
Abstract
We aimed to evaluate the effect of social interaction on learning in juvenile jack mackerel Trachurus japonicus. We first compared the learning process between single fish and groups of fish. Reward-conditioned learning was established in eight trials in both treatments, whereas individuals in the group treatment responded to stimuli more frequently in the feeding area than in the single fish. This implies that information about the feeding area was shared in the group and pursuing other individuals gave them a behavioral advantage for feeding. We then investigated whether information on the feeding area can be transmitted through observation of other individuals in aligned tanks. Fish in the control group required six trials to be conditioned to aeration stimuli and feeding location, whereas those in the observation treatment required only three trials for this learning. This result implies that information on the feeding area was transmitted through visual observations. The present research suggests that sharing and transmission of information occur in schools of jack mackerel. Schooling behavior would thus enable optimization of the foraging behavior in this species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
About 50% of teleost fishes in the world form schools at least for a part of their life history [1]. Schooling may well provide substantial advantages for fishes, i.e., defense by intimidating or confusing predators [2], reducing energy requirements for swimming [1], and increasing the chance of reproduction [3].
Schooling behavior would also be useful for sharing social information within a group of fish [3, 4]. For example, chinook salmon Oncorhynchus tshawytscha use geomagnetic and olfactory cues for homing to their natal river, where grouping behavior is suggested to improve accuracy for homing by balancing out errors by individuals [5]. Schooling would also enable the transmission of information between individuals by observational learning, which is the learning through observation of other individuals [6–8]. Observational learning is likely to occur more often in schooling fish than between solitary individuals.
Learning in animals plays an important role in adaptation to changes in environmental conditions and thus enhances survival potential. Fish would acquire survival skills by learning in various life history contexts [9, 10], such as predator avoidance [11, 12], homing [13, 14], and foraging and capture of food items [15, 16]. Therefore, quick and effective learning of such information would be advantageous.
Jack mackerel Trachurus japonicus is an obligatory schooling fish which starts to form schools at ca. 12 mm in standard length (SL) (Masuda R. et al., unpublished data, 2009). Jack mackerel juveniles have learning capability of a specific feeding area in an experimental tank even when they are treated in solitary conditions, indicating that learning would play a substantial role for the adaptation to environmental conditions experienced in this species [17]. They form large schools during their life history and therefore they have opportunities to acquire information from conspecifics when they are schooling. For fish showing such behavior, the social effect of schooling for learning may be substantial.
The social effect for learning has been studied mostly in freshwater and coral reef fishes [18–21], whereas there are few studies for pelagic marine fishes on this subject. We conducted two experiments to investigate the advantages of schooling for learning using jack mackerel as a target species. Experiment 1 compared the learning capability between single fish and groups of fish to elucidate the effect of schooling on their learning. Experiment 2 examined the effect of social transmission between groups of fish.
Materials and methods
Experiment 1: Learning process in single fish and groups of fish
Jack mackerel (mean ± standard deviation, 105 ± 9 mm SL) were captured by angling at a pontoon of the Maizuru Fisheries Research Station (MFRS; 35° 49′ N, 135° 36′ E), Kyoto University. Fish were kept in 500-l black polyethylene tanks supplied with filtered sea water at a rate of 4 l/min and with strong aeration (600 ml/min). Rearing tanks were set indoors and water temperature was kept at about 20°C using a heater and thermostat from November to December 2008. The fish were fed with commercial pellets (Otohime S2, Marubeni Nisshin Feed) to saturation once a day until being used for the experiment. All the fish were kept in a tank for at least 1 month to be weaned on pellets. When fish were confirmed to forage actively on pellets near the water surface, individuals were transferred to an experimental tank and the experiment was started.
Transparent acrylic tanks (length × width × height 90 × 30 × 30 cm; Fig. 1) were set in a temperature-controlled room and were covered with black vinyl sheets. The tanks were visually divided into three even sections by marking their walls, then the central part was defined as the feeding area where pellets were dropped. Sea water was delivered into the experimental tanks from both ends and drained out from the feeding area to minimize the effect of olfactory cues from food. Tanks were gently aerated and temperature was maintained at ca. 20°C using a heater and thermostat. Tanks were separated to reduce disturbance by the experimenter using a black vinyl sheet and a PVC feeding pipe was set from above the feeding area through the sheet. A video camera (Handycam DCR-PC350, Sony) set above the experimental tank allowed recording of the behavior of fish during the experiment. Experiments were conducted either with a single fish treatment or with a group treatment consisting of four individuals. A total of 20 individuals were used for each treatment, i.e., 20 single fish and 5 groups. Fish were introduced into the experimental tanks on the day prior to the experiment, and were allowed to acclimatize overnight. A few pellets were provided prior to the experiment, and the experiment was started if the fish accepted the pellets, and by all of the four fish in the group treatment. If not, the experiment was postponed to the next day. Fish that did not feed on the next day of acclimatization were not used in further experiments. The experiments were conducted and the video camera recorded the behavior of fish soon after feeding. Introduction of a floating object made of black polyethylene plastic mesh (length × width 9 × 9 cm) into the experimental tank before feeding was used as the conditioned stimulus (CS) for reward training. One min after presenting the CS, three or four food pellets per fish were provided around the floating object through the feeding pipe as a reward stimulus. The video camera recorded the fish behavior for 2 min, from 1 min prior to the CS to 1 min after the CS. The reward was then provided. This process was defined as one trial. Five sequential trials at 5-min intervals comprised one session and four sessions at 1-h intervals comprised the whole test; therefore, the reward training was conducted for a total of 20 trials in both treatments. Fish body length was measured when fish accomplished the reward training after anesthesia, using ca. 5 ml of 2-phenoxyethanol per 10 l of seawater.

Schematic drawing of an experimental tank in experiment 1. Black floating mesh was used as a conditioned stimulus and 4–16 pellets were used as an unconditioned stimulus. The central area of the tank was defined as the feeding area
The duration of fish staying in the feeding area (stay duration) was used as the criterion for learning. The duration was measured in each 1 min of the pre- and post-signal period. In the single fish treatment, data of four randomly selected individuals were combined to make five groups of four individuals. This way the number of replications, and so the statistical power, was equalized between the two treatments. Total stay duration of all individuals was divided by the number of individuals (i.e., 4). The maximum stay duration would thus be 60 s in both treatments. The stay duration in the feeding area was log-transformed to achieve near normality and was compared between the pre- and post-signal period by paired t test in each treatment. When the stay duration between the pre- and post-signal period differed significantly in two consecutive trials, it was considered that the fish had learned the conditioning. Learning capability of single fish and groups of fish was compared by the number of trials required by the fish to learn the CS. Aggregating behavior to CS was calculated as the aggregation index, by subtracting the stay duration of pre-signal from that of post-signal for each trial. Aggregation index was subjected to two-way repeated measures analyses of variance (ANOVA) to compare the effects of treatments (single fish vs. groups of fish) during all trials (first to twentieth trial), as well as the first (first to fifth trial), second (sixth to tenth trial), third (eleventh to fifteenth trial), and fourth session (sixteenth to twentieth trial) to detect any tendency of aggregating behavior.
Experiment 2: Social transmission of learning behavior between schools
Jack mackerel (60.2 ± 6.7 mm SL) were captured from those associated with the giant jellyfish Nemopilema nomurai using a pair of hand nets while snorkeling ca. 10 km offshore area of Maizuru, Kyoto (35° 68′ N, 135° 44′ E). We used smaller individuals than those in the experiment 1 because the smaller individuals would accept pellets more readily in captivity. Collection by hand nets helped minimize accidental injury which they otherwise occasionally would suffer through angling. Fish were kept in 500-l black polyethylene tanks supplied with filtered sea water at a rate of 4 l/min and with strong aeration (600 ml/min). Rearing tanks were set inside a building and water temperature was kept at about 20°C using a heater and thermostat from November 2009 to February 2010. These fish were fed with commercial pellets (Otohime S2, Marubeni Nisshin Feed) to saturation once a day until being used for the experiment. All the fish were kept in a tank for at least 1 month to be weaned on pellets. When fish were confirmed to forage actively on pellets near the water surface, individuals were transferred to an experimental tank and the experiment was started.
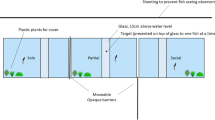
Transparent acrylic tanks (length × width × height 60 × 30 × 35 cm; Fig. 2) were set in a temperature-controlled room and were covered with black vinyl sheets except for one side. Sea water was added into the experimental tanks and drained out to keep a depth of 20 cm. Tanks were separated by a black sheet to reduce disturbance. A video camera (Dimage X1, Minolta Co., Tokyo, Japan) set above the experimental tank allowed recording of the fish behavior during the experiment.

Schematic drawing of experimental tanks in experiment 2. Aeration was used as a conditioned stimulus and 4–16 pellets were used as an unconditioned stimulus. The tanks were placed side by side, and a black board was put between the demonstrator tank and observer tank except during the observation period
The learning capability was compared between “observer fish” and “demonstrator fish” (i.e., non-observer fish). Fish were conditioned to aeration stimulus as a CS and feeding pellets as a reward stimulus. We used aeration as CS this time as opposed to a floating object because this is a more efficient stimulus (Takahashi K., unpublished data, 2010) and could reduce the total number of trials (see below). An air stone was put in the center of each tank. Aeration was remotely controlled and was turned off except during the experiment. The experiment was conducted using a group of four individuals, and six groups were used for both observer and demonstrator fish. Fish were introduced into the experimental tanks on the day prior to the experiment, and were allowed to acclimatize overnight. The tank of observer fish and that of demonstrator fish were arranged to face each other with the uncovered sides of the tanks, and a removable black board (60 × 35 cm) was placed between these two tanks except during an observation trial. A few pellets were provided prior to the experiment to confirm acclimatization, and the experiment was started if all fish in both tanks accepted them. A video camera recorded the behavior of fish as in experiment 1.
Firstly, the innate response to aeration was investigated in demonstrator fish in no-reward trials. Aeration was gently turned on to provide ca. 12 ml/min of air. Fish behavior was video recorded for 2 min, from 1 min prior to turning on the aeration to 1 min after that. This process was defined as one trial and ten trials were conducted at a minimum of 30-min intervals each for the innate response. Pellets were provided ca. 30 min before the first trial and after the tenth trial during the innate response test. For observer fish, the innate response was investigated on the day before the observation trials.
Training was started on the following 1 or 2 days in the demonstrator fish conditioned to aeration as a CS and pellets as an unconditioned stimulus. One minute after presenting aeration, ca. 20 pellets (Otohime S2, Nisshin Marubeni) divided into three times per group of fish were provided near the air stone from above the tank. Fish behavior was video recorded for 2 min, from 1 min prior to CS to 1 min after that; the reward was then provided thereafter. This process was defined as one trial and ten trials with at least 30-min intervals each comprised the whole test in a day. The training was conducted for ten trials in each group.
After completing the training of demonstrator fish, the observation trials were commenced on the following day. The black board between the tanks was removed before the observation trials, thereby fish in both tanks were visible to each other. Observation trials were conducted in the same manner as the conditioning of demonstrator fish by feeding pellets to the demonstrator fish. Ten observation trials were conducted with at least 30-min intervals a day. After ten trials, the black board was replaced and demonstrator fish were measured in body length after anesthesia. The experiment with observation fish was started on the following day with the same procedure as the demonstrator fish. The data of demonstrator fish were used as a control because of the limitation of available fish. Therefore, acclimatization duration before conditioning differed between demonstrator and observer fish (1.33 and 2.17 day on average, respectively). Brown [22] suggested that familiarity with the test environment would affect the performance of avoidance learning. However, all the fish used in experiment 2 were confirmed to feed on pellets actively, and thus should have been well acclimatized in the tank. Therefore, the difference of the timing used in conducting trials of those two groups should have had minimum effects on the learning ability. Fish body length was measured when fish accomplished the reward training after anesthesia.
The number of fish staying near the aeration stone was used as the index of innate and learned responses, the area being defined as 17 × 17 cm surrounding the aeration stone. The number of fish in this area was counted every 5 s in each 1 min of the pre- and post-signal period. This method substantially reduced the labor for analysis without reducing detectability of behavioral change compared to the one in experiment 1. To compare the tendency of the innate response in both treatments, increases of number of fish from the pre- to post-signal during the first ten trials were subjected to two-way repeated measures ANOVA to compare the effects of treatments (observer fish vs. demonstrator fish) and trials. The number of fish near the aeration stone combining the data of all individuals in each treatment was compared between the pre- and post-signal period from the first to tenth trial. Wilcoxon signed-rank test was used because these data did not achieve normality even with transformation. When the number between the pre- and post-signal period differed significantly between two consecutive trials, it was considered that the fish had accomplished the conditioning in the previous trial. Learning capability of the observer fish and demonstrator fish was compared by the number of trials required by the fish in order to accomplish the conditioning. Aggregating behavior to aeration was calculated as the aggregation index, by subtracting the stay duration of pre-signal from that of post-signal for each trial. Aggregation index during the first to tenth trial was compared by repeated measures two-way ANOVA between treatments.
Results
Experiment 1: Learning process in single fish and groups of fish
Both single and groups of fish required eight trials to accomplish the conditioning (n = 5; Fig. 3). Two-way repeated measures ANOVA of the aggregation index revealed that the effects of treatments and trials were significant, whereas that of the treatments × trials interaction was not (Table 1). The aggregation index during 20 trials increased significantly more in the group of fish than in the single fish (p < 0.05; Fig. 4). Comparing each of the four sessions, the group of fish had a significantly greater increase in the third session than single fish (p < 0.05), whereas there was no such difference in the first, second, and fourth sessions (p > 0.05; Table 1).

Average duration spent in the feeding area in the single fish and groups of fish. Symbols represent pre-signal (open circles) and post-signal period (closed circles), and asterisks indicate that the duration significantly increased from the pre- to post-signal period (p < 0.05; by paired t test). Bars indicate standard error (n = 5)

Average aggregation index in the single fish and groups of fish. Symbols represent single fish (open circles) and groups of fish (closed circles), and asterisks indicate that the increment significantly differed between single fish and groups of fish in each session (p < 0.05; by two-way repeated measures ANOVA). Bars indicate standard error (n = 5)
Experiment 2: Social transmission of learning behavior between schools
All the effects were non-significant in the innate response (treatments, p = 0.74; trials, p = 0.83; treatments × trials, p = 0.66; Table 1). To accomplish the conditioning, the demonstrator fish required six trials, and the observer fish required three trials; the latter thus learned twice as fast as the former (Fig. 5). Two-way repeated measures ANOVA of the aggregation index revealed that the effects of trials and treatments × trials interaction were significant (trials, p < 0.001; treatments × trials interaction p < 0.05; Fig. 6), whereas that of treatments was not (p > 0.05).

Average number of fish in the feeding area in the demonstrator fish and observer fish. Symbols represent pre-signal (open circles) and post-signal period (closed circles), and asterisks indicate that the number significantly increased from the pre-signal to post-signal period (p < 0.05; by Wilcoxon signed-rank test). Bars indicate standard error (n = 6)

Average aggregation index in the demonstrator fish and observer fish. Symbols represent demonstrator fish (open circles) and observer fish (closed circles). Bars indicate standard error (n = 6)
Discussion
Both single fish and groups of four individuals learned that floating objects are stimuli for a feeding area in experiment 1. Fish that typically show grouping behavior often show abnormal behavior when individuals are kept in solitary conditions. Masuda and Ziemann [23] indeed described that Pacific threadfin Polydactylus sexfilis had difficulty in learning when maintained in solitary conditions. Some individuals in the single fish treatment did not forage for pellets and were omitted from this study. In our previous study, 50 out of 90 individuals (i.e., 56%) trained in solitary conditions failed to be conditioned because of inability to forage [17]. These findings imply that jack mackerel juveniles kept as single fish may suffer substantial stress which could interact with their learning capability.
This experiment revealed that reward-conditioned learning was equally established in eight trials both in single and in groups of jack mackerel juveniles. This indicates that the number of trials required by most quick learners was equal between these two groups consisting of 20 fish for each treatment. This result suggests that learning itself would not be enhanced by schooling. In contrast to our study, Welty [18] reported that groups of 2, 4, or 8 individuals of goldfish Carassius auratus learned a feeding area more quickly than single fish. Gleason et al. [24] also indicated that five individuals of zebrafish learned an avoidance task in fewer trials than solitary fish. Although these studies suggested the possibility that the grouping behavior generates advantages for learning, these could also have resulted from followers (below as individuals showing ‘following behavior’) of quick learners.
On the other hand, the aggregation index was different between single and groups of fish, and individuals in the group treatment aggregated more in the feeding area than single fish. This would be caused by the effect of following behavior of learned individuals within the group. This tendency was clear after the eighth trial, implying that many individuals in the group treatment aggregated from the period when the first fish learned the feeding area. This result suggests that behavior to follow learned fish enhanced the ability of individuals to aggregate in the feeding area as also shown in Welty [18] and Gleason et al. [24]. Previous studies showed that following learned fish in a group enhances the feeding efficiency in pollock [25, 26]. In our study, unlearned fish in a group could get food by following a fish in the same group that had learned that floating objects are stimuli for a feeding area. As a result, the information that a floating object is a feeding area would be shared across the group members.
In fish, it is also reported that observation of other individuals enhances the efficiency of learning [6–8, 18]. Brown et al. [7] showed that fish acquired the information of feeding area through observations of fish in an adjacent tank. Experiment 2 in our study showed that the learning rate was improved by observations of other individuals. On aggregation index, there was significant interaction between treatments and trials. The result also suggests that the learning process of observer fish differed from demonstrator fish. These findings suggest that information that aeration was related to feeding was transmitted between groups via observation of fish in the adjacent tank. Jack mackerel form large groups in the wild; therefore, there would be frequent opportunities to observe other individuals within the school. Jack mackerel would efficiently acquire a behavioral advantage by observational learning in the natural environment.
Welty [18] showed that naïve goldfish acquired the route to the feeding area through only the observation of the behavior of learned fish without their own experience of the route, whereas in our experiment 2, observer fish did not show the association behavior to aeration on the first trial after observation. This discrepancy may be attributable to the personal information, defined as information attained through prior experience of the individual. The observer fish in our experiment might have learned that aeration was not an expected stimulus with foods as personal information. This is likely to differ from the social information provided by observation of the demonstrator, i.e., aeration evokes the concept of food. Indeed, Kendal et al. [27] showed in guppies that individual fish with prior experience of a feeding site ignored an alternative feeding site indicated by other individuals, suggesting that personal information is more important than social information. Furthermore, in our preliminary experiment, jack mackerel juveniles without experience of aeration before training learned that aeration indicated a feeding area in only one training trial (Takahashi K., unpublished data, 2010). Therefore, if they had not had an experience of aeration before the observational trial, the transmission of behavior might have occurred through observation in jack mackerel.
There are many potential stimuli that would be associated with feeding in the natural environment for jack mackerel; therefore, a quick response to such stimuli would increase the chance of feeding and thus enhance the survival rate. In this study, the sharing and transmission of feeding information in a school occurred through following and observation of learned fish, and these social effects facilitated the foraging efficiency of jack mackerel. From these results, we can say that schooling behavior would generate advantages that many individuals in a school can share optimum behavioral patterns. Schooling would therefore provide benefits for information gathering [28]. Ward et al. [29] showed that mosquitofish Gambusia holbrooki optimized their avoidance response to a predator model in a larger school more efficiently than as a solitary fish or in a small group, and Ward and colleagues suggested that the ‘many eyes’ effect of a large group enhanced both the speed and accuracy of their behavior. For learning in schooling fish species, ‘many eyes’ would be useful for quick learning.
The advantage of schooling behavior is also reported in learning of feeding skills, migration routes, and predator avoidance [6] in addition to feeding. For example in French grunt Haemulon flavolineatum living on a coral reef, a migration route was transmitted to naïve fish by accompanying a learned fish [21]. Schooling is also advantageous for the learning of alarm responses; Nakayama et al. [30] showed in chub mackerel Scomber japonicus juveniles that the startle response evoked by an electronic stimulus was transmitted among individuals in an adjacent tank through only visual information. Naïve zebrafish accompanied by learned individuals learned avoidance behavior from a net more quickly than unaccompanied fish [31]. Learning of predation-related information would be too costly; therefore, the sharing and transmission of the alarm information is more crucial for survival in natural environments.
Studies on the learning by fish have implications for several aspects of fisheries, e.g., it may enable the enhancement of the survival of released fish by conditioning the essential life skills in reared fish before release [32, 33]. However, training in hatcheries would not be realistic because of the cost and equipment required for the conditioning of fish. On the other hand, this study suggests that it may be sufficient to train only a subset of the fish to be released, because learned information would be shared in a school. Also, the transmission by observational learning has the possibility to reduce the cost of the conditioning procedure; indeed, red seabream Pagrus major after observational learning acquired avoidance behavior into shelter more rapidly than control fish without observational trial (Takahashi K., unpublished data, 2011). This study indicates the potential that the learning would be applicable to fisheries science, whereas there are few studies that have actually applied the learning of fish to fisheries fields [34]. In future, this kind of work will be extended to applied studies for fish learning in fisheries science, such as the behavioral improvement of reared fish.
References
Shaw E (1978) Schooling fishes. Am Sci 66:166–175
Landeau L, Terborgh J (1986) Oddity and the ‘confusion effect’ in predation. Anim Behav 34:1372–1380
Hoare DJ, Krause J (2003) Social organization, shoal structure and information transfer. Fish Fish 4:269–279
Croft DP, Krause J, Couzin ID, Pitcher TJ (2003) When fish shoals meet: outcomes for evolution and fisheries. Fish Fish 4:138–146
Quinn TP, Fresh K (1984) Homing and straying in chinook salmon (Oncorhynchus tshawytscha) from Cowlitz River Hatchery, Washington. Can J Fish Aquat Sci 41:1078–1082
Brown C, Laland KN (2003) Social learning in fishes: a review. Fish Fish 4:280–288
Brown C, Markula A, Laland KN (2003) Social learning of prey location in hatchery-reared Atlantic salmon. J Fish Biol 63:738–745
Arai T, Tominaga O, Seikai T, Masuda R (2007) Observational learning improves predator avoidance in hatchery-reared Japanese flounder Paralichthys olivaceus juveniles. J Sea Res 58:59–64
Keifer JD, Colgan PW (1992) The role of learning in fish behavior. Rev Fish Biol Fish 2:125–143
Laland KN, Brown C, Krause J (2003) Learning in fishes: from three-seconds memory to culture. Fish Fish 4:192–202
Brown GE (2003) Learning about danger: chemical alarm cues and local risk assessment in prey fishes. Fish Fish 4:227–234
Kelley JL, Magurran AE (2003) Learned predator recognition and antipredator responses in fishes. Fish Fish 4:216–226
Dodson JJ (1988) The nature and role of learning in the orientation and migratory behaviour of fishes. Environ Biol Fish 23:161–182
Odling-Smee L, Braithwaite AV (2003) The role of learning in fish orientation. Fish Fish 4:235–246
Ehlinger TJ (1989) Learning and individual variation in bluegill foraging: habitat specific techniques. Anim Behav 38:643–658
Warburton K (2003) Learning of foraging skills by fish. Fish Fish 4:203–215
Takahashi K, Masuda R, Yamashita Y (2010) Ontogenetic changes in the spatial learning capability of jack mackerel Trachurus japonicus. J Fish Biol 77:2315–2325
Welty JC (1934) Experiments in group behaviour of fishes. Physiol Zool 7:85–127
Reader SM, Kendal JR, Laland KN (2003) Social learning of foraging sites and escape routes in wild Trinidadian guppies. Anim Behav 66:729–739
Karplus I, Katzenstein R, Goren M (2006) Predator recognition and social facilitation of predator avoidance in coral reef fish Dascyllus marginatus juveniles. Mar Ecol Prog Ser 319:215–223
Helfman GS, Schultz ET (1984) Social transmission of behavioural traditions in a coral reef fish. Anim Behav 32:379–384
Brown C (2001) Familiarity with the test environment improves escape responses in the crimson spotted rainbowfish, Melanotaenia duboulayi. Anim Cogn 4:109–113
Masuda R, Ziemann DA (2000) Ontogenetic changes of learning capability and stress recovery in Pacific threadfin juveniles. J Fish Biol 56:1239–1247
Gleason PE, Weber PG, Weber SP (1977) Effect of group size on avoidance learning in zebrafish, Brachydanio rerio (Pisces: Cyprinidae). Anim Learn Behav 5:213–216
Troy AB, Clifford HR, Olla BL (1991) Social enhancement of foraging on an ephemeral food source in juvenile walleye pollock, Theragra chalcogramma. Environ Biol Fish 31:307–311
Clifford HR, Olla BL (1992) Social mechanisms facilitating exploitation of spatially variable ephemeral food patches in a pelagic marine fish. Anim Behav 44:69–74
Kendal RL, Coolen I, Laland KN (2004) The role of conformity in foraging when personal and social information conflict. Behav Ecol 15:269–277
Conradt L (2011) When it pays to share decisions. Nature 471:40–41
Ward AJW, Herbert-Read JE, Sumpter DJT, Krause J (2011) Fast and accurate decisions through collective vigilance in fish shoals. Proc Natl Acad Sci U S A 108:2312–2315
Nakayama S, Masuda R, Tanaka M (2007) Onsets of schooling behavior and social transmission in chub mackerel Scomber japonicus. Behav Ecol Sociobiol 61:1383–1390
Lindeyer CM, Reader SM (2010) Social learning of escape routes in zebrafish and the stability of behavioural traditions. Anim Behav 79:827–834
Suboski MD, Templeton JJ (1989) Life skills training for hatchery fish: social learning and survival. Fish Res 7:343–352
Brown C, Laland K (2001) Social learning and life skills training for hatchery reared fish. J Fish Biol 59:471–493
Mizra RS, Chivers DP (2000) Predator-recognition training enhances survival of brook trout: evidence from laboratory and field-enclosure studies. Can J Zool 78:2198–2208
Acknowledgments
We thank colleagues of the MFRS who assisted the collection of jack mackerel juveniles. We are also grateful to Dr. M. Sano and two anonymous reviewers for providing constructive comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Takahashi, K., Masuda, R. & Yamashita, Y. School for learning: sharing and transmission of feeding information in jack mackerel Trachurus japonicus juveniles. Fish Sci 78, 269–276 (2012). https://doi.org/10.1007/s12562-011-0454-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12562-011-0454-4