
Abstract
In the cystic fibrosis transmembrane conductance regulator (CFTR) chloride channel, lyotropic anions with high permeability also bind relatively tightly within the pore. However, the location of permeant anion binding sites, as well as their relationship to anion permeability, is not known. We have identified lysine residue K95 as a key determinant of permeant anion binding in the CFTR pore. Lyotropic anion binding affinity is related to the number of positively charged amino acids located in the inner vestibule of the pore. However, mutations that change the number of positive charges in this pore region have minimal effects on anion permeability. In contrast, a mutation at the narrow pore region alters permeability with minimal effects on anion binding. Our results suggest that a localized permeant anion binding site exists in the pore; however, anion binding to this site has little influence over anion permeability. Implications of this work for the mechanisms of anion recognition and permeability in CFTR are discussed.
Similar content being viewed by others
Introduction
Cystic fibrosis is caused by loss-of-function mutations in the cystic fibrosis transmembrane conductance regulator (CFTR) [1]. CFTR is a member of the ATP-binding cassette family of membrane proteins and functions as a phosphorylation-regulated, ATP-gated anion channel in the apical membrane of many different epithelial cell types [2]. The anion-selective channel pore is formed from a subset of CFTR’s 12 transmembrane (TM) α-helices [3].
As with most anion channels, CFTR shows only a moderate level of ion selectivity, such that its ability to discriminate between different ions appears much less than, for example, highly selective Ca2+, K+ and Na+ channels [4]. In fact, not only Cl− but also other small anions such as HCO3 −, SCN−, I− and glutathione may be physiological substrates of CFTR-mediated transport [3]. Ion selectivity in channels can be defined in a number of different ways. For example, ion permeability considers the relative probability that a specific ion species will pass through the channel pore, whereas ion binding considers the relative affinity of ion:pore interactions. In very simple terms, permeability selectivity is determined by how easily an ion will enter the channel pore, and binding selectivity is determined by how tightly it will bind inside the pore [5, 6]. In some channel types these two facets of the ion recognition process may be interrelated; for example, in some cation channels high permeability of the physiological substrate ion is thought to be ensured by selective binding of that ionic species [6–9].
The most commonly reported facet of ion selectivity is permeability selectivity, and among small monovalent anions, CFTR shows a so-called “lyotropic” or Hofmeister anion permeability sequence [10, 11]. Lyotropic selectivity suggests that relative permeability is inversely related to the anion free energy of hydration [12]. As a result, in CFTR lyotropic anions that lose their waters of hydration relatively easily—such as SCN− and Au(CN) −2 —tend to show the highest relative permeabilities [3]. This lyotropic anion permeability selectivity pattern is, in fact, common to most anion channels [13, 14], perhaps hinting at some common factor influencing anion relative permeability in unrelated anion channel proteins.
Lyotropic anions with high permeability also appear to bind relatively tightly within the pore of CFTR and other anion channels [3, 13, 14], suggesting that these channels show similar patterns of anion binding and anion permeability selectivity. One manifestation of this tight anion binding is that low concentrations of highly lyotropic anions, including SCN− and Au(CN) −2 , can block Cl− permeation through the CFTR channel [3, 13, 15–17]. However, the relationship between binding and permeability is not clear. For example, does tight binding of specific ions result in their high permeability, as is the case, for example, in highly selective Ca2+ channels [7]? Or are tight binding and high permeability the result of separate kinds of anion-pore interactions?
Most mutations in the TMs of CFTR have minimal effects on relative anion permeabilities [3]. However, mutations at one site—F337 in TM6—are associated with major changes in anion permeability ratios [18]. Specifically, mutations that reduce side-chain volume (F337A, F337S) disrupt the relationship between the anion permeability and anion free energy of hydration [18]. This led to the suggestion that F337 contributes to a localized anion “selectivity filter” in the narrow region of the pore [3]. In contrast, the location of sites inside the pore that result in tight binding of lyotropic anions are not known. Larger, impermeant anions also block Cl− permeation through the open CFTR pore, and they appear to do so predominantly by binding to a site within a relatively wide inner vestibule of the pore that is readily accessed from the cytoplasm [19]. In particular, a positively charged amino acid side chain, K95 in TM1, appears to be an important determinant of the open channel blocker binding site [19–22]. Blocker binding to this site can further be strengthened by introduction of additional positive charges at other, nearby pore-lining sites in TMs 6 and 12 [22–24]. Additional blocker binding sites may also exist, for example, in the outer vestibule of the pore [26, 27].
In the present study we compared the effects of mutations at the cytoplasmic blocker binding site (at K95, I344, V345 and S1141) and narrow pore region (F337) on lyotropic anion binding and anion relative permeability in open CFTR channels. Mutations at the blocker binding site drastically alter the apparent affinity of binding of lyotropic anions (determined by their ability to block Cl− permeation) while having minimal effects on anion permeability. In contrast, the F337A mutation disrupts the normal lyotropic anion permeability pattern as expected [18] but has much lesser effects on lyotropic anion binding. These results indicate that the apparent relationship between high lyotropic anion permeability and tight lyotropic anion binding can be disrupted by mutations within the pore, suggesting that these different facets of the anion permeation mechanism are not strongly interdependent but instead have distinct and independent molecular bases.
Materials and methods
Experiments were carried out on baby hamster kidney cells transiently transfected with CFTR, as described in detail previously [22]. In order to isolate anion effects on Cl− permeation from effects on channel gating [17, 24], experiments were carried out on constitutively active E1371Q-CFTR channels [22, 24]. Additional mutations were introduced into this E1371Q background using the QuikChange site-directed mutagenesis system (Agilent Technologies, Santa Clara, CA, USA) and verified by DNA sequencing.
Macroscopic E1371Q-CFTR current-voltage (I–V) relationships were recorded using depolarizing voltage ramps during patch clamp recordings from inside-out membrane patches, as described in detail recently [24]. Current traces were filtered at 100 Hz using an 8-pole Bessel filter, digitized at 1 kHz, and analyzed using pCLAMP software (Molecular Devices, Sunnyvale, CA, USA). For experiments with channel blockers (Figs. 1, 2, 3, 4), both intracellular (bath) and extracellular (pipette) solutions contained (mM): 150 NaCl, 2 MgCl2, 10 N-tris[hydroxymethyl]methyl-2-aminoethanesulfonate (TES), pH 7.4. For experiments at different cytoplasmic pH (Fig. 3a–c), the pH buffer in the intracellular solution was changed from TES to either 2-(N-morpholino)ethanesulfonic acid (MES) for pH 5.5 or [(2-hydroxy-1,1-bis[hydroxymethyl]ethyl)amino]-1-propanesulfonic acid (TAPS) for pH 9.0. Different concentrations of blockers (KAu(CN)2, NaSCN, KC(CN)3) were applied directly to the cytoplasmic face of inside-out patches from stock solutions made up in normal intracellular solution. Blocker concentration-inhibition relationships were fitted by the equation:
where [B] is the blocker concentration and K D the apparent blocker dissociation constant.

Block by intracellular Au(CN)2 − is weakened in K95Q/E1371Q channels. Example macroscopic I–V relationships for E1371Q (a) and K95Q/E1371Q (b) CFTR channels recorded before (control) and after the addition of Au(CN) −2 to the intracellular (bath) solution at the concentrations stated. c Mean fraction of control current remaining after addition of different concentrations of Au(CN) −2 at a membrane potential of −100 mV for both these channel constructs. Data have been fitted as described in “Materials and methods.” d Mean K D values obtained from such fits at different membrane potentials. Mean K D values were significantly different between the two channel constructs at all voltages examined (P < 0.005). Mean of data from 7 to 8 patches

Block by intracellular SCN− and C(CN) −3 is weakened in K95Q/E1371Q channels. Example macroscopic I–V relationships for E1371Q (a) and K95Q/E1371Q (b) CFTR channels recorded before (control) and after the addition of 10 mM SCN− to the intracellular (bath) solution. c Mean K D values for SCN− block for these channel constructs, obtained as described for Au(CN) −2 in Fig. 1. Mean K D values were significantly different between the two channel constructs at all voltages examined (P < 0.05). d Mean K D values for C(CN) −3 block, obtained from similar experiments (not illustrated). Mean K D values were significantly different between the two channel constructs at all voltages examined (P < 0.002). Mean of data from 3 to 5 patches

Strength of Au(CN) −2 block is dependent on the number of positive charges in the pore inner vestibule. a Example macroscopic I–V relationships for K95H/E1371Q CFTR channels recorded using bath solutions at pH 5.5 (left) or pH 9.0 (right, different patch). Currents were recorded before (control) and after the addition of 1 mM Au(CN) −2 to the intracellular (bath) solution. b Mean K D values for Au(CN) −2 block of K95H/E1371Q at these two different pHs, obtained as described for Au(CN) −2 in Fig. 1. Mean K D values were significantly different between the two pHs at all voltages examined (P < 0.01). c Relationship between the observed K D values for Au(CN) −2 block (at −100 mV) and bath solution pH in K95H/E1371Q and E1371Q. d Example macroscopic I–V relationships for I344K/E1371Q (left) and K95Q/I344K/E1371Q (right) CFTR channels recorded before (control) and after the addition of a low concentration (10 μM) of Au(CN) −2 to the intracellular (bath) solution. e Mean K D values for Au(CN) −2 block for these channel constructs, as well as the additional positive charge mutants V345K/E1371Q and S1141K/E1371Q, obtained as described in Fig. 1. f Relationship between the observed K D values for Au(CN) −2 block (at −100 mV) and the expected number of fixed positive charges in the pore inner vestibule in different channel constructs. K95H0 refers to the unprotonated form of the histidine side chain (as expected at pH 9.0) and K95H+ the protonated form (at pH 5.5). Asterisks indicate a significant difference from E1371Q (*P < 0.05, **P < 0.0001). Mean of data from 3 to 8 patches

Block of F337A/E1371Q channels by intracellular lyotropic permeant anions. a, b Example macroscopic I–V relationships for F337A/E1371Q CFTR channels recorded before (control) and after the addition of Au(CN) −2 (1 mM) or SCN− (10 mM) to the intracellular (bath) solution. c Mean K D values for Au(CN) −2 , SCN−, and C(CN) −3 (estimated at −100 mV as described in Figs. 1 and 2) compared in E1371Q, K95Q/E1371Q, and F337A/E1371Q. Asterisks indicate a significant difference from E1371Q (P < 0.01). Mean of data from 3 to 8 patches
For anion permeability experiments (Fig. 5), NaCl in the bath solution was replaced by NaBr, NaF, NaNO3 or NaSCN. The macroscopic current reversal potential (V REV) was estimated by fitting a polynomial function to the I–V relationship and used to calculate the permeability of different anions (X−) relative to that of Cl− (P X/P Cl) according to:
where [X]i and [Cl]i (intracellular concentrations) are 150 and 4 mM, respectively, and [Cl]o (extracellular) is 154 mM; R, T and F have their usual thermodynamic meanings. For presentation of current traces (Fig. 5a), currents from different patches were normalized to a common scale in order to emphasize changes in the current reversal potential with different test anions; the actual amplitude of the currents under these conditions is dependent on the number of channels present in the inside-out membrane patch and so contains no relevant information.

Anion permeability in different CFTR channel variants. a Normalized I–V relationships for the CFTR variants indicated, recorded with Cl−-containing extracellular solutions and intracellular solutions containing F−, Cl−, Br−, NO3 − or SCN− as indicated. Note that the range of current reversal potentials was greatly reduced in F337A/E1371Q, suggesting a relative loss of permeability selectivity in this mutant. b Mean P X/P Cl values calculated from current reversal potential measurements under these conditions as described in “Materials and methods.” Asterisks indicate a significant difference from E1371Q (*P < 0.05; **P < 0.002). c Relationship between P X/P Cl and anion free energy of hydration (G h, taken from Marcus [31] ). Note that the normal lyotropic relationship between relative permeability and G h is greatly reduced in F337A/E1371Q but retained in K95Q/E1371Q and I344K/E1371Q. Mean of data from 3 to 8 patches
Experiments were carried out at room temperature, 21–24 °C. Values are presented as mean ± SEM. For graphical presentation of mean values, error bars represent SEM, and where no error bars are shown SEM is smaller than the size of the symbol. Tests of significance were carried out using Student’s two-tailed t test, with P < 0.05 being considered statistically significant. All chemicals were from Sigma-Aldrich (Oakville, ON, Canada) or Strem Chemicals (Newburyport, MA, USA).
Results
Intracellular Au(CN) −2 ions cause a voltage-dependent block of open CFTR channel pores [17], a finding that is recapitulated in constitutively open E1371Q channels (Fig. 1). The voltage-dependent apparent K D under these conditions ranged from 38.5 ± 11.5 μM (n = 7) at −100 mV to 306.3 ± 67.6 μM (n = 7) at +60 mV, similar to results obtained previously for wild-type CFTR [17]. However, the blocking effects of Au(CN) −2 were dramatically weakened when a key positive charge in the pore inner vestibule was neutralized by mutagenesis (Fig. 1b–d). Thus, the mean K D was increased between 92-fold (at −100 mV) and 19-fold (at +60 mV) in K95Q/E1371Q (Fig. 1d). Similar weakening of block in K95Q/E1371Q was also observed for other highly lyotropic permeant anions SCN− and C(CN) −3 (Fig. 2). These results suggest that the positive charge at K95 plays an important role in permeant anion binding inside the pore.
To investigate the role of positive charge in permeant anion binding more directly and independently of other amino acid side chain properties, we mutated K95 to histidine and then toggled the side-chain charge by using intracellular solutions with different pH values. As shown in Fig. 3a and b, block of K95H/E1371Q by intracellular Au(CN) −2 was drastically stronger at pH 5.5 compared to pH 9.0, with the mean K D at −100 mV being increased 244-fold at the more alkaline pH. In contrast, Au(CN) −2 block of E1371Q was not pH-dependent (Fig. 3c).
We also used mutagenesis to increase the number of positive charges available to interact with Au(CN) −2 ions from one (as in wild type) to two. Previously it has been shown that a positively charged side chain introduced at different positions in TM6 (I344, V345) and TM12 (S1141) can substitute for the endogenous K95 side chain in interacting with cytoplasmic open channel blocking anions [22, 23], suggesting that these residues have side chains that are located close to K95 in the inner vestibule of the open channel pore [19]. Consistent with this, mutation of any of these three residues to lysine (I344K, V345K, S1141K) led to a significant strengthening of Au(CN) −2 block (Fig. 3d–f), with mean K D values at −100 mV being reduced by 18-fold (I344K/E1371Q), 17-fold (V345K/E1371Q) and 7-fold (S1141K/E1371Q), respectively. “Moving” the key positive charge from TM1 to TM6 by mutagenesis (in the K95Q/I344K/E1371Q mutant) resulted in Au(CN) −2 block that was intermediate in strength between E1371Q and I344K/E1371Q (Fig. 3e–f). Overall, these results supported a model in which the strength of Au(CN) −2 binding in the pore is directly related to the number of positive charges lining the pore inner vestibule and available to interact with Au(CN) −2 ions inside the pore (Fig. 3f).
As well as showing tight binding of lyotropic permeant anions, CFTR also shows a lyotropic anion permeability sequence (see “Introduction”). This permeability sequence is disrupted by mutations at the putative narrow region of the pore, located more extracellularly in the pore than K95, in particular F337A and F337S (see “Introduction”). As shown in Fig. 4, the F337A mutation had only a minor effect on binding of lyotropic Au(CN) −2 , SCN− and C(CN) −3 ions when compared to the K95Q mutation, suggesting that these anions can still bind relatively tightly in the pore even when lyotropic permeability selectivity is compromised.
To investigate anion permeability directly, we used intracellular (bath) solutions containing different test anions. As shown in Fig. 5, macroscopic current reversal potential measurements with different anions present in the intracellular solution gave a permeability selectivity sequence SCN− > NO3 − > Br− > Cl− > F− for E1371Q-CFTR, with very similar relative permeabilities for those previously reported for wild type CFTR under similar conditions [11]. Consistent with the proposed role of F337 in controlling lyotropic anion permeability, the permeability of all anions tested in F337A/E1371Q was significantly changed relative to E1371Q (Fig. 5b), with the permeability selectivity sequence being changed to NO3 − ≥ SCN− ≥ Cl− > Br− > F−. Again, relative permeability values for F337A/E1371Q were similar to those reported previously for F337A [18]. In contrast, neither K95Q nor I344K altered the permeability selectivity sequence; in fact, the only significant difference between either K95Q/E1371Q or I344K/E1371Q compared to E1371Q was a small increase in P F/P Cl in K95Q/E1371Q.
Discussion
A key piece of evidence for permeant anion binding within the CFTR pore is the finding that some permeant anions—such as Au(CN) −2 , SCN− and C(CN) −3 —can inhibit Cl− permeation through the channel [13, 15–17]. In the open CFTR channel pore, these anions block Cl− currents with apparent K Ds in the range of tens of micromolar to a few millimolar (Figs. 1, 2), consistent with relatively tight binding inside the permeation pathway. Removal of a positive charge in the pore inner vestibule by mutagenesis (in the K95Q mutant) or by increasing pH (in K95H) dramatically increases the apparent K D (Figs. 1, 2, 3, 4), suggesting that this positive charge is required for tight binding of permeant anions. In fact, since Au(CN) −2 , SCN− and C(CN) −3 are permeant anions and presumably have access to the entire pore, it is possible that the residual block observed in K95Q and in K95H (at pH 9.0) reflects interactions with a different part of the pore. For example, permeant anions may also bind to site(s) near the outer mouth of the pore [25–27]. We propose that permeant anions bind relatively tightly in the inner vestibule of the pore, close to the positively charged side chain of K95, in much the same way as has been described previously for larger, impermeant blocking ions [19–21]. The idea that permeant anions bind in this region is supported by the finding that apparent binding affinity can be increased by additional positive charges introduced into sites that are thought to line a similar region of the open channel pore (I344, V345, S1141; Fig. 3d–f).
Irrespective of the exact location of permeant anion binding site(s), our results cast new light on the relationship between anion binding and anion permeability in CFTR. Thus, mutations that dramatically decrease (K95Q) or increase (I344 K) the strength of permeant anion binding inside the pore had only minimal effects on anion relative permeability (Fig. 5). This suggests that binding to this site is not crucial to the anion permeability selectivity process. Conversely, a mutation that is known to have a strong effect on relative permeability—F337A (Fig. 5)—had relatively minor effects on permeant anion binding (Fig. 4). This suggests that CFTR permeability selectivity is not the result of selective ion binding. Overall, although wild-type CFTR demonstrates lyotropic anion permeability selectivity and lyotropic anion binding, these two apparently parallel pore functional properties are independent since the apparent relationship between the two can be broken by mutagenesis; mutations at K95 (and I344) alter binding without affecting permeability, whereas mutations at F337 disrupt permeability selectivity with a more modest effect on binding. We propose that whereas tight binding occurs mainly in the pore inner vestibule, anion permeability is determined at a narrow pore region located closer to the outer end of the pore (Fig. 6). Since lyotropic selectivity patterns are thought to reflect differences in anion:water and anion:protein interaction energies [13, 28], these similar patterns occurring at distinct regions of the CFTR channel pore and controlling different permeation properties may simply reflect different physical processes that are influenced by anion:water interactions.

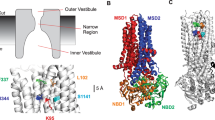
Molecular bases for CFTR channel lyotropic anion binding and lyotropic anion permeability. a Atomic homology model of CFTR [21] indicating the approximate extent of the membrane spanning domains (MSDs), intracellul ar loops (ICLs) and cytoplasmic nucleotide binding domains (NBDs). Transmembrane helices TM1 and TM6, the sites of mutations used in the present study, are highlighted in red and blue, respectively. b Detailed view of TMs 1 and 6 in this same homology model, indicating the relative location of amino acid side chains proposed to contribute to the lyotropic anion binding site in the pore inner vestibule (“energy well”; K95, I344) and to the lyotropic anion permeability selectivity filter in the pore narrow region (“energy barrier”; F337). c Cartoon model showing the proposed overall architecture of the pore [3] and indicating the approximate locations of these same amino acids within this overall cartoon model. In both a and b, CFTR model structure was based on coordinates provided by Dalton et al. [21] and visualized using PyMol (Schrödinger, LLC, Portland, OR, USA) (color figure online)
Lyotropic anion binding and anion permeability selectivities of CFTR have previously been modeled as energy wells (reflecting binding) and energy barriers (reflecting anion entry into the pore), respectively [13]. Using such a formulism, our results suggest that the pore inner vestibule (close to K95 and I344) represents a key energy well and that the depth of this well can be increased or decreased by manipulations that change the number of positively charged amino acid side chains in this part of the pore (Fig. 6). Conversely, the narrow pore region around F337 is an important energy barrier (Fig. 6). Our results therefore suggest that well depths and barrier heights can vary more or less independently to selectively alter anion binding or anion permeability.
Does K95 also contribute to Cl− ion binding inside the pore? Chloride itself is a mildly lyotropic anion that presumably experiences similar forces to other permeant anions as it passes through the pore. Removing the positive charge at this position does not have a major impact on anion permeability selectivity (Fig. 5) but it is known to drastically reduce single-channel Cl− conductance [22, 23, 29]. Initially the role of the positive charge at K95 was described as electrostatic attraction of cytoplasmic Cl− ions to the pore [20, 22]. However, as pointed out by Hwang and Kirk [30], since removal of this positive charge dramatically decreases both Cl− efflux and Cl− influx, a purely electrostatic attractive effect is unlikely. An alternative possibility is that Cl− binding in the pore inner vestibule, supported by the positive charge of K95, is essential to normal Cl− conductance. Ion binding inside ion channel pores is often associated with high flux rates, especially in multi-ion pores that can hold more than one permeant ion at a time [7–9]. We speculate that permeant anion binding in the pore inner vestibule is important for optimizing Cl− conductance rather than for controlling anion permeability selectivity.
References
Lubamba B, Dhooghe B, Noel S, Leal T (2012) Cystic fibrosis: insight into CFTR pathophysiology and pharmacotherapy. Clin Biochem 45:1132–1144
Frizzell RA, Hanrahan JW (2013) Physiology of epithelial chloride and fluid secretion. Cold Spring Harb Perspect Med 3:a009498
Linsdell P (2014) Functional architecture of the CFTR chloride channel. Mol Membr Biol 31:1–16
Hille B (2001) Ion channels of excitable membranes, 3rd edn. Sinauer Associates, Sunderland
Eisenman G, Horn R (1983) Ionic selectivity revisited: the role of kinetic and equilibrium processes in ion permeation through channels. J Membr Biol 76:197–225
Horn R, Roux B, Åqvist J (2014) Permeation redux: thermodynamics and kinetics of ion movement through potassium channels. Biophys J 106:1859–1863
Sather WA, McCleskey EW (2003) Permeation and selectivity in calcium channels. Annu Rev Physiol 65:133–159
Roux B (2005) Ion conduction and selectivity in K+ channels. Annu Rev Biophys Biomol Struct 34:153–171
Armstrong CM (2007) Life among the axons. Annu Rev Physiol 69:1–18
Tabcharani JA, Linsdell P, Hanrahan JW (1997) Halide permeation in wild-type and mutant cystic fibrosis transmembrane conductance regulator chloride channels. J Gen Physiol 110:341–354
Linsdell P, Hanrahan JW (1998) Adenosine triphosphate-dependent asymmetry of anion permeation in the cystic fibrosis transmembrane conductance regulator chloride channel. J Gen Physiol 111:601–614
Wright EM, Diamond JM (1977) Anion selectivity in biological systems. Physiol Rev 57:109–157
Liu X, Smith SS, Dawson DC (2003) CFTR: what’s it like inside the pore? J Exp Zool 300:69–75
Linsdell P (2006) Mechanism of chloride permeation in the cystic fibrosis transmembrane conductance regulator chloride channel. Exp Physiol 91:123–129
Dawson DC, Smith SS, Mansoura MK (1999) CFTR: mechanism of anion conduction. Physiol Rev 79(Suppl 1):S47–S75
Linsdell P (2001) Thiocyanate as a probe of the cystic fibrosis transmembrane conductance regulator chloride channel pore. Can J Physiol Pharmacol 79:573–579
Linsdell P, Gong X (2002) Multiple inhibitory effects of Au(CN) −2 ions on cystic fibrosis transmembrane conductance regulator Cl− channel currents. J Physiol 540:29–38
Linsdell P, Evagelidis A, Hanrahan JW (2000) Molecular determinants of anion selectivity in the cystic fibrosis transmembrane conductance regulator chloride channel pore. Biophys J 78:2973–2982
Linsdell P (2014) Cystic fibrosis transmembrane conductance regulator chloride channel blockers: pharmacological, biophysical and physiological relevance. World J Biol Chem 5:26–39
Linsdell P (2005) Location of a common inhibitor binding site in the cytoplasmic vestibule of the cystic fibrosis transmembrane conductance regulator chloride channel pore. J Biol Chem 280:8945–8950
Dalton J, Kalid O, Schushan M, Ben-Tal N, Villà-Freixa J (2012) New model of cystic fibrosis transmembrane conductance regulator proposes active channel-like conformation. J Chem Inf Model 52:1842–1853
Zhou J-J, Li M-S, Qi J, Linsdell P (2010) Regulation of conductance by the number of fixed positive charges in the intracellular vestibule of the CFTR chloride channel pore. J Gen Physiol 135:229–245
El Hiani Y, Linsdell P (2012) Tuning of CFTR chloride channel function by location of positive charges within the pore. Biophys J 103:1719–1726
Linsdell P (2014) State-dependent blocker interactions with the CFTR chloride channel: implications for gating the pore. Pflügers Arch 466:2243–2255
Gong X, Linsdell P (2003) Molecular determinants and role of an anion binding site in the external mouth of the CFTR chloride channel pore. J Physiol 549:387–397
Zhou J-J, Fatehi M, Linsdell P (2007) Direct and indirect effects of mutations at the outer mouth of the cystic fibrosis transmembrane conductance regulator chloride channel pore. J Membr Biol 216:129–142
Zhou J-J, Fatehi M, Linsdell P (2008) Identification of positive charges situated at the outer mouth of the CFTR chloride channel pore. Pflügers Arch 457:351–360
Collins KD (1997) Charge density-dependent strength of hydration and biological structure. Biophys J 72:65–76
Ge N, Muise CN, Gong X, Linsdell P (2004) Direct comparison of the functional roles played by different membrane spanning regions in the cystic fibrosis transmembrane conductance regulator chloride channel pore. J Biol Chem 279:55283–55289
Hwang T-C, Kirk KL (2013) The CFTR ion channel: gating, regulation, and anion permeation. Cold Spring Harb Perspect Med 3:a009498
Marcus Y (1997) Ion properties. Marcel Dekker, New York
Acknowledgments
We would like to thank Christina Irving and Dr. Yassine El Hiani for their assistance with this work. Supported by the Canadian Institutes of Health Research and Cystic Fibrosis Canada.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
About this article

Cite this article
Rubaiy, H.N., Linsdell, P. Location of a permeant anion binding site in the cystic fibrosis transmembrane conductance regulator chloride channel pore. J Physiol Sci 65, 233–241 (2015). https://doi.org/10.1007/s12576-015-0359-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12576-015-0359-6