
Abstract
Late Miocene sediments from the Beli Breg Coal Basin, Western Bulgaria, were investigated using spore-pollen analysis. Based on palynological characteristics, we describe dynamics and development of vegetation in the studied basin. The main types of palaeocoenoses are distinguished. The fossil flora is characterised by a variable structure of plant communities and diversity of dominant species. In general, the warm temperate representatives of the genera Quercus, Castanea, Corylopsis, Ulmus, and Carya dominated the composition of mixed mesophytic forest palaeocoenoses. Climate data reconstructed using the Coexistence Approach method show that the climate was of a moderately warm type, with a mean annual temperature of ca. 16 °C and temperatures ca. 4 °C by mean in the winter season, thus providing very favourable climatic conditions for the distribution of warm temperate vegetation. The established values for precipitation of about 1000 mm annually indicate the presence not only of a warm but also humid climate, with low seasonality and relatively short dry period.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The specific palaeogeographical position of the Balkan Peninsula which was located between Tethys and Paratethys during the Neogene is important in terms of understanding the evolution of the recent flora and vegetation of this region. The territory of Bulgaria, with its numerous Miocene freshwater basins, appears as a key region to understand the Neogene evolution of the connection between Central and Eastern Europe and Asia Minor (Meulenkamp et al. 1996; Rögl 1998, 1999; Meulenkamp and Sissingh 2003; Kvaček et al. 2006; Popov et al. 2006; Akgün et al. 2007; Akkiraz et al. 2008; Ivanov et al. 2011; Kováčová et al. 2011; Alçiçek and Jiménez-Moreno 2013; Biltekin et al. 2015; Durak and Akkiraz 2016; Ivanov and Worobiec 2017; Kayseri-Özer 2017; Kayseri-Özer et al. 2017, 2019; Yavuz et al. 2017, Vasiliev et al. 2018). Apparently, this area plays a major role in the evolution and migration of the Mediterranean vegetation, in the survival of numerous Palaeogene taxa in refuges, and in the processes of plant speciation (Palamarev 1989; Palamarev and Ivanov 1998, 2001, 2004; Palamarev et al. 1999).
Miocene freshwater basins are widespread on the territory of Bulgaria. Their sediments often contain different plant fossils: carpoids, leaf prints, cuticles, and palynomorphs. During the last decades, palynological studies on late Miocene sediments from Karlovo, Sandanski, Staniantsi, Sofia, Gotse Delchev, and Tundzha Basins have been conducted. Thus, new data on the composition of plant assemblages and vegetation dynamics were obtained (Ivanov 2010, 2015; Ivanov and Lazarova 2005, 2019; Ivanov et al. 2007a, b, 2010, 2012; Hristova and Ivanov 2014). This article is a continuation of previous studies on fossil flora and vegetation in Western and Southern Bulgaria and aims to elucidate the development of vegetation and the evolution of the climate during the late Miocene–early Pliocene, which are still poorly understood so far.
As the spatial distribution of plants and vegetation strongly depends on climate, the reconstruction of fossil vegetation can provide conclusions about past climates. Several quantitative methods have been developed during the last few decades based on this assumption, e.g. the Climate Leaf Analysis Multivariate Programme (CLAMP) (Wolfe 1993), the Coexistence Approach (CA) (Mosbrugger and Utescher 1997; Utescher et al. 2014), the Leaf Margin Analysis (Wilf 1997), the Climatic Amplitude method (Fauquette et al. 1998), and the European Leaf Physiognomic Approach (ELPA) (Traiser et al. 2005). In this way, a number of local, regional and global climatic reconstructions have been proposed for the Neogene period (Bertini 2010, 2002, 2006; Bruch and Gabrielyan 2002; Ivanov et al. 2002, 2007a, b, c, 2011, 2019; Bruch and Kovar-Eder 2003; Fauquette and Bertini 2003; Uhl et al. 2003, 2006, 2007a, b; Bruch et al. 2002, 2004, 2006, 2007, 2011; Mosbrugger et al. 2005; Traiser et al. 2005, 2007; Fauquette et al. 2006, 2007; Jiménez-Moreno 2006; Jiménez-Moreno and Suc 2007; Jiménez-Moreno et al. 2007a, b, c, 2008a, b, 2009; Utescher et al. 2007, 2009a, b, 2011a, b, 2013, 2015; Alçiçek and Jiménez-Moreno 2013; Ivanov 2015; Ivanov and Worobiec 2017; Yavuz et al. 2017, Popova et al. 2017, 2019, Ivanov and Lazarova 2019).
Geological settings
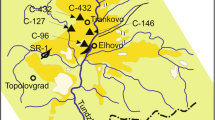
The Beli Breg Basin is located about 50 km west of the city of Sofia (Figs. 1 and 2.). It is an elongated NW-SE structure, with a maximal length of ca. 9 km and a width of 2 to 5 km (Vatsev and Zdravkov 2004). It is surrounded by Jurassic (Polaten and Slivnitsa Formations: Ivanova et al. 2000, Ivanova and Koleva-Rekalova 2004) and Lower Cretaceous rocks as well as Upper Cretaceous volcano-clastic sediments, andesitic to trachyandesitic volcanites, and tuffs. The Neogene sediments are represented by sandstones, carbonates, sandy clays, and up to five brown coal seams, of which only one has an exploitable thickness and wider distribution within the basin.

Geological map of the Beli Breg Lignite Basin (redrawn after Yovchev 1960, with corrections). 1 Neogene sediments; 2 outlines of the coal horizon; 3 Pleistocene sediments; 4 andesites; 5 location of the studied section

Panoramic view of Beli Breg open coal mine and surveyed profile (red numbers indicate the main depths of the profile; GPS coordinates of the profile are shown in the lower right corner)
The lithological subdivision of the sediments in the Beli Breg Basin was discussed by Ivanov et al. (2007b). Yovchev (1960) recognised five informal litostratigraphic units (horizons) in the basin: the first (from bottom to top) is represented by green sandy clays, sands, and conglomerates (up to 50 m in thickness); the second (coal-bearing) is represented by a thick brown coal seam; and the III, IV, and V units lay on top of the above-mentioned units and are represented mainly by dark-green and green clays and sandy clays, not evenly distributed. Later, Vatsev and Zdravkov (2004) introduced three official lithostratigraphic units (from bottom to top): Nedelishte, Kaisiinitsa, and Tranerska Formations. The sediments of the Kaisiinitsa Formation include the main coal seam with the III and IV units described by Yovchev (1960) on top and are subject to our study.
The age of sediments is also under discussion, and according to different authors (Konjarov 1932; Yovchev 1960; Zagorchev et al. 1995; Palamarev 1972; Palamarev and Kitanov 1988; Nikolov 1985; Ognjanova-Rumenova and Yaneva 2000), it varies within a wide frame (see Ivanov et al. 2007b and the discussion therein). Vatsev and Zdravkov (2004) attempted to summarise all available data and stated a Pontian to early Dacian age (late Miocene to early Pliocene) for the Kaisiinitsa Formation.
The biochrononologic data based on the mammalian remains recovered from the strata (Maisch 2014) are generally in line with recent results based on micro- and macropalaeobotanical studies (Ivanov et al. 2007b; Bozukov et al. 2011). The mammalian remains point to a Turolian age at least for the basal part (lignite layer) of the Kaisiinitsa Formation, from where the fossils were recovered. More specifically, the apparent co-occurrence of Tapirus avernensis and Tetralophodon longirostris at Beli Breg indicates an age not younger than MN 12 (Maisch 2014). This conclusion differs in detail from the conclusions of Vatsev and Zdravkov (2004), who assigned a late Turolian age corresponding to MN 13 (a Pontian age) to the coal bearing beds of the Kaisiynitsa Formation. Both palaeobotanical and mammalian data indicate a late Messinian (Pontian) to early Zanclean (early Dacian) age (roughly a middle to late Turolian age) of the Kaitsiynitsa Formation of the Beli Breg Basin.
Palaeogeographically, the Beli Breg Basin represents one of the numerous wetland areas surrounding the Sofia Neogene Basin. In the initial stage of its development, coarse-grained sediments were deposited and subsequently sandy clays. Under ongoing subsidence, swamping and development of marsh vegetation occurred. The development of the peatland was not uniform in space, which is why the thickness of the coal seam varies. Further subsidence of the basin led to flooding of the peat, and lacustrine environments established, with deposition of fine greenish clays, marls, and sandy clays terminating the succession.
Materials and methods
Studied materials and applied techniques
Fossil microbotanical materials have been collected and studied from an outcrop in the central part of the Beli Breg Basin, in the so-called Tsatsarovtsi part (Figs. 1 and 2). The main coal horizon (0.0–14.0 m) is exposed at the base of the section (Fig. 3). The coals are lignitic, so-called xylite-rich coal (Zdravkov and Kortenski 2003, 2004). They contain slightly coalified plant fragments of trunks and branches and other large plant remains, mixed with detrital materials. At places, thin carbonate and clay layers are observed, partly with numerous shells of freshwater molluscs. Remains of large mammals, fish, and birds are less common. The coal complex is overlain by grey marls and clays, often with layers of finely laminated calcareous clays or clays rich in organic matter. Locally, coal layers with a thickness of 10 to 30–40 cm are also observed. In the studied section, the part with siliciclastic sedimentation includes the interval from 14.0 to 67.0 m and is covered with a soil layer on top. The clays contain a variety of macroflora and remains of other fossils—bivalves, fish, and even comparatively well-preserved skeleton parts of a fossilised bird (probably a waterfowl bird acc. to Prof. Z. Boev, NMNH-BAS, Sofia).

Lithological column of the profile from Beli Breg Basin
Palynological data are based on a total of 69 samples taken from a measured section in the Beli Breg open cast mine at a sample rate of approximately 50 cm. Fifty seven out of all analysed samples contained enough palynomorphs for drawing a pollen diagram (more than 500 grains per sample). A single, only moderately rich sample at depth level 5 m has been considered in climate reconstructions and PFT analysis for more completeness of the record. Thus, the data obtained revealed details on the climate and vegetation evolution in the Beli Breg Basin during the late Pontian–early Dacian (Messinian–Zanclean).
The samples were processed according to the standard technique for disintegrating Cenozoic sediments. On the basis of pollen/spore counts, a percentage pollen diagram is plotted (Fig. 4) showing the palynological record of the complete section. The percentage of each pollen taxon identified in the pollen spectra was calculated with respect to the total pollen sum (AP + NAP = 100%), with spores excluded. Local elements (e.g. spores, aquatic plants) were calculated on the basis of the sum AP + NAP+L = 100%.


a/b Pollen diagram of studied section in Beli Breg Basin
Vegetation reconstructions
Vegetation reconstructions are based on both autecological analysis and Plant Functional Types (PFT analysis).
The principles of autecology were used for the reconstruction of vegetation, as well as the data on ecological requirements of the modern analogues of the fossil taxa. As many Neogene European floras, the flora of the Beli Breg Basin includes taxa whose Nearest Living Relatives (NLR) now grow in distant areas, e.g. East Asia and North America. The palaeocoenoses were reconstructed with the help of autecological analysis, assuming that the ecological requirements of fossil taxa are similar to those of their recent analogues; taxa with similar ecological and edaphic requirements were grouped; and the main palaeocommunities were identified.
Charts showing the results of the pollen analyses are illustrated by two types of pollen diagrams: detailed and synthesized. The first diagrams include all identified plants and show their individual presence. In the second type of diagrams, the plants were ordered into ecological groups following Suc (1984) and Jiménez-Moreno et al. (2005) and thus provide information on the general trends in vegetation change. The synthesized pollen diagram was plotted with pollen taxa arranged in different groups on the basis of ecological criteria to clearly show the temporal changes in vegetation.
The groups used are the following (Nix 1982):
Mega-mesothermic (subtropical) elements: “taxodioid” Cupressaceae pollen, Taxodium-type, Symplocos, Engelhardia, Platycarya, Myrica, Sapotaceae, Distylium, Hamamelis, Corylopsis, Castanea-Castanopsis type, Cyrillaceae-Clethraceae, Reevesia, Theaceae, Alangium, Chloranthaceae, Parthenocissus, Araliaceae, Arecaceae and others;
Cathaya: pollen of Cathaya sp.;
Mesothermic elements: (Quercus, Carya, Pterocarya, Carpinus betulus, Carpinus orientalis, Ostrya, Parrotia, Eucommia, Juglans, Zelkova, Ulmus, Tilia, Acer, Liquidambar, Alnus, Salix, Populus, Rhus, Celtis, Platanus, Nyssa, Ilex, Lonicera, Caprifoliaceae, Vitaceae, Fraxinus, Betula, Sequoia-type, Fagus, Hedera, Ilex, Tilia, etc.;
Pinus + Pinaceae: Pinus diploxylon type and undetermined Pinaceae pollen;
Mid-altitude trees (Meso-microthermic elements): Tsuga, Cedrus, Sciadopitys;
High-altitude trees (microthermic elements): Abies, Picea, Keteleeria; Cupressaceae: Cupressus-Juniperus-type and/or pollen irrespective of environmental interpretations, including unspecified pollen grains;
Xerophytes: xerophyte taxa e.g. Quercus ilex-coccifera type, Olea-type (Oleaceae), Caesalpiniaceae, Pistacia, Rhus and others;
Herbs: Poaceae, Amaranthaceae, Asteraceae-Asteroideae, Asteraceae-Cichorioideae, Centaurea, Plantago, Brassicaceae, Lamiaceae, Valerianaceae, Polygonaceae, Knautia (Dipsacoideae), Rosaceae, Malvaceae, Geraniaceae, Erodium, Caryophyllaceae, etc.;
Steppe elements: Artemisia, Ephedra.
Plant Functional Type (PFT) Approach
The PFT approach has been frequently applied in the reconstruction of Cenozoic vegetation patterns and their spatio-temporal changes (e.g. Utescher and Mosbrugger 2007; Popova et al. 2013; Utescher et al. 2017) and is applicable on all types of plant organs. The approach was developed in close connection to biome modelling approaches using the CARAIB dynamic vegetation model because it employs the same classes and hence enables proxy model data comparison (François et al. 2011; Forrest et al. 2015; Henrot et al. 2017). Currently, we use a system including 26 herbaceous to arboreal PFT classes and aquatics (PFT 27) (cf. Table 1) defined in each case by using traits and climatic thresholds (Popova et al. 2013; Henrot et al. 2017). The classification of the fossil record is based on the autecology of the Nearest Living Relatives known for a fossil flora.
In the PFT Approach, only the presence/absence of components is considered which makes the method more robust against taphonomic signals. This is not only useful when integrating over data originating from different plant organs but also in filtering effects of changes of the sedimentary facies in microfloral records. Depending on the taxonomic level of NLR identification, a taxon may contribute to a single or to various different PFTs. In the application of the approach, ecospectra are obtained for each microflora showing the diversity proportions of the single functional types. A list containing palynomorph taxa, NLRs, and PFTs for each sample is provided in the Supplement 1.
Cluster analysis
We performed cluster analysis (clustering of variables) using 43 palynomorph taxa frequently present in the record. The analysis uses Pearson correlation coefficients and average linkage between groups (SPSS software). The dendrogam is subdivided into 8 groups interpreted as vegetation associations.
Palaeoclimate reconstruction
To quantify climate conditions, we apply the Coexistence Approach (CA), a method frequently applied on all types of Cenozoic palaeofloras using the climatic requirements of the Nearest Living Relatives known for a fossil assemblage. In the procedure ranges are identified for each studied variable in which a maximum number of NLRs may coexist. These ranges are denoted Coexistence Intervals and are considered to represent the most probable climatic spans in which the fossil flora existed. More details on the method are given in Mosbrugger and Utescher (1997) and Utescher et al. (2014). The Palaeoflora Database (Utescher and Mosbrugger 1990-2017) is used as source for climate requirements of extant taxa. As climate variables mean annual temperature (MAT), coldest and warmest month means (CMT, WMT), mean annual precipitation (MAP), and mean precipitation of the driest, warmest, and wettest month are reconstructed (MPdry, MPwarm, MPwet). To assess seasonality of climate, mean annual ranges of temperature (MART) and precipitation (MARP) are calculated as WMT-CMT and MPwet-MPdry, respectively, using coexistence interval means. Climate data for all depth levels are given in Supplement 2. A list of fossil palynomorph taxa recovered from the Beli Breg section; their NLR concept and climatic requirements of extant plant taxa are provided in Supplement 3. The CA requires a minimum diversity of 10 taxa contributing with climate data. In our record, 54 samples meet this premise; depth levels 15.25 m and 62.30 m were included but are less reliable having eight taxa only.
Results
Characteristics of fossil flora and vegetation
The first report on plant fossils from the Beli Breg Basin (charophytes) was published in 1972 (Palamarev 1972), but detailed floristic data and their analysis were performed much later (Palamarev and Kitanov 1988). The authors described a total of 34 different taxa and outlined the ecological and coenotical characteristics of the macroflora. Palamarev et al. (1999, 2002) analysed fossil flora from the palaeocological and biostratigraphic point of view aiming to summarise the vegetation evolution in Bulgaria. Later, Bozukov et al. (2011) identified 19 species, 13 of which newly described for that site. The studied material was collected in the upper part of the same profile: from grey-green clayey marl at 53 m and marls at 60 m. On the basis of geological and palaeobotanical data, authors suggested a Dacian age for the fossil flora. Ivanov et al. (2007a) analysed the fossil microflora from a well near the village of Tsatsarovtsi, aiming to obtain the main features of palaeovegetation and climate conditions during the sedimentation process. The above cited authors paid also attention to the palaeoclimatic analysis of the fossil flora and provided first palaeoclimate data for that site.
The present pollen analysis of the sediments from the new section in the Beli Breg Basin reveals the nature and peculiarities of the fossil flora and vegetation during their accumulation. Vegetation and palaeoecological analysis, including a simplified description of vegetation units recognised in the study area, as well as changes in plant communities and vegetation dynamics observed are drawn. The basic floristic diversity of the relatively rich pollen flora is due to the angiosperms. They are represented by 74 species of 49 families (tree and shrub species predominating); gymnosperms are registered with 16 pollen species and ferns with 9 taxa.
The most commonly occurring taxa are species of Betula, Quercus, Fagus, Ulmus, Zelkova, and Carya. The percentage of most taxa varies within relatively narrow ranges (mainly between 1 and 3%) and refers to representatives of Corylus, Carpinus, Cornus, Fraxinus, Acer, Tilia, Eucommia, Pterocarya, Juglans, Engelhardia, and others. In many cases, sporadic occurrence of individual pollen types like Castanea, Corylopsis, Parotia, Reevesia, Buxus, Symplocos, Periploca, Ilex, Sapotaceae, Arecaceae, Vitaceae, Caprifoliaceae, Pistacia, Celtis, Rhus, Oleaceae, and others is observed.
Herbaceous pollen is represented by 19 pollen species (22.5% of the palaeoflora composition) but with low values in the sections. Typically, NAP is in the range of 2–4%, except for individual pollen spectra, where it reaches 6–7%. The pollen of wood and shrub components (AP) is predominant. This implies the dominance of forest-type vegetation in the surroundings of the basin.
Spore plants have been identified with nine types, but their participation in the composition of the vegetation is extremely limited. They appear sporadically in separate pollen spectra. Only representatives of Laevigatosporites (Thelypteridaceae/Polypodiaceae) and Osmunda have a more permanent presence in the spore-pollen spectra, but usually they do not exceed the symbolic 1–2%, apart from an Osmunda peak in the uppermost part of the section (Fig. 4b).
Other microfossils present in the pollen spectra include fungal spores, zygospores (Mougeotia sp., Spirogyra sp., and Zygnema sp.), green algae Botryococcus and Pediastrum, and several types of undetermined algal cysts.
The participation of thermophillous elements in the composition of the flora is significantly limited, both in terms of their floristic diversity and quantitative values (Engelhardia, Platycarya, Symplocos, Reevesia, Sapotaceae, Arecaceae). Only Engelhardia pollen is represented relatively constantly in the pollen spectra, but in only a few cases, it reaches values of 1.5% (usually around or below 1%). The total amount of mega-mesothermal elements confirms their limited role in the formation of plant communities. The highest values of 5–6% are observed in the sediment samples above the main coal seam.
In the studied section, components of different types of palaeocoenoses are recorded: euhydrophytic and hydrophytic grasses, hygrophytic forest (including riparian forest), mixed mesophytic to hygromesophytic forest, as well as subxerophytic elements. The herbaceous component plays a minor role only in the pollen spectra (Figs. 4 and 5).

Synthetic pollen diagram of studied section in Beli Breg Basin. Shaded grey: cycles from Fig. 8
The palaeoecological analysis of the fossil flora and the quantitative participation of the individual taxa shows that the mesophytic forest plant communities that determined the appearance of the zonal vegetation were dominant. This vegetation was of mosaic character, composed of different plant associations that inhabited the lowlands and hilly areas surrounding the Beli Breg Basin. These were basically deciduous forest communities with accessory evergreen components. As mentioned above, palaeotropical elements are poorly represented in the pollen spectra, while the Arctotertiary ones dominate and determine the nature of the vegetation cover. The dominant role in these floristically rich plant associations played the species of Quercus, Fagus, Carya, Betula, and Ulmus, accompanied by species belonging to the following genera: Carpinus, Ostrya, Corylus, Zelkova, Fraxinus, Eucommia, Acer, Juglans, Pterocarya, Platycarya, Tilia, Buxus, Ilex, Parrotia, and others. Deciduous and evergreen representatives of Corylus, Cornus, Ilex, and Ericaceae formed an undergrowth, together with species of the Pteridaceae, Osmundaceae, Polypodiaceae, and Lycopodiaceae families. Lianas of Vitaceae, Periploca, and Humulus also were part of these communities.
Studies on the macroflora of the Beli Breg Coal Basin (Palamarev et al. 2002; Palamarev and Kitanov 1988; Bozukov et al. 2011) point out the importance of species of the Fagaceae family for vegetation structure. They probably represented the dominant plants in the forest communities and formed the main tree layer under which evergreen shrubs such as Buxus, Ilex, and Daphne developed (Palamarev et al. 2002; Bozukov et al. 2011).
The high percentages of conifer pollen in all spectra underline their importance in the vegetation. Pinus pollen predominates over Cathaya (Figs. 4 and 5). It should be emphasised that pollen of conifers and Pinus in particular may occur in high abundance in pollen spectra for two reasons: (1) the high potential of airborne bubbles for long-range transport and (2) the high resistance of the exine of conifer pollen, which favours them during fossilisation processes. The high proportion of Pinus is a constant feature throughout the late Miocene–early Pliocene successions in southeast Romania (Casas-Gallego et al. 2020). The authors suggest these conifers occupied upland areas or were integrated within the deciduous forest. Among the other gymnosperms, Tsuga and Cedrus are more common (over 10%), while Picea, Abies, and Keteleeria are less common, usually in the range of 3–5%. Their record in the pollen spectra indicates the presence of coniferous plants in communities which develop in low- to high-mountain habitats probably with the participation of some representatives of flowering plants such as Corylus, Betula, Fagus, Carpinus, and Pterocarya.
The hygrophytic forest communities (swamp forests), as part of the vegetation in the study area, developed on wet lowlands and swampy coastal territories that provided the necessary water level for their development. These communities have specific environmental requirements. The factors controlling their spread are the presence of a very moist substrate and a constant (or long enough) high water level, whereby the soil remains submerged for most of the growing season. The specific ecological conditions of growth and development are also controlled by their taxonomic composition, which in principle is not distinguished by great floristic diversity.
The elements of the swamp vegetation are poorly represented in the pollen spectra. In all samples analysed, taxodioid Cupressaceae had very low values, rarely reaching up to 2%. While macroremains of Glyptostrobus (leaves or cone twigs) are common in clay sediments (Palamarev et al. 2002; Palamarev and Kitanov 1988), pollen that can be attributed to Glyptostrobus type is very rare. The same applies to the pollen of the other components characteristic of swamp forests such as Alnus, Sciadopitys, Nyssa, Myrica, and Salix.
The main part of the analysed samples originates from clay sediments above the main coal seam. During this period of the basin development, lacustrine conditions with a high water level prevailed. The lacustrine environment was probably not conducive to the development of the swamp forests—there was a lack of sufficiently large wetlands that were suitable for their development. The low pollen content in the coal seam samples does not allow a proper assessment of the distribution of these forests during peat accumulation and their importance for the genesis of the coal. The few available pollen complexes of the coal seam (Fig. 4) show higher values for Alnus—10–15%, and in two of the samples even above 30%. However, the other components do not show increased values. Only the spores of Osmunda and Laevigatosporites (cf. Thelypteridaceae/Polypodiaceae) have slightly higher values of up to 3–4%, apart from one count for Osmunda (6.8% at 41.00 m) and one for Laevigatosporites (12.9% at 32.00 m).
In addition to the floodplain forests, river valley palaeocoenoses were of limited distribution. Pterocarya, Platanus, Liquidambar, Salix, and Itea, which are part of the riparian forests and/or communities in the transitional zones between the floodplain forests and the coastal ones, are also rare. This implies the absence of a highly developed river network in the vicinity of the studied site.
Aquatic vegetation is more or less related to wetlands, especially with regard to the hydrohydrophytic herbaceous palaeocoenoses of the hеlophytes Typha, Sparganium, Alisma, and Stratiotes, which inhabit shallow coastal areas. Considering the percentages of their pollen in the samples examined, it can be assumed that they were of limited distribution. Nuphar and Potamogeton thrived in the open water areas. Pediastrum and Botryococcus both occurring at relatively high percentages likewise indicate the presence of open water. The low quantity of algal spores indicates that habitats suitable for their development (shallow sections with eutrophic stagnant waters) were rare in the lake.
AP pollen (trees and shrubs) dominates the pollen spectra, while grasses are very poorly represented. This determines the dominance of forest vegetation. The pollen of herbaceous plants (mainly Poaceae, Chenopodiaceae, Artemisia, Caryophyllaceae, Polygalaceae, Apiaceae, and Asteraceae) does not exceed 6–7%. In most pollen spectra, NAP ranges from 2 to 4%. These data indicate that herbaceous vegetation and the presence of open areas were very limited in the study area and did not play a significant role in the vegetation structure.
Associations obtained from cluster analysis
- Mixed Mesophytic Forest 1 (MMF 1): This association (Figs. 6 and 7) comprises typical zonal taxa such as Carpinus, Quercus, Fagus, Eucommia, Zelkova, and Oleaceae, as well as Picea and Cathaya and resembles an association described from the Staniantsi section (cf. Utescher et al. 2009a, b: MMF 1). MMF 1 may reach proportions of up to 50% and reaches its highest presence in the middle part of the profile.

Dendrogram classifying main vegetation units using vegetation structure

Synthesized diagram showing groups obtained from cluster analysis. High richness of MMF 1 components relates to warm and humid phases (highlighted grey, cf. Fig. 8)
- Mixed Mesophytic Forest 2 (MMF 2): MMF 2 combines the frequently occurring Pinus diploxylon with other, probably allochthonous elements such as Cornus, Tilia, and Poaceae (Fig. 6). MMF 2 attains the highest richness in the record (partly > 50%) and is regularly present throughout the profile (Fig. 7).
- Riparian community: This association includes many taxa presently common along streams (Alnus, Ulmus) but also hardwood taxa (Acer, Engelhardia, Juglans, Carya, Carpinus, Platanus, Betula). The composition is close to the Staniantsi riparian groups I and II (Utescher et al. 2009a, b) but has less herbaceous plants (Chenopodiaceae). As in Staniantsi, Artemisia is grouped here by cluster analysis. This could point to similar environments in the vicinities of both basins. The record of Artemisia does not necessarily indicate dry conditions but may point to the presence of pioneer herbaceous communities on deforested areas and open shrubs. The riparian community displays cyclic variations and may attain up to 20% (Fig. 7). This is in accordance with the autecological interpretations of vegetation dynamics (see previous chapter).
- Swamp forest with ferns. This association includes Osmunda and Polypodiaceae combined with Taxodioideae, while Tsuga heterophylla type and Asteroideae are of minor importance (Fig. 6). The swamp forest association is most common in the uppermost part of the profile containing brown coal levels where it may attain > 50% (Fig. 7).
- Wetlands: Nyssa, Typha, and Glyptostrobus all indicate a higher water table. However, percentages of all these taxa stay low (Fig. 7). The wetland association is restricted to the uppermost profile part and attains very minor percentages only.
- Conifer association: Very frequent members are Abies, Cedrus, and Tsuga canadensis (Fig. 6). However, low correlation coefficients obtained for the subgroup with Abies, Cedrus, and Ericaceae may indicate transport of the components from outside the basin. This association is throughout present showing cyclic variability and attaining up to 25% (Fig. 7).
- Aquatic elements Botryococcus and Pediastrum. Partially, very high percentages are observed. As evident from their poor correlation, both taxa obviously existed under different palaeoecological conditions. In the obtained cluster combine, both are grouped among the conifers (partly allochthonous?) and the wetland association. Botryococcus is regularly present, while Pediastrum is confined to depth levels 35–60 m.
Diversity of PFTs
Herbaceous PFTs (Fig. 8) are regularly present throughout the section at a mean of ca. 16%. At 43 m and in the topmost part of the profile, from 63 m on, diversity percentages of well over 20% are obtained. In most levels, mesic herbs (PFT 1) dominate; at a few levels (18 m; 47 m), diversity of xeric herbs predominates. C4 herbs (PFT 3) are regularly recorded at minor proportions. Aquatics may attain up to 10.5% of total diversity and tend to be more diverse in the upper part of the section.

Diversity of arboreal PFTs (scaled to 100%) and diversities of herbs and shrubs (% of total diversity). Shorter-term cyclicity: raised diversity of warm temperate conifer PFTs (highlighted grey) and intervals with higher diversity of broadleaved deciduous PFTs (no colour)
Shrubs are represented by PFTs 4–11. Broadleaved evergreen boreal/temperate cold shrubs (PFT 7) and subdesertic shrubs (PFT 10) are rarely recorded. The highest diversity in the shrub fraction is obtained for broadleaved deciduous cool and warm temperate shrubs, regularly present in our record (Fig. 8). Evergreen broadleaved temperate and xeric shrubs (PFTs 8, 9) likewise regularly occur but at lower proportions. With a mean proportion of 7.2% of total diversity (max 14.9%), shrubs may have played a minor role in the late Miocene vegetation at Beli Breg.
Arboreal diversity in the section comprises conifers and broadleaved trees in about equal parts, except for the level 26.5 m and the topmost part from 62 m on, a level with brown coal, in which conifers dominate the spectra. Conifer PFTs regularly include the cold to warm temperate PFTs 12–16, while needleleaved evergreen temperate cool trees (PFT 13) and needleleaved evergreen subtropical trees (PFT 16) represent the most diverse fractions. While needleleaved summergreen boreal/temperate cold trees (PFT 17) are absent from the spectra, needleleaved summergreen subtropical swamp trees regularly occur from ca. 25 m onward, coinciding with increased presence of aquatics. The broadleaved arboreal fraction is mainly composed of cool to warm temperate PFTs 23 and 24. Broadleaved evergreen trees are regularly present at low diversities (< 7% of total diversity; mean 3.7%), except for some levels of the topmost part (Fig. 8). Moreover, there is a sporadic occurrence of raingreen and tropical PFTs (PFTs 25, 26), but all taxa contributing to both classes may likewise represent non-tropical PFTs.
The arboreal PFT record displays a shorter-term cyclicity in the order of 5–12 m, expressed by the ratio of conifers and broadleaved trees. A similar cyclicity is observed in the MMF 1 community described above (Fig. 8) as well as in the dynamic of mesothermic elements and mid-altitude trees (Fig. 5). Seven cycles can be identified in the profile based on PFT record.
Palaeoclimate reconstruction
Palaeoclimate records for the profile sampled in the Beli Breg open cast are presented graphically in Figs. 9 and 10. A total of 56 spore-pollen complexes were analysed using the CA for a total of nine palaeoclimatic variables.

Dynamics of main climate parameters: mean annual, winter and summer temperatures. Temperature records: shorter-term cyclicity; grey, cycles from Fig. 8; red, warm climatic phases; blue, distinctly cool phases. Dotted line: number of palynomorph taxa

Precipitation records. Shorter-term cyclicity: grey, cycles from Fig. 8; blue, humid climatic phases; yellow, distinctly dry phases
The palaeoclimate quantification using the Coexistence Approach points to the persistence of warm temperate conditions during the timespan regarded, with MAT mean values varying between 14–17 °C, CMT at 3–7 °C, and WMT at 23–25 °C. With MAP at 800–1300 mm, MPdry at 20–30 mm, MPwarm at 80–170 mm, and MPwet at 130–220 mm, climate conditions were humid but had a distinct seasonality (MART at 17–21 °C, MARP at 100–190 mm).
The temperature record has only few reliable data up to depth level 14 m, then displays a part with a more stable climate (up to depth level 26 m, cf. WMT variability), and more variable conditions in the upper part of the section. Longer-term trends are expressed in the CMT record showing slightly declining values up to depth level 50 m and increasing again towards the top. With a more or less constant WMT level, MART shows the opposite trends. Moreover, the temperature record displays two cooler phases at 47–53 m and 63–66 m, respectively. The sample at 5 m may also represent a transient cool phase, but too few productive samples are available from that part for clear evidence. The temperature records show smaller-scale cyclicity in the order of 3–6 m that is best expressed in the upper part of the profile.
As regards precipitation, MAP means vary between 800 and 1200 mm, with MPdry at 30–40 mm, MPwarm at 60–170 mm, and MPwet at 130–220 mm. While MPdry stayed at about the same level, MPwarm and MPwet both show some short-term variability. A phase with reduced MPwarm is observed at depth level 28–31 m; various wetter intervals with higher MPwarm and MPwet are recorded at 15–18 m, 38 m, 48–50 m, 55–57 m, and 62 m. With an almost constant MPdry, the highly variable MPwet at the same time points to the existence of pointed variations in MARP throughout the observed timespan. The precipitation data reconstructed for the Beli Breg section clearly show that seasonal drought existed but did not occur in the warm season. At some levels, e.g. at ca. 50 m, MPwarm and MPwet intervals largely overlap, and means are close together; hence, the period with the highest rainfall was in the warmest part of the year.
Discussion
Vegetation dynamics
The relationships between fossil palynomorphs whose modern analogues develop under certain environmental conditions indicate that fossil representatives with modern analogues evolving in warm temperate climates are the most numerous. Subtropical representatives are more restricted in the pollen spectra, typically represented by single pollen grains. Most of them are probably preserved as relics of the middle Miocene subtropical palaeoflora. The group of mega-mesothermic floral elements is poorly represented (Fig. 4). On the other hand, meso-microthermic taxa are better represented. In general, the Arctotertiary component is predominant in the structure of the palaeoflora of Beli Breg, while the palaeotropical component is less prevalent.
The vegetation dynamics in the Beli Breg Basin region are well traced in the pollen diagrams of the studied section (Figs. 4 and 5), due to the more complete fossil record when compared to previous studies by Ivanov et al. (2007a). As noted above, information on fossil vegetation and changes therein is mainly available from clay sediments above the main coal seam (third horizon according to Yovchev 1960). Information on vegetation during peat forming is limited, as only single samples of the lignites (second horizon according to Yovchev 1960) contain enough spores and pollen. As can be assumed, the swamp forests are more widespread during this period (Fig. 5). Higher values are recorded for Alnus species (10–15%), which indicate that its representatives were important vegetational elements during this period.
The spores of Osmunda and Laevigatosporites (cf. Thelypteridaceae/Polypodiaceae) are also slightly more frequent in the brown coal samples. The fragmented nature of this information does not allow for a more accurate assessment of their importance in the formation of peat vegetation, as well as to trace the dynamics in their distribution and to look for patterns similar to those observed in other basins (e.g. Staniantsi–see Utescher et al. 2009a). Data on mesophytic forest communities show dominance of Betula, Quercus, and Ulmus, while Fagus and Carya are less prevalent. Higher values are also obtained for Pterocarya (Fig. 4a), which is an indication of the widespread distribution of representatives of this genus and is probably related to the widespread distribution of adequate wetland environments.
The low content of spores and pollen in the coals does not provide the possibility to properly assess the situation during the accumulation of lignites and the nature of the vegetation that provided the necessary organic material. For this reason, the conclusion of Zdravkov and Kortenski (2003, 2004) that the coal was formed in an environment with predominantly grassy vegetation, mixed with woody plants and with limited distribution of open water in the marsh, cannot be confirmed.
Species of Ephedra, Olea, Celtis, Rhus, Quercus ilex-coccifera type, and Carpinus orientalis/Ostrya type probably formed a kind of mesoxerophytic forest and shrub communities with limited distribution (very low rates in the pollen spectra: Fig. 4b). Probably this type of vegetation only occurred in specific environments such as eroded soils and under specific microclimatic conditions. This coincides with macrofloristic data presented by Palamarev and Kitanov (1988) and Palamarev et al. (2002).
Immediately after the flooding of the peatland and the transition from the marsh to the lake regime in the Beli Breg Basin, the mesothermal and mega-mesothermal floristic elements are represented with the highest values of the whole profile (Fig. 5). During this period, the microthermal elements attain low values, but thereafter tend to slightly increase, and then retain relatively constant values until almost the end of the study period—only at the top of the profile they are again at a low level (Fig. 5). In the composition of forest vegetation, the share of Fagus and Carya increased, while Betula and Quercus maintained their participation without significant changes, but for Ulmus, a decrease in percentage values is observed (Fig. 4a). The further development of the vegetation is again characterised by declining importance of Fagus and Quercus as dominant elements in the forest ecosystems, combined with an increasing the role of Ulmus and Zelkova. This change recorded in the studied section may refer to a climate change that was weak enough not to be resolved by palaeoclimatic reconstruction methods (see next section).
At the top of the profile, there is again a significant change in the proportions of pollen types with the highest participation of Quercus and Ulmus. Abies, Picea, and Tsuga, which may be indicative of changes in the composition of mountain vegetation, all decline again in the uppermost levels. The high Pinus percentage in the upper part of the profile distorts the picture and does not allow deducing a clear signal related to vegetation change.
The dynamics of the vegetation in the studied time period shows relatively small changes in the plant communities, probably of cyclic nature. This points to the absence of longer-term dramatic changes in palaeogeography, environment, and climate. Apart from the above-mentioned frequency changes of the main pollen types, no significant changes are observed in the pollen of many other genera and families; e.g. Carpinus betulus, C. orientalis/Ostrya, Cornus, Acer, Fraxinus, Pterocarya, Juglans, Engelhardia, Oleaceae, Poaceae, and others are present in the pollen spectra with constant values, without sharp deviations. This confirms the assumption of a relatively quiet and stable natural environment.
General climate conditions
The current climate in Western Bulgaria is characterised by the following values: average annual temperature (MAT) 10.0 °C (10.2 °C), coldest month temperature (CMM) − 1.9 °C (− 1.5 °C), warmest month temperature (TWM) 20.5 °C (19.8 °C) and average annual rainfall (MAP) 589 (571) mm, MPwet 90 mm, MPwarm 60 mm, and MPdry 31 mm, according to data from the Sofia Weather Station, located at 584 m asl. (Stringmeteo 2018; Velev 1997). The data for the Dragoman meteorological station (716 m asl) for the period 2000–2007 are MAT 9.4 °C; TWM 19.9 °C; CMM − 1.2 °C; and MAP 516 mm.
Based on mean values Maeotian to Pontian temperature conditions, the study area was characterised by MAT at 15 °C, CMT at 5 °C, and WMT at 24 °C and thus was about 5 °C above the present-day level, but having a close to modern temperature seasonality. The higher-than-present temperature level may in parts be attributable to a then lower elevation and post-Miocene uplift of the study area. With MAP at 1150 mm, MPwet at 170 mm, MPwarm at 124 mm, and MPdry at 25 mm conditions were considerably wetter than present but tended to be more seasonal. Like today, the warm season received a considerable amount of rainfall. All in all, our data coincide with numerous other quantitative reconstructions from about time-equivalent palynomorph records of the Eastern Paratethys (Utescher et al. 2009a, b; Ivanov et al. 2002, 2011). Interregional comparisons with published climate data from neighbouring regions are detailed in Cyclicities in climate and vegetation evolution. Macrofloras commonly providing a more precise reconstruction (Mosbrugger and Utescher 1997) point to somewhat lower CMT (1.7–4.0 °C) in the neighbouring Peripannonian realm (Danube Valley: Utescher et al. 2007) and less rainfall in the warm and wet season. The published data for the Beli Breg macroflora (Bozukov et al. 2011) provide a more precise reconstruction for the study area itself, with narrow climate intervals for MAT and CMT (Table 2). However, the microflora-based temperature means are very close to the values obtained from the macroflora: MAT at 15 °C, CMT at 5 °C, and WMT at 24 °C.
Apart from somewhat cooler conditions at the base and the top of the section, distinct longer-term climatic trends are not resolved by our data. However, our temperature records (Fig. 9) indicate a more stable climatic phase between ca. 14–27 m in the profile and distinct variability from ca. 45 m upwards. Moreover, a long-term increasing trend of seasonality is displayed culminating at ca. 50 m and declining again towards the top.
CMT, WMT, MPwet, and MPwarm all show distinct short-term variability (periodicity at 2–5 m) throughout the section. The observed variability is partly irregular and cannot be strictly related to the cyclic lithological changes recorded in the section but is most probably due to orbital forcing. The cyclic patterns observed in the Beli Breg section are discussed in Cyclicities in climate and vegetation evolution.
Cyclicities in climate and vegetation evolution
A synoptic view on the variability of arboreal diversity and palaeoclimate throughout the studied section reveals a shorter-term cyclic pattern. Phases characterised by higher proportions of conifers in the arboreal spectra are confined to warmer and in most cases wetter climate conditions, best reflected by CMT and WMT as regards temperature, and MPwarm and MPwet for precipitation, respectively. As an exception, a distinctly cool phase recorded at 60 m correlates with humid conditions, also in the dry season. Further evidence for alternating warm/wet–cool/drier phases comes from the ration of drought-tolerant by intolerant conifer PFTs (Fig. 11) that about co-varies with the CA derived precipitation records. Minor non-simultaneity observed for some peaks may point to an asymmetrical reaction of the biosphere on climate change. These signals are, however, close to or beyond the sample resolution, and hence, a reliable interpretation is out of reach.

Moreover, the arboreal PFT record displays a cyclicity in the order of 5–12 m, related to the ratio of conifers and broadleaved trees. The same cyclicity is observed in the Mixed Mesophytic Forest 1 community (MMF 1) described above (Fig. 7). This trend is illustrated by changes in the percentage proportion of mesothermic elements and mid-altitude trees (see Fig. 5 for details). The pollen diagram (Fig. 4a, b) also supports the assumption of cyclicity in the vegetation record. This is best visible it the curves of Cathaya, Cedrus, and Fagus. All these data came in agreement for the cyclic character of the climate as a main driver in the vegetation change, probably due to orbital scale variability (100 kyr).
Comparable longer-term cycles of a 6–12 m periodicity were reported from an about time-equivalent section in the Staniantsi Basin (Utescher et al. 2009a). There, warmer and wetter phases have been related to a higher proportion of the upland vegetation (including conifers), while in cooler, drier phases, pollen from the local wetland vegetation dominated. According to their estimated periods, the longer-term cycles recorded at Staniantsi were interpreted as 100 kyr eccentricity (Laskar et al. 2004). It can be assumed that the cooler, drier phases are correlated with a high eccentricity, while phases with a lower eccentricity correspond to warmer, more equable climate phases. Although 100-kyr eccentricity cycles are less common in Neogene records (cf. Steenbrink et al. 2006), fluctuations referred to eccentricity are reported from the early Pliocene of Italy (Bertini 1994), late Miocene to early Pliocene of southern Romania (Popescu 2001; Popescu et al. 2006), and from the Miocene of the Pannonian Basin (Jiménez-Moreno et al. 2005).
Particularly, late Miocene lacustrine deposits of the Western Pannonian Basin display sedimentary cycles that can be correlated with short eccentricity cycles (~ 100 kyr), among other orbital scale variability (Sacchi and Müller 2004). These studied sequences comprise aggradational stacks formed in a delta plain environment with the occurrence of grain-size minima (lignites or fine organic-rich siliciclastic sediments) at the top of each unit, a lithological pattern resembling the succession of sedimentary units at Beli Breg, even though the patterns observed here are more irregular, a fact that can be explained by field interpretation of grain-size and colour and missing of a proper grain-size analysis. Considering the integrated astronomical calibration obtained for the Iharosberéni 1 well of the Pannonian Basin (Sacchi and Müller 2004) and the astronomical polarity time scale (Laskar 1990), the timespan between 7.3 and 7.0 Ma represents a promising candidate in search of suitable boundary conditions that would explain patterns of climate and vegetation change at Beli Breg. According to this, the lower, climatically more stable part of our section (up to ~ 27 m) would correspond with the timespan 7.3–7.2 Ma characterised by minor insolation changes, while the upper profile part could be correlated with the pointed insolation changes between 7.2 and 6.9 Ma. Consequently, the lower 50 m of the profile would correspond to ca. 5 short eccentricity cycles and represent ca. 500 kyr. Although independent time control at Beli Breg is not precise enough, mammal data from the basal part of the profile indicate a Turolian age not younger than MN12 (~ 7 Ma) and thus are not in the odds with this stratigraphic interpretation.
An interesting phenomenon is the positive correlation between conifer diversity and warmer and wetter phases. The Beli Breg Basin was a lake situated between mountains. This may suggest that the vegetation of the lowlands and low hilly terrains near the water basin was distributed on relatively limited area. Namely, these were the terrains suitable for mesophytic forests. It is very likely that, in warmer and wetter climates, the lake would flood and destroy some of the mesophytic forests in the close vicinity. This was immediately reflected in the pollen spectra, such as the increase in the pollen of the conifers from the mid- and high-altitude areas and the reduction of the pollen from mesophytic forests. On the other hand, in the case of expansion of the lake and increase of the flooded areas, tree pollen with better opportunities for long-range transport was enriched in the central part of the lake (where the studied section originates). Due to their morphology, conifer pollen is usually of high capability for long-distance transport. These two scenarios sufficiently explain the relationship between the frequency of conifers and climate change to warmer and wetter phases.
Another important feature of the fossil flora is the low quantity of herbaceous plants. For the Eastern Paratethys, the emergence of open habitats and the distribution of herbaceous vegetation during the late Miocene characterised the flora and the vegetation turnover (Ivanov et al. 2002, 2007a; Feurdean and Vasiliev 2019; Ivanov and Lazarova 2019). That is not registered in the present section. Herbs occur with significant taxonomic diversity, but in very low quantity. The NAP does not exceed 6–7%, and in most pollen spectra, it ranges from 2 to 4%. Apparently, herb communities were quite limited, and thus, there is no evidence for the existence of open habitats. This specific feature of the flora cannot be explained with certainty. It is worth to mention that, according to the chart presented by Feurdean and Vasiliev (2019): Fig. 2.b), a decrease in the distribution of herbs was recorded between 6.12 and 5.8 Ma in the area of Southeast Europe. Herbaceous pollen percentages increased abruptly to ~ 80%, later on at ~ 4.3 Ma. The absence of significant changes in NAP values throughout the profile makes it impossible to trace the dynamics of grass vegetation and the occurrence of changes in its distribution pattern.
Regional comparisons
Climate data calculated for the Beli Breg microflora are well within the range of values reconstructed for upper Miocene succession in neighbouring areas, e.g. the Forecarpathian Basin (Ivanov et al. 2002), Staniantsi Basin (Utescher et al. 2009a), Tundzha Basin (Ivanov and Lazarova 2019), and Danube Valley/Belgrade (Utescher et al. 2007). The climate record of the Forecarpathian Basin (Drenovets) shows comparative values in the lower part of the Pontian sediments, while in the upper part, the record displays a trend to higher temperature and annual precipitation when compared to our data. Quite similar are the climate data for the pollen flora from the Izgrev Member of Elhovo Formation (Tundzha Basin: Ivanov and Lazarova 2019), but the results from the Sinapovska River outcrop (uppermost part of Elhovo Formation) show slightly lower temperature levels. It is worth to mention that for this outcrop, a high abundance of herbs (NAP = 48%) is a characteristic indicating open habitats.
The comparison of the Beli Breg climate data to coeval results from the Danube Valley in Serbia shows very similar MAT but with a significant difference in the WMT and CMT values. Lower CMT in combination with higher WMT, characteristic for the Serbian flora, probably relate to a more continental type of climate resulting from climate change caused by changes in the boundaries of the water basins located in the close vicinity. Closest to our climatic values are those obtained by Kvaček et al. (2002) for fossil floras from Northern Greece (Table 2). The close similarity climatic requirements of the analysed fossil floras could also be due to their close geographical position and possibly synchronous age.
Comparing palaeoclimate data for the late Miocene from more recent studies on Eastern and Central Europe (Table 2), it is clear that the Beli Breg region was characterised by favourable climate conditions with comparatively high annual temperature and mild winters. In the study area, precipitation rates were apparently higher when compared to other European regions. However, mean precipitation of the driest month was among the lowest. As in the Staniantsi Basin, where the climate record displays cycles of more humid and drier phases (Utescher et al. 2009a), there is also evidence for such fluctuations from the Beli Breg record.
Conclusions
Our multi-method approach applied on a palynomorph record recovered from a measured section sampled in Beli Breg open cast mine reveals new insights into regional late Miocene climate and vegetation dynamics in this part of the Eastern Paratethys. Based on the observed cyclicities, a refined regional stratigraphic concept can be suggested.
Thermophilous elements are poorly represented in the late Miocene vegetation of NW Bulgaria, both in terms of their floristic diversity and in terms of their abundance, thus confirming the previously reported temperate character of the regional vegetation and limited role of mega-mesothermal elements in the formation of plant communities.
The palaeoecological analysis of the fossil flora and abundances of individual palynomorphs show that mesophytic forest plant communities dominated the appearance of the zonal vegetation. Trees and shrub pollen dominate the palynomorph spectra, while herbs are recorded in small quantity, thus testifying to the dominance of forest-type vegetation in the areas surrounding the Beli Breg palaeo-lake.
Vegetation dynamics in the studied late Miocene timespan show relatively minor longer-term changes in the plant communities. This points to the absence of dramatic changes regarding palaeogeography, environment, and climate. The shorter-term dynamics observed in some palynomorph assemblages and individual components imply cyclic changes triggered by climate oscillations.
Cluster analysis suggests the existence of two types of Mixed Mesophytic Forest associations. MMF 1 comprises typical zonal taxa and reaches its highest presence in the middle part of the profile. MMF 2 combines the frequently occurring Pinus and some allochthonous elements. It attains the highest richness in the record (partly > 50%) and is regularly present throughout the profile.
The arboreal PFT record displays a shorter-term cyclicity in the order of 5–12 m, related to the ratio of conifers and broadleaved trees. Comparable cyclicities are observed in MMF 1 community described above and in the dynamic of mesothermic elements and mid-altitude trees. Based on the PFT diversity record, seven cycles in total can be identified in the studied profile.
Based on mean values, the temperature conditions in the study area were about 5 °C above the present-day level, under about twice as high annual precipitation. While seasonality of temperature was close to modern seasonality of rainfall tended to be higher (distinctly high precipitation also in the warm season). CMT, WMT, MPwet, and MPwarm all show distinct short-term variability (periodicity at 2–5 m) throughout the section, most probably related to orbital forcing. The PFT diversity record and MMF 1 association display cyclicity in the order of 5–12 m, related to the ratio of conifers and broadleaved trees. These cycles also coincide with changing proportions of mesothermic elements and mid-altitude trees (e.g. Cathaya, Cedrus, and Fagus). All these data come in agreement with a cyclic character of the climate as a main driver in the vegetation change, due to orbital scale variability, most probably responding to long eccentricity (100 kyr). Cyclostratigraphic considerations suggest ca. 500 kyr for the deposition of the studied sedimentary succession and an age of ca. 7.3–6.8 Ma which coincides with mammal dating.
Palaeoclimate data for the late Miocene from the Beli Breg region point to favourable climate conditions with comparatively high annual temperatures and mild winters. Annual precipitation rates were apparently high when compared to other European regions, but precipitation means of the driest month were distinctly lower. This allows us to consider a more pronounced seasonality when compared to the Central European record.
References
Akgün, F., Kayseri, M. S., & Akkiraz, M. S. (2007). Palaeoclimatic evolution and vegetational changes during the late Oligocene-Miocene period in Western and Central Anatolia (Turkey). Palaeogeography, Palaeoclimatology, Palaeoecology, 253(1–2), 56–90.
Akkiraz, M. S., Kayseri, M. S., & Akgün, F. (2008). Palaeoecology of coal-bearing Eocene sediments in Central Anatolia (Turkey) based on quantitative palynological data. Turkish Journal of Earth Sciences, 17, 317–360.
Alçiçek, H., & Jiménez-Moreno, G. (2013). Late Miocene to Pliocene fluvio-lacustrine system in Karacasu Basin (SW Anatolia, Turkey): Depositional, palaeogeographic and palaeoclimatic implications. Sedimentary Geology, 291, 62–83.
Bertini, A. (1994). Messinian-Zanclean vegetation and climate in North-Central Italy. Historical Biology, 9, 3–10.
Bertini, A. (2002). Palynological evidence of upper Neogene environments in Italy. Acta Universitatis Carolinae, Geologica, 46, 15–25.
Bertini, A. (2006). The northern Apennines palynological record as a contribute for the reconstruction of the Messinian palaeoenvironments. Sedimentary Geology, 188-189, 235–258.
Bertini, A. (2010). Pliocene to Pleistocene palynoflora and vegetation in Italy – State of the art. Quaternary International, 225, 5–24.
Biltekin, D., Popescu, S.-M., Suc, J.-P., Quézel, P., Jiménez-Moreno, G., Yavuz, N., & Çağatay, M. N. (2015). Anatolia: A long-time plant refuge area documented by pollen records over the last 23 million years. Review of Palaeobotany and Palynology, 215, 1–22.
Bozukov, V., Utescher, T., Ivanov, D., Tsenov, B., Ashraf, A. R., & Mosbrugger, V. (2011). New results for the macroflora of the Beli Breg Lignite Basin, West Bulgaria. Phytologia Balcanica, 17(1), 3–19.
Bruch, A. A., & Gabrielyan, I. (2002). Quantitative data of the Neogene climatic development in Armenia and Nakhichevan. Acta Universitatis Carolinae, Geologica, 46, 41–48.
Bruch, A. A., & Kovar-Eder, J. (2003). Climatic evaluation of the flora from Oberdorf (Styria, Austria, Early Miocene) based on the Coexistence Approach. Phytologia Balcanica, 9(2), 175–185.
Bruch, A. A., Fauquette, S., & Bertini, A. (2002). Quantitative climate reconstructions on Miocene palynofloras of the Velona Basin (Tuscany, Italy). Acta Universitatis Carolinae, Geologica, 46, 27–37.
Bruch, A. A., Utescher, T., Olivares, C. A., Dolakova, N., Ivanov, D., & Mosbrugger, V. (2004). Middle and Late Miocene spatial temperature patterns and gradients in Europe — Preliminary results based on palaeobotanical climate reconstructions. Courier Forschungsinstitut Senckenberg, 249, 15–27.
Bruch, A. A., Utescher, T., Mosbrugger, V., Gabrielyan, I., & Ivanov, D. (2006). Late Miocene climate in the circum-Alpine realm — A quantitative analysis of terrestrial palaeofloras. Palaeogeography, Palaeoclimatology, Palaeoecology, 238, 270–280.
Bruch, A. A., Uhl, D., & Mosbrugger, V. (2007). Miocene climate in Europe — Patterns and evolution: A first synthesis of NECLIME. Palaeogeography, Palaeoclimatology, Palaeoecology, 253(1–2), 1–7.
Bruch, A. A., Utescher, T., & Mosbrugger, V. (2011). Precipitation patterns in the Miocene of Central Europe and the development of continentality. Palaeogeography, Palaeoclimatology, Palaeoecology, 304, 202–211.
Casas-Gallego, M., Marza, A., & Tudor, E. (2020). Palaeovegetation and palaeoclimate evolution during the late Miocene to early Pliocene of SE Romania. Geological Journal. https://doi.org/10.1002/gj.3734.
Durak, S. D. Ü., & Akkiraz, M. S. (2016). Late Oligocene-early Miocene palaeoecology based on pollen data from the Kalkım-Gönen Basin (Northwest Turkey). Geodinamica Acta, 28, 295–310.
Fauquette, S., & Bertini, A. (2003). Quantification of the northern Italy Pliocene climate from pollen data: Evidence for a very peculiar climate pattern. Boreas, 32(2), 361–369.
Fauquette, S., Guiot, J., & Suc, J.-P. (1998). A method for climatic reconstruction of the Mediterranean Pliocene using pollen data. Palaeogeography, Palaeoclimatology, Palaeoecology, 144(1–2), 183–201.
Fauquette, S., Suc, J.-P., Bertini, A., Popescu, S.-M., Warny, S., Bachiri Taoufiq, N., Perez Villa, M.-J., Chikhi, H., Feddi, N., Subally, D., Clauzon, G., & Ferrier, J. (2006). How much did climate force the Messinian salinity crisis? Quantified climatic conditions from pollen records in the Mediterranean region. Palaeogeography, Palaeoclimatology, Palaeoecology, 238(1–4), 281–301.
Fauquette, S., Suc, J.-P., Jiménez-Moreno, G., Micheels, A., Jost, A., Favre, E., Bachiri-Taoufiq, N., Bertini, A., Clet-Pellerin, M., Diniz, F., Farjanel, G., Feddi, N., & Zheng, Z. (2007). Latitudinal climatic gradients in the Western European and Mediterranean regions from the Mid-Miocene (c. 15 Ma) to the Mid-Pliocene (c. 3.5 Ma) as quantified from pollen data. Geological Society Special Publication, 481–502.
Feurdean, A., & Vasiliev, I. (2019). The contribution of fire to the late Miocene spread of grasslands in eastern Eurasia (Black Sea region). Scientific Reports, 9(1), 6750.
Forrest, M., Eronen, J. T., Utescher, T., Knorr, G., Stepanek, C., Lohmann, G., & Hickler, T. (2015). Climate-vegetation modelling and fossil plant data suggest low atmospheric CO2 in the late Miocene. Climate of the Past, 11, 1701–1732.
François, L., Utescher, T., Favre, E., Henrot, A.-J., Warnant, P., Micheels, A., Erdei, B., Suc, J.-P., Cheddadi, R., & Mosbrugger, V. (2011). Modelling Late Miocene vegetation in Europe: Results of the CARAIB model and comparison with palaeovegetation data. Palaeogeography, Palaeoclimatology, Palaeoecology, 304, 359–378.
Henrot, A. J., Utescher, T., Erdei, B., Dury, M., Hamon, N., Ramstein, G., Krapp, M., Herold, N., Goldner, A., Favre, E., Munhoven, G., & François, L. (2017). Middle Miocene climate and vegetation models and their validation with proxy data. Palaeogeography, Palaeoclimatology, Palaeoecology, 467, 95–119.
Hristova, V., & Ivanov, D. (2014). Late Miocene vegetation and climate reconstruction based on pollen data from the Sofia Basin (West Bulgaria). Palaeoworld, 23(3–4), 357–369.
Ivanov, D. (2010). Palaeoclimate reconstructions for the late Miocene in the Southeast Bulgaria using pollen data from the Tundzha Basin. In G. Christofides, N. Kantiranis, D.S. Kostopoulus, & A.A. Chatzipetros (Eds.) Proceedings of the XIX CBGA Congress (pp. 269–278). Thessaloniki, Greece: Scientific Annals, School of Geology, Aristotle University of Thessaloniki, Special Volume 100.
Ivanov, D. (2015). Climate and vegetation change during the late Miocene in Southwest Bulgaria based on pollen data from the Sandanski Basin. Review of Palaeobotany and Palynology, 221, 128–137.
Ivanova, D., & Koleva-Rekalova E. (2004). Agglutinated foraminifers in the framework of southwestern Bulgarian palaeoenvironmental evolution during the Late Jurassic and Early Cretaceous. In M. Bubik, & M.A. Kaminski (Eds.) Proceedings of the Sixth International Workshop on Agglutinated Foraminifera. Grzybowski Foundation, spec. Publ., 8, 217–227.
Ivanov, D., & Lazarova, M. (2005). Late Miocene flora from Tundzha Basin. Preliminary palynological data. Comptes Rendus de l'Académie Bulgare des Sciences, 58(7), 799–804.
Ivanov, D., & Lazarova, M. (2019). Past climate and vegetation in Southeast Bulgaria — A study based on the late Miocene pollen record from the Tundzha Basin. Journal of Palaeogeography, 8(1), 43–67.
Ivanov, D., & Worobiec, E. (2017). Middle Miocene (Badenian) vegetation and climate dynamics in Bulgaria and Poland based on pollen data. Palaeogeography, Palaeoclimatology, Palaeoecology, 467, 83–94.
Ivanova, D., Stoykova, K., & Lakova, I. (2000). New microfossil data on the age relationship between Slivnitsa and Salash Formation in Dragoman region, Western Bulgaria. Comptes Rendus de l'Académie Bulgare des Sciences, 53(4), 71–74.
Ivanov, D., Ashraf, A. R., Mosbrugger, V., & Palamarev, E. (2002). Palynological evidence for Miocene climate change in the Forecarpathian Basin (Central Paratethys, NW Bulgaria). Palaeogeography, Palaeoclimatology, Palaeoecology, 178(1–2), 19–37.
Ivanov, D., Ashraf, A. R., Utescher, T., Mosbrugger, V., & Slavomirova, E. (2007a). Late Miocene vegetation and climate of the Balkan region: Palynology of the Beli Breg Coal Basin sediments. Geologica Carpathica, 58(4), 367–381.
Ivanov, D., Bozukov, V., & Koleva-Rekalova, E. (2007b). Late Miocene flora from SE Bulgaria: Vegetation, landscape and climate reconstruction. Phytologia Balcanica, 13(3), 281–292.
Ivanov, D. A., Ashraf, A. R., & Mosbrugger, V. (2007c). Late Oligocene and Miocene climate and vegetation in the Eastern Paratethys area (northeast Bulgaria), based on pollen data. Palaeogeography, Palaeoclimatology, Palaeoecology, 255(3–4), 342–360.
Ivanov, D., Djorgova, N., & Slavomirova, E. (2010). Palynological subdivision of Late Miocene sediments from Karlovo Basin, Central Bulgaria. Phytologia Balcanica, 16(1), 23–42.
Ivanov, D., Utescher, T., Mosbrugger, V., Syabryaj, S., Djordjević-Milutinović, D., & Molchanoff, S. (2011). Miocene vegetation and climate dynamics in eastern and central Paratethys (southeastern Europe). Palaeogeography, Palaeoclimatology, Palaeoecology, 304, 262–275.
Ivanov, D., Utescher, T., Ashraf, A. R., Mosbrugger, V., Bozukov, V., Djorgova, N., & Slavomirova, E. (2012). Late Miocene palaeoclimate and ecosystem dynamics in southwestern Bulgaria a study based on pollen data from the Gotse-Delchev Basin. Turkish Journal of Earth Sciences, 2, 187–211.
Ivanov, D., Tsenov, B., Utescher, T., Kováčová, M., Mosbrugger, V., & Ashraf, A. R. (2019). Climate reconstructions based on Miocene leaf flora from NW Bulgaria: Comparing leaf physiognomy and nearest living relative approach. Phytologia Balcanica, 25(2), 137–146.
Jiménez-Moreno, G. (2006). Progressive substitution of a subtropical forest for a temperate one during the middle Miocene climate cooling in Central Europe according to palynological data from cores Tengelic-2 and Hidas-53 (Pannonian Basin, Hungary). Review of Palaeobotany and Palynology, 142, 1–14.
Jiménez-Moreno, G., & Suc, J.-P. (2007). Middle Miocene latitudinal climatic gradient in Western Europe: Evidence from pollen records. Palaeogeography, Palaeoclimatology, Palaeoecology, 253, 208–225.
Jiménez-Moreno, G., Rodriguez-Tovar, F. J., Pardo-Iguzquiza, E., Fauquette, S., Suc, J.-P., & Muller, P. (2005). High-resolution palynological analysis in late early-middle Miocene core from the Pannonian Basin, Hungary: Climatic changes, astronomical forcing and eustatic fluctuations in the Central Paratethys. Palaeogeography, Palaeoclimatology, Palaeoecology, 216(1), 73–97.
Jiménez-Moreno, G., Aziz, H. A., Rodriguez-Tovar, F. J., Pardo-Iguzquiza, E., & Suc, J.-P. (2007a). Palynological evidence for astronomical forcing in early Miocene lacustrine deposits from Rubielos de Mora Basin (NE Spain). Palaeogeography, Palaeoclimatology, Palaeoecology, 252(3), 601–616.
Jiménez-Moreno, G., Fauquette, S., Suc, J.-P., & Aziz, H. A. (2007b). Early Miocene repetitive vegetation and climatic changes in the lacustrine deposits of the Rubielos de Mora Basin (Teruel, NE Spain). Palaeogeography, Palaeoclimatology, Palaeoecology, 250(1), 101–113.
Jiménez-Moreno, G., Popescu, S.-M., Ivanov, D., & Suc, J.-P. (2007c). Neogene flora, vegetation and climate dynamics in southeastern Europe and the northeastern Mediterranean. In M. Williams, A. M. Haywood, F. J. Gregory, & D. N. Schmidt (Eds.) Deep-time perspectives on climate change: Marrying the signal from computer models and biological proxies (pp. 503–516). London: The Micropalaeontological Society, Geological Society, Special Publications.
Jiménez-Moreno, G., Fauquette, S., & Suc, J.-P. (2008a). Vegetation, climate and palaeoaltitude reconstructions of the eastern Alps during the Miocene based on pollen records from Austria, Central Europe. Journal of Biogeography, 35(9), 1638–1649.
Jiménez-Moreno, G., Mandic, O., Harzhauser, M., Pavelic, D., & Vranjkovic, A. (2008b). Vegetation and climate dynamics during the early middle Miocene from Lake Sinj (Dinaride Lake system, SE Croatia). Review of Palaeobotany and Palynology, 152(3–4), 270–278.
Jiménez-Moreno, G., de Leeuw, A., Mandic, O., Harzhauser, M., Pavelic, D., Krijgsman, W., & Vranjkovic, A. (2009). Integrated stratigraphy of the Early Miocene lacustrine deposits of Pag Island (SW Croatia): Palaeovegetation and environmental changes in the Dinaride Lake system. Palaeogeography, Palaeoclimatology, Palaeoecology, 280, 193–206.
Kayseri-Özer, M. S. (2017). Cenozoic vegetation and climate change in Anatolia — A study based on the IPR-vegetation analysis. Palaeogeography, Palaeoclimatology, Palaeoecology, 467, 37–68.
Kayseri-Özer, M. S., Karadenizli, L., Akgün, F., Oyal, N., Saraç, G., Şen, Ş., Tunoğlu, C., & Tuncer, A. (2017). Palaeoclimatic and palaeoenvironmental interpretations of the late Oligocene, late Miocene-early Pliocene in the Çankırı-Çorum Basin. Palaeogeography, Palaeoclimatology, Palaeoecology, 467, 16–36.
Kayseri-Özer, M.S., Atalar, M., & Kováčová, M. (2019). Palaeovegetation evolution of the Çankırı Basin during the Mio-Pliocene (Central Anatolia) based on the IPR analysis method. In L. W. van den Hoek Ostende, S. Mayda, & T. Kaya (Eds.) Taking the Orient Express? The role of Anatolia in mediterranean Neogene palaeobiogeography palaeobiodiversity and palaeoenvironment, 99(4), 571–590.
Konjarov, G. (1932). Die Braunkohlen Bulgariens. Sofia: Royal Printing House. [in Bulgarian, German abstract]
Kováčová, M., Doláková, N., & Kováč, M. (2011). Miocene vegetation pattern and climate change in the northwestern Central Paratethys domain (Czech and Slovak Republic). Geologica Carpathica, 62(3), 251–266.
Kvaček, Z., Velitzelos, D., & Velitzelos, E. (2002). Late Miocene flora of Vegora, Macedonia, N. Greece: University of Athens.
Kvaček, Z., Kováč, M., Kovar-Eder, J., Doláková, N., Jechorek, H., Parashiv, V., Kováčová, M., & Sliva, Ľ. (2006). Miocene evolution of the landscape and vegetation in the Central Paratethys. Geologica Carpathica, 57(4), 295–310.
Laskar, J. (1990). The chaotic motion of the solar system: A numerical estimate of the size of the chaotic zones. Icarus, 88(2), 266–291. https://doi.org/10.1016/0019-1035(90)90084-M.
Laskar, J., Robutel, P., Joutel, F., Gastineau, M., Correia, A. C. M., & Levrard, B. (2004). A longterm numerical solution for the insolation quantities of the earth. Astronomy and Astrophysics, 428, 261–285.
Maisch, M. W. (2014). Mammalian remains (Mammalia: Perrissodactyla, Proboscidea) from the late Miocene Kaisiynitsa Formation of the Beli Breg Basin (Bulgaria). Neues Jahrbuch für Geologie und Paläontologie (Abhandlungen), 272(1), 109–114.
Meulenkamp, J. E., & Sissingh, W. (2003). Tertiary palaeogeography and tectonostratigraphic evolution of the northern and southern peri-Tethys platforms and the intermediate domains of the African–Eurasian convergent plate boundary zone. Palaeogeography, Palaeoclimatology, Palaeoecology, 196, 209–228.
Meulenkamp, J. E., Kovac, M., & Cicha, I. (1996). On late Oligocene to Pliocene depocentre migrations and the evolution of the Carpathian-Pannonian system. Tectonophysics, 266, 301–317.
Mosbrugger, V., & Utescher, T. (1997). The Coexistence Approach — a method for quantitative reconstructions of Tertiary terrestrial palaeoclimate data using plant fossils. Palaeogeography, Palaeoclimatology, Palaeoecology, 134(1–4), 61–86.
Mosbrugger, V., Utescher, T., & Dilcher, D. L. (2005). Cenozoic continental climatic evolution of Central Europe. Proceedings of the National Academy of Sciences, 102(42), 14964–14969.
Nikolov, I. (1985). Catalogue of the localities of Tertiary mammals in Bulgaria. Palaeontology, Stratigraphy and Litholology, 21, 43–62.
Nix, H. (1982). Environmental determinants of biogeography and evolution in Terra Australis. In W. R. Barker & P. J. M. Greenslade (Eds.) Evolution of the Flora and fauna of arid Australia (pp. 47–66). Frewville: Peacock Publishing.
Ognjanova-Rumenova, N., & Yaneva, M. (2000). Preliminary data on the palaeoecological development of Beli Brjag Basin, SW Bulgaria. (pp. 160–161). In Abstracts Geological Conference. Sofia: Bulgarian Geological Society.
Palamarev, E. (1972). Die Gattung Tectochara im Pliozänbecken der Grube “Bolschewik”. Izvestiya na Botanicheskaya Institut (Sofia) 22, 127-133. [in Bulgarian with German abstract]
Palamarev, E. (1989). Paleobotanical evidences of the Tertiary history and origin of the Mediterranean sclerophyll dendroflora. Plant Systematics and Evolution, 162, 93–107.
Palamarev, E., & Ivanov, D. (1998). Über einige Besonderheiten der tertiären Floren in Bulgarien und ihre Bedeutung für die Entwicklungsgeschichte der Pflanzenwelt in Europa. Acta Palaeobotanica, 38, 147–165.
Palamarev, E., & Ivanov, D. (2001). Charakterzüge der Vegetation des Sarmatien (Mittel- bis Obermiozän) im südlichen Teil des Dazischen Beckens (Südost Europa). Palaeontographica B, 259, 209–220.
Palamarev, E., & Ivanov, D. (2004). Badenian vegetation of Bulgaria: Biodiversity, palaeoecology and palaeoclimate. Courier Forschungsinstitut Senckenberg, 249, 63–69.
Palamarev, E., & Kitanov, G. (1988). The fossil macroflora of the Beli Brjag Coal-Basin. In 100th Anniversary of Acad. Nikolaj A. Stojanov (pp.183-206). Sofia: Publishing House of Bulgarian Academy of Sciences. [in Bulgarian with English abstract]
Palamarev, E., Ivanov, D., & Bozukov, V. (1999). Paläoflorenkomplexe im Zentralbalkanischen Raum und ihre Entwicklungsgeschichte von der Wende Oligozän/Miozän bis ins Villafranchien. Flora Tertiaria Mediterranea, 6(5), 1–99.
Palamarev, E., Bozukov, V., & Ivanov, D. (2002). Late Neogene floras from Bulgaria: Vegetation and palaeoclimate estimates. Acta Universitatis Carolinae, Geologica, 46, 57–63.
Popescu, S.-M. (2001). Repetitive changes in early Pliocene vegetation revealed by high resolution pollen analysis: Revised cyclostratigraphy of southwestern Romania. Review of Palaeobotany and Palynology, 120, 181–202.
Popescu, S.-M., Krijgsman, W., Suc, J.-P., Clauzon, G., Mărunţeanu, M., & Nica, T. (2006). Pollen record and integrated high-resolution chronology of the early Pliocene Dacic Basin (southwestern Romania). Palaeogeography, Palaeoclimatology, Palaeoecology, 238, 78–90.
Popova, S., Utescher, T., Gromyko, D. V., Mosbrugger, V., Herzog, E., & François, L. (2013). Vegetation change in Siberia and the northeast of Russia during the Cenozoic cooling: A study based on diversity of plant functional types. Palaios, 28, 418–432.
Popova, S., Utescher, T., Gromyko, D. V., Bruch, A. A., Henrot, A.-J., & Mosbrugger, V. (2017). Cenozoic vegetation gradients in the mid- and higher latitudes of Central Eurasia and climatic implications. Palaeogeography, Palaeoclimatology, Palaeoecology, 467, 69–82.
Popova, S., Utescher, T., Gromyko, D. V., Mosbrugger, V., & François, L. (2019). Dynamics and evolution of Turgay-type vegetation in Western Siberia throughout the early Oligocene to earliest Miocene - a study based on diversity of plant functional types in the carpological record: Oligocene vegetation of western Siberia. Journal of Systematics and Evolution, 57(2), 129–141.
Popov, S. V., Shcherba, I. G., Ilyina, L. B., Nevesskaya, L. A., Paramonova, N. P., Khondkarian, S. O., & Magyar, I. (2006). Late Miocene to Pliocene palaeogeography of the Paratethys and its relation to the Mediterranean. Palaeogeography, Palaeoclimatology, Palaeoecology, 238(1–4), 91–106.
Rögl, F. (1998). Palaeogeographic considerations for Mediterranean and paratethys seaways (Oligocene to Miocene). Annalen des Naturhistorischen Museums in Wien (A), 99, 279–310.
Rögl, F. (1999). Mediterranean and paratethys. Facts and hypotheses of an Oligocene to Miocene paleogeography (short overview). Geologica Carpathica, 50, 339–349.
Sacchi, M., & Müller, P. (2004). Orbital cyclicity and astronomical calibration of the upper Miocene continental succession cored at the Iharosbereny-I well site, Western Pannonian basin, Hungary. In Cyclostratigraphy: Approaches and Case Histories, 275-294, https://doi.org/10.2110/pec.04.81.0275.
Steenbrink, J., Hilgen, F., Krijgsman, W., Wijbrans, J. R., & Meulenkamp, J. E. (2006). Late Miocene to early Pliocene depositional history of the intramontane Florina-Ptolemais-Servia Basin, NW Greece: Interplay between orbital forcing and tectonics. Palaeogeography, Palaeoclimatology, Palaeoecology, 238(1), 151–178.
Stringmeteo. (2018). Climate data for reference Bulgarian stations (1961–1990, Monthly weather-fore-cast of NIMH). https://www.stringmeteo.com/synop/bg_climate.php?m1=7&m2=8&station. Accessed 8 Nov 2018. [in Bulgarian]
Suc, J.-P. (1984). Origin and evolution of the Mediterranean vegetation and climate in Europe. Nature, 307, 429–432.
Syabryaj, S., Utescher, T., Molchanoff, S., & Bruch, A. A. (2007). Vegetation and palaeoclimate in the Miocene of Ukraine. Palaeogeography, Palaeoclimatology, Palaeoecology, 253(1–2), 153–168.
Traiser, C., Klotz, S., Uhl, D., & Mosbrugger, V. (2005). Environmental signals from leaves — A physiognomic analysis of European vegetation. New Phytologist, 166, 465–484.
Traiser, C., Uhl, D., Klotz, S., & Mosbrugger, V. (2007). Leaf physiognomy and palaeoenvironmental estimates – An alternative technique based on an European calibration. Acta Palaeobotanica, 47(1), 181–201.
Uhl, D., Mosbrugger, V., Bruch, A. A., & Utescher, T. (2003). Reconstructing palaeotemperatures using leaf floras – Case studies for a comparison of Leaf Margin Analysis and the coexistence approach. Review of Palaeobotany and Palynology, 126, 49–64.
Uhl, D., Bruch, A., Traiser, C., & Klotz, S. (2006). Palaeoclimate estimates for the middle Miocene Schrotzburg flora (S Germany): A multi-method approach. International Journal of Earth Sciences, 95(6), 1071–1085.
Uhl, D., Klotz, S., Traiser, C., Thiel, C., Utescher, T., Kowalski, E., & Dilcher, D. L. (2007a). Cenozoic paleotemperatures and leaf physiognomy – A European perspective. Palaeogeography, Palaeoclimatology, Palaeoecology, 248, 24–31.
Uhl, D., Traiser, C., Griesser, U., & Denk, T. (2007b). Fossil leaves as palaeoclimate proxies in the Palaeogene of Spitsbergen (Svalbard). Acta Palaeobotanica, 47(1), 89–107.
Utescher, T., & Mosbrugger, V. (1990-2018). The Palaeoflora Database at http://www.palaeoflora.de.
Utescher, T., & Mosbrugger, V. (2007). Eocene vegetation patterns reconstructed from plant diversity — A global perspective. Palaeogeography, Palaeoclimatology, Palaeoecology, 247(3–4), 243–271. https://doi.org/10.1016/j.palaeo.2006.10.022.
Utescher, T., Djordjevic-Milutinovic, D., Bruch, A., & Mosbrugger, V. (2007). Palaeoclimate and vegetation change in Serbia during the last 30 ma. Palaeogeography, Palaeoclimatology, Palaeoecology, 253(1–2), 141–152. https://doi.org/10.1016/j.epsl.2017.05.019.
Utescher, T., Ivanov, D., Harzhauser, M., Bozukov, V., Ashraf, A. R., Rolf, C., Urbat, M., & Mosbrugger, V. (2009a). Cyclic climate and vegetation change in the late Miocene of Western Bulgaria. Palaeogeography, Palaeoclimatology, Palaeoecology, 272(1–2), 99–114.
Utescher, T., Mosbrugger, V., Ivanov, D., & Dilcher, D. L. (2009b). Present-day climatic equivalents of European Cenozoic climates. Earth and Planetary Science Letters, 284(3–4), 544–552.
Utescher, T., Böhme, M., & Mosbrugger, V. (2011a). The Neogene of Eurasia: Spatial gradients and temporal trends — The second synthesis of NECLIME. Palaeogeography, Palaeoclimatology, Palaeoecology, 304, 196–201.
Utescher, T., Bruch, A. A., Micheels, A., Mosbrugger, V., & Popova, S. (2011b). Cenozoic climate gradients in Eurasia — A palaeo-perspective on future climate change? Palaeogeography, Palaeoclimatology, Palaeoecology, 304, 351–358.
Utescher, T., Böhme, M., Hickler, T., Liu, Y.-S.(C.), Mosbrugger, V., & Portmann, F. (2013). Continental climate and vegetation patterns in North America and Western Eurasia before and after the closure of the Central American Seaway. In GSA 125th Anniversary Annual Meeting. Denver. Abstracts with Programs. Geological Society of America 45(7), 302.
Utescher, T., Bruch, A. A., Erdei, B., François, L., Ivanov, D., Jacques, F. M. B., Kern, A. K., & Liu, Y.-S.(C.), Mosbrugger, V., & Spicer, R.A. (2014). The coexistence approach — Theoretical background and practical considerations of using plant fossils for climate quantification. Palaeogeography, Palaeoclimatology, Palaeoecology, 410, 58–73.
Utescher, T., Bondarenko, O. V., & Mosbrugger, V. (2015). The Cenozoic cooling – Continental signals from the Atlantic and Pacific side of Eurasia. Earth and Planetary Science Letters, 415, 121–133.
Utescher, T., Dreist, A., Henrot, A.-J., Hickler, T., Liu, Y.-S., & (C.), Mosbrugger, V., Portmann, F. T., & Salzmann, U. (2017). Continental climate gradients in North America and Western Eurasia before and after the closure of the Central American Seaway. Earth and Planetary Science Letters, 472, 120–130.
Vasiliev, I., Karakitsios, V., Bouloubassi, I., Agiadi, K., Kontakiotis, G., Antonarakou, A., Triantaphyllou, M., Gogou, A., Kafousia, N., de Rafélis, M., Zarkogiannis, S., Kaczmar, F., Parinos, C., & Pasadakis, N. (2018). Large sea surface temperature, salinity, and productivity-preservation changes preceding the onset of the Messinian salinity crisis in the eastern Mediterranean Sea. Paleoceanography and Paleoclimatology, 34, 182–202.
Vatsev, M., & Zdravkov, A. (2004). On the stratigraphy of the Neogene sediments from Beli Breg Basin, West Sredna Gora zone (Preliminary results). In Annual Scientific Conference “Geology 2004” (pp. 95-96). Sofia: Bulgarian Geological Society.
Velev, S. (1997). Contemporary air temperature and precipitation fluctuations in Bulgaria. In M. Jordanova, & D. Donchev (Eds.) Geography of Bulgaria (pp. 145–150). Sofia: Publishing House Bulgarian Academy Sciences. [in Bulgarian with English abstract]
Wilf, P. (1997). When are leaves good thermometers? A new case for Leaf Margin Analysis. Paleobiology, 23, 373–390.
Wolfe, J. A. (1993). A method of obtaining climatic parameters from leaf assemblages. U.S. Geological Survey Bulletin, 2040, 1–71.
Yavuz, N., Culha, G., Demirer, Ş. S., Utescher, T., & Aydın, A. (2017). Pollen, ostracod and stable isotope records of palaeoenvironment and climate: Upper Miocene and Pliocene of the Çankırı Basin (Central Anatolia, Turkey). Palaeogeography, Palaeoclimatology, Palaeoecology, 467, 149–165.
Yovchev, Y. S. (1960). Mineral resources of P. R. Bulgaria. Coals and bituminous clays. Sofia: Tehnika. [in Bulgarian]
Zagorchev, I., Kostadinov, V., Chunev, D., Dimitrova, R., Sapunov, I., Tchoumatchenko, P., & Yanev, S. (1995). Explanatory notes to the geological map of Bulgaria, Scale 1: 100,000. Vlasotnitze and Breznik map sheet (pp. 1–72). Sofia: Geological and Geophysical Corporation. [in Bulgarian, English abstract]
Zdravkov, A., & Kortenski, J. (2003). Petrology and depositional environment of the coals from Beli Breg Basin, Bulgaria. In Abstracts Annual Scientific Conference 2003. Sofia: Bulgarian Geological Society. Sofia. https://www.bgd.gea.uni-sofia.bg/summanual_2003_6.htm#39.
Zdravkov, A., & Kortenski, J. (2004). Maceral composition and depositional environment of the coals from Beli Breg Basin, Bulgaria. Review of the Bulgarian Geological Society, 65, 157–166.
Acknowledgements
Our sincere thanks go to Marianna Kováčová and Funda Akgün (reviewers), and Angela Bruch (guest-editor) who greatly improved the paper by their comments and suggestions.
Funding
This work was supported by the Project 436 Bul. 113/139/0-1/AS 103/3-1 (DFG, Germany) and by the National Science Program “Environmental Protection and Reduction of Risks of Adverse Events and Natural Disasters”, approved by the Resolution of the Council of Ministers № 577/17.08.2018 and supported by the Ministry of Education and Science (MES) of Bulgaria (Agreement № D01- 230/06.12.2018 and № DO1-322/18.12.2019). It is a contribution to the International Network Programme NECLIME (www.neclime.de).
Author information
Authors and Affiliations
Contributions
All authors read and approved final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is a contribution to the special issue “Palaeobotanical contributions in honour of Volker Mosbrugger”
Rights and permissions
About this article

Cite this article
Ivanov, D., Utescher, T., Djorgova, N. et al. The late Miocene Beli Breg Basin (Bulgaria): palaeoecology and climate reconstructions based on pollen data. Palaeobio Palaeoenv 101, 79–102 (2021). https://doi.org/10.1007/s12549-020-00475-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-020-00475-8