
Abstract
Alternaria molds produce a variety of chemically diverse secondary metabolites with potentially adverse effects on human health. However, data on occurrence in food and human exposure is inconsistent for some of these mycotoxins. Membrane filtration is a frequent step in many sample preparation procedures for LC-MS-based methods analyzing food contaminants. Yet, little is known about the possibility of adsorptive phenomena that might result in analyte losses. Thus, we treated a complex extract of Alternaria toxins with several types of syringe filters and unraveled the impact on its chemical composition by LC-MS/MS. We observed significant, and in some cases complete, losses of compounds due to filtration. Particularly, two key Alternaria toxins, alternariol (AOH) and its monomethyl ether (AME), were heavily affected. As a comparison with published food surveys indicating a correlation of the type of filtration used with lower incidence reports in food, our results point at a possible underestimation of AME in past exposure assessment. Also, perylene quinones were greatly affected by filtration, underlining the importance to take this into consideration during analytical method development. Furthermore, we applied the comet assay in HT-29 cells to elucidate the impact of filtration on the genotoxicity of the extract. We observed strong coincidences with the loss of epoxide-carrying metabolites and also an intriguing induction of oxidative DNA damage by yet toxicologically uncharacterized Alternaria toxins. In conclusion, we highlight potential issues with sample filtration and call for a critical re-evaluation of previous food occurrence data in the light of the results at hand.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Black molds of the genus Alternaria are ubiquitous microorganisms, which can grow on a variety of substrates, such as soil, decaying organic materials, and agricultural crops. The infestation of grains, vegetables, and fruits can lead to the occurrence of potentially toxic fungal secondary metabolites in food and feed (Ostry 2008).
Alternaria molds are known to be able to produce more than 70 mycotoxins of a wide diversity of chemical classes (Fig. 1), which are until today not regulated by respective authorities and are thus considered to belong to the “emerging mycotoxins” (Fraeyman et al. 2017). Tenuazonic acid (TeA), an amine/amide metabolite, is the most frequently reported Alternaria toxin in food samples (Marin et al. 2013). Due to its rather mild acute toxic properties and lack of chronic toxicity, it is considered of little concern for humans (EFSA 2016).

Chemical structures of selected Alternaria toxins
However, two toxins belonging to the group of dibenzo-α-pyrones, namely alternariol (AOH) and alternariol monomethyl ether (AME), are considered a higher risk due to their potential impact on DNA integrity. Both toxins are very similar in chemical structure (Fig. 1) and biological activity. They are known to target human topoisomerase II, thus leading to DNA double-strand breaks in vitro (Fehr et al. 2009). Furthermore, they were reported to modulate inflammatory responses (Kollarova et al. 2018; Solhaug et al. 2015) and to interact with steroid receptors (Dellafiora et al. 2018; Frizzell et al. 2013; Lehmann et al. 2006; Stypuła-Trębas et al. 2017; Vejdovszky et al. 2017). Regarding their in vivo activity, the few available data are inconsistent and limited by technical problems which prevented clear evidence on systemic genotoxicity (Schuchardt et al. 2014). On this base, and the reported prevalence in food samples, the EFSA CONTAM panel lately estimated the 95th percentile dietary exposure of the European population to exceed toxicological threshold of concern (TTC) values (EFSA 2016).
Epoxide-carrying perylene quinones such as altertoxin (ATX)-II or stemphyltoxin-III (STTX-III) were demonstrated to possess a high potential for genotoxicity and other adverse effects in vitro (Aichinger et al. 2018; Del Favero et al. 2020; Fleck et al. 2012; Schwarz et al. 2012b; Tiessen et al. 2013). Even so, they have not yet been a primary focus of risk assessment, probably due to their high reactivity and thus low estimated bioavailability and relevance in vivo. However, perylene quinones were identified as the main contributors to DNA-damaging properties of extracts from Alternaria molds grown under laboratory conditions (Fleck et al. 2016; Schwarz et al. 2012b).
By applying single-cell gel electrophoresis, we recently reported ATX-II and STTX-III to be responsible for a good part of DNA damage caused by a naturally occurring mixture of Alternaria toxins in human cancer cells. However, a certain proportion of genotoxicity—particularly involving the induction of formamidopyrimidine-DNA glycosylase (FPG)–sensitive sites—was maintained after removing those two compounds from the mixture (Aichinger et al. 2019). Thus, other, yet toxicologically uncharacterized, Alternaria metabolites might cause DNA damage as well, which is a hypothesis that has urgently to be enlightened to ensure completeness of the respective risk assessment.
For assessing mycotoxins in biological matrices, state-of-the-art analytics mostly involve liquid chromatography coupled to tandem mass spectrometry (LC-MS/MS). This allows the simultaneous and sensitive quantification of multiple analytes (Warth et al. 2013). Many of these methods (Prelle et al. 2013; Siegel et al. 2010; Tölgyesi et al. 2015) use membrane filtration during sample preparation to reduce matrix effects, but primarily to protect LC systems from small particles.
However, it might be necessary to take possible adsorptive effects at membrane filters into account. Although related literature reports are rather limited, such a scenario does not seem unlikely, as previously published literature demonstrates the possibility of analyte-membrane interactions. For example, a study carried out by Carlson and Thompson (2000), who filtrated chemicals commonly encountered in drug preparations with cellulose, nylon, or polyvinylidene difluoride (PVDF) filters, found all analytes—and particularly those with rather acidic properties (e.g., sodium saccharine or salicylic acid)—to be adsorbed by the membrane matrices.
Mycotoxins, including AOH, were previously described to be absorbed by materials like cyclodextrins, which can be used to remove them from aqueous solutions (Fliszár-Nyúl et al. 2019). With materials commonly used for filtration steps during sample preparation, knowledge about such adsorptive effects is yet scarce for mycotoxins and other natural contaminants. However, the high chemical diversity of Alternaria toxins presupposes a considerable likelihood for such phenomena. To shed light on this issue would be of great importance for analytical method development and also for the evaluation of the analytical quality of previously published food surveys that used filtration techniques during sample preparation. Moreover, the high chemical reactivity of epoxide-carrying perylene quinones might cause an additional loss of those compounds due to the spontaneous formation of covalent adducts with filter membranes.
Thus, in this study, we used six different material types of syringe filters to assess the impact of filtration on the chemical composition of a complex mixture of Alternaria metabolites. Furthermore, we applied in vitro genotoxicity testing of the filtrates to collect hints on the identity of yet undescribed DNA-damaging mycotoxins.
Materials and methods
Materials
Ethidium bromide, Triton X-100, and solutions of NH4Ac (5 mM) and ammonia were purchased from Sigma-Aldrich (Schnelldorf, Germany). Normal and low-melting agarose were purchased from Bio-Rad (Frankfurt, Germany). LC-MS grade acetonitrile and methanol were purchased from Honeywell (Seelze, Germany). Alternaria toxins were purchased as reference materials: TeA, ATX-I, and tentoxin from Sigma-Aldrich (Steinheim, Germany), AOH and AME from Toronto Research Chemicals (Ontario, Canada), altenusin from Eubio (Vienna, Austria), altersetin from AnalytiCon Discovery GmbH (Potsdam, Germany). Altenuene was provided by Prof. Joachim Podlech (Institute of Organic Chemistry, Karlsruhe Institute of Technology, Germany). ATX-I, ATX-II, STTX-III, and alterperylenol were isolated from Alternaria alternata grown on rice by an optimized protocol based on Schwarz et al. (2012a). FPG was bought from New England Biolabs (Frankfurt, Germany). A complete extract (CE) originating from rice infected with the Alternaria alternata strain DSM 62010 was acquired as stated in Puntscher et al. (2019c) and Puntscher et al. (2019a).
Polytetrafluorethylene (PTFE, pore size 0.22 μm/diameter 30 mm), PVDF (0.45/15), and nylon (0.2/30) syringe filters were purchased from Carl Roth (Karlsruhe, Germany); polyethersulfone (PES, 0.2/30), glass fiber/cellulose acetate (GF/CA, 0.2/30), and regenerated cellulose (RC, 0.45/15) syringe filters were obtained from Sarstedt (Nümbrecht, Germany).
Filtration
The ethanolic complete Alternaria toxin extract (CE) was diluted to 100 μg/mL in phosphate-buffered saline (PBS) solution (pH 7.4), resulting in a final concentration of approximately 0.04% ethanol. The resulting solution was used to perform filtration with six different syringe filters applying 3 mL each of the aqueous extract solution. The obtained aqueous filtrates were aliquoted and stored at − 80 °C for later genotoxic testing. For quantitative purposes, the filtrates were diluted by factors of 0, 10, or 70 with a 1:1 mixture of acetonitrile and methanol, vortexed, and subsequently injected into the LC-MS/MS system.
Since significant losses of compounds by filtration were observed (see Results section), the filtration experiment was repeated and after the filtration of the aqueous extract solution, the filters were washed with 3 mL of acetonitrile/methanol (1:1) to assess the recovery of potentially adsorbed toxins. The relative amount of toxins in the aqueous and the organic filtrate was determined by LC-MS/MS.
LC-MS/MS
Aqueous and organic filtrates were measured by LC-MS/MS, applying a method originally developed for food matrices with minor modifications (Puntscher et al. 2018). In brief, a Dionex Ultimate 3000 ultra-high-performance liquid chromatography (UHPLC) system coupled to a TSQ Vantage triple quadrupole mass spectrometer (Thermo Scientific) was used. The MS was equipped with a heated electrospray ionization (HESI) interface (Thermo Scientific) and measurements were performed in the negative electrospray ionization mode. Chromatographic separation was performed using an Ascentis Express column (C18, 100 × 2.1 mm, 2.7 μm, Supelco, Sigma-Aldrich) equipped with a SecurityGuard™ (C18, 2 mm, Phenomenex, Aschaffenburg, Germany). The mobile phases consisted of 5 mM NH4Ac solution in water adjusted to a pH of 8.7 with 25% ammonia solution (eluent A) and methanol (eluent B). (For the detailed gradient, as well as mass transitions, please see Puntscher et al. (2018).) The mass transitions of AST can be found at Puntscher et al. (2019a).
Cell culture
HT-29 colon carcinoma cells were acquired from the German Collection of Microorganisms and Cell Cultures (DSMZ, Braunschweig, Germany) and cultivated in Dulbecco’s Modified Eagle’s Medium (DMEM), supplemented with 10% (v/v) of heat-inactivated fetal calf serum and 1% (v/v) of penicillin/streptomycin. Dulbecco’s phosphate-buffered saline (DPBS), cell culture media, and supplements were obtained from GIBCO Invitrogen (Karlsruhe, Germany); cell culture flasks and dishes were purchased from Sarstedt (Nümbrecht, Germany). Cells were grown to 70–90% confluence at 37 °C in humidified conditions in 5% CO2 atmosphere.
Single-cell gel electrophoresis (“comet assay”)
The comet assay was carried out following the guidelines of Tice et al. (2000) with minor modifications as published previously (Aichinger et al. 2017). Briefly, 150,000 cells were seeded in petri dishes and allowed to grow for 48 h. Cells were incubated for 1 h with the aqueous filtrates or the solvent control (PBS), diluted by a factor of 10 with non-supplemented DMEM. UV-B radiation was used to create a positive control. Cells were harvested, embedded in agarose, and lysed by immersion in a detergent-containing buffer. After treating half of the slides with FPG, electrophoresis (25 V, 300 mA) was performed under strong alkaline conditions (pH 13) for 20 min. Subsequently, slides were neutralized and stained with a solution of 10 μg/mL ethidium bromide. The “Comet Assay IV” system (Perceptive Instruments, Suffolk, UK) was used to score the tail intensity of 100 cells per object slide with a Zeiss Axioskop (λex = 546 ± 1 nm; λem = 590 nm), the average of which was used as a measure for DNA damage.
Statistical evaluation
Statistical analysis of results was performed using the Origin2019 software. Data was confirmed to be normally distributed with the Shapiro-Wilk test. Subsequently, we carried out calculations for assessing significant differences using either one-way ANOVA with Fisher LSD post hoc testing, or Student’s t test.
Results
Chemical composition
Filtration of the CE with different syringe filters resulted in a diverse pattern of compound loss depending on both the type of filter and the analyte (Table 1). The potentially hazardous dibenzo-α-pyrones AOH and AME were significantly reduced by RC and completely removed by nylon, PES, or GF/CA filtration. A prominent difference between the two compounds appeared with PVDF membranes, as it adsorbed AME to a much higher extent (almost completely) as compared with AOH (24% reduction). The structurally similar altenuene (ALT) was seemingly less affected by filtration, with recoveries of 85% for PVDF, 29% for nylon, 18% for PES, 59% for GF/CA, and 103% for RC membranes. Tentoxin (TEN) was moderately affected by all filter types, with losses between 40% (PES) and 60% (GF/CA). Of particular interest, the perylene quinone family—ATX-I and ATX-II, alterperylenol (ALP), STTX-III—exerted similar adsorptive properties. They were not susceptible to PTFE or RC treatment and moderately impacted by PVDF, but almost completely eliminated by nylon, PES, and GF/CA filtration. The less studied metabolites altersetin (AST) and altenusin (ALS) showed distinct adsorptive behaviors. AST was moderately affected by PTFE, significantly reduced by RC, and completely eliminated by PVDF, nylon, and PES membranes. ALS was almost not affected by PTFE, PVDF, GF/CA, and RC filtration, but significantly adsorbed at nylon and PES membranes.
Recovery of adsorbed toxins
The used filters were washed with methanol/acetonitrile (1:1) to allow for an estimation of whether organic solvents could recover the adsorbed toxins. A total recovery was calculated by summing up the peak areas measured in aqueous and organic filtrates, taking into account respective dilution factors and filtrate volumes. Of note, a certain error should be expected to occur due to unavoidable residues of the aqueous filtrate in the syringe filters after the first filtration step, as well as slight variations in passage volumes. Thus, it is important to state that the obtained recovery values should be considered as approximation.
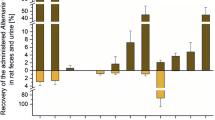
The respective results revealed the ability of methanol/acetonitrile to elute part of the adsorbed mycotoxins (Fig. 2), at least for some toxins. Roughly a third of the lost AOH was recovered from nylon, PES, GC/CA, and RC filters. The total recovery for AME from the PVDF membrane was about 50% higher as compared with the aqueous filtrate, but AOH was not washed from this filter at all (Fig. 2(a)). Furthermore, approximately 50% of the adsorbed AME were recovered from nylon, PES, GC/CA, and RC filters (Fig. 2(a)). The total recovery of ALT was comparable with the other dibenzo-α-pyrones, with the exception that the recovered level from PTFE filters did not exceed 58% even after organic elution (Fig. 2(a)). The perylene quinones ATX-I, ATX-II, ALP, and STTX-III showed very similar recovery patterns (Fig. 2(b)). All of them were partially (25–36%) eluted from nylon and PES and to a lesser extent (14–16%) from GF/CA filters, while the washing of PVDF and RC filters had little to no effect on the total recovery (< 8%). Small amounts of TEN were elutable from nylon and GF/CA, but not from PTFE filters (Fig. 2(c)). Regarding ALS, 29% of the adsorbed toxin was recovered by washing the nylon filter, 13% from GF/CA, 16% from PES, and 7% from PVDF filters. Of note, the compound adsorbed at the PTFE membrane was again only recovered in insignificant doses (Fig. 2(c)). Furthermore, moderate amounts of AST (11–40%) were recovered from all filters (Fig. 2(c)).

Recovery of (a) dibenzo-α-pyrones, (b) perylene quinones, (c) the miscellaneous toxins TEN, AST, and ALS after aqueous filtration and an additional washing step with methanol/acetonitrile (1:1). Bars show peak areas, corrected for dilution factors and sample volumes, and related to the unfiltered sample, which is indicated by a dashed line
Genotoxicity
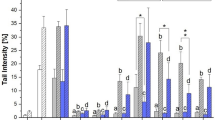
DNA damage was assessed by conducting comet assays after a 1 h incubation of HT-29 cells with 1:10 dilutions of the aqueous filtrates or the unfiltered extract in DMEM, resulting in a final concentration of 5 μg/mL. Each sample was measured with and without FPG treatment to discriminate for the induction of FPG-sensitive sites. Tail intensities (TI) for both the solvent (SC) and the positive control were in line with our expectations and confirmed the functionality of our test system (Fig. 3). In line with recently published data (Aichinger et al. 2019), the unfiltered extract caused mean TIs of 11.5% (untreated) and 26.1% (+ FPG). Filtration of the CE with PTFE did not have an impact on its genotoxicity. Filtrations with PVDF or RC reduced the induction of DNA strand breaks without enzymatic treatment to 4.9% and 8.0% TI, respectively, but did not impact the overall genotoxicity including FPG-sensitive sites. The strongest detoxifying effects were observed for filtrations with nylon, PES, and GF/CA membranes. In these cases, the levels of strand breaks were reduced to not being distinguishable from the SC independent of FPG treatment. However, re-running one-way ANOVA only for the low-damaged groups (SC, nylon, PES, GF/CA) revealed the GF/CA-treated filtrate to induce significantly higher genotoxicity as compared with the SC, after enzymatic treatment.

Impact of filtration on the genotoxicity of the Alternaria extract in HT-29 cells, as measured by comet assay. Bars are depicting mean values + SD of the measured tail intensities of at least 4 independent biological replicates. Normal distribution was confirmed with the Shapiro-Wilk test. Significant differences were calculated by one-way ANOVA, followed by Fisher LSD post hoc testing (p < 0.05), with “a” indicating a significant difference to the respective solvent control (SC) and “b” to the unfiltered extract. A separate one-way ANOVA was carried out for the samples revealing very low tail intensities to again test for significant differences to the respective solvent control, which are indicated by “c.” The positive control was tested against the SC with Student’s t test, observed significant differences therewith are indicated by “d.” The same was used for testing the impact of FPG treatment for each sample, which significantly enhanced tail intensities of all samples, including SC, except for the PES filtrate (not indicated). The underlying table shows LC-MS data for the corresponding aqueous filtrates, in relation to the respective toxin concentration in the unfiltered sample
Discussion
Syringe filters are widely applied in sample preparation for LC-MS based food safety surveys, yet little is known about possible interactions between the membrane material and potential analyte losses. Crop-infesting Alternaria spp. may produce a cocktail of more than 70 toxic metabolites for which up to date no regulations exist and which are thus commonly classified as “emerging mycotoxins” (Gruber-Dorninger et al. 2017). The high chemical diversity in this mixture of contaminants gives rise to the suspicion that at least some of its substances might be affected by adsorptive phenomena during filtration. If occurring during food monitoring studies, this could lead to an underestimation of humane exposure with potentially problematic consequences for risk assessment. Also, knowing such interactions would be of great value for the future development of appropriate analytical methods for Alternaria toxins.
To address these pending questions, we used a well-characterized complex extract from an Alternaria alternata strain grown on rice (Puntscher et al. 2019c) and subjected it to microfiltration by using six distinct syringe filter types. The subsequent analysis of the filtrates’ chemical composition by LC-MS/MS revealed significant adsorptive phenomena that vary with both the membrane material and the monitored analyte (Table 1).
TeA, the most abundant Alternaria toxin, was not adsorbed in significant proportions at any of the tested filters. The genotoxic compound AOH, which is of particular concern for human health according to EFSA (2016), was not affected by PTFE filtration, but partially adsorbed by PVDF and RC and completely bound by nylon, PES, and GF/CA filters. Of note, we found filtration to affect the structurally and mechanistically similar derivative AME in a stronger way, as the toxin was removed completely by filtration with PVDF and even adsorbed to a minor extent by PTFE membrane (Table 1). The lost dibenzo-α-pyrones could only partially be recovered by washing the filters with acetonitrile/methanol (Fig. 2), indicating the relevance of adsorptive phenomena also in these eluents.
Interestingly, and potentially of concern, there seems to be a connection between our results and published occurrence data sets, in which food surveys employing filtration with PVDF or nylon filters report conspicuously less frequent contaminations with AOH and AME as compared with studies using PTFE, RC, or no filters at all (Table 2). Moreover, studies screening food types particularly affected by Alternaria toxin contamination, e.g., tomato products, tend to report AOH, but no AME at all, when using PVDF filters (De Berardis et al. 2018; Prelle et al. 2013). However, in the absence of filtration, the latter compound is commonly reported to occur even more frequently and in higher concentrations as compared with AOH (Noser et al. 2011; Puntscher et al. 2019b; Zhao et al. 2015).
This discrepancy might indicate a potential filtration-derived underestimation of AME’s risk for human health. In their last dietary exposure assessment, members of the EFSA CONTAM panel stated that “reported levels of AME were lower than those reported for AOH” and that AME was “quantified in a few samples of tomato-based products, although at the lower levels than those reported for AOH” (EFSA 2016). The results of the study at hand suggest that the respective occurrence data should be critically re-evaluated on the basis of used filtration techniques and this step should be particularly well-monitored during future method development and validation. In the light of this, accurate specification of the used membrane filters is crucial for data interpretation and should be further encouraged (Table 2). Providing manufacturer and a respective trademark is not sufficient since this does not define the filter material and, e.g., PTFE and PCDF filters exert significant differences in their adsorption of analytes.
Another particularly intriguing question arising from our results is whether the described adsorptive phenomena could be responsible for the rare reports of mycotoxins belonging to the perylene quinone family in food samples. Despite their low occurrence data in food, they are evidently among the predominantly formed secondary metabolites of Alternaria molds grown under laboratory conditions (Puntscher et al. 2019c; Zwickel et al. 2018). For epoxide-carrying representatives like ATX-II and STTX-III, the chemical reactivity towards co-occurring food constituents and fibers might at least contribute to this mystery (Aichinger et al. 2018; Crudo et al. 2020). However, for non-reactive metabolites like ATX-I or ALP, our data suggests that filtration might be a major source of compound loss. Even the considerable concentrations present in the used extract were completely lost by filtration with nylon, PES, and GF/CA filters, and significantly reduced by PVDF filtration (Table 1). Furthermore, the absence of differences in the adsorptive behavior between epoxide and non-epoxide containing perylene quinones, and the possibility to recover parts of all the compounds by washing with organic solvents (Fig. 2), rules out a chemical reaction of ATX-II/STTX-III with membrane materials.
Our data underlines the importance of considering filtration with caution when screening for these compounds. In line with this concern, so far, the only report of ATX-II in naturally contaminated food samples applied a sample preparation protocol without a filtration step (Puntscher et al. 2020). Nevertheless, and on the contrary to AOH/AME, a clear connection between filtration technique and reporting in food samples could not be established from the available data, probably also due to the little number of studies incorporating methods which include those toxins.
An additional aim of this study was to assess the impact of filtration on the DNA-damaging properties of the used complex extract, as previous experiments suggested a contribution of compounds yet not characterized as genotoxic agents (Aichinger et al. 2019). Using HT-29 colon carcinoma cells, we performed single-cell gel electrophoresis to observe a pattern of genotoxicity that was largely following our expectations based on the measured chemical composition of the filtrates (Fig. 3). In line with previous data, DNA damage—both with and without FPG treatment—seemed proportional to the present concentration of ATX-II and STTX-III, the two most potent genotoxic chemicals of the extract (Fleck et al. 2016; Schwarz et al. 2012b). No significant induction of DNA strand breaks was determined for any of the filtrates hardly containing those compounds (nylon, GF/CA, PES). Thus, additional genotoxic compounds can be expected to exert similar adsorptive properties and could possibly be structurally related, such as ATX-III that was not included in the analytical method, and thus may be present in the extract in unknown concentrations.
Intriguingly, the GF/CA-treated extract was still able to significantly induce FPG-sensitive sites (Fig. 3), indicating the presence of compounds responsible for oxidative stress. With AOH, AME, and the perylene quinones practically absent and TeA and TEN concentrations not being higher than in PES-treated filtrates, the possible identifiable candidates to exert this effect are limited to ALT, ALS, and AST. ALT was previously analyzed in the comet assay with respect to DNA-damaging potential in HT-29 cells and showed no effect up to a concentration of 100 μM (Fehr et al. 2009). Data on the impact of AST and ALS on human cells are scarce, with AST—the compound occurring in much higher concentrations in our extract (Table 1)—being described as a weak topoisomerase II inhibitor under cell-free conditions (Jarolim et al. 2016). Its potential to induce oxidative stress and possibly oxidative DNA damage should be examined by further studies. However, it also cannot be excluded that the observed induction of FPG-sensitive sites was caused by further, yet completely unknown, Alternaria metabolites.
Taken together, this study demonstrates the high potential of adsorptive filter-analyte interactions, which might result in an underestimation of toxin occurrence, and thus exposure, of certain Alternaria toxins, particularly the genotoxic AME as well as perylene quinones. We conclude that both the future LC-MS/MS method development and the retrospective data evaluation in the course of risk assessment should take such phenomena into consideration. Furthermore, we collected additional hints on yet undescribed genotoxic Alternaria metabolites and recommend respective testing, including AST.
Data availability
All data and existing sample material will be stored at the Department of Food Chemistry and Toxicology, University of Vienna, and can be made available upon reasonable request.
Abbreviations
- ALP:
-
Alterperylenol
- ALS:
-
Altenusin
- ALT:
-
Altenuene
- AME:
-
Alternariol monomethyl ether
- AOH:
-
Alternariol
- AST:
-
Altersetin
- ATX:
-
Altertoxin
- FPG:
-
Formamidopyrimidine-DNA glycosylase
- GF/CA:
-
Glass fiber/cellulose acetate
- PES:
-
Polyethersulfone
- PTFE:
-
Polytetrafluorethylene
- PVDF:
-
Polyvinylidene difluoride
- RC:
-
Regenerated cellulose
- STTX:
-
Stemphyltoxin
- TeA:
-
Tenuazonic acid
- TEN:
-
Tentoxin
- TI:
-
Tail intensity
References
Aichinger G, Beisl J, Marko D (2017) Genistein and delphinidin antagonize the genotoxic effects of the mycotoxin alternariol in human colon carcinoma cells. Mol Nutr Food Res 61:1600462. https://doi.org/10.1002/mnfr.201600462
Aichinger G, Puntscher H, Beisl J, Kutt ML, Warth B, Marko D (2018) Delphinidin protects colon carcinoma cells against the genotoxic effects of the mycotoxin altertoxin II. Toxicol Lett 284:136–142. https://doi.org/10.1016/j.toxlet.2017.12.002
Aichinger G, Kruger F, Puntscher H, Preindl K, Warth B, Marko D (2019) Naturally occurring mixtures of Alternaria toxins: anti-estrogenic and genotoxic effects in vitro. Arch Toxicol 93(10):3021–3031. https://doi.org/10.1007/s00204-019-02545-z
Carlson M, Thompson RD (2000) Analyte loss due to membrane filter adsorption as determined by high-performance liquid chromatography. J Chromatogr Sci 38:77–83. https://doi.org/10.1093/chromsci/38.2.77
Crudo F, Aichinger G, Mihajlovic J et al (2020) Gut microbiota and undigested food constituents modify toxin composition and suppress the genotoxicity of a naturally occurring mixture ofAlternaria toxins in vitro. Arch Toxicol. https://doi.org/10.1007/s00204-020-02831-1
De Berardis S, De Paola EL, Montevecchi G, Garbini D, Masino F, Antonelli A, Melucci D (2018) Determination of four Alternaria alternata mycotoxins by QuEChERS approach coupled with liquid chromatography-tandem mass spectrometry in tomato-based and fruit-based products. Food Res Int 106:677–685. https://doi.org/10.1016/j.foodres.2018.01.032
Del Favero G, Hohenbichler J, Mayer RM, Rychlik M, Marko D (2020) Mycotoxin altertoxin II induces lipid peroxidation connecting mitochondrial stress response to NF-κB inhibition in THP-1 macrophages. Chem Res Toxicol 33(2):492–504. https://doi.org/10.1021/acs.chemrestox.9b00378
Dellafiora L, Warth B, Schmidt V, Del Favero G, Mikula H, Fröhlich J, Marko D (2018) An integrated in silico/in vitro approach to assess the xenoestrogenic potential of Alternaria mycotoxins and metabolites. Food Chem 248:253–261. https://doi.org/10.1016/j.foodchem.2017.12.013
Diana Di Mavungu J, Monbaliu S, Scippo M-L, Maghuin-Rogister G, Schneider Y-J, Larondelle Y, Callebaut A, Robbens J, Van Peteghem C, De Saeger S (2009) LC-MS/MS multi-analyte method for mycotoxin determination in food supplements. Food Addit Contam A 26:885–895. https://doi.org/10.1080/02652030902774649
EFSA – European Food Safety Authority, Panel on Contaminants in the Food Chain (2016) Dietary exposure assessment to Alternaria toxins in the European population. EFSA J 14:e04654. https://doi.org/10.2903/j.efsa.2016.4654
Fehr M, Pahlke G, Fritz J, Christensen MO, Boege F, Altemöller M, Podlech J, Marko D (2009) Alternariol acts as a topoisomerase poison, preferentially affecting the IIalpha isoform. Mol Nutr Food Res 53:441–451. https://doi.org/10.1002/mnfr.200700379
Fleck SC, Burkhardt B, Pfeiffer E, Metzler M (2012) Alternaria toxins: altertoxin II is a much stronger mutagen and DNA strand breaking mycotoxin than alternariol and its methyl ether in cultured mammalian cells. Toxicol Lett 214:27–32. https://doi.org/10.1016/j.toxlet.2012.08.003
Fleck SC, Sauter F, Pfeiffer E, Metzler M, Hartwig A, Koberle B (2016) DNA damage and repair kinetics of the Alternaria mycotoxins alternariol, altertoxin II and stemphyltoxin III in cultured cells. Mutat Res 798-799:27–34. https://doi.org/10.1016/j.mrgentox.2016.02.001
Fliszár-Nyúl E, Lemli B, Kunsági-Máté S, Szente L, Poór M (2019) Interactions of mycotoxin alternariol with cyclodextrins and its removal from aqueous solution by beta-cyclodextrin bead polymer. Biomolecules 9:428. https://doi.org/10.3390/biom9090428
Fraeyman S, Croubels S, Devreese M, Antonissen G (2017) Emerging Fusarium and Alternaria mycotoxins: occurrence, toxicity and toxicokinetics. Toxins 9:228. https://doi.org/10.3390/toxins9070228
Frizzell C, Ndossi D, Kalayou S, Eriksen GS, Verhaegen S, Sørlie M, Elliott CT, Ropstad E, Connolly L (2013) An in vitro investigation of endocrine disrupting effects of the mycotoxin alternariol. Toxicol Appl Pharmacol 271:64–71. https://doi.org/10.1016/j.taap.2013.05.002
Gotthardt M, Asam S, Gunkel K, Moghaddam AF, Baumann E, Kietz R, Rychlik M (2019) Quantitation of six Alternaria toxins in infant foods applying stable isotope labeled standards. Front Microbiol 10:109. https://doi.org/10.3389/fmicb.2019.00109
Gruber-Dorninger C, Novak B, Nagl V, Berthiller F (2017) Emerging mycotoxins: beyond traditionally determined food contaminants. J Agric Food Chem 65:7052–7070. https://doi.org/10.1021/acs.jafc.6b03413
Hickert S, Bergmann M, Ersen S, Cramer B, Humpf H-U (2016) Survey of Alternaria toxin contamination in food from the German market, using a rapid HPLC-MS/MS approach. Mycotoxicol Res 32:7–18. https://doi.org/10.1007/s12550-015-0233-7
Janić Hajnal E, Orčić D, Torbica A, Kos J, Mastilović J, Škrinjar M (2015) Alternaria toxins in wheat from the autonomous province of Vojvodina, Serbia: a preliminary survey. Food Addit Contam A 32:361–370. https://doi.org/10.1080/19440049.2015.1007533
Jarolim K, Del Favero G, Ellmer D, Stark TD, Hofmann T, Sulyok M, Humpf H-U, Marko D (2016) Dual effectiveness of Alternaria but not Fusarium mycotoxins against human topoisomerase II and bacterial gyrase. Arch Toxicol 91:2007–2016. https://doi.org/10.1007/s00204-016-1855-z
Juan C, Chamari K, Oueslati S, Mañes J (2016) Rapid quantification method of three Alternaria mycotoxins in strawberries. Food Anal Methods 9:1573–1579. https://doi.org/10.1007/s12161-015-0338-9
Kollarova J, Cenk E, Schmutz C, Marko D (2018) The mycotoxin alternariol suppresses lipopolysaccharide-induced inflammation in THP-1 derived macrophages targeting the NF-kappaB signalling pathway. Arch Toxicol 92:3347–3358. https://doi.org/10.1007/s00204-018-2299-4
Lehmann L, Wagner J, Metzler M (2006) Estrogenic and clastogenic potential of the mycotoxin alternariol in cultured mammalian cells. Food Chem Toxicol 44:398–408. https://doi.org/10.1016/j.fct.2005.08.013
López P, Venema D, de Rijk T, de Kok A, Scholten JM, Mol HGJ, de Nijs M (2016) Occurrence of Alternaria toxins in food products in The Netherlands. Food Control 60:196–204. https://doi.org/10.1016/j.foodcont.2015.07.032
Marin S, Ramos AJ, Cano-Sancho G, Sanchis V (2013) Mycotoxins: occurrence, toxicology, and exposure assessment. Food Chem Toxicol 60:218–237. https://doi.org/10.1016/j.fct.2013.07.047
Noser J, Schneider P, Rother M, Schmutz H (2011) Determination of six Alternaria toxin with UPLC-MS/MS and their occurrence in tomatoes and tomato products from the Swiss market. Mycotoxin Res 27:265–271. https://doi.org/10.1007/s12550-011-0103-x
Ostry V (2008) Alternaria mycotoxins: an overview of chemical characterization, producers, toxicity, analysis and occurrence in foodstuffs. World Mycotoxin J 1:175–188. https://doi.org/10.3920/WMJ2008.x013
Prelle A, Spadaro D, Garibaldi A, Gullino M (2013) A new method for detection of five Alternaria toxins in food matrices based on LC-APCI-MS. Food Chem 140:161–167. https://doi.org/10.1016/j.foodchem.2012.12.065
Puntscher H, Kütt M-L, Skrinjar P, Mikula H, Podlech J, Fröhlich J, Marko D, Warth B (2018) Tracking emerging mycotoxins in food: development of an LC-MS/MS method for free and modified Alternaria toxins. Anal Bioanal Chem 410:4481–4494. https://doi.org/10.1007/s00216-018-1105-8
Puntscher H, Aichinger G, Grabher S, Attakpah E, Krüger F, Tillmann K, Motschnig T, Hohenbichler J, Braun D, Plasenzotti R, Pahlke G, Höger H, Marko D, Warth B (2019a) Bioavailability, metabolism, and excretion of a complex Alternaria culture extract versus altertoxin II: a comparative study in rats. Arch Toxicol 93:3153–3167. https://doi.org/10.1007/s00204-019-02575-7
Puntscher H, Cobankovic I, Marko D, Warth B (2019b) Quantitation of free and modified Alternaria mycotoxins in European food products by LC-MS/MS. Food Control 102:157–165. https://doi.org/10.1016/j.foodcont.2019.03.019
Puntscher H, Hankele S, Tillmann K, Attakpah E, Braun D, Kütt ML, del Favero G, Aichinger G, Pahlke G, Höger H, Marko D, Warth B (2019c) First insights into Alternaria multi-toxin in vivo metabolism. Toxicol Lett 301:168–178. https://doi.org/10.1016/j.toxlet.2018.10.006
Puntscher H, Marko D, Warth B (2020) First determination of the highly genotoxic fungal contaminant altertoxin II in a naturally infested apple sample. Emerg Contam 6:82–86. https://doi.org/10.1016/j.emcon.2020.01.002
Qiao X, Yin J, Yang Y, Zhang J, Shao B, Li H, Chen H (2018) Determination of Alternaria mycotoxins in fresh sweet cherries and cherry-based products: method validation and occurrence. J Agric Food Chem 66:11846–11853. https://doi.org/10.1021/acs.jafc.8b05065
Rodriguez-Carrasco Y, Mañes J, Berrada H, Juan C (2016) Development and validation of a LC-ESI-MS/MS method for the determination of Alternaria toxins alternariol, alternariol methyl-ether and tentoxin in tomato and tomato-based products. Toxins 8(11):328. https://doi.org/10.3390/toxins8110328
Rubert J, Dzuman Z, Vaclavikova M, Zachariasova M, Soler C, Hajslova J (2012) Analysis of mycotoxins in barley using ultra high liquid chromatography high resolution mass spectrometry: comparison of efficiency and efficacy of different extraction procedures. Talanta 99:712–719. https://doi.org/10.1016/j.talanta.2012.07.010
Schuchardt S, Ziemann C, Hansen T (2014) Combined toxicokinetic and in vivo genotoxicity study on Alternaria toxins. EFSA Support Publ 11:679E. https://doi.org/10.2903/sp.efsa.2014.EN-679
Schwarz C, Kreutzer M, Marko D (2012a) Minor contribution of alternariol, alternariol monomethyl ether and tenuazonic acid to the genotoxic properties of extracts from Alternaria alternata infested rice. Toxicol Lett 214:46–52. https://doi.org/10.1016/j.toxlet.2012.08.002
Schwarz C, Tiessen C, Kreutzer M, Stark T, Hofmann T, Marko D (2012b) Characterization of a genotoxic impact compound in Alternaria alternata infested rice as altertoxin II. Arch Toxicol 86:1911–1925. https://doi.org/10.1007/s00204-012-0958-4
Siegel D, Feist M, Proske M, Koch M, Nehls I (2010) Degradation of the Alternaria mycotoxins alternariol, alternariol monomethyl ether, and altenuene upon bread baking. J Agric Food Chem 58:9622–9630. https://doi.org/10.1021/jf102156w
Solhaug A, Wisbech C, Christoffersen TE, Hult LO, Lea T, Eriksen GS, Holme JA (2015) The mycotoxin alternariol induces DNA damage and modify macrophage phenotype and inflammatory responses. Toxicol Lett 239:9–21. https://doi.org/10.1016/j.toxlet.2015.08.1107
Ssepuuya G, Van Poucke C, Ediage EN, Mulholland C, Tritscher A, Verger P, Kenny M, Bessy C, De Saeger S (2018) Mycotoxin contamination of sorghum and its contribution to human dietary exposure in four sub-Saharan countries. Food Addit Contam A 35:1384–1393. https://doi.org/10.1080/19440049.2018.1461253
Stypuła-Trębas S, Minta M, Radko L, Jedziniak P, Posyniak A (2017) Nonsteroidal mycotoxin alternariol is a full androgen agonist in the yeast reporter androgen bioassay. Environ Toxicol Pharmacol 55:208–211. https://doi.org/10.1016/j.etap.2017.08.036
Tice RR, Agurell E, Anderson D, Burlinson B, Hartmann A, Kobayashi H, Miyamae Y, Rojas E, Ryu JC, Sasaki YF (2000) Single cell gel/comet assay: guidelines for in vitro and in vivo genetic toxicology testing. Environ Mol Mutagen 35:206–221. https://doi.org/10.1002/(sici)1098-2280(2000)35:3<206::aid-em8>3.0.co;2-j
Tiessen C, Gehrke H, Kropat C, Schwarz C, Bächler S, Fehr M, Pahlke G, Marko D (2013) Role of topoisomerase inhibition and DNA repair mechanisms in the genotoxicity of alternariol and altertoxin-II. World Mycotoxin J 6:233–244. https://doi.org/10.3920/wmj2013.1592
Tölgyesi Á, Stroka J, Tamosiunas V, Zwickel T (2015) Simultaneous analysis of Alternaria toxins and citrinin in tomato: an optimised method using liquid chromatography-tandem mass spectrometry. Food Addit Contam A 32:1512–1522. https://doi.org/10.1080/19440049.2015.1072644
Vejdovszky K, Hahn K, Braun D, Warth B, Marko D (2017) Synergistic estrogenic effects of Fusarium and Alternaria mycotoxins in vitro. Arch Toxicol 91:1447–1460. https://doi.org/10.1007/s00204-016-1795-7
Walravens J, Mikula H, Rychlik M, Asam S, Devos T, Ediage EN, Diana Di Mavungu J, Jacxsens L, Van Landschoot A, Vanhaecke L, De Saeger S (2016) Validated UPLC-MS/MS methods to quantitate free and conjugated Alternaria toxins in commercially available tomato products and fruit and vegetable juices in Belgium. J Agric Food Chem 64:5101–5109. https://doi.org/10.1021/acs.jafc.6b01029
Warth B, Sulyok M, Krska R (2013) LC-MS/MS-based multibiomarker approaches for the assessment of human exposure to mycotoxins. Anal Bioanal Chem 405:5687–5695. https://doi.org/10.1007/s00216-013-7011-1
Zhao K, Shao B, Yang D, Li F (2015) Natural occurrence of four Alternaria mycotoxins in tomato- and citrus-based foods in China. J Agric Food Chem 63:343–348. https://doi.org/10.1021/jf5052738
Zwickel T, Kahl SM, Rychlik M, Müller MEH (2018) Chemotaxonomy of mycotoxigenic small-spored Alternaria fungi – do multitoxin mixtures act as an indicator for species differentiation? Front Microbiol 9:1368. https://doi.org/10.3389/fmicb.2018.01368
Acknowledgments
We thank the Mass Spectrometry Core Facility (MSC) of the University of Vienna for their support, as well as Hannes Puntscher for his aid in discussing methodical issues.
Funding
Open access funding provided by University of Vienna. This study was funded by the University of Vienna, Austria.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethics approval
Not applicable.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Aichinger, G., Živná, N., Varga, E. et al. Microfiltration results in the loss of analytes and affects the in vitro genotoxicity of a complex mixture of Alternaria toxins. Mycotoxin Res 36, 399–408 (2020). https://doi.org/10.1007/s12550-020-00405-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12550-020-00405-9