
Abstract
We examined the effect of docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) on the rates of abnormal morphology in juvenile brown sole Pseudopleuronectes herzensteini. Larvae during the D–E stages (15–24 days post hatching) were fed live food containing various amounts of DHA and/or EPA prepared using emulsified oils (DHA ethyl ester, EPA ethyl ester, and corn oil). Larvae during the F–I stages were fed Artemia enriched with a commercial diet supplement. We found that DHA and EPA promoted larval development and improved the incidence of morphological abnormalities in brown sole juveniles to a similar extent. However, DHA was more effective than EPA in preventing the appearance of morphological abnormalities in brown sole. The incidence of normal morphology was clearly improved by an increase of the DHA content in brown sole larvae at 25 days post-hatching. These results suggest that it is important to promote larval development and feed larvae with live food containing high levels of DHA during the D–E stages to prevent morphological abnormalities in brown sole juveniles.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Abnormalities in ocular-side pigmentation (pseudoalbinism) and eye position are commonly observed in hatchery-reared brown sole Pseudopleuronectes herzensteini [1]. It has recently been proposed that these abnormalities are caused by abnormal mechanisms of metamorphic asymmetry [2–4] but, to date, technological tools for preventing morphological abnormalities in brown sole have not been established. In addition, very little information is available on the correlation between essential fatty acids (EFAs) and the incidence of morphological abnormalities in brown sole. Previous research carried out by the authors has shown that the docosahexaenoic acid (DHA) requirement of brown sole larvae is approximately 0.6% on a dry weight basis (DWB) for the rotifer-feeding period and 1.6–2.8% (DWB) for the Artemia-feeding period in terms of survival and growth [5]. It has also been determined that the developmental stages when the DHA content of live food is likely to be the most effective in preventing morphological abnormalities in brown sole are the D–E stages [6].
It has recently been suggested that the effect of both DHA and eicosapentaenoic acid (EPA) on pigmentation differs between flatfish species [7–10]. Although the incidence of morphological abnormalities in brown sole is reduced by feeding larvae with Artemia enriched with commercial enrichment materials [11], it is not clear whether this preventative effect of the commercial enrichment materials is caused by DHA only or also by other nutritional factors. In addition, the effect of the contents of DHA and EPA in live food on the development of abnormal morphology of brown sole has not been investigated. Thus, in the study reported here, we have examined the effect of live foods enriched with various amounts of DHA and/or EPA on the incidence of morphological abnormalities in brown sole during the D–E stages in order to determine the effect of these compounds on normal morphology at the juvenile stage in brown sole.
Materials and methods
Live food treatments
Larval brown sole during the D–E stages [15–24 days post-hatching (dph)] were fed rotifers and Artemia nauplii in five different feed compositions (Table 1). Rotifers and Artemia nauplii were enriched with various amounts of corn oil (Wako Pure Chemical Industries, Japan), DHA ethyl ester, and EPA ethyl ester (Taiyo Yushi, Japan) to give the following experimental treatments: (1) control; (2) EPA; (3) DHA < EPA; (4) DHA > EPA. As commercial enrichment material, the rotifers and Artemia were fed “Super V12” (1 l/1000 individuals/1000 l culture medium at 20–23°C for 48 h; Chlorella Industries, Japan.) and “Super capsule” powder (70 g/t culture medium at 20°C for 17–23 h; Chlorella Industries), respectively. The purity of the corn oil and of the DHA and EPA ethyl esters was 99, 95 and 95%, respectively. The final volume of the enrichment oils was adjusted to 500 μl with corn oil, and 1000 rotifers/ml or 100–150 Artemia/ml were placed in 5-l enrichment tanks. Then, 0.05 ml of chicken egg yolk and varying amounts of corn oil, DHA, and EPA ethyl esters (Table 1) were added to the enrichment tanks after being emulsified using a household mixer and enriched at 24°C for 17–23 h.
Test fish and rearing methods
Fertilized eggs were obtained using broodstock caught on the coast of Tomakomai City, Hokkaido, in June 2006 and transported to the Hokkaido Mariculture Fisheries Station in Muroran, Hokkaido. Naturally spawned fertilized eggs were placed in a 200-l tank and incubated at 15°C until 3 dph. These hatchlings were reared in a 4000-l tank until 14 dph and fed rotifers enriched with Super V12. Experimental fish (2000 larvae at 15 dph) were then each placed in a separate tank for each treatment. The experiment was conducted in duplicate (EPA, DHA < EPA and DHA > EPA treatments) and in triplicate (control and commercial treatments). Feeding experiments using live foods for each experimental treatment were conducted during 15–24 dph. After 24 h starving at 25 dph, Artemia enriched with Super capsule powder was fed to the fish of all groups until 50 dph. Water temperature was set at 15°C. The conditions for the rearing period are shown in Table 2.
Assessments of growth, development, and morphology
Larvae were sampled from each experimental tank at 14 (n = 30), 26 (n = 20), and 50 dph (n = 15) for the measurement of body length (standard length). In addition, 20 larvae were sampled from each experimental tank at 14 and 26 dph for classification of the developmental stages. As reported previously [5], the standard established by Aritaki and Seikai [12] was used to classify developmental stages and abnormal morphology. In brief, developmental stages were classified into five categories (stage C: preflexion larva, mouth open; stage D: preflexion larva; stage E: flexion larva; stage F: postflexion larva, onset of metamorphosis; stage G: post flexion larva, early phase of metamorphosis). Approximately 400 fish from each experimental treatment were then sampled at 50 dph in order to observe the morphological types (type A: normal fish; type B + B′: pseudoalbino fish; type C: ambicolorate fish). Values for both trials were expressed as the mean.
Fatty acid analysis
The enriched rotifers and Artemia nauplii used in the experiments were sampled twice during the rearing period. For the fatty acid composition analysis, 500 larvae were sampled at 25 dph and the data pooled from each experimental treatment. These samples were stored at −80°C in a freezer prior to analysis. The fatty acid composition was analyzed as reported previously [5, 6, 11].
Statistical analysis
Average survival rate, body length, and incidence of each morphological pattern were analyzed by one-way analysis of variance (ANOVA), followed by Tukey–Kramer’s multiple comparison test using a 5% level of significance. Comparative data were analyzed after inverse sine transformation [13].
Results
Fatty acid composition of live food and larval body
The fatty acid compositions in the live food are shown in Table 3. Total lipid content was 16.8–21.9% for rotifers and 22.6–27.7% for Artemia nauplii,. The highest EPA content was found in live food of the EPA treatment, followed by the DHA < EPA treatment and the DHA > EPA treatment, in decreasing order. The EPA content in live food of the commercial treatment was 0.2% in the rotifer and 2.1% in Artemia nauplii. The DHA content in the live food was highest for the DHA > EPA treatment, and there was no significant difference between live food in the DHA < EPA and commercial treatments. The DHA and EPA contents in live foods were directly affected by the content of EPA and/or DHA ethyl esters in the enrichment material in each treatment.
Table 4 shows the results obtained for the lipid and fatty acid contents of the reared larvae at 14 (initial) and 25 dph. Total lipid content in the larvae at 14 dph was 19.5% and that in the larvae of all experimental treatments at 25 dph ranged from 17.0 to 20.5%. The EPA content of larvae in treatments EPA and DHA < EPA at 25 dph was higher than the initial values. The EPA content in larvae reared in the EPA treatment at 25 dph was the highest of all the experimental treatments, and there was no apparent difference between the EPA content of larvae in the DHA > EPA treatment and those in the commercial treatment at 25 dph. The DHA content of larvae reared in the DHA > EPA treatment was the highest of all the experimental treatments at 25 dph; in contrast, the DHA content of larvae reared in the control and EPA treatments at 25 dph was considerably decreased compared to initial value, while that in the DHA > EPA treatment only increased. There was no marked difference between the DHA content in larvae reared in the DHA < EPA treatment and that in the commercial treatment at 25 dph. The DHA/EPA ratio in the DHA > EPA treatment at 25 dph was the highest among all treatments. The DHA and EPA contents in larvae at 25 dph were clearly reflected by those in the live food.
Survival and growth
The mean value of survival rate in larvae fed control live foods was lower than those in the other treatments, although there was no significant difference among all treatments (Table 5). The body length of larvae reared in the commercial treatment was the highest at 26 dph among all experimental treatments, while that of the control treatment was the lowest; at 50 dph, the body length of larvae was highest in those reared in the DHA < EPA treatment (Table 5).
Composition of developmental stage
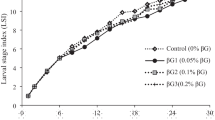
At 14 dph, all larvae in all experimental treatments had only reached the D developmental stage. At 26 dph, the development of the larvae in the control treatment was the slowest among all experimental treatments, while that of those in the commercial treatment was the fastest (Fig. 1). The development of larvae in the EPA, DHA < EPA, and DHA > EPA treatments was clearly faster than that of the control treatment larvae. There was no clear difference in terms of the composition of developmental stage between treatments EPA, DHA < EPA, and DHA > EPA (Fig. 1).

Composition of developmental stages at 26 dph in each treatment. Developmental stages (D–G) were modified from Aritaki and Seikai [12]. EPA Eicosapentaenoic acid, DHA docosahexaenoic acid
Incidence of abnormal morphology
The incidence of morphological types of each treatment is shown in Table 5. The incidence of type A in the DHA > EPA treatment was significantly higher than that in the control and EPA treatments. Conversely, the incidence of the type B + B′ morphological type in the DHA > EPA treatment was significantly lower than that in the control and EPA treatments. No significant differences in terms of the occurrence of type A and type B + B′ larval morphological types were observed between treatments DHA < EPA, DHA > EPA, and commercial. The mean value of type A morphology in the EPA treatment tended to be higher than that observed in the control treatment, although there was no significant difference between these two treatments. Type C larval morphology was infrequently observed in all of the experimental treatments.
Discussion
Survival, growth, development, and normal morphology
To date, many studies have been conducted to determine the effects of DHA and EPA as EFAs on the life-cycle parameters of various marine fish species [14–16]. For larval Japanese flounder Paralichthys olivaceus, the effects of DHA and EPA are similar as those of EFAs in terms of survival and growth [17, 18]. In our study, we found that not only DHA but also EPA improved survival and growth and that there was no clear difference between DHA and EPA as EFAs in terms of survival and growth in larval brown sole. These results are in agreement with those obtained in an earlier study in which only commercial enrichment products were used [11].
The DHA levels in live food have a marked effect on the development of larval brown sole [5, 6]. Our results demonstrate that not only DHA but also EPA in live food promoted the development of larval brown sole and that DHA and EPA were similar as EFAs on the basis of larval development. In comparison, larval development in the Commercial treatment was clearly faster than that in treatments EPA, DHA < EPA, and DHA > EPA, suggesting that also other nutritional factors, in addition to DHA or EPA, promote the development of larval brown sole.
Docosahexaenoic acid has been reported to be effective in preventing morphological abnormalities in flatfish species, including brown sole [6], marbled sole Pseudopleuronectes yokohamae [19], turbot Scophthalmus maximus [20], Japanese flounder [21], and Atlantic halibut Hippoglossus hippoglossus [22]. In our study, the fatty acid composition of the whole larval fish body reflected the composition of the live food (Tables 3, 4). The incidence of normal morphology (type A) was significantly improved by an increased DHA content in larvae at 25 dph (Tables 3, 4). The incidence of the type A morphology in the DHA > EPA treatment was significantly higher than that in the EPA and control treatments, although the mean value in the EPA treatment was 15% higher than that in control treatment. At 25 dph, the DHA content in fish fed control and EPA live foods was considerably lower than the initial value, while that in fish fed DHA-enriched live foods was the same or higher than the initial value. These findings clearly demonstrate that DHA is superior to EPA in preventing morphological abnormalities in brown sole.
It has recently been reported that the EPA contents of Artemia given to Senegal sole Solea senegalensis contributed to normal pigmentation, while DHA did not [6, 7]. Our results show that EPA is effective in preventing morphological abnormalities, although the efficacy of EPA is inferior to that of DHA. It is thus likely that in flatfish, the efficacy of DHA and EPA may vary in terms of the prevention of morphological abnormalities. Future studies in this area should focus on the EFA efficacy of morphological abnormalities.
The average incidence of type A morphology in the commercial treatment was about 12% lower than that in the DHA > EPA treatment. In terms of the DHA content of brown sole larvae at 25 dph, larvae in the commercial treatment had 0.7% less DHA content than those in the DHA > EPA treatment. These results suggest that a DHA level that is higher than that provided by the commercial diet may improve the morphological abnormalities of brown sole juveniles.
Key factors for the prevention of morphological abnormalities in brown sole
It has been suggested that the incidence of normal morphology in the juvenile brown sole is not strongly affected by survival and growth until the completion of metamorphic development [6]. We also observed a similar correlation between the incidence of normal morphology and survival and growth.
The incidence of normal morphology in flatfish species may be affected by the rate of development [12, 23]. Studies on this topic have been carried out using DHA-enriched live foods [12, 23]. In a previous study, we also reported that normal morphology and the progression of larval development of brown sole is promoted by feeding larvae live foods containing high level of DHA during the D–E stages—the so-called DHA window [6]. The results of our study demonstrate that DHA and EPA promoted larval development in a similar fashion and improved the incidence of morphological abnormalities in brown sole, although the effect of DHA on the prevention of morphological abnormalities in brown sole was superior to that of EPA. Therefore, we conclude that the two important factors affecting the normal morphology of brown sole are the promotion of larval development and the feeding of live foods containing high levels of DHA during the DHA window in larval brown sole.
In Japanese flounder, histological studies found that the retro-orbital vesicle and the pseudomesial bar are essentially important components for right eye migration [24]. The quality of the live food affects ossification and eye migration in Atlantic halibut larvae [25]. Moreover, Hamre and Harboe [26] reported that feeding Artemia enriched with high n-3 highly unsaturated fatty acids may improve the impaired eye migration. However, the relationship between histological development during eye migration and the nutrition of metamorphosing larvae has never been investigated in brown sole. Further research is necessary to identify the mechanisms underlying how DHA is utilized for the normal left–right asymmetrical development during metamorphosis.
References
Saotome K, Aritaki M (1988) Magarei jinkoushubyou no taisyokuijyou to keitaiijyou [Abnormalities of body colour and eye position in hatchery-reared brown sole (Pseudopleuronectes herzensteini) juveniles]. Saibai Giken 17:9–17 (in Japanese)
Seikai T (2005) Expression of anomalous pigmentation in flatfishes caused by wavered asymmetry. Nippon Suisan Gakkaishi 71:996–997 (in Japanese)
Aritaki M (2008) Studies on the morphological abnormalities related to metamorphosis and its prevention for flatfishes. Nippon Suisan Gakkaishi 74:772–775 (in Japanese)
Tagawa M, Aritaki M (2005) Production of symmetrical flatfish by controlling the timing of thyroid hormone treatment in spotted halibut Verasper variegatus. Gen Comp Endocrinol 141:184–189
Satoh N, Takeuchi T (2009) Docosahexaenoic acid requirement of larval brown sole Pseudopleuronectes herzensteini. Nippon Suisan Gakkaishi 75:28–37 (in Japanese with English abstract)
Satoh N, Takeuchi T (2009) Estimation of sensitive period for the abnormal morphology in hatchery-reared brown sole Pseudopleuronectes herzensteini fed live food enriched with docosahexaenoic acid. Fish Sci (in press)
Reitan KI, Rainuzzo JR, Olsen Y (1994) Influence of lipid composition of live feed on growth, survival and pigmentation of turbot larvae. Aquacult Int 2:33–48
Hamre K, Harboe T (2008) Critical levels of essential fatty acids for normal pigmentation in Atlantic halibut (Hippoglossus hipoglossus L.) larvae. Aquaculture 227:101–108
Villalta M, Estevez A, Bransden MP, Bell JG (2005) The effect of graded concentrations of dietary DHA on growth, survival and tissue fatty acid profile of Senegal sole (Solea senegalensis) larvae during the Artemia feeding period. Aquaculture 249:353–365
Villalta M, Estevez A, Bransden MP, Bell JG (2007) Effects of dietary eicosapentaenoic acid on growth, survival, pigmentation and fatty acid composition in Senegal Sole (Solea senegalensis) larvae during the Artemia feeding period. Aquacult Nutr 14:232–241
Satoh N, Fujioka T, Shimizu Y, Takeuchi T (2006) Effect of live food enrichment on survival, growth, pigmentation and starvation resistance in larval brown sole (Peudopleuronectes herzensteini). Aquacult Sci 54:305–312 (in Japanese with English abstract)
Aritaki M, Seikai T (2004) Temperature effects on early development and occurrence of metamorphosis-related morphological abnormalities in hatchery-reared brown sole Pseudopleuronectes herzensteini. Aquaculture 240:517–530
Yamada S, Kitada S (2003) Seibutsushigen toukeigaku (Statistics of fisheries resources). Seizando, Tokyo, pp 93–121 (in Japanese)
Watanabe T (1993) Importance of docosahexaenoic acid in marine larval fish. J World Aquacult Soc 24:152–161
Takeuchi T (1997) Essential fatty acid requirements of aquatic animals with emphasis on fish larvae and fingerlings. Rev Fish Sci 5:1–25
Takeuchi T (2001) A review of feed development for early life stages of marine finfish in Japan. Aquaculture 200:203–222
Furuita H, Takeuchi T, Uematsu K (1998) Effects of eicosapentaenoic and docosahexaenoic acids on growth, survival and brain development of larval Japanese flounder (Paralichthys olivaceus). Aquaculture 161:269–279
Furuita H, Konishi K, Takeuchi T (1999) Effect of different levels of eicosapentaenoic acid and docosahexaenoic acid in Artemia nauplii on growth, survival, and salinity tolerance of larvae of the Japanese flounder, Paralichthys olivaceus. Aquaculture 170:59–69
Kanazawa A (1993) Nutritional mechanisms involved in the occurrence of abnormal pigmentation in hatchery-reared flatfish. J World Aquacult Soc 24:162–166
Song X, Zhang X, Guo N, Zhu L, Kuang C (2007) Assessment of marine thraustochytrid Schizochytrium limacinum OUC88 for maliculture by enriched feeds. Fish Sci 73:565–573
Estevez A, Kanazawa A (1996) Fatty acid composition of neural tissues of normally pigmented and unpigmented juveniles of Japanese flounder using rotifer and Artemia enriched with –3 HUFA. Fish Sci 62:88–93
Shields RJ, Bell JG, Luizi FS, Gara B, Bromage NR, Sargent JR (1999) Natural copepods are superior to enriched Artemia nauplii as feed for halibut larvae (Hippoglossus hipoglossus) in terms of survival, pigmentation and retinal morphology: relation to dietary essential fatty acids. J Nutr 129:1186–1194
Kayaba T (2005) Studies on techniques for reliable production of seed of barfin flounder Verasper moseri (doctoral thesis). Sci Rep Hokkaido Fish Exp Stn 69:1–116 (in Japanese with English abstract)
Okada N, Takagi Y, Seikai T, Tanaka M, Tagawa M (2001) Asymmetrical development of bones and soft tissues during eye migration of metamorphosing Japanese flounder, Paralichthys olivaceus. Cell Tissue Res 304:59–66
Sæle O, Solbakken JS, Watanabe K, Hamre K, Pittman K (2003) The effect of diet on ossification and eye migration in Atlantic halibut larvae (Hippoglossus hippoglossus L.). Aquaculture 220:683–696
Hamre K, Harboe T (2008) Artemia enriched with high n-3HUFA may give a large improvement in performance of Atlantic halibut (Hippoglossus hippoglossus L.) larvae. Aquaculture 277:239–243
Acknowledgments
The authors extend their thanks to Shoji Yoshida of the Tomakomai Fisheries Association in Hokkaido, Japan, for capturing brown sole brood stock. Mitsuru Sannohe and Kosuke Haga from the Erimo town in Hokkaido, Japan, are thanked their assistance with rearing techniques. In addition, Dr. Yutaka Haga, Tokyo University of Marine Science and Technology, is thanked for his valuable advice with this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Satoh, N., Takaya, Y. & Takeuchi, T. The effect of docosahexaenoic and eicosapentaenoic acids in live food on the development of abnormal morphology in hatchery-reared brown sole Pseudopleuronectes herzensteini . Fish Sci 75, 1001–1006 (2009). https://doi.org/10.1007/s12562-009-0125-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12562-009-0125-x