.05), the A. hydrophilla caused the first mortalities at 20 h post-infection (hpi) in all infected groups. The mortality rate of fish fed with diet supplemented with L. bulgaricus, L. plantarum, and L. casei was significantly higher than that of fish fed with the control diet. Mortalities continuously occurred until 48 hpi. The highest mortality rate was recorded at 30 hpi in all groups (Table 2).
Relative mRNA Expression of Immune-Related Genes
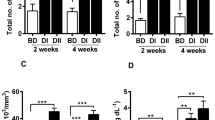
The results of expression of immune-related genes (IL-1β, IL-8, and TNF-α) in the head kidneys of T. grypus have been shown in Figs. 1–3.

Gene expression of TNF-α from juvenile T. grypus. Fish were fed with diet containing L. plantarum, L. bulgaricus subsp. Bulgaricus, L. casei, and control (diet/diets without probiotics). Samples were taken at different time after challenge with A. hydrophila. Values are shown as means ± standard error (n = 3). Different lowercase superscript letters denote a significant difference between values in each row (P < 0.05). Different capital superscripts denote significant differences within columns (P < 0.05)
As shown in Fig. 1, in all probiotic-treated groups, a mild upregulation in TNF-α gene expression was observed at 6 and 12 h post A. hydrophila injection (P > 0.005). A significant upregulation in the level of expression of TNF-α gene in fish fed diet supplemented with L. plantarom, L. bulgaricus, or L. casei was seen in both 24 and 48 h post-injection (P < 0.05). A slight increase occurred in the expression of TNF-α gene in all probiotic-treated groups 7 days after challenge (P = 0.085).
Moreover, the level of IL-1β gene expression was low in the head kidneys of all groups before infection at day zero. A statistically significant increase in the expression level of IL-1β was observed in all three probiotic-treated groups at 24 and 48 h after infection A. hydrophila (P < 0.05). The highest upregulation (3.27 ± 1.32) belongs to L. plantarum group at 24 h after challenge (Fig. 2).

Gene expression of IL1-β from juvenile T. grypus. Fish were fed with diet containing L. plantarum, L. bulgaricus subsp. Bulgaricus, L. casei, and control (diet/diets without probiotics). Samples were taken at different time after challenge with A. hydrophila. Values are shown as means ± standard error (n = 3). Different lowercase superscript letters denote a significant difference between values in each row (P < 0.05). Different capital superscript letters denote significant differences within columns (P < 0.05)
An insignificant increase in IL-8 gene expression of probiotic-treated groups at 6, 12, and 48 h post-challenge was reported (P > 0.05). IL-8 gene expression in the plantarum and bulgaricus groups was significantly (P < 0.05) higher at 24 h after infection in comparison to the control group (Fig. 3).

Gene expression of IL-8 from juvenile T. grypus. Fish were fed with diet containing L. plantarum, L. bulgaricus subsp. Bulgaricus, L. casei, and control (diet/diets without probiotics). Samples were taken at different time after challenge with A. hydrophila. Values are shown as means ± standard error (n = 3). Different lowercase superscript letters denote a significant difference between values in each row (P < 0.05). Different capital superscript letters denote significant differences within columns (P < 0.05)
There was no significant difference in the levels of IL-1β and IL-8 gene expression among the groups 7 days after challenge (P > 0.05).
Non-specific Immune Responses
The results of two-way ANOVA of non-specific immune response parameters of fish fed with a diet containing different probiotic contents have been presented in Table 3.
Serum lysozyme activity gradually increased at 12, 24, and 48 h post-infection with A. hydrophila in all probiotic-treated groups (Table 4). Meanwhile, the highest increase in serum lysozyme activity in different sampling intervals was observed in the L. plantarum, followed by L. bulgaricus and then L. casei groups which were statistically higher than the control group (P < 0.05).
Serum bactericidal activity increased in all three probiotic-treated groups compared to the control group in sampling intervals except on day 7 post-challenge (P < 0.05). The probiotic species did not affect serum bactericidal activity after the challenge with A. hydrophila of T. grypus in different sampling intervals (P > 0.05).
Although the NBT reduction was enhanced in L. plantarum and L. casei groups 24, 48 h, and 7 days after challenge (P < 0.05), no significant change occurred in other sampling intervals and other groups compared to the control (P > 0.05).
In comparison to the controls, complement activity was significantly higher in the L. plantarum and L. bulgaricus supplemented groups in all sampling intervals (P < 0.05). The highest complement activity was recorded in the L. plantarum supplemented group at 48 h after A. hydrophila infection. A graduate increase pattern was observed in the complement activity almost in all probiotic-treated groups (Table 4).
Discussion
The involvement of probiotics in nutrition, disease resistance, and other beneficial activities in fish has been proven beyond any doubt. Among the numerous health benefits attributed to probiotics, modulation of immune system is one of the most commonly purported benefits of the probiotics and their potency to stimulate the immunity under in vitro and in vivo conditions is noteworthy [20, 35].
Results of present study showed that both native isolated Lactobacilli were not only successful in increasing the expression of immune-related gene in head kidney of T. grypus, but they also deeply impacted on selected serum immune parameters and their resistance to bacterial infection of A. hydrophila. Besides, a high positive correlation was recorded in expression of immune-related gene in head kidneys and serum non-specific immune response parameters which confirmed the stimulation of primary immune organs in the gene level and humeral immune response in serum. These findings may be very useful and promising for commercial aquaculture and may help to protect the fish against bacterial infection because aquatic animals are continually vulnerable to numerous opportunistic pathogens [22]. Our results were in agreement with results of previous studies, demonstrating that oral administration of probiotics impacted on immune regulatory proteins which resulted in enhanced protection against pathogens [36, 62].
Probiotics are sometimes unable to colonize viably and predominantly in the host’s intestine and relatively ineffective as transient flora [33] but probably indigenous Lactobacillus sp. can colonize in the intestine of fish. At this study, T grypus fed with a diet containing the indigenous probiotic bacteria L. bulgaricus and L. plantarum showed immune stimulation after challenge with A. hydrophila, possibly due to the promotion in the immune-related protein level in immune organs [49]. We predict that these bacteria are highly adhesive to GI tract of T. grypus because these bacteria are autochthonous. In previous studies, the beneficial effects of probiotic administration against A. hydrophila infection in fish have been demonstrated with dietary probiotic supplementations of Aeromonas sobria GC2 [10], Leuconostoc mesenteroides LFP 196 and L. plantarum CLFP 238 [61], L. plantarum subsp. plantarum CLFP 3, Lactococcus lactis subsp. cremoris CLFP 25 and L. mesenteroides CLFP68 [43], L. acidophilus and L. brevis [30], and L. acidophilus [2]. As far as we are aware, this study is the first report of a high positive correlation between immunological parameters and the respective immune-related gene expression against bacterial infection in fish treated with probiotics.
Cytokines are the most important modulators for initiation and developing a perfect immune response; moreover, the investigation of cytokines functions through studying their expression profile may provide valuable data that can clarify the immunostimulatory mechanisms of probiotics in aquaculture [58]. Probiotic benefits, more specifically the use of autochthonous probiotics as immune enhancers, have not been tested in the Tor (Barbus) genus fish; however, extensive studies regarding other fish species have been done. In the current study, a series of immune-related genes was used as a primary biomarker to characterize the effects of indigenous probiotics on innate immune responses of T. grypus after intraperitoneal injection (ip injection) of A. hydrophila. Besides, the correlation between expression of these cytokines’ genes and humeral non-specific immune defense parameters was evaluated. Involvement of probiotics in upregulating the gene expression of immune relevant cytokines, the first line of immune defense mechanism, is already recorded [58]. The bacterial mono-association studies on gnotobiotic fish also indicated the upregulation of serum amyloid A1, C-reactive protein, and complement components [48].
IL-1β is a prototypic pro-inflammatory cytokine which accelerates additional inflammatory processes by inducing other inflammatory molecules like TNF and IL-8 [30, 53, 63]. In the present study, a significant upregulation in IL-1β expression was observed in all probiotic-treated groups, but the highest expression of IL-1β was seen in L. plantarum-treated group followed by L. bulgaricus- and then L. casaei-treated groups, respectively (P < 0.05). The subsequent decline in these transcripts at day 7 post-infection may be well correlated with the reduction in activity of serum immune defense parameters. Probably, probiotics induced more expression of IL-1β in leukocytes and macrophages of head kidneys [64, 65]. The lymphocytes and macrophages have binding sites for peptidoglycan (a cell wall component of LAB) which can stimulate the secretion of IL-1β. Although no significant differences were observed in all probiotic-treated groups at 6 and 12 h post-injection, upregulation in probiotics groups (after 6 h) is possibly related to IL-1β which is one of the initiators and drivers of cytokines released during inflammatory response in fish to this bacterium [19]. An increased level of IL-1β expression was reported in carp injected with bacterial LPS [23] and in Atlantic cods injected with IPNV after 24 and 48 h, respectively [56]. On the other hand, Zebrafish and Puntius sarana injected with A. hydrophila showed upregulation in IL1-β expression at 1 h post-challenge and it began to decline to control levels at 6 hpi [16, 50]. Significant upregulation of IL1-β in fish treated with probiotics provides evidence that early inflammatory immune response is more stimulated in probiotic-treated groups infected with A. hydrophila, and this stimulation of IL-1β showed a high positive correlation with other serum innate immune defense parameters like lysozyme (R 2 = 0.572; P < 0.0001), complement (R 2 = 0.596; P < 0.0001), and NBT reduction (R 2 = 0.634; P < 0.0001) which seem to play a key role in controlling bacterial pathogenesis (Table 5).
In this study, the highest upregulation in TNFα expression was obtained in L. bulgaricus-treated groups and afterward in L. plantarum group both at 24 and 48 h post-infection with A. hydrophila, compared to the control group, but a gradual reduction in TNFα expression occurred in all groups treated with probiotic after 24 hpi. The pleiotropic cytokine TNFα has been shown to be an important component of the innate immunity and pro-inflammatory response of fish [24]. In this regard, several immune gene expression studies have shown the increased levels of expression of TNFα in relation to different probiotic diets, such as Canrobacterium maltaromaticum, Carnobacterium divergens [27], L. plantarum [43], and Lactobacillus rhamnosus [41].
As other researchers suggested, this study also revealed that pro-inflammatory cytokines such as IL-1β, and TNFα could reach to the maximum level in systemic immune tissues in comparison to local tissue where pathogens proliferate [51]. Likewise, IL-1β and TNFα transcripts increased in all injected groups and even in the challenge control group, but they slightly increased in non-challenge control, indicating a trend toward upregulation of inflammatory genes after bacterial infection. On the other hand, the higher expression of cytokines genes in the head kidneys of T. grypus fed with indigenous probiotics supplemented food was compared to that of T. grypus fed with exogenous probiotic (L. casei); therefore, this comparison demonstrated the influence of probiotic bacterial origin on its immunomodulatory effects. The results also highly correlated with the increase of IL-1β and TNFα.
The expression of IL-8 in the head kidneys at days 1 and 2 indicates chemotaxis in immune response at the site of infection. IL-8, also known as neutrophil chemotactic factor, has two primary functions. It induces chemotaxis in target cells, primarily neutrophils, then other granulocytes, causing them to migrate toward the site of infection. IL-8 also induces phagocytosis once they have arrived. IL-8 production is stimulated by the expression of IL-1β and TNF-α, so it is not surprising to see the simultaneous expression of these three cytokines. A high positive correlation was seen in the expression of IL-8 with two other immune-related genes, IL-1β (R 2 = 0.736; P < 0.0001) and TNFα (R 2 = 0.723; P < 0.0001). It shows that whole immune stimulation occurred in probiotic-treated fish and also all immune-related genes in fish head kidneys upregulated in a similar pattern. Interestingly, food supplementation of T. grypus, fed with indigenous probiotic bacteria (L. bulgaricus and L. plantarom), caused higher expression of IL-8 than that of T. grypus fed with exogenous probiotic bacteria (L. caseai). A more appropriate indigenous bacterial colonization in the intestine may explain this increase in IL-8 [27]. The alteration of the intestinal microflora may explain this increase in IL-8. However, further tests are needed to verify this hypothesis. As reported in this study, the magnitude of the IL-8 transcriptional response to a range of inflammatory stimuli was found to be less than the IL-1β and TNFα transcriptional response [32]. Results obtained in the present research are in agreement with previous studies in rainbow trout (Oncorhynchus mykiss) that showed supplemented diet with probiotics increased the expression of proinflammatory cytokines, including IL-8 [43] in haddock (Melanogrammus aeglefinus) tissues injected with bacterial LPS [15]. It is interesting to find that upregulation in the expression of IL-1β, TNF-α, and IL-8 genes was observed at early stages of infection (24 and 48 h post-challenge); whereas, downregulation of this gene expression occurred 7 days after challenge. The immune response of fish against A. hydrophila depends on the co-operation of both humeral and cellular immune responses which initially are conducted in primary immunocompetent organs by upregulating the expression of immunomudulator cytokines. Autochthonous probiotics indicated that host immunological responses were responsible for mediating high immune responses of these treatments. According to gene expression assay of study, we found not only significant upregulation of immune relevant genes in probiotic-treated fish, but also faster upregulation, more stable expression, and slower decline of three immune-related genes (IL-1β, TNF-α, and Il-8) in fish receiving indigenous probiotics (L. plantarum and L. bulgaricus) compared with fish fed with exogenous probiotics. Besides, a high positive correlation was seen within expression of three immune relevant genes or between these genes expression and serum humeral immune response parameters. Despite these in vivo studies, some in vitro studies are necessary to better understand the interaction between these probiotic strains and the immune cell and organs in fish.
A result of this study showed that food supplementation with L. plantarum and L. bulgaricus were successful in stimulating of serum immune parameters of T. grypus. The serum lysozyme activity of probiotic-treated groups significantly increased (P < 0.001) from 12 h until 7 days post-challenge. Besides, a statistically significant correlation between immune relevant genes expression and lysozyme activity was reported in the probiotic-treated groups. Lysozyme, one of the important bactericidal enzymes of innate immunity, is an indispensable tool for fish to fight against infectious agents. Probiotics are found to trigger the lysozyme level in teleost [6]. Probiotics like L. rhamnosus, Carnobacterium maltaromaticum, C. divergens in O. mykiss [38], L. lactis ssp. Lactis, and L. mesenteroides in Salmo trutta [6] enhanced the lysozyme level; NBT reduction, an indicator for respiratory burst activity of immune-related cells, increased in indigenous probiotic-treated groups compared to the control groups (P < 0.05). Superoxide anion production during the respiratory burst of phagocytes can be induced by a variety of phagocyte activating agents [25, 29, 54]. The findings of respiratory burst activity following the probiotics treatment in fish are often contradictory, while some studies indicated probiotics did not have any significant impact on this non-specific defense mechanism of fish [17, 57]. Several in vitro and in vivo studies showed a significant increase in respiratory burst activity by various probiotics in many aquatic animals including fish. Probiotics like Bacillus subtilis and certain members of LAB group can stimulate respiratory burst activity in fish [64]. Heat inactivated L. delbrueckii and B. subtilis under in vitro condition and also enhanced activity of head kidney leucocytes of gilt-head sea bream [52]. This study further confirmed that the probiotics might be responsible for degrading free radicals production by host phagocytic cells. The NBT level before infection and on day 7 post-infection was almost similar. The highest correlation between the immune relevant genes expression and serum immune parameters was recorded between IL 1β expression and NBT reduction (R 2 = 0. 634, P < 0.0001).
There is a general consensus that probiotics from autochthonous sources have a greater chance of competing with resident microbes and of becoming predominant within a short period of intake and persisting in the colonic environment for a period of time after the withdrawal of probiotics [52]. For instance, Carnevali et al. [13] recorded a significant decrease in larva and fry mortalities using Lactobacillus fructivorans, isolated from gut of S. aurata. Furthermore, it is assumed that the host’s immune cells do not naturally react with bacteria on the surfaces and in nature [52]. Complement, a component of the non-specific immune response, may have effector mechanisms such as the direct killing of microorganisms by lysis. Probiotics can enhance natural complement activity of fish; moreover, it was reported that the diet and water along with probiotics treatments could stimulate fish different complement components.
It is also worth noting that non-viable probiotics can stimulate complement components in fish. Choi and Yoon [14] found an increased complement activity in O. mykiss from 4th week of feeding with the heat-inactivated probiotics. The hemolytic activity of serum of T. grypus, which increased significantly after 6 h challenge with A. hydrophila, was estimated by the total complement activity (P < 0.001). Complement activation is usually beneficial to the host. However, persistent activation of complement in severe bacterial infection could lead to adverse effects and immunosuppression to the host [42]. However, it returned to the normal level quickly in the survivors 7 days after challenge. The elevation of immune status in the probiotic fish as demonstrated here might indirectly reflect the effect of indigenous probiotics on outcomes in long-term protection against A. hydrophila or on a decrease in bacterial load from the circulation or body in the survivors. The higher serum complement activity and the enhanced serum bactericidal power corresponded with the TNF-α and IL-1β expressions in the probiotic supplemented groups.
In the innate humeral response, bactericidal activity of serum played an important role in the immune system. Additional evidence exists in rainbow trout where the complement activity and IL-1β gene expression were enhanced in fish fed with the Enterococcus faecium and L. rhamnosus supplemented diet [38, 40, 47]. Das et al. [16] suggested that Bacillus amyloliquefaciens was a potential probiotic species and could improve the immune response.
Total mortalities following experimental infection with A. hydrophila were significantly lower in fish fed with the probiotic supplemented diet (23 to 36% mortality), compared to the control group (70% mortality). There were no remarkable differences in resistance against bacterial infection in different groups treated with probiotics (P > 0.05). Our results were similar to the findings of Giri et al. [22], who recorded a lower mortality rate in fish fed on a diet containing probiotics compared with those fed on normal diet. Dietary supplementation of food with defined probiotics may be effective bio-therapeutic or prophylactic means in aquaculture [37, 39]. Our findings suggested that feeding with probiotics for 8 weeks is effective in increasing the resistance against Aeromonas infections in fish. In similar work L. rhamnosus was successfully used as a feed additive in tilapia [46] and rainbow trout [14] to prevent Edwardsiellosis. The probiotic bacteria can induce inflammatory responses and increase phagocytosis [6, 28]. It can be expected that the increase of immune relevant gene expression and humeral immune parameters in probiotic-treated fish can simultaneously improve resistance against infection of A. hydrophila.
Based on the findings of this work, it can be concluded that food supplementation with autochthonous probiotics (L. plantarum and L. bulgaricus) not only stimulate non-specific immune parameters and resistance to bacterial infection, but also upregulate expression of immune relevant genes with a high positive correlation. Then, probiotic-treated fish possess more effective immunity status against various pathogens especially A. hydrophila infection.
References
Adams M (1999) Safety of industrial lactic acid bacteria. J Biotechnol 68(2):171–178. https://doi.org/10.1016/S0168-1656(98)00198-9
Aly SM, Ahmed YA-G, Ghareeb AA-A, Mohamed MF (2008) Studies on Bacillus subtilis and Lactobacillus acidophilus, as potential probiotics, on the immune response and resistance of Tilapia nilotica (Oreochromis niloticus) to challenge infections. Fish Shellfish Immunol 25(1):128–136. https://doi.org/10.1016/j.fsi.2008.03.013
Amann RI (1995) In situ identification of micro-organisms by whole cell hybridization with rRNA-targeted nucleic acid probes. In: Molecular microbial ecology manual. Springer, Berlin, pp 331–345. https://doi.org/10.1007/978-94-011-0351-0_23
Askarian F, Matinfar A, Kousha A, Bahmani M, Khorshidi K, Shenavar A, Ringo E (2008) Diversity of lactic acid bacteria in the gastrointestinal tracts of reared beluga (Huso huso) and Persian sturgeon (Acipenser persicus): a comparative study. J Fish Aquat Sci 3(5):302–311. https://doi.org/10.3923/jfas.2008.302.311
Balcázar JL, De Blas I, Ruiz-Zarzuela I, Cunningham D, Vendrell D, Múzquiz JL (2006) The role of probiotics in aquaculture. Vet Microbiol 114(3):173–186. https://doi.org/10.1016/j.vetmic.2006.01.009
Balcázar JL, De Blas I, Ruiz-Zarzuela I, Vendrell D, Calvo AC, Márquez I, Gironés O, Muzquiz JL (2007) Changes in intestinal microbiota and humoral immune response following probiotic administration in brown trout (Salmo trutta). Br J Nutr 97(03):522–527. https://doi.org/10.1017/S0007114507432986
Balcázar JL, Vendrell D, de Blas I, Ruiz-Zarzuela I, Muzquiz JL, Girones O (2008) Characterization of probiotic properties of lactic acid bacteria isolated from intestinal microbiota of fish. Aquaculture 278(1):188–191. https://doi.org/10.1016/j.aquaculture.2008.03.014
Banerjee G, Ray AK (2017) The advancement of probiotics research and its application in fish farming industries. Res Vet Sci 115:66–77. https://doi.org/10.1016/j.rvsc.2017.01.016
Barta O (1993) Veterinary clinical immunology laboratory. Bar-lab Inc., PO Box 11819
Brunt J, Austin B (2005) Use of a probiotic to control lactococcosis and streptococcosis in rainbow trout, Oncorhynchus mykiss (Walbaum). J Fish Dis 28(12):693–701. https://doi.org/10.1111/j.1365-2761.2005.00672.x
Bustin SA, Benes V, Garson JA, Hellemans J, Huggett J, Kubista M, Mueller R, Nolan T, Pfaffl MW, Shipley GL (2009) The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin Chem 55(4):611–622. https://doi.org/10.1373/clinchem.2008.112797
Campos-perez J, Ward M, Grabowski P, Ellis A, Secombes C (2000) The gills are an important site of iNOS expression in rainbow trout Oncorhynchus mykiss after challenge with the Gram-positive pathogen Renibacterium salmoninarum. Immunology 99(1):153–161. https://doi.org/10.1046/j.1365-2567.2000.00914.x
Carnevali O, Zamponi MC, Sulpizio R, Rollo A, Nardi M, Orpianesi C, Silvi S, Caggiano M, Polzonetti AM, Cresci A (2004) Administration of probiotic strain to improve sea bream wellness during development. Aquacult Int 12(4–5):377–386. https://doi.org/10.1023/B:AQUI.0000042141.85977.bb
Choi S-H, Yoon T-J (2008) Non-specific immune response of rainbow trout (Oncorhynchus Mykiss) by dietary heat-inactivated potential probiotics. Immune Network 8(3):67–74. https://doi.org/10.4110/in.2008.8.3.67
Corripio-Miyar Y, Bird S, Tsamopoulos K, Secombes C (2007) Cloning and expression analysis of two pro-inflammatory cytokines, IL-1β and IL-8, in haddock (Melanogrammus aeglefinus). Mol Immunol 44(6):1361–1373. https://doi.org/10.1016/j.molimm.2006.05.010
Das A, Sahoo P, Mohanty B, Jena J (2011) Pathophysiology of experimental Aeromonas hydrophila infection in Puntius sarana: early changes in blood and aspects of the innate immune-related gene expression in survivors. Vet Immunol Immunopathol 142(3):207–218. https://doi.org/10.1016/j.vetimm.2011.05.017
Díaz-Rosales P, Arijo S, Chabrillón M, Alarcón F, Tapia-Paniagua S, Martínez-Manzanares E, Balebona M, Moriñigo M (2009) Effects of two closely related probiotics on respiratory burst activity of Senegalese sole (Soleasenegalensis, Kaup) phagocytes, and protection against Photobacteriumdamselae subsp. piscicida. Aquaculture 293(1):16–21. https://doi.org/10.1016/j.aquaculture.2009.03.050
Dimitroglou A, Merrifield DL, Carnevali O, Picchietti S, Avella M, Daniels C, Güroy D, Davies SJ (2011) Microbial manipulations to improve fish health and production—a Mediterranean perspective. Fish Shellfish Immunol 30(1):1–16. https://doi.org/10.1016/j.fsi.2010.08.009
Engelsma MY, Huising MO, van Muiswinkel WB, Flik G, Kwang J, Savelkoul HF, Verburg-van Kemenade BL (2002) Neuroendocrine–immune interactions in fish: a role for interleukin-1. Vet Immunol Immunopathol 87(3):467–479. https://doi.org/10.1016/S0165-2427(02)00077-6
Gatesoupe F (1999) The use of probiotics in aquaculture. Aquaculture 180(1):147–165. https://doi.org/10.1016/S0044-8486(99)00187-8
Geng X, Dong X, Tan B, Yang Q, Sy C, Hy L, Xq L (2012) Effects of dietary probiotic on the growth performance, non-specific immunity and disease resistance of cobia, Rachycentron canadum. Aquac Nutr 18(1):46–55. https://doi.org/10.1111/j.1365-2095.2011.00875.x
Giri SS, Sukumaran V, Oviya M (2013) Potential probiotic Lactobacillus plantarum VSG3 improves the growth, immunity, and disease resistance of tropical freshwater fish, Labeo rohita. Fish Shellfish Immunol 34(2):660–666. https://doi.org/10.1016/j.fsi.2012.12.008
Gonzalez SF, Buchmann K, Nielsen ME (2007) Real-time gene expression analysis in carp (Cyprinus carpio L.) skin: inflammatory responses caused by the ectoparasite Ichthyophthirius multifiliis. Fish Shellfish Immunol 22(6):641–650. https://doi.org/10.1016/j.fsi.2006.08.011
Grayfer L, Walsh JG, Belosevic M (2008) Characterization and functional analysis of goldfish (Carassius auratus L.) tumor necrosis factor-alpha. Dev Comp Immunol 32(5):532–543. https://doi.org/10.1016/j.dci.2007.09.009
Ji P-F, Yao C-L, Wang Z-Y (2009) Immune response and gene expression in shrimp (Litopenaeus vannamei) hemocytes and hepatopancreas against some pathogen-associated molecular patterns. Fish Shellfish Immunol 27(4):563–570. https://doi.org/10.1016/j.fsi.2009.08.001
Kajita Y, Sakai M, Atsuta S, Kobayashi M (1990) The immunomodulatory effects of levamisole on rainbow trout, Oncorhynchus mykiss. Fish Pathol 25(2):93–98. https://doi.org/10.3147/jsfp.25.93
Kim D-H, Austin B (2006) Innate immune responses in rainbow trout (Oncorhynchus mykiss, Walbaum) induced by probiotics. Fish Shellfish Immunol 21(5):513–524. https://doi.org/10.1016/j.fsi.2006.02.007
Korkea-aho T, Papadopoulou A, Heikkinen J, Von Wright A, Adams A, Austin B, Thompson K (2012) Pseudomonas M162 confers protection against rainbow trout fry syndrome by stimulating immunity. J Appl Microbiol 113(1):24–35. https://doi.org/10.1111/j.1365-2672.2012.05325.x
Li C, Yao C-L (2013) Molecular and expression characterizations of interleukin-8 gene in large yellow croaker (Larimichthys crocea). Fish Shellfish Immunol 34(3):799–809. https://doi.org/10.1016/j.fsi.2012.12.019
Liu W, Ren P, He S, Xu L, Yang Y, Gu Z, Zhou Z (2013) Comparison of adhesive gut bacteria composition, immunity, and disease resistance in juvenile hybrid tilapia fed two different Lactobacillus strains. Fish Shellfish Immunol 35(1):54–62. https://doi.org/10.1016/j.fsi.2013.04.010
Merrifield DL, Dimitroglou A, Foey A, Davies SJ, Baker RT, Bøgwald J, Castex M, Ringø E (2010) The current status and future focus of probiotic and prebiotic applications for salmonids. Aquaculture 302(1):1–18. https://doi.org/10.1016/j.aquaculture.2010.02.007
Miettinen M, Vuopio-Varkila J, Varkila K (1996) Production of human tumor necrosis factor alpha, interleukin-6, and interleukin-10 is induced by lactic acid bacteria. Infect Immun 64(12):5403–5405
Mohammadian T, Alishahi M, Tabandeh MR, Ghorbanpoor M, Gharibi D, Tollabi M, Rohanizade S (2016) Probiotic effects of Lactobacillus plantarum and L. delbrueckii ssp. bulguricus on some immune-related parameters in Barbus grypus. Aquacult Int 24(1):225–242. https://doi.org/10.1007/s10499-015-9921-8
NavinChandran M, Iyapparaj P, Moovendhan S, Ramasubburayan R, Prakash S, Immanuel G, Palavesam A (2014) Influence of probiotic bacterium Bacillus cereus isolated from the gut of wild shrimp Penaeus monodon in turn as a potent growth promoter and immune enhancer in P. monodon. Fish Shellfish Immunol 36(1):38–45. https://doi.org/10.1016/j.fsi.2013.10.004
Nayak S (2010) Probiotics and immunity: a fish perspective. Fish Shellfish Immunol 29(1):2–14. https://doi.org/10.1016/j.fsi.2010.02.017
Nikoskelainen S, Ouwehand A, Salminen S, Bylund G (2001) Protection of rainbow trout (Oncorhynchus mykiss) from furunculosis by Lactobacillus rhamnosus. Aquaculture 198(3):229–236. https://doi.org/10.1016/S0044-8486(01)00593-2
Nikoskelainen S, Ouwehand AC, Bylund G, Salminen S, Lilius E-M (2003) Immune enhancement in rainbow trout (Oncorhynchus mykiss) by potential probiotic bacteria (Lactobacillus rhamnosus). Fish Shellfish Immunol 15(5):443–452. https://doi.org/10.1016/S1050-4648(03)00023-8
Panigrahi A, Kiron V, Kobayashi T, Puangkaew J, Satoh S, Sugita H (2004) Immune responses in rainbow trout Oncorhynchus mykiss induced by a potential probiotic bacteria Lactobacillus rhamnosus JCM 1136. Vet Immunol Immunopathol 102(4):379–388
Panigrahi A, Kiron V, Puangkaew J, Kobayashi T, Satoh S, Sugita H (2005) The viability of probiotic bacteria as a factor influencing the immune response in rainbow trout Oncorhynchus mykiss. Aquaculture 243(1):241–254. https://doi.org/10.1016/j.aquaculture.2004.09.032
Panigrahi A, Kiron V, Satoh S, Hirono I, Kobayashi T, Sugita H, Puangkaew J, Aoki T (2007) Immune modulation and expression of cytokine genes in rainbow trout Oncorhynchus mykiss upon probiotic feeding. Dev Comp Immunol 31(4):372–382. https://doi.org/10.1016/j.dci.2006.07.004
Panigrahi A, Viswanath K, Satoh S (2011) Real-time quantification of the immune gene expression in rainbow trout fed different forms of probiotic bacteria Lactobacillus rhamnosus. Aquac Res 42(7):906–917. https://doi.org/10.1111/j.1365-2109.2010.02633.x
Peakman M, Senaldi G, Vergani D (1989) Review: assessment of complement activation in clinical immunology laboratories: time for reappraisal? J Clin Pathol 42(10):1018–1025. https://doi.org/10.1136/jcp.42.10.1018
Pérez-Sánchez T, Balcázar JL, Merrifield DL, Carnevali O, Gioacchini G, de Blas I, Ruiz-Zarzuela I (2011) Expression of immune-related genes in rainbow trout (Oncorhynchus mykiss) induced by probiotic bacteria during Lactococcus garvieae infection. Fish Shellfish Immunol 31(2):196–201. https://doi.org/10.1016/j.fsi.2011.05.005
Pfaffl MW (ed) (2009) QPCR 2009 event: proceedings. Technical University of Munich, Munich
Picchietti S, Fausto AM, Randelli E, Carnevali O, Taddei AR, Buonocore F, Scapigliati G, Abelli L (2009) Early treatment with Lactobacillus delbrueckii strain induces an increase in intestinal T-cells and granulocytes and modulates immune-related genes of larval Dicentrarchus labrax (L.) Fish Shellfish Immunol 26(3):368–376. https://doi.org/10.1016/j.fsi.2008.10.008
Pirarat N, Kobayashi T, Katagiri T, Maita M, Endo M (2006) Protective effects and mechanisms of a probiotic bacterium Lactobacillus rhamnosus against experimental Edwardsiella tarda infection in tilapia (Oreochromis niloticus). Vet Immunol Immunopathol 113(3):339–347. https://doi.org/10.1016/j.vetimm.2006.06.003
Pirarat N, Pinpimai K, Endo M, Katagiri T, Ponpornpisit A, Chansue N, Maita M (2011) Modulation of intestinal morphology and immunity in nile tilapia (Oreochromis niloticus) by Lactobacillus rhamnosus GG. Res Vet Sci 91(3):e92–e97. https://doi.org/10.1016/j.rvsc.2011.02.014
Rawls JF, Mahowald MA, Goodman AL, Trent CM, Gordon JI (2007) In vivo imaging and genetic analysis link bacterial motility and symbiosis in the zebrafish gut. Proc Natl Acad Sci 104(18):7622–7627. https://doi.org/10.1073/pnas.0702386104
Robertson P, O'Dowd C, Burrells C, Williams P, Austin B (2000) Use of Carnobacterium sp. as a probiotic for Atlantic salmon (Salmo salar L.) and rainbow trout (Oncorhynchus mykiss, Walbaum). Aquaculture 185(3):235–243. https://doi.org/10.1016/S0044-8486(99)00349-X
Rodríguez I, Novoa B, Figueras A (2008) Immune response of zebrafish (Danio rerio) against a newly isolated bacterial pathogen Aeromonas hydrophila. Fish Shellfish Immunol 25(3):239–249. https://doi.org/10.1016/j.fsi.2008.05.002
Salazar-Mather TP, Hokeness KL (2003) Calling in the troops: regulation of inflammatory cell trafficking through innate cytokine/chemokine networks. Viral Immunol 16(3):291–306. https://doi.org/10.1089/088282403322396109
Salinas I, Díaz-Rosales P, Cuesta A, Meseguer J, Chabrillón M, Morinigo MA, Esteban MA (2006) Effect of heat-inactivated fish and non-fish derived probiotics on the innate immune parameters of a teleost fish (Sparus aurata L.) Vet Immunol Immunopathol 111(3):279–286. https://doi.org/10.1016/j.vetimm.2006.01.020
Savan R, Sakai M (2006) Genomics of fish cytokines. Comp Biochem Physiol Genom Proteom 1(1):89–101
Secombes C, Fletcher T (1992) The role of phagocytes in the protective mechanisms of fish. Annu Rev Fish Dis 2:53–71. https://doi.org/10.1016/0959-8030(92)90056-4
Secombes C, Wang T, Hong S, Peddie S, Crampe M, Laing K, Cunningham C, Zou J (2001) Cytokines and innate immunity of fish. Dev Comp Immunol 25(8):713–723. https://doi.org/10.1016/S0145-305X(01)00032-5
Seppola M, Larsen AN, Steiro K, Robertsen B, Jensen I (2008) Characterisation and expression analysis of the interleukin genes, IL-1β, IL-8 and IL-10, in Atlantic cod (Gadus morhua L.) Mol Immunol 45(4):887–897. https://doi.org/10.1016/j.molimm.2007.08.003
Sharifuzzaman S, Austin B (2009) Influence of probiotic feeding duration on disease resistance and immune parameters in rainbow trout. Fish Shellfish Immunol 27(3):440–445. https://doi.org/10.1016/j.fsi.2009.06.010
Shima JS, Osenberg CW (2003) Cryptic density dependence: effects of covariation between density and site quality in reef fish. Ecology 84(1):46–52. https://doi.org/10.1890/0012-9658(2003)084[0046:CDDEOC]2.0.CO;2
Talpur AD, Ikhwanuddin M, Abdullah MDD, Bolong A-MA (2013) Indigenous Lactobacillus plantarum as probiotic for larviculture of blue swimming crab, Portunus pelagicus (Linnaeus, 1758): effects on survival, digestive enzyme activities and water quality. Aquaculture 416:173–178
Torfi Mozanzadeh M, Marammazi J, Yaghoubi M, Yavari V, Agh N, Gisbert E (2015) Somatic and physiological responses to cyclic fasting and re-feeding periods in sobaity sea bream (Sparidentex hasta, Valenciennes 1830). Aquacult Nutr 23(1):181–191
Vendrell D, Balcazar JL, de Blas I, Ruiz-Zarzuela I, Gironés O, Muzquiz JL (2008) Protection of rainbow trout (Oncorhynchus mykiss) from lactococcosis by probiotic bacteria. Comp Immunol Microbiol Infect Dis 31(4):337–345. https://doi.org/10.1016/j.cimid.2007.04.002
Yasui H, Nagaoka N, Mike A, Hayakawa K, Ohwaki M (1992) Detection of Bifidobacterium strains that induce large quantities of IgA. Microb Ecol Health Dis 5(3):155–162. https://doi.org/10.3109/08910609209141310
Zhao L, Sun J-S, Sun L (2011) The g-type lysozyme of Scophthalmus maximus has a broad substrate spectrum and is involved in the immune response against bacterial infection. Fish Shellfish Immunol 30(2):630–637. https://doi.org/10.1016/j.fsi.2010.12.012
Zhou X, Tian Z, Wang Y, Li W (2010) Effect of treatment with probiotics as water additives on tilapia (Oreochromis niloticus) growth performance and immune response. Fish Physiol Biochem 36(3):501–509. https://doi.org/10.1007/s10695-009-9320-z
Zhou Y, Yang H, Hu H, Liu Y, Mao Y, Zhou H, Xu X, Zhang F (2006) Bioremediation potential of the macroalga Gracilaria lemaneiformis (Rhodophyta) integrated into fed fish culture in coastal waters of north China. Aquaculture 252(2):264–276. https://doi.org/10.1016/j.aquaculture.2005.06.046
Acknowledgements
This work was funded by a Grant from Shahid Chamran University of Ahvaz Research Council (Grant No: 27176, 1393.3.2). The authors of this research followed instructions of the university in Iran and performed experiments based on Ethical Guideline of laboratory animals.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
All institutional and national guidelines for the care and use of laboratory animals were followed.
Conflict of Interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Mohammadian, T., Alishahi, M., Tabandeh, M.R. et al. Changes in Immunity, Expression of some Immune-Related Genes of Shabot Fish, Tor grypus, Following Experimental Infection with Aeromonas hydrophila: Effects of Autochthonous Probiotics. Probiotics & Antimicro. Prot. 10, 616–628 (2018). https://doi.org/10.1007/s12602-017-9373-8
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12602-017-9373-8