
Abstract
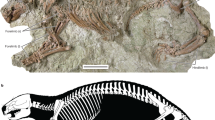
A disarticulated, though still roughly associated partial Deinotherium skeleton from the late Middle Miocene (late Sarmatian sensu stricto; 12.2–12.0 Ma) Gratkorn locality (Austria) is described. Based on dimensions and morphology of the material it can be determined as a medium-sized taxon of Deinotheriidae and definitively assigned to the genus Deinotherium. This specimen from Gratkorn confirms the osteological differences in the postcrania between Prodeinotherium and Deinotherium. As the diagnostically important p/3 is missing on the specimen it can only be assigned to Deinotherium levius vel giganteum. The Gratkorn specimen is one of not many skeletons of a medium-sized taxon of Deinotheriidae and one of only a few well-dated late Middle Miocene occurrences in Central Europe with associated dental and postcranial material.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Deinothere remains are frequent findings in the Miocene of Europe and a useful tool for biochronological and biostratigraphical considerations (see, e.g. Dehm 1960; Huttunen 2002a, b; Böhme et al. 2012; Pickford and Pourabrishami 2013). Following recent works (Böhme et al. 2012; Pickford and Pourabrishami 2013) on the stratigraphic range of the different species, the genus Prodeinotherium Éhik, 1930 can be considered as indicative for the Early to middle Middle Miocene, while Deinotherium Kaup, 1829 first occurs in Europe during the Middle Miocene (Mottl 1969; Svistun 1974) and is recorded up to the terminal Late Miocene (Markov 2008b). Unfortunately, in most cases the findings comprise only isolated remains, and very often only isolated teeth [e.g. abundant tooth material from the famous Eppelsheim Formation (Eppelsheim Fm)]. In contrast to this, a fairly well preserved, disarticulated, partial Deinotherium skeleton (Fig. 1) of late Sarmatian age (12.2–12.0 Ma) was discovered in the clay pit St. Stefan near Gratkorn (Styria, Austria; Gross et al. 2011; 2014, this issue) during geological mapping of the region in 2005. It is one of very few skeleton findings of a medium-sized deinothere taxon described so far. The remains were excavated by the Universalmuseum Joanneum, Graz, from 2005 to 2008. All elements could be assigned to one individual except for some tooth remains detected about 30 m NW of the skeleton that represent a second individual. The fragmentary preservation of the latter allowed stable isotope analyses (δ18OCO3, δ13C; see Aiglstorfer et al. 2014, this issue). The excavation of the Deinotherium skeleton led to the discovery of an abundant and rich vertebrate fauna, which has been excavated in continuous campaigns in a cooperative project of the Universalmuseum Joanneum, Graz, the Eberhard Karls Universität Tübingen and the Ludwig-Maximillians-Universität München (see other publications in this special issue).

Taxonomy of European Deinotheriidae
Taxonomy of Deinotheriidae has been under discussion for long (see, e.g. Gräf 1957; Bergounioux and Crouzel 1962; Harris 1973; Gasparik 1993, 2001; Antoine 1994; Huttunen 2000; Ginsburg and Chevrier 2001; Duranthon et al. 2007; Markov 2008a, b; Vergiev and Markov 2010; Böhme et al. 2012; Pickford and Pourabrishami 2013). At the moment, one, Deinotherium (Ginsburg and Chevrier 2001; Pickford and Pourabrishami 2013), respectively two genera, Prodeinotherium and Deinotherium (Gasparik 1993; Antoine 1994; Huttunen 2000; Duranthon et al. 2007; Vergiev and Markov 2010), are considered valid. While a gradual size increase within Deinotheriidae from the Early to the Late Miocene is generally accepted, Antoine (1994), Huttunen (2000), Vergiev and Markov (2010) and others argue that Prodeinotherium and Deinotherium do not only differ in size but also in dental and postcranial morphology. Huttunen (2000) gives an overview of distinguishing characters between the smaller genus Prodeinotherium and the larger genus Deinotherium, discussing and evaluating the characters given by Harris (1973) and others on specimens from Central Europe. As noted by Huttunen and also observed in this study (see Discussion below), genus diagnostic characters can indeed be identified in the postcranial material and therefore support the separation of two genera Prodeinotherium and Deinotherium as proposed by Éhik (1930). In addition to the on-going discussion on valid genera, different concepts concerning species validity are also held at the moment. While some authors accept five valid morphospecies (Böhme et al. 2012) or chronospecies (Pickford and Pourabrishami 2013), others tend to reduce the number to four (Gasparik 1993, 2001; Markov 2008a; Vergiev and Markov 2010) or even only two species (Huttunen 2002a). Species determination is hindered considerably by the difficulty in identifying stratigraphically mixed faunas, the great dimensional and morphological overlap between the species and the impossibility to evaluate intraspecific variation (Huttunen 2000). Huttunen (2002a), for example, synonymized Deinotherium levius Jourdan, 1861 with Deinotherium giganteum Kaup, 1829 due to the assumed contemporaneous occurrence of D. giganteum and D. levius morphotypes in the Eppelsheim Fm. Furthermore, the mentioned gradual size increase (Gasparik 1993; Böhme et al. 2012; Pickford and Pourabrishami 2013) and the stepwise morphological modification of the characteristic features (Antoine 1994; Gasparik 2001) aggravate a clear species differentiation. Huttunen (2002a), like others before her, considered Deinotherium gigantissimum Stefanescu, 1892 only “a large variety of D. giganteum” (Huttunen 2002a, p. 244). Dating of deinothere findings and identification of stratigraphically mixed faunas are the keys for evaluation of inter- and intraspecific variations and for determination of the role of sexual dimorphism or the sympatric occurrence of different species. In the modern Loxodonta africana Blumenbach, 1797, for example, the average weight of females (about 2.8 t) reaches only about 56 % of the males’ average weight (5 t; Joger 2010). Such a scope would include specimens from Prodeinotherium bavaricum von Meyer, 1831 to D. giganteum. The large dimensional and morphological variability in D. giganteum observed by Huttunen (2000) that led her to a supposed synonymy with D. levius could thus be a consequence of faunal mixing or uncertainty in stratigraphic positions of localities, and also biased by a certain degree of sexual dimorphism (Huttunen 2000, 2002b). The mixed and time-averaged faunal assemblage from the “Dinotheriensande” (Eppelsheim Fm; at that time considered stratigraphically uniform) in particular has biased her observations and those of others for a long time. Böhme et al. (2012) and Pickford and Pourabrishami (2013) were able to show, however, that the Eppelsheim Fm also covers a considerable amount of the Middle Miocene and therefore comprises several non-co-occurring Deinotherium species. In contrast to the observations of Huttunen (2000, 2002a, b), Gräf (1957) gives a morphospecies differentiation of D. giganteum and D. levius based on differences in dental material. She already observed variability concerning dental features but as her comparison material was limited (Pickford and Pourabrishami 2013) some of her features were found to be more variable than she considered (see, for example, Huttunen 2000 for discussion), while others show a smaller variability than she estimated due to mixed faunal assemblages (see, for example, Pickford and Pourabrishami 2013 for discussion). Gräf (1957) further underestimated the dimensional range sometimes (Pickford and Pourabrishami 2013). Pickford and Pourabrishami (2013) based their work on a large number of deinothere dental material and tried to focus their considerations on well-dated material and to avoid faunal assemblages likely to result from a considerable extent of faunal mixing, such as fluvial deposits. These researchers classify different size groups in combination with their stratigraphic range while being well aware that these groups cannot be strictly separated due to a gradual size increase. Böhme et al. (2012) mention D. bavaricum, D. levius and D. giganteum as morphospecies recorded from the Eppelsheim Fm based on comparisons with dental material from rich and well-documented localities from Europe.
We follow the morphospecies concept of Böhme et al. (2012) with five European species, which differs from other concepts, such as those of Gasparik (1993, 2001) and Vergiev and Markov (2010) in the acceptance of the species D. levius, based on the diagnostic features in the p/3 described by Gräf (1957) and referred to, for example, by Mottl (1969) and Böhme et al. (2012). We could observe the generic differences on the postcranial material from Gratkorn in comparison to Prodeinotherium from several localities, and therefore follow the two genera concept as proposed by Éhik (1930) and used by Gasparik (1993, 2001), Huttunen (2000, 2002a, b), Duranthon et al. (2007), Vergiev and Markov (2010) and others, in contrast to Böhme et al. (2012) and Pickford and Pourabrishami (2013). In this work, we therefore consider the following European morphospecies to be valid: Prodeinotherium cuvieri, P. bavaricum, Deinotherium levius, D. giganteum and D. proavum Eichwald, 1831. Codrea (1994), Gasparik (2001) and Pickford and Pourabrishami (2013) stated that D. proavum should have priority over D. gigantissimum Stefanescu, 1892 and that the latter should be considered a junior synonym.
Institutional abbreviations
- GPIT:
-
Paläontologische Sammlung der Universität Tübingen, Tübingen, Germany
- IGM:
-
Montanuniversität Leoben, Leoben, Austria
- MNHN:
-
Muséum National d’Histoire Naturelle, Paris, France
- NHMM:
-
Naturhistorisches Museum Mainz, Mainz, Germany
- NHMW:
-
Naturhistorisches Museum Wien, Vienna, Austria
- NMNHS:
-
National Museum of Natural History, Sofia, Bulgaria
- PMSU:
-
Paleontological Museum of Sofia University “St. Klimt Ochrdisky”, Department of Geology and Paleontology, Sofia, Bulgaria
- SMNS:
-
Staatliches Museum für Naturkunde Stuttgart, Stuttgart, Germany
- SNSB-BSPG:
-
Staatliche Naturwissenschaftliche Sammlungen Bayerns Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany
- SSN:
-
Paläontologisches Museum Nierstein, Nierstein, Germany
- UMJGP:
-
Universalmuseum Joanneum, Graz, Austria
Anatomical abbreviations
- prc/prcd:
-
protocone/protoconid
- pac:
-
paracone
- mc/mcd:
-
metacone/metaconid
- hyc/hycd:
-
hypocone/hypoconid
- ecd:
-
entoconid
- Mc:
-
metacarpal
- Mt:
-
metatarsal
- sin.:
-
sinistral
- dex.:
-
dextral
- lmax:
-
maximal length
- wmax:
-
maximal width
Material
Dental and cranial material
UMJGP 204078 (P4/ sin.); UMJGP 203690 (P4/ dex.); UMJGP 204081 (M1/ sin.); UMJGP 204079 (M2/ sin.); UMJGP 203628 (M2/ dex.); UMJGP 204080 (M3/ sin.); UMJGP 203624 (i/2 dex.?); UMJGP 203670 (p/4 sin.); UMJGP 203669 (m/1 dex.); UMJGP 203689 (m/3 sin.); UMJGP 203654 (fragment of skull ?); UMJGP 203435 (p/4 sin.); 203460 (tooth fragment, buccal wall of 203435?); UMJGP203420-21 (tooth fragments).
Postcranial material
Vertebral column and ribs
UMJGP 204654 (atlas); UMJGP 203623, 204111, 203605 (vertebrae cervicales); 203638, 203653, 203659, 203680 (vertebrae thoracicae or lumbales); UMJGP 203663 (fragment of vertebra caudalis?); UMJGP 204681 (processus spinosus of vertebra cervicalis 6 or 7); UMJGP 203693 (fragment of processus spinosus of vertebra cervicalis 7 or vertebra thoracica 1); UMJGP 203642 (processus spinosus of vertebrae thoracicae 1 or 2); UMJGP 203655, 203649, 203647, 203602, 203694 and 203603 (processus spinosi of cranial series of vertebrae thoracicae); UMJGP 203687 (fragments of processus spinosus (?));UMJGP 203681 (?), UMJGP 204684 (?), UMJGP 203716, UMJGP 203646 (?), UMJGP 203675(?) (fragments of arcus vertebrarum); UMJGP 203604, 203608, 203610 (two crushed costa fragments ?), 203634, 203643, 203644, 203648 (with fragment 203645), 203660(?), 203687, 203696, 203692, 203697, 203703, 203717, 203666, 203658, 203629, 203630, 203635, 203617, 204673(?) (fragments of costae); UMJGP 203657 (costa 1/2? dex.); UMJGP 203606 (costa 2/3? dex.); UMJGP 203639, 203650, 203695, 203633 (costae dex. of central to caudal series of the thorax); UMJGP 204110, 203618 and 203614-5 (fragment of the same costa?), 203631, 203632, 203607 (costae sin. of central to caudal series of the thorax).
Limb elements
UMJGP 203662, 203664, 203667, 203668, 203671, 203672, 203676, 203677, 203678(?), 203679, 203691, 204103 (fragments of scapula?); UMJGP 203674 (humerus dex.? with part of scapula?); UMJGP 203665 (radius sin.); UMJGP 203621 (fragment of radius dex.); UMJGP 203688 (os carpi ulnare sin.); UMJGP 203640 (os carpale secundum sin.); UMJGP 203685 (distal epiphysis of metacarpal II or III sin. or IV dex.); UMJGP 203684 (phalanx proximalis of manus?); UMJGP 204112 (femur dex., distal epiphysis); UMJGP 203601 (femur dex., fragment of proximal shaft); UMJGP 203612, 203613 (fragments of fibula dex.); UMJGP 203622 (fibula sin.); UMJGP 203611 (os tarsi centrale sin.); UMJGP 203683 (os tarsi centrale dex.); UMJGP 204696 (distal trochlea of metatarsal II?); UMJGP 203625 (? metatarsal IV dex.); UMJGP 203708 (phalanx proximalis II, III, IV dex. of pes?); UMJGP 203709 (os sesamoideum); UMJGP 203710 (os sesamoideum); UMJGP 203620 (lateral fragment of metacarpal I or metatarsal I dex.?); UMJGP 203616 (metapodial?).
Methods
For comparison of postcranial material we used the Prodeinotherium skeleton from Langenau (SMNS 41562; Germany; Early Miocene; MN 4; 17.2–17.1 Ma), the partial Prodeinotherium skeletons from Franzensbad (NHMW2000z0047/0001; Czech Republic; Early Miocene; MN 5; 16.9 Ma) and Unterzolling (SNSB-BSPG 1977 I 229; Germany; early Middle Miocene; 15–14.5 Ma) described by Huttunen (2000, 2004) and Huttunen and Göhlich (2002), the partial skeleton of D. levius from Gusyatin (also Husyatyn) (Ukraine; Middle Miocene; early late Badenian; 13.1–13.4 Ma; marine sediments dated with foraminifera by Didkovsk in Svistun 1974) described by Svistun (1974) and the skeleton of Deinotherium proavum from Ezerovo (Bulgaria; Late Miocene; MN 12; Kovachev and Nikolov 2006) mounted at the PMSU, as well as descriptions of postcranial elements by Huttunen (2000).
Comparison material for teeth comprises Prodeinotherium remains from Falun de la Touraine and Anjou (both France; early Middle Miocene; Langhian; MN 5; 15 ± 0.5 Ma), Unterzolling, Sprendlingen 2 (Germany; Middle Miocene), the Eppelsheim Formation and localities from the North Alpine Foreland Basin (NAFB) described by Antoine (1994), Ginsburg and Chevrier (2001), Huttunen and Göhlich (2002), Huttunen (2004), Duranthon et al. (2007) and Böhme et al. (2012). For Deinotherium, dental material from the Middle Miocene sites La Grive, St. Gaudens, Tournan (all France; late Middle Miocene; MN 7/8; 13–11.5 Ma), Massenhausen, Hinterauerbach, Sprendlingen 2 (all Germany; late Middle Miocene; MN 7/8; 13–11.5 Ma), St. Oswald near Gratwein (Austria; Middle Miocene; early Badenian), Oberdorf near Weiz (Austria; late Middle Miocene; late Sarmatian; 12.2–11.6 Ma), Breitenhilm near Hausmannstetten (Austria; late Middle Miocene; late Sarmatian; 12.2–11.6 Ma) and Dietersdorfberg near Mureck (Austria; late Middle Miocene; Sarmatian; 12.7–11.6 Ma) described by Peters (1871), Depéret (1887), Gräf (1957), Mottl (1969, 1970), Ginsburg and Chevrier (2001) and Böhme et al. (2012) was compared with the Gratkorn specimen. Furthermore, we considered Deinotherium giganteum specimens described by Gräf (1957) and Tobien (1988) from Montredon (France; Late Miocene; late Vallesian; MN 10; 9.5 Ma) and Frohnstetten (Germany; Late Miocene), as well as the type of D. giganteum from Eppelsheim (HLMD Din. 466), described by Kaup (1829, 1832). Due to the stratigraphic mixture of the rich Deinotherium material from the Eppelsheim Formation, it is excluded besides the type of D. giganteum. Deinotherium remains from Austria described or referred to by Mottl (1969), Hilber (1914) and Huttunen (2000) and general observations on dental material by Tobien (1988), Antoine (1994), Ginsburg and Chevrier (2001) and Duranthon et al. (2007) on deinothere material from France are included in the discussion. As unfortunately no description on the dental material of D. levius from Gusyatin is given in Svistun (1974), we only took the tooth metrics into consideration here. Furthermore, tooth metrics of (?)D. levius from Opatov (formerly Abtsdorf; Czech Republic; Middle Miocene; Badenian) given by Zázvorka (1940) are considered.
Measurements were accomplished with a calliper (precision if possible 0.1 mm in teeth; 1 mm in postcranial material) and are modified after Göhlich (1998).
Systematic palaeontology
Order Proboscidea Illiger, 1811
Family Deinotheriidae Bonaparte, 1845
Genus Deinotherium Kaup, 1829
Type species: Deinotherium giganteum Kaup, 1829
Valid European species: Deinotherium levius Jourdan, 1861, D. giganteum Kaup, 1829, D. proavum Eichwald, 1835
Deinotherium levius vel giganteum
Deinotherium levius Jourdan, 1861
Lectotype: toothrow with P3/ to M3/ (Lyon, Muséum des Sciences Naturelles, Nr. L.Gr. 962)
Type locality: La Grive Saint-Alban, France (late Middle Miocene)
Deinotherium giganteum Kaup, 1829
Holotype: Left mandible with tusk, m/2 - 3, right mandible fragment: symphysis with tusk fragment (HLMD Din. 466)
Type locality: Eppelsheim, Germany (Miocene)
Description
The partial deinothere skeleton from Gratkorn (Fig. 1), which is preserved in a disarticulated but roughly associated situation, consists of elements of the vertebral column, of the anterior and posterior limbs, and of some teeth. Most of the bones are fragmentary. This partial skeleton represents one individual, while a second individual can be identified by some additional cheek teeth fragments found 30 m NW of the skeleton.
With not fully fused epiphyses in longbones and permanent, lightly worn dentition, the partial skeleton represents a not fully grown “young” adult. It could already have reached sexual maturity. A delayed fusion of the longbones and continuation of growth beyond sexual maturity has been observed in the modern Loxodonta africana (Poole 1996; in males even till the age of 30–45 years).
Dentition and cranial material
Dental remains comprise ten teeth of one individual (P4/ sin., P4/ dex., M1/ sin., M2/ sin., M2/ dex., M3/ sin., i/2 dex.?, p/4 sin., m/1 dex., m/3 sin.) and one p/4 sin., with some cheek teeth fragments (UMJGP 203420, 203421, 203460) of a second individual. A poorly preserved fragment of a pneumatized (?) bone (UMJGP 203654) of the skull cannot be described in detail due to limitations of preservation.
Upper dentition
P4/ (P4/ sin.: UMJGP 204078; P4/ dex.: UMJGP 203690; Fig. 2a–c): P4/ sin. enamel damaged anterobuccally, P4/ dex. enamel damaged posterobuccally, both slightly worn. Subrectangular in occlusal view being wider than long; bilophodont; protoloph complete (reaching paracone); metaloph incomplete (no contact with metacone); ectoloph complete with moderate ectoflexus; blunt postprotocrista weak and short; praehypocrista moderate and crenulated; median valley open lingually; anterior cingulum strong ascending at paracone and forming a well-developed cone; posterior cingulum strong, ascending both to hypo- and metacone (fusion with ectoloph posterior to metacone); small posterobuccal cingulum present at metacone; three roots.

Cheek teeth of D. levius vel giganteum from Gratkorn in occlusal view and with dental terminology. a P4/ dex. (UMJGP 203690), b sketch of c with terminology used for upper premolars, c P4/ sin. (UMJGP 204078), d p/ 4 sin. (UMJGP 203670), e sketch of d with terminology used for lower premolars, f sketch of g with terminology used for upper molars, g M1/ sin. (UMJGP 204081), h m/ 1 dex. (UMJGP 203669), i sketch of h with terminology used for lower molars, j M2/ sin. (UMJGP 204079), k m/ 3 sin. (UMJGP 203689), l M2/ dex. (UMJGP 203628), m M3/ sin. (UMJGP 204080)
Comparison: After Gräf (1957) a P4/ with fused metaloph and ectoloph is typical for D. levius. In the Gratkorn specimen, metaloph and ectoloph are not fully fused, but with a fused protoloph and a clearly developed praehypocrista they show a similar pattern as described by Huttunen (2000) for D. giganteum from Mannersdorf near Angern (NHMW2000z0013/000; Austria; Late Miocene; Pannonian H/F), which is slightly larger in dimensions than the latter or than the range for D. levius given by Gräf (1957) or Pickford and Pourabrishami (2013). D. levius from St. Oswald near Gratwein (Middle Miocene) described by Mottl (1969, fig. 3) is heavily worn, but shows a metaloph not fully fused with the ectoloph as well. It is smaller in dimensions than generally observed for D. levius. Meta- and ectoloph are also not fully fused in a Middle Miocene Deinotherium specimen from Massenhausen (SNSB-BSPG 1951 I 47), which should be D. levius following Gräf (1957), and not in all figures for D. levius given by Depéret [1887; see, for example, D. levius from La Grive (late Middle Miocene) figured on pl. 20, fig. 3]. Tobien (1988) observed fusion and non fusion of ecto- and metaloph, as well as variability in the presence of a well-developed praehypocrista for D. giganteum from Montredon (Late Miocene). Antoine (1994) and Ginsburg and Chevrier (2001) describe a rectangular shape and a weak ectoflexus as being typical for P. bavaricum, a trapezoid shape and a strong ectoflexus for “D. giganteum” (including D. levius). As shape and ectoflexus vary in D. giganteum from Montredon (Tobien 1988) and as, for example, a P4/ of P. bavaricum from Sprendlingen 2 (SSN12SP10; Middle Miocene) shows a stronger ectoflexus than the specimens from Montredon figured by Tobien (1988), this feature is considered variable as well. Therefore, we agree with Huttunen (2000) that a certain variability concerning the fusion of lophs in the P4/ exists and that the morphology of the P4/ does not provide a significant feature for species separation.
M1/ (M1/ sin.: UMJGP 204081; Fig. 2f–g): slightly worn, incomplete, missing anterior and lingual wall of protoloph, buccal cone of tritoloph damaged posterobuccally. Subrectangular shape and longer than wide; trilophodont; all three lophs complete and concave posteriorly; tritoloph linguobuccally less wide than protoloph and metatoloph; buccal posterior cristae (postparacrista, postmetacrista and posterior crista of the buccal cone of the tritoloph) short and pointing posteriorly; blunt lingual posterior cristae (postprotocrista, posthypocrista and posterior crista of the lingual cone of the tritoloph) pointing posteromedian; praecrista only present at metacone (very weak) and at buccal cone of tritoloph, running anteriorly and contacting postmetacrista at its base; anterior valley anteroposteriorly wider than the posterior and with a small tubercle at its buccal side; buccal cingulum present ascending occlusally at cones; posterior cingulum descends from lingual to buccal ascending at buccal cone of tritoloph.
Comparison: Due to fragmentation it cannot be verified whether the metaloph in M1/ is wider than the protoloph on the Gratkorn specimen, which would be characteristic for D. levius after Gräf (1957), but seems to be more variable following the observations of Tobien (1988) and Huttunen (2000). Comparable to the specimen from Gratkorn, for a specimen from St. Oswald near Gratwein (Middle Miocene) Mottl (1969) observed a stronger incision on the buccal wall between protoloph and metaloph than between metaloph and tritoloph, which she states as common for D. levius from La Grive (late Middle Miocene) but less common in D. giganteum. Indeed, the incision is more pronounced in figures of D. levius from La Grive (Depéret 1887, pls. 18–20), and can be observed as strong only in one single specimen of D. giganteum figured by Tobien (1988, pl. 2, fig. 9) from Montredon (Late Miocene), but comparably strong in specimens from Massenhausen (SNSB-BSPG 1951 I 47; late Middle Miocene) and Hinterauerbach (SNSB-BSPG 1951 I 90; late Middle Miocene). The more developed incision between proto- and metaloph seems to be more common in D. levius, but is variable in its extant as well in D. levius [see, for example, SSN12SP15 and 16 from Sprendlingen 2 (Middle Miocene)]. The morphology of the M1/ thus makes an assignation to D. levius more likely but does not exclude a determination as D. giganteum.
M2/(anterior part of M2/ sin.: UMJGP 204079; M2/ dex.: UMJGP 203628; Fig. 2j, l): both slightly worn, M2/ sin. incomplete (only anterior half preserved), M2/ dex. incomplete (anterolingual quarter missing). Subquadratic shape in occlusal view; bilophodont; lophs complete and concave posteriorly; postparacrista pointing posterior and crenulated; postmetacrista long and pointing posteromedially, crenulated as well; weak praemetacrista present, connected to postparacrista at its base; blunt postprotocrista long and pointing posteromedially; posthypocrista short and pointing posteriorly; weak ridge present posterior to metaloph at lingual side on top of large but weak elevation pointing posterobuccally and fusing with postmetacrista by forming a small convolute and enclosing a clear depression anterior to it; anterior and posterior cingula strong; anterior cingulum ascends slightly at protocone forming a small elevation, but ascends strongly at paracone forming a pronounced apex; posterior cingulum descends from lingual to buccal ascending at metacone forming a small apex; posterior cingulum ascends lingual at hypocone; weak lingual cingulum.
Comparison: The postmetaloph morphology of the M2/ dex. from Gratkorn fits well in the description of Gräf (1957) for D. levius and to D. levius from Sprendlingen 2 (MNHM PW2013/29-LS; Middle Miocene). With a clearly present (though small) convolute and the stronger postmetaloph incision it clearly differs from the specimen assigned to D. giganteum by Gräf (1957) from Frohnstetten (GPIT/1035; Late Miocene). Mottl (1969) describes as well the presence of a convolute in specimens from St. Oswald near Gratwein (Middle Miocene), but the posthypocrista in the specimens she figures (Mottl 1969, pl. 3, fig.2) is more strongly developed than in the specimen from Gratkorn. Huttunen (2000) showed that the morphology of the postmetaloph is highly variable, that it does not significantly change with tooth size and that all morphological variations are recorded in teeth of lengths 59–88 mm. Tobien (1988) even observed an intraindividual variation for D. giganteum from Montredon (Late Miocene) concerning this feature (see, for example, Tobien 1988, pl. 4). Thus, the morphology of M2/ cannot be used at the moment for species determination of the Gratkorn specimen.
M3/ (M3/ sin.: UMJGP 204080; Fig. 2m): not worn (tooth germ), enamel missing at protocone. Trapezoid (widening anteriorly) shape in occlusal view, wider than long; bilophodont; lophs complete and concave posteriorly; protoloph linguobuccally wider than metaloph; postparacrista long, crenulated, and pointing posteriorly; postmetacrista long, crenulated, pointing posteromedially, and terminating at midline of tooth; postprotocrista and posthypocrista short, crenulated and pointing posteriorly; lingual half of posterior wall of protoloph and metaloph with blunt elevation; anterior and posterior cingulum present (anterior more strongly developed); anterior cingulum slightly ascending at protocone forming a small elevation but stronger at paracone forming a pronounced apex; anterior cingulum ascending lingually at protocone; posterior cingulum descending from lingual to buccal ascending at metacone twice forming two small peaks; weak lingual cingulum.
Comparison: The M3/ from Gratkorn strongly resembles D. giganteum from Frohnstetten (GPIT/1035; Late Miocene) but also D. levius from Sprendlingen 2 (SSN12SP22; late Middle Miocene). It differs from the specimen from St. Oswald near Gratwein (Middle Miocene) by a less strongly developed posthypocrista (see e.g. Mottl 1969, pl. 3, fig. 3). Gräf (1957) described a long postmetacrista turning to anterior at midline and tapering in the postmetaloph valley parallel to the posthypocrista as typical for D. levius. Tobien (1988) did not observe such a long postmetacrista for D. giganteum from Montredon (Late Miocene) and considered it a typical feature for D. levius as well. In any case, the specimens of D. giganteum figured by him (Tobien 1988, pl. 4 and 5) resemble more closely D. levius from Hinterauerbach (SNSB-BSPG 1951 I 90; late Middle Miocene) than the specimen from Gratkorn. In Depéret (1887) the extension and morphology of the postmetacrista seem to vary as well (see, for example, D. levius from La Grive (late Middle Miocene; Depéret 1887, pl. 18, fig. 1 and pl. 20, fig. 3). We thus consider the development of the postmetacrista not useful as a diagnostic feature for the determination of the Gratkorn specimen.
Lower dentition
tusk (i/2 dex.?: UMJGP 203624; Fig. 3): basal part of lower tusk including deep pulpa, very fragmentary, missing tip and complete caudal wall. Basal ovoid cross section [maximal diameter (DAP) of 90–100 mm reconstructed) with a shallow longitudinal furrow along the lateral side; flattened medial side; no enamel band; no “guillochage”.

Lower tusk (i/2 dex.?) (UMJGP 203624) in caudal (a) and rostral view (b). Scale bar 20 mm
Comparison: As typical for Deinotheriidae the tusk does not possess an enamel band and no “guillochage” (Göhlich 1999; Duranthon et al. 2007). In terms of its size it fits well with D. levius or giganteum (see, for example, values in Duranthon et al. 2007). As it is only a fragment of a young adult and diameters of tusks are highly variable among the two genera [for comparison, see, for example, diameter for P. bavaricum from Unterzolling (early Middle Miocene) in Huttunen and Göhlich (2002)], the assignation is mainly based on the association with the specimen.
p/4 (p/4 sin.: UMJGP 203670; Fig. 2d–e): slightly worn. Subrectangular shape longer than wide; bilophodont; metalophid and hypolophid complete and concave anteriorly, the latter being more straight and slightly longer than the first; ectolophid low and descending anteriorly; strongly crenulated paracristid ascending lingually and ending in anterior cingulid; cingulid present anterobuccal of paracristid; posterior cingulid straight and low and fusing with weak posthypocristid; low buccal cingulid at median valley; two roots.
p/4 sin.: UMJGP 203435 (isolated tooth from different specimen): very fragmentary, smaller and stronger worn than UMJGP 203670.
Comparison: In the p/4, the reduced metalophid compared to the hypolophid is used as a character by Gräf (1957) to distinguish D. giganteum from D. levius [although her values for D. giganteum vary between 87.9 and 98.9 % and therefore overlap with D. levius (99.4–103.2 %)]. The Deinotherium from Gratkorn fits well in morphology with D. levius from Sprendlingen 2 (MNHM PW2013/28-LS, SSN12SP34; Middle Miocene) and to the specimen from Dietersdorfberg near Mureck (UMJGP 3699; late Middle Miocene; see also description in Mottl 1969) but differs from the specimen from St. Oswald near Gratwein (Middle Miocene; Mottl 1969, pl. 4, fig. 1) by a less wide hypolophid and from one specimen from Oberdorf near Weiz (UMJGP 9641; late Middle Miocene) by a less wide metalophid. D. giganteum from Montredon (Late Miocene; Tobien 1988) shows a relatively wide metalophid in the p/4 of some specimens. Duranthon et al. 2007 observed that a trapezoid shape is more frequent in D. giganteum than in P. bavaricum. Comparing different specimens of P. bavaricum (e.g. SNSB-BSPG 1952 I 36; SNSB-BSPG 1959 XIII 12; GPIT/1035-34 and 37) and D. levius (SNSB-BSPG 1951 I 90) with specimens of D. giganteum figured by Tobien (1988), it can be observed that the ratio of meta-/hypolophid width is variable and does not show any significant differences between the species. Furthermore, Tobien (1988) showed a more or less constant ratio between metalophid and hypolophid width (with higher variability for D. giganteum; Tobien 1988, fig. 6). We therefore agree with Huttunen (2000), who observed no morphological change for this tooth position.
m/1 (m/1 dex.: UMJGP 203669; Fig. 2h–i): slightly worn, damaged anterobuccal wall of metalophid and posterolingual wall of tritolophid. Trilophodont; elongated anteroposterior in occlusal view with maximal width at second lophid; all three lophids concave anteriorly; blunt praeprotocristid, praehypocristid and anterior cristid of buccal tritolophid conid pointing anteromedially; praehypocristid ending in small tubercle; anterior cingulid weak; posterior cingulid well pronounced; both valleys open on both sides, deeper at buccal sides; two roots.
Comparison: The feature on m/1 for distinguishing D. levius and D. giganteum given by Gräf (1957; length of posterior cristid/length of tritolophid) cannot be verified on the specimen from Gratkorn as the latter misses the posterior cristid. Taking into consideration the observations of Tobien (1988) for D. giganteum and of Huttunen (2000) for Deinotherium from Lower Austria, the ratios seem to show a greater overlap than expected by Gräf. Duranthon et al. (2007) observed a tendency of tritolophid enlargement from P. bavaricum to D. giganteum. Though varying as well, a general tendency can be observed upon comparison of the different specimens of the species with the specimen from Gratkorn (though fragmented), fitting well with D. levius from Hinterauerbach (SNSB-BSPG 1951 I 90; late Middle Miocene) and Massenhausen (late Middle Miocene).
m/3 (m/3 sin.: UMJGP 203689; Fig. 2k): not worn (tooth germ). Elongated widening anteriorly in occlusal view being longer than wide; bilophodont; lophs complete and concave anteriorly; metalophid linguobuccally wider than hypolophid; praeprotocristid and praehypocristid crenulated, long, and pointing anteromedially; praehypocristid longer than praeprotocristid; praemetacristid and praeprotocristid pronounced, mirror-inverted, both descending in a curve pointing medially recurving anteriorly to lingual and buccal side, respectively; praeentocristid pronounced but short pointing anteriorly; median valley deeper at buccal side; anterior cingulid low and very weak with small peak at buccal side; posterior cingulid (positioned buccally) strongly developed with a strong apex.
Comparison: In the type of D. giganteum (Kaup 1832; add. pl. I, figs. 3, 5 and pl. IV) the posterior cingulid is wider and not positioned buccally as it is in the Gratkorn specimen. However, based on the figures and observations in Tobien (1988; pl. 3, fig. 20, pl. 5. figs. 23–25) for D. giganteum from Montredon (Late Miocene), the width and position of the posterior cingulid is variable. In comparison to other material from Styria, the m/3 from Gratkorn is similar to the specimen from St. Oswald near Gratwein (Middle Miocene; Mottl 1969, pl. 4, fig. 1), differing only in its less wide hypolophid. The m/3 in the Deinotherium from Breitenhilm near Hausmannstetten (UMJGP 1756; late Middle Miocene) is also similar in morphology to the Gratkorn specimen. UMJGP 1756 was assigned to D. giganteum by Mottl (1969). However, due to the strong wear of the p/3 in the specimen an assignation to D. levius cannot be excluded, and based on its dimensions the specimen is well in accordance with this species as well [see Fig. 6; furthermore, the well-developed anterior cingulid of the p/3 in the specimen points rather to a more primitive evolutionary stage, as it is the case in D. levius (Gräf 1957; Böhme et al. 2012)]. In the specimen from Dietersdorfberg near Mureck (UMJGP 3699; late Middle Miocene) the posterior cingulid is more set off than in the specimen from Gratkorn. As the morphology of the m/3 thus seems to be quite variable, no distinguishing characters can be recognised for species differentiation at the moment, as also observed by Huttunen (2000) and Duranthon et al. (2007).
Postcranial material
Columna vertebralis: Of the vertebral column the atlas, eight fragmentary vertebrae and 12 processus spinosi/arcus vertebrarum are preserved (Fig. 4).

Elements of vertebral column of D. levius vel giganteum from Gratkorn. a Atlas in cranial view (UMJGP204654), b vertebra cervicalis in cranial view (UMJGP 203605), c vertebra cervicalis in cranial view (UMJGP 204111), d vertebra cervicalis in cranial view (UMJGP 203623), e vertebra thoracica or lumbalis (UMJGP 203659), f vertebra thoracica or lumbalis (UMJGP 203653), g fragment of vertebra caudalis? (UMJGP 203663), h processus spinosus of vertebra cervicalis 6 or 7 (UMJGP 204681), i processus spinosus of vertebra thoracica from cranial series (UMJGP 203602), j processus spinosus of vertebra thoracica from cranial series (UMJGP 203603). Scale bar 10 cm (a–f, h–j), 1 cm (g)
Atlas (UMJGP 204654; Fig. 4a): poorly preserved; relatively wide arcus vertebrae; on cranial side two suboval foveae articulares craniales for the articulation with the occipital condyles still visible; dorsal of articulation facets depression on each side; lateral median walls of foramina transversaria still observable.
Comparison: The atlas from Gratkorn is similar in dimensions to D. giganteum from Brunn-Vösendorf (Austria; Late Miocene; Pannonian E; MN 9) described by Huttunen (2000), to D. levius from Gusyatin (Middle Miocene; Svistun 1974) and to the specimen from Holzmannsdorfberg (UMJGP 61634; Austria; Late Miocene; Pannonian C/D; MN 9), but it is clearly larger than Prodeinotherium from Langenau (Early Miocene). Due to poor preservation, a morphological comparison is not possible.
In addition to the atlas, eight further vertebrae (more or less badly preserved) could be identified. Following comparisons with the skeletons of Prodeinotherium from Franzensbad and Langenau (both Early Miocene) and the descriptions of Göhlich (1998) and Huttunen and Göhlich (2002), these vertebrae remains were tentatively identified as cervicales, thoracicale or lumbales. UMJGP 203623, 204111, 203605 comprise vertebrae cervicales (Fig. 4b–d): corpora vertebrarum relatively large and craniocaudally flat (enhanced flattening likely due to sediment compaction) as typical for vertebrae cervicales, comprising more or less preserved arcus vertebrarum; UMJGP 203605 still showing convex right cranial articulation facet, concave, kidney-shaped and caudoventrally facing right caudal articulation facet, and a nearly complete arcus vertebrae; basal part of processus spinosus recognisable as being cranially convex and caudally concave; UMJGP 204111 more poorly preserved, slightly larger than UMJGP 203605, with complete arcus vertebrae and both kidney-shaped caudal articulation facets still preserved; foramen vertebrae possibly slightly higher dorsoventrally than in UMJGP 203605; concave base of processus spinosus inclined cranially; UMJGP 203623 largest and best preserved vertebra cervicalis with both the convex cranial articulation facets facing craniomedially (axis inclined medially) and concave caudal articulation facets facing laterally; UMJGP 203638, 203653 (with small bone fragment), 203659, 203680 represent vertebrae thoracicae or lumbales (Fig. 4e–f): smaller corpus vertebrae than in vertebrae cervicales with a subtriangular (UMJGP 203638, 203659, 203680) to transverse-oval shape (UMJGP 203653) and less flattened craniocaudally than vertrebrae cervicales; UMJGP 203663 badly preserved and quite small, but due to its transversal subrounded shape and its small cranial caudal width it could be a fragment of a vertebra caudalis (non-fused extremitas; Fig. 4g).
Several more or less fragmented processus spinosi (Fig. 4h–j) could be tentatively assigned to certain parts of the vertebral column: processus spinosus of vertebra cervicalis 6 or 7 (UMJGP 204681; Fig. 4h): slender processus spinosus [assigned to caudal part of cervical vertebral column due to length and slender habitus and based on comparison with the skeleton of Prodeinotherium from Langenau (Early Miocene) and figures in Huttunen and Göhlich (2002)]; in cross section triangular (pointing anterior); only slight cranial inclination (nearly vertical); fragment of processus spinosus of vertebra cervicalis 7 or vertebra thoracica 1 (UMJGP 203693): slender and similar in dimensions to UMJGP 204681 but with stronger developed triangular cross section, more pronounced cranial crest and more concave caudal side [following Huttunen and Göhlich (2002) the processus spinosi become more concave from caudal part of cervical vertebrae to cranial part of thoracic vertebrae]; processus spinosi of vertebrae thoracicae from cranial series {UMJGP 203642, 203655, 203649 [with fragment of arcus vertebrae (? UMJGP 203646)], 203647, 203602, 203694 and 203603}: mediolaterally wider than processus spinosi of vertebrae cervicales; ordered from cranial to caudal due to increase in mediolateral width [in accordance with the skeleton of Prodeinotherium from Langenau (Early Miocene)]: processus spinosus of vertebra thoracica 1 or 2 (UMJGP 203642): with small fragment of right arcus and fragmented right processus lateralis; processus spinosus with triangular cross section, caudally slightly concave and decreasing in mediolateral width from proximal to distal (minimum preserved width distally: 30 mm); other processus spinosi of vertebrae thoracicae from cranial series {UMJGP 203655, 203649 [with fragment of arcus vertebralis (? UMJGP 203646)], 203647, 203602, 203694 and 203603} strongly increase in mediolateral width; craniocaudally flattened; longitudinal crest along the midline on the cranial surface opposed by a concave caudal surface; cranial crest more pronounced in UMJGP 203655 and 203602; mediolateral width and dorsoventral height of arcus vertebrae increases from UMJGP 203602 (Fig. 4i) to 203603 (Fig. 4j); UMJGP 203603 caudally not concave but with crest; fragment of one processus spinosus with clear bite mark (UMJGP 203694). Further fragments of processus spinosi [UMJGP 203687(?)] and arcus vertebrarum [UMJGP 203681 (?), UMJGP 204684(?), UMJGP 203716, UMJGP 203675(?)] are preserved but cannot be assigned to specific vertebrae due to fragmentary preservation and do not allow any detailed description.
Costae: Most costae are fragmentary and allow no specific diagnosis [UMJGP 203604, 203608, 203610 (two crushed fragments?), 203634, 203643, 203644, 203648 (with fragment 203645), 203660 (?), 203687, 203696, 203692, 203697, 203703, 203717, 203666, 203658, 203629, 203630, 203635, 203617, 204673 (?)]. They were assigned to the Deinotherium skeleton due to their large dimensions and their finding position. Eleven costae were more complete and could be determined as elements of the cranial [UMJGP 203657 (costa 1/2? dex.), UMJGP 203606 (costa 2/3? dex.), and central-caudal part of the thorax (costae dex.: UMJGP 203639, 203650, 203695, 203633; costae sin.: 204110, 203631, 203618 and 203614-5 (fragment of the same rib), 203632, 203607]. Costae 1/2? and 2/3? in contrast to more caudal costae less curved but straight and shorter, craniocaudally flattened (stronger distal than proximal) and mediolaterally expanded, widening distally; cross section of costa 1/2? (UMJGP 203657) proximally ovoid (pointing caudolaterally) to distally strongly flattened and more acute caudolaterally; costae of central to caudal part of thorax decrease in mediolateral width from cranial to caudal (UMJGP 203639 mediolaterally wider than UMJGP 203695) and gain a more rounded cross section from cranial to caudal; on the proximal part of corpus costae more or less developed sulcus costae on the cranial side and crest on caudal side; on craniolateral side ellipsoid shaped plane surface developed; sulcus costae more pronounced along distal part of corpus on caudal plane; costae mediolaterally flattened distally.
Scapula: represented by several blade-like bone fragments, the largest being 100–200 mm [UMJGP 203662, 203664, 203667, 203668, 203671, 203672, 203676, 203677, 203678(?), 203679, 203691, 204103]. The affiliation to the scapula is due to the flatness and rather constant thickness (5–25 mm) of the bone-blades and due to their finding position (Fig. 7). All fragments are supposed to represent a single scapula, although completely compressed and fractured. No anatomical details or diagnostic characters are preserved. An additional, small blade-like bone fragment, probably also belonging to the scapula, is attached to the humerus fragment (UMJGP 203674). On fragment UMJGP 204103 chewing marks are preserved.
Fragment of humerus dex.? (UMJGP 203674): very fragmentary, with plane surface on one side and convex one on the other; epiphyseal surface on plane side; in size and morphology the convex bone fits best to a proximal articulation surface of a humerus; due to poor preservation a more detailed description and reasonable affiliation not possible.
Radius (radius sin. missing distal end (UMJGP 203665; Fig. 5d): radius dex. proximal fragment with articulation facet for humerus (UMJGP 203621)): slender, tapering proximally and bent concave laterally; distal half of corpus radii mediolaterally flattened; cross-section at level of collum subtriangular; torsion of radius not very pronounced; caput radii subtriangular in proximal view; collum radii with pronounced incision dorsally; proximal articular facet for humerus subdivided in two slightly concave facets, facing proximolaterally and proximomedially, and enclosing an obtuse angle (Fig. 5d1); lateropalmar on caput radii large triangular facet for articulation with ulna (Fig. 5d1; due to preservation no detailed description can be given, though) distally bordered by a ridge running from lateroproximal to mediodistal; medial and lateral tuberosity on collum radii; distal to facet for the ulna on the lateropalmar side of the diaphysis longitudinal depression extending distally, becoming less deep in the middle part of the bone but deepening and widening again more distally; minimum width of the corpus radii in dorsal view in its middle part, broadening both distally and proximally.

Elements of anterior and posterior limbs of D. levius vel giganteum from Gratkorn with affiliation of articulation facets: a os carpi ulnare sin. [UMJGP 203688; 1 proximal view (os pisif. = os pisiforme), 2 distal view, 3 dorsal view, 4 medial view], b os carpale secundum sin. [UMJGP 203640; 1 lateral view, 2 sketch of lateral view with identified articulation facets for os carpale tertium (III), 3 dorsal view (articulation facet for os carpale primum on medial side)], 4 proximal view with articulation facet for ossa carpi radiale and intermedium, c phalanx proximalis of pes? (UMJGP 203708; 1 dorsal view, 2 plantar view, 3 lateral/medial view), d radius sin. (UMJGP 203665; 1 lateropalmar view, 2 mediodorsal view); e fibula sin. (UMJGP 203622; 1 lateroplantar view, 2 mediodorsal view), f fragments of femur dex. in caudal view with sketch of outline (fragment of proximal shaft: UMJGP 203601; distal epiphysis: UMJGP 204112), g os tarsi centrale dex. [UMJGP 203683; 1 proximal view, 2: distal view, 3 sketch of distal view with identified articulation facets for os tarsale secundum (II), tertium (III) and quartum (IV)]. Scale bar 5 cm (a–c, g), 10 cm (d–f)
Comparison: The radius sin. (UMJGP 203665) is mediodorsal-lateropalmar more flattened at the proximal diaphysis than in P. bavaricum from Franzensbad (Early Miocene) or Unterzolling (early Middle Miocene; Huttunen and Göhlich 2002) which show a more triangular proximal diaphysis. In overall shape, the radius from Gratkorn stronger resembles that of D. proavum from Ezerovo (Late Miocene) mounted at the University of Sofia. With the latter it also shares the generally more flattened corpus radii and the reduced torsion. Svistun (1974) unfortunately does not give any information concerning the degree of the torsion of the radius in comparison to other species. Though varying in its extent [in the specimen from Langenau (Early Miocene) it is more weakly developed than in the specimens from Unterzolling and Franzensbad] the torsion of the radius in the genus Prodeinotherium is stronger than in the Gratkorn specimen and in other specimens of Deinotherium.
Os carpi ulnare sin. (UMJGP 203688; Fig. 5a): quite large with pronounced lateropalmar processus (mostly broken off); proximal articulation surface for ulna large, subtriangular (pointing palmar) and dorsopalmar concave with a slightly convex medial half and a slightly concave lateral half (Fig. 5a1); triangular articulation facet for os pisiforme located at the lateral half of palmar surface and extending on lateral processus, facing lateropalmar forming a right angle with the proximal facet and tapering off medially (Fig. 5a1); distal articulation facet for articulation with os carpale quartum (damaged laterally) comprising two concave facets (axes dorsopalmarly) divided by central convexity (Fig. 5a2); due to fragmentariness of lateral processus only small part of articulation facet for Mc V preserved distally on the process, separated from distal facet by a distinct ridge; medial surface with a proximal and a distal longitudinal facet for articulation with os carpi intermedium (Fig. 5a4).
Comparison: The distal surface of the os carpi ulnare comprises two concave facets (axes dorsopalmarly) divided by central convexity as observed in Deinotherium from Paasdorf near Mistelbach (NHMW; Austria; Late Miocene) and described by Svistun (1974) for D. levius from Gusyatin (Middle Miocene). Following Huttunen (2000) this is typical for the genus. It can be distinguished from the concavo-convex or concave distal surface in Prodeinotherium (Huttunen 2000; Huttunen and Göhlich 2002).
Os carpale secundum sin. (UMJGP 203640; Fig. 5b): triangular shaped in proximal and distal view, narrowing palmarly (here damaged); proximal articulation facet for os carpi radiale and intermedium large and triangular, concave and tapering palmarly; facet for carpi radiale and intermedium enclosing an obtuse angle with facet for os carpale tertium; distal articulation facet for Mc II slightly convex (preserved only medially, damaged laterally); medial side damaged palmarly; round (three-quarters of circle), and slightly convex facet for articulation with os carpale primum on dorsodistal quarter of medial side (enclosing a nearly right angle with distal articulation facet); on lateral side three facets for articulation with the os carpale tertium not well preserved but still recognisable (Fig. 5b1, b2): large facet located proximodorsally, semicircular facet in proximopalmar part, only a small portion of the elongated distal facet preserved.
Comparison: Comparison material for the os carpale secundum consisted of one specimen of D. cf. giganteum from Wien XII Oswaldgasse (NHMW SK 2810; Austria; Late Miocene; Pannonian E; 10.4–10 Ma), which is larger and differs morphologically from the Gratkorn specimen by a less rounded dorsal side and the facet for articulation with os carpale primum, which comprises only a semi circle in the specimen from Wien XII Oswaldgasse. Following the description by Svistun (1974) the os carpale secundum of D. levius is in general of similar shape as the Gratkorn specimen but differs from the latter as it seems to possess only two facets for the articulation to the os carpale tertium.
Distal epiphysis with articulation facet of Mc II or III sin. or IV dex. (UMJGP 203685): due to its relatively large size it can be assigned to the manus rather than to the pes; due to fragmentary preservation most of the articulation facet missing; distal articulation facet dorsopalmar convex with small oblique ridge slightly shifted from the central line on palmar part of the trochlea, but not as asymmetric as it would be expected for Mc V.
Phalanx proximalis? of manus (UMJGP 203684) of unidentified digit: dorsal surface not preserved and phalanx missing its distal part; epiphysis not entirely closed proximally; proximal facet for articulation with metacarpal dorsopalmarily concave with a general inclination to proximopalmar; palmar side convex.
Comparison: Morphology alone does not allow affiliation to manus or pes, but dimensions in comparison with UMJGP 203708 render a determination as phalanx proximalis of manus more likely.
Femur dex. (distal epiphysis (UMJGP 204112), fragment of proximal shaft (UMJGP 203601); Fig. 5f): portion of proximal femur shaft with basis of trochanter minor (distinct depression on shaft caudal of trochanter minor); caudolateral edge of shaft subrectangular at base of trochanter minor; both condyles on distal epiphysis damaged, the articulation surface of the condylus lateralis femoris damaged, except for its caudalmost part; only distal part of the trochlea ossis femoris preserved and showing a deep distal incision between the two condyles widening caudally; pronounced mediolateral depressions proximal to both condyles.
Comparison: Due to fragmentary preservation of the Gratkorn femur no comparison to other specimens can be given.
Fibula sin. (UMJGP 203622; Fig. 5e) and dex. (UMJGP 203613 (proximal portion of shaft without facet) 203612 (distal portion of shaft)): fibula sin. almost complete though lacking proximal and distal articulation facets; corpus fibulae triangular proximally (here smallest circumference); distal half mediolaterally flattened with slightly concave medial side; diagonal crest running from smallest circumference proximodorsally along the lateral side of the proximal fourth of the shaft.
Comparison: The morphological difference concerning the fibula between Prodeinotherium and Deinotherium as observed by Huttunen [“form of shaft proximally flattened in dorsoplantar direction” in Deinotherium (Huttunen 2000, p. 91)] cannot be confirmed based on the specimen from Gratkorn, as the cross section of the proximal shaft is triangular. The proximal cross section of both Gratkorn specimens is not more dorsoplantarily flattened than in Prodeinotherium from Langenau (Early Miocene), but its distal shaft seems to be more flattened mediolaterally than the latter.
Os tarsi centrale sin. (UMJGP 203611) and dex. (UMJGP 203683; Fig. 5g): both ossa tarsorum centralia badly preserved and missing most of dorsal, medial and plantar surfaces; proximal articulation facet for astragalus large, concave and oval shaped (mediolaterally elongated); small, proximoplantar oriented facet for articulation with the calcaneum located in the lateral half of the plantar side forming an obtuse angle with proximal articulation facet; on distal surface three articulation facets for the tarsals II–IV identified (from lateral to medial for os tarsale quartum (oriented distoplantolateral); os tarsale tertium; os tarsale secundum); most medial distal facet for Mt I not traceable, all preserved distal facets slightly concave separated by dorsomedial-plantolateral oriented ridges diverging in dorsomedial direction; no plantomedial process.
Comparison: With only three distal facets and no articulation facet for the Mt I the os tarsi centrale differs from that of Prodeinotherium (which shows four facets) but fits well with the situation in Deinotherium (Huttunen 2000). Furthermore, the os tarsi centrale differs from that of P. bavaricum from Unterzolling (early Middle Miocene) in the lack of a plantomedial process (Huttunen and Göhlich 2002).
Distal trochlea of Mt II? (UMJGP 204696): due to its smaller size in comparison to the Mc described above (UMJGP 203685) trochlea assigned to a metatarsal; allocation of trochlea to digit II based on only slightly asymmetric shape.
? Mt IV dex. (UMJGP 203625): fragmentary assumed metatarsal missing most of the proximal and the complete distal end; elongated rectangular shape in dorsal view; cross section of diaphysis subtriangular widening medially; large trapezoid proximal articulation facet slightly declining laterally and smaller proximal facet (due to preservation shape cannot be reconstructed) declining medially; two facets enclosing an obtuse angle of about 130°; lateral side of shaft with pronounced proximodistal elongated sulcus weakening distally.
Comparison: identification as Mt IV dex. with uncertainty due to fragmentary preservation; overall shape of fragment also fitting to morphology of Mc II and III sin., but comparing dimensions with anterior and posterior metapodials of P. bavaricum from Franzensbad (Early Miocene), determination as Mt IV dex. is more likely.
Lateral fragment of Mc I or Mt I dex.? (UMJGP 203620): missing distal end; proximal articulation facet slightly dorsopalmarly concave and distinctly declining dorsally.
Phalanx proximalis II, III or IV? of pes (UMJGP 203708; Fig. 5c): subquadratic shape in dorsal view with proximal epiphyseal suture not entirely closed; proximal facet for articulation with metatarsal oval and dorsoplantar concave; distal trochlea slightly concave on plantar side; plantar surface concave; dorsal surface more plane.
Comparison: Quite symmetric shape of the phalanx indicates assignment to central digits II, III or IV, affiliation to pes is due to dimension in comparison with UMJGP 203684.
Os sesamoideum (UMJGP 203709 (almost complete; hmax = 61 mm); UMJGP 203710 (only distal half): morphology does not permit affiliation to manus or pes nor to any digit.
An additional small shaft fragment (UMJGP 203616) might represent another metapodial, which is similar in its dimension to UMJGP 203625; shaft with rectangular cross-section and slight concavity on lateral side;
Discussion
In terms of size and morphology, the teeth of the Gratkorn specimen fit well with both medium-sized species D. giganteum and D. levius from the type localities and other well-documented sites (Fig. 6). Differentiation between the two species D. levius and D. giganteum has been in discussion for a considerable time, and the validity of D. levius is often questioned, due to aforementioned supposed morphological, dimensional and stratigraphic overlap with D. giganteum (Huttunen 2002a). Gräf (1957) provided a comprehensive description and comparison of dental material of D. levius and D. giganteum. However, most of the species characteristics for D. levius described by her were shown to be more variable (see also discussions in Bergounioux and Crouzel 1962; Tobien 1988; Huttunen 2000; Pickford and Pourabrishami 2013). Unfortunately, a p/3, so far “the only tooth that has clearly differential morphology in different size classes and different MN Zones” (Huttunen 2000, p. 42; see also discussion in Gasparik 2001), is not preserved from the Gratkorn specimen. This tooth is generally accepted to be species specific (Mottl 1969; Gasparik 2001; Huttunen and Göhlich 2002; Duranthon et al. 2007; Böhme et al. 2012) and distinguishes D. levius (proto- and metaconid separated) and D. giganteum (proto- and metaconid fused) (Gräf 1957; Mottl 1969; Böhme et al. 2012). Gasparik (2001) described in detail the morphology of the p/3 and especially the degree of fusion for proto- and metaconid in the species differentiation he gave for the material from Hungary. He figured a p/3 of “D. giganteum” from Sopron (Hungary; Late Miocene; Pannonian B; MN 9), which shows not fully fused proto- and metaconid (which would be typical for D. levius). Furthermore, measurements for this tooth given by Huttunen (2000) would not contradict an assignation to D. levius. The specimen from Sopron would thus be the youngest representative of the species D. levius, as the locality Sopron, Boór’s sandpit, can be correlated to Pannonian B, based on the occurrence of Melanopsis impressa (Vendl 1930 cited in Thenius 1948). The assumption of Huttunen and Göhlich (2002) that the separation of proto- and metaconid in the p/3 is a typical feature in Prodeinotherium distinguishing it from Deinotherium cannot be confirmed, taking into consideration the p/3s from Massenhausen (e.g. SNSB-BSPG 1955 I 43 and 47; late Middle Miocene), Hinterauerbach (SNSB-BSPG 1951 I 90; late Middle Miocene) and Sprendlingen 2 (Middle Miocene; Böhme et al. 2012), which all show separated proto- and metaconid, but are not in the dimensional variability of Prodeinotherium and should be assigned to D. levius. The separation of proto- and metaconid in the p/3 has thus to be considered a primitive dental character, still present in the oldest representative of the genus Deinotherium, D. levius, but lost in the younger representatives, such as D. giganteum.

Bivariate plots [wmax versus lmax (mm)] of dental material of D. levius vel giganteum from Gratkorn in comparison to other Deinotheriidae: Prodeinotherium bavaricum from Falun de la Touraine and Anjou (both France; early Middle Miocene; Langhian; MN 5; 15 ± 0.5 Ma; data from Ginsburg and Chevrier 2001); P. bavaricum and Deinotherium levius from Sprendlingen 2 (Germany; Middle Miocene; data from Böhme et al. 2012 and own measurements); D. levius from Middle Miocene sites [from France and Germany: St. Gaudens, Tournan (both France; late Middle Miocene; MN 7/8; 13–11.5 Ma); Massenhausen, Hinterauerbach (both Germany; late Middle Miocene; MN 7/8; 13–11.5 Ma; data from Gräf 1957; Ginsburg and Chevrier 2001); D. levius from St. Oswald near Gratwein (Austria; Middle Miocene; early Badenian), Oberdorf near Weiz (Austria; late Middle Miocene; late Sarmatian; 12.2–11.6 Ma) and Dietersdorfberg near Mureck (Austria; late Middle Miocene; Sarmatian; 12.7–11.6 Ma) after Mottl 1969 and own measurements; D. levius from La Grive (France; late Middle Miocene; MN 7/8; 13–11.5 Ma; data from Huttunen 2000) and from Gusyatin (also Husyatyn) (Ukraine; Middle Miocene; early late Badenian; 13.1–13.4 Ma; data from Svistun 1974); D. levius(?) from Opatov (formerly Abtsdorf; Czech Republic; Middle Miocene; Badenian; data from Zázvorka 1940); D. levius(?) from Sopron (Hungary; Late Miocene; Pannonian B/C; MN 9; data from Huttunen 2000); Deinotherium from Breitenhilm near Hausmannstetten (Austria; late Middle Miocene; late Sarmatian; 12.2–11.6 Ma; data from Peters 1871); holotype of D. giganteum from Eppelsheim (Germany; Miocene; data from Gräf 1957) and D. giganteum from Montredon (France; Late Miocene; late Vallesian; MN 10; 9.5 Ma; data from Tobien 1988; Ginsburg and Chevrier 2001)
The skeletal deinothere elements from Gratkorn fit with the larger genus Deinotherium in size and morphology and show some distinct differences from the smaller genus Prodeinotherium. The specimen therefore corresponds well with the genus separation proposed by Éhik (1930). The weak torsion of the radius, a mediodorsal-lateropalmar flattened proximal diaphysis and the generally more flattened corpus radii are typical of Deinotherium and distinguish the radius from that of Prodeinotherium (Huttunen 2000 and personal observation). The distal articulation facet of the os carpi ulnare comprises two concave facets (axes dorsopalmarly) divided by a central convexity in the Gratkorn specimen and is not flat concave like in Prodeinotherium (Huttunen 2000). Furthermore, the Gratkorn specimen shares an os tarsi centrale with only three distal articulation facets and no facet for the articulation with the Mt I with Deinotherium, whereas Prodeinotherium shows four distal articulation facets (Huttunen 2000).
Summing up, from size and dimensions of the postcranial elements the specimen from Gratkorn fits well to the larger genus Deinotherium. As the teeth show most dimensional and morphological overlap with D. levius, which is described from other localities of the same age, it most likely represents this species. However, it cannot be clearly distinguished from D. giganteum due to the absence of the diagnostic p/3 and it is thus determined as Deinotherium levius vel giganteum.
Ecology
In contrast to the bunodont gomphotheres, deinotheriids with their more primitive lophodont dentition, are considered to represent typical browsers (Harris 1975) well adapted to the consumption of soft foliage (Göhlich 1999). Calandra et al. (2008) showed that in comparison to two different Gomphotherium species, D. giganteum fed on less abrasive food. Harris (1975) observed only slight striation on the molar wear facets, which he interpreted as an indication for feeding on soft vegetation, while Calandra et al. (2008) found a higher scratch density on grinding than on shearing facets and therefore assume that each facet had two different functions during mastication. Harris (1996) observed a strict feeding on C3-plants for deinotheres through their evolutionary history, while Miocene gomphotheres in Africa switched from a C3 to a C4-diet (Harris 1996; Huttunen 2000; Lister 2013). Stable isotope analyses (δ18OCO3, δ13C) from Gratkorn (Aiglstorfer et al. 2014, this issue) show a C3-diet for D. levius vel giganteum as well and indicate canopy browsing.
The Gratkorn specimen was a not fully grown “young” adult, but could have reached sexual maturity. Due to the fact that most deinotheres occur as isolated finds or in fluviatile accumulated (and often stratigraphically mixed) assemblages, estimations on sexual dimorphism in terms of general size and tusk dimensions cannot be given so far (see also Huttunen 2000 for discussion). Therefore, gender determination for the partial skeleton from Gratkorn cannot be assessed.
Following estimations of Christiansen (2004) a body mass of about 6 t was calculated for the Gratkorn specimen based on the minimal circumference of the radius. As the animal was not fully grown lower values than for a fully grown specimen would be expected. However, this weight estimation has to be considered rather as a minimum value as it is based on modern elephants and following Christiansen (2007) can be applied to primitive proboscideans with reservations only [with more elongate bodies they could have reached higher body masses with the same shoulder height than the more compact modern elephants (Christiansen 2007)]. Other body mass estimations for Deinotherium giganteum vary between 11 t [Fortelius 2013 (NOW database)] and 19 t (representing an assumed fully-grown specimen; Merceron et al. 2012). In any case, Deinotherium levius vel giganteum was by far the largest herbivorous mammal at the Gratkorn locality.
The significantly different Sr87/Sr 86 values in D. levius vel giganteum from Gratkorn in comparison to the local mammal fauna indicate that it was not a permanent resident of the locality but had a different habitat, such as the Styrian Basin, at least during tooth enamel formation (Aiglstorfer et al. 2014, this issue). Migration was most likely necessary for the animal as the environs around the Gratkorn locality presumably could not provide enough biomass during all seasons to support such a large animal. Comparable to modern elephants (Galanti et al. 2006), the Deinotherium from Gratkorn thus presumably had a large habitat range.
Taphonomy
The partial skeleton of Deinotherium levius vel giganteum is spread over an area of about 140 m2, with most of the material concentrated in the northern 50 m2 (Fig. 7). Rough anatomical associations are preserved in some cases, such as the assemblage of posterior extremities comprising both fibulae, os tarsi centrale dex. and metatarsals in the western part of the excavation. Most costae are accumulated in the central part, and fragments of scapula, humerus, radius and os carpi ulnare sin. are deposited in the eastern part. Teeth of the sinistral upper jaw (though dislocated from the rest) or dextral part of the skull and mandible are still roughly associated as well, while the sinistral lower jaw is torn apart, as is the dextral femur, of which two parts have been excavated more than 6 m apart. Besides the sinistral upper jaw, the atlas is dislocated from the rest of the skeleton by more than 6 m. The rough association of the specimen and the lack of long bone or rib alignment indicate no significant water transport of the carcass after death and decomposition, but rather fragmentation, disintegration and finally burial at the actual place of death. Havlik et al. (2014, this issue) were able to show that the large mammal assemblage from Gratkorn was a preferred feeding place for scavengers. Scavenging by carnivores or trampling by large herbivorous mammals (such as, for example, Rhinocerotidae or Deinotheriidae) could explain dislocation and breakage of some skeletal parts. In studies on death and deposition of modern elephants in Africa, dislocation of the long bones of more than 100 m by lions, hyenas or even other elephants was observed (Haynes 1988). Furthermore, African elephants show a high degree of interest in skulls of their kin, touching them with trunks or feet, turning them over or even carrying them away (McComb et al. 2006). The strong demolition and dislocation of the dextral femur could thus simply result from such a treatment through other deinotheres, similar to what has also been described for modern elephants in Shabi Shabi (Zimbabwe; Conybeare and Haynes 1984). The strong breakage of most deinothere bones and biting and chewing marks of carnivores on several bones [e.g. costa fragment with bite marks at distal part (UMJGP 203630), radius sin. with bite marks at lateral tuberosity of collum radii (UMJGP 203665) and chewing marks on fragment of scapula (UMJGP 204103)] fit well with an intense feeding by scavengers on the carcass. The general preservation of most bones of this partial skeleton is rather bad and very fragmentary and shows traits of weathering (see, for example, os tarsi centrale), which indicates no fast burial of the carcass but exposure on the surface for a considerable amount of time.

Excavation plan of the partial Deinotherium skeleton from Gratkorn with identification of skeletal elements (modified after excavation plan by M. Gross (excavations 2005–2008); coordinates are in Austrian Grid (BMN M34 – GK)
One p/4 sin. (UMJGP 203435) and some tooth fragments from lower and/or upper molars (UMJGP 203420, 203421, 203460) were found on the surface about 30 m NW of the partial skeleton. As there is a p/4 sin. preserved from the partial skeleton described above, the second p/4 sin. has to be assigned to a second specimen. Due to the position in the field and the general taphonomic situation (see, for example, Havlik et al. 2014, this issue) it is most likely that the tooth fragments belong to the same individual like the p/4. The tooth remains differ from the teeth assigned to the skeleton by smaller dimensions, and stronger tooth wear (see Table 1 for different lengths of p/4). Sampling for isotopic measurements was done on this second specimen (see Aiglstorfer et al. 2014, this issue).
Conclusions
Besides the partial skeleton from Gusyatin (Middle Miocene; Svistun 1974) the specimen from Gratkorn, though partial, is the only one of a medium-sized deinothere taxon described so far. Other deinothere skeletons recorded are the mentioned Prodeinotherium skeletons from Langenau (Germany; Early Miocene), Franzensbad (Czech Republic; Early Miocene) and Unterzolling (Germany; early Middle Miocene) (Huttunen 2000, 2004; Huttunen and Göhlich 2002), the Prodeinotherium skeleton from Česká Třebová (Czech Republic; Middle Miocene; Badenian; Musil 1997) and the Deinotherium proavum skeletons from Ezerovo (Bulgaria; Late Miocene; Kovachev and Nikolov 2006), from Obuhovka (Russia; Late Miocene; Turolian; pre-Pontian; Bajgusheva and Titov 2006), from Pripiceni (Moldava; Late Miocene; Turolian; post-Bessarabian; Tarabukin 1968) and from Mânzaţi (Romania; Late Miocene; Stefanescu 1894). The assignation of deinothere remains from Opatov (Middle Miocene; Zázvorka 1940; Musil 1997; most likely representing at least two skeletons) to D. levius could not be verified during the investigation for this publication. However, the dental measurements (Zázvorka 1940), fit with a medium-sized deinothere.
With a generally more flattened corpus radii, a mediodorsal-lateropalmar flattened proximal diaphysis and the weaker torsion of the bone, a distal surface on the os carpi ulnare with two concave facets (axes dorsopalmarly) divided by a central convexity, three distal articulation facets and none for the articulation to the Mt I in the os tarsi centrale, the deinothere from Gratkorn fits well in postcranial morphology to the larger genus Deinotherium and clearly differs from the smaller genus Prodeinotherium. It thus confirms well to the genus separation. In dental dimensions the specimen fits with the medium-sized species D. levius and D. giganteum. For most tooth positions it overlaps with the lower dimensional range of D. giganteum and for all positions it nests well in the variability observed for D. levius (Fig. 6). The Gratkorn specimen is thus well in accordance with the gradual size increase observed for European Deinotheriidae mentioned above and most likely represents D. levius. However, due to the lack of a p/3 a distinction from D. giganteum cannot be given, and the specimen is determined as Deinotherium levius vel giganteum. Although the specimen cannot be clearly assigned to a certain species, it is of scientific value. It possesses a clearly defined stratigraphic age and represents one of the rare records of associated postcranial and dental material of a medium sized deinothere taxon.
References
Aiglstorfer M, Bocherens H, Böhme M (2014) Large mammal ecology from the late Middle Miocene locality Gratkorn (Austria). In: Böhme M, Gross M, Prieto J (eds) The Sarmatian vertebrate locality Gratkorn, Styrian Basin. Palaeobio Palaeoenv 94(1). doi 10.1007/s12549-013-0145-5
Antoine PO (1994) Tendances évolutives de Deinotheriidae (Mammalia, Proboscidea) miocènes du domaine sous-pyrénéen. Master thesis. Université Paul-Sabatier, Toulouse
Bajgusheva SV, Titov V (2006) About teeth of Deinotherium giganteum Kaup from eastern paratethys. Hellenic J Geosci 41:177–182
Bergounioux F-M, Crouzel F (1962) Les déinothéridés d’Europe. Ann Paléontol 48:13–56
Böhme M, Aiglstorfer M, Uhl D, Kullmer O (2012) The antiquity of the Rhine river: stratigraphic coverage of the Dinotheriensande (Eppelsheim Formation) of the Mainz basin (Germany). PLoS ONE 7(5):e36817
Calandra I, Göhlich UB, Merceron G (2008) How could sympatric megaherbivores coexist? Example of niche partitioning within a proboscidean community from the Miocene of Europe. Naturwissenschaften 95(9):831–838. doi:10.1007/s00114-008-0391-y
Christiansen P (2004) Body size in proboscideans, with notes on elephant metabolism. Zool J Linn Soc 140(4):523–549. doi:10.1111/j.1096-3642.2004.00113.x
Christiansen P (2007) Long-bone geometry in columnar-limbed animals: allometry of the proboscidean appendicular skeleton. Zool J Linn Soc 149(3):423–436. doi:10.1111/j.1096-3642.2007.00249.x
Codrea V (1994) A priority issue: Deinotherium proavum Eichwald or Deinotherium gigantissimum Stefanescu? In: Petersen (ed) The Miocene from the Transylvanian Basin. Cluj-Napoca, Romania, pp 105–110
Conybeare A, Haynes G (1984) Observations on elephant mortality and bones in water holes. Quat Res 22(2):189–200. doi:10.1016/0033-5894(84)90039-5
Dehm R (1960) Zur Frage der Gleichaltrigkeit bei fossilen Säugerfaunen. Int J Earth Sci 49(1):36–40
Depéret C (1887) Recherches sur la succession des faunes des vertebres miocenes de la vallee du Rhone. Arch Mus Natl Hist Nat (Lyon) 4:45–313
Duranthon F, Antoine PO, Laffont D, Bilotte M (2007) Contemporanéité de Prodeinotherium et Deinotherium (Mammalia, Proboscidea) à Castelnau-Magnoac (Hautes Pyrénées, France). Rev Paléobiol 26(2):403–411
Éhik J (1930) Prodinotherium hungaricum n.g., n.sp. Institutum Regni Hungariae Geologicum 6:1–21
Fortelius M (2013) New and Old Worlds database of fossil mammals (NOW). University of Helsinki. Available at: http://www.helsinki.fi/science/now/. Accessed 2 Sept 2013
Galanti V, Preatoni D, Martinoli A, Wauters LA, Tosi G (2006) Space and habitat use of the African elephant in the Tarangire–Manyara ecosystem, Tanzania: Implications for conservation. Mamm Biol 71(2):99–114. doi:10.1016/j.mambio.2005.10.001
Gasparik M (1993) Deinotheres (Proboscidea, Mammalia) of Hungary. Ann Hist-Nat Mus Natl Hung 85:3–17
Gasparik M (2001) Neogene proboscidean remains from Hungary; an overview. Fragm Pal Hung 19:61–77
Ginsburg L, Chevrier F (2001) Les Dinothères du bassin de la Loire et l’évolution du genre Deinotherium en France. Symbioses 5:9–24
Göhlich UB (1998) Elephantoidea (Proboscidea, Mammalia) aus dem Mittel- und Obermiozan der oberen Süsswassermolasse Süddeutschlands: Odontologie und Osteologie. Münchner Geowiss Abh A 36:1–245
Göhlich UB (1999) Order Proboscidea. In: Rössner G, Heissig K (eds) The Miocene land mammals of Europe. Verlag Dr. Friedrich Pfeil, München, pp 157–168
Gräf IE (1957) Die Prinzipien der Artbestimmung bei Dinotherium. Palaeontogr Abt A Palaeozool-Stratigr 108(5/6):131–185
Gross M, Böhme M, Prieto J (2011) Gratkorn: A benchmark locality for the continental Sarmatian s.str. of the Central Paratethys. Int J Earth Sci (Geol Rundsch) 100(8):1895–1913. doi:10.1007/s00531-010-0615-1
Gross M, Böhme M, Havlik P, Aiglstorfer M, (2014) The late Middle Miocene (Sarmatian s.str.) fossil site Gratkorn - the first decade of research, geology, stratigraphy and vertebrate fauna. In: Böhme M, Gross M, Prieto J (eds) The Sarmatian vertebrate locality Gratkorn, Styrian Basin. Palaeobio Palaeoenv 94(1). doi:10.1007/s12549-013-0149-1
Harris JM (1973) Prodeinotherium from Gebel Zelten, Libya. Bull Br Mus (Nat Hist) Geol 23:285–350
Harris JM (1975) Evolution of feeding mechanisms in the family Deinotheriidae (Mammalia: Proboscidea). Zool J Linn Soc 56:331–362
Harris JM (1996) Isotopic changes in the diet of African Proboscideans. J Vertebr Paleontol 16(40A)
Havlik P, Aiglstorfer M, Beckman A, Gross M, Böhme M (2014) Taphonomical and ichnological considerations on the late Middle Miocene Gratkorn locality (Styria, Austria) with focus on large mammal taphonomy. In: Böhme M, Gross M, Prieto J (eds) The Sarmatian vertebrate locality Gratkorn, Styrian Basin. Palaeobio Palaeoenv 94(1). doi 10.1007/s12549-013-0142-8
Haynes G (1988) Longitudinal studies of african elephant death and bone deposits. J Archaeol Sci 15(2):131–157. doi:10.1016/0305-4403(88)90003-9
Hilber V (1914) Steirische Dinotherien. Mitt Naturwiss Ver Steiermark 51:111–132
Huttunen KJ (2000) Deinotheriidae (Probosidea, Mammalia) of the Miocene of Lower Austria, Burgenland and Czech Republic: Systematics, Odontology and Osteology. PhD thesis. Universität Wien, Vienna
Huttunen KJ (2002a) Systematics and Taxonomy of the European Deinotheriidae (Proboscidea, Mammalia). Ann Naturhist Mus Wien A 103A:237–250
Huttunen KJ (2002b) Deinotheriidae (Proboscidea, Mammalia) dental remains from the Miocene of Lower Austria and Burgenland. Ann Naturhist Mus Wien A 103A:251–285
Huttunen KJ (2004) On a Prodeinotherium bavaricum (Proboscidea, Mammalia) skeleton from Franzensbad, Czech Republic. Ann Naturhist Mus Wien A 105A:333–361
Huttunen KJ, Göhlich UB (2002) A partial skeleton of Prodeinotherium bavaricum (Proboscidea, Mammalia) from the Middle Miocene of Unterzolling (Upper Freshwater Molasse, Germany). Geobios 35:489–514
Joger U (2010) The way of life and ecology of modern elephants: Are conclusions about straight-tusked elephants and their habitat possible? [in German]. In: Meller H (ed) Elefantenreich. Eine Fossilwelt in Europa. Landesamt für Denkmalpflege und Archäologie Sachsen-Anhalt. Landesmuseum für Vorgeschichte, Halle, pp 314–321
Kaup JJ (1829) Neues Säugethier, Deinotherium: Deinotherium giganteum. Isis 22(4):401–404
Kaup JJ (1832) Description d’ossements fossiles de Mammifères inconnus jusquá present, qui se trouvent au Muséum grand ducal de Darmstadt. C. Stahl, Darmstadt
Kovachev D, Nikolov I (2006) Deinotherium thraceiensis sp. nov. from the Miocene near Ezerovo, Plovdiv District. Geol Balc 35(3–4):5–40
Lister AM (2013) The role of behaviour in adaptive morphological evolution of African proboscideans. Nature 500:331–334. doi:10.1038/nature12275
Markov GN (2008a) Fossil proboscideans (Mammalia) from the vicinities of Varna: a rare indication of middle Miocene vertebrate fauna in Bulgaria. Hist Nat Bulg 19:137–152
Markov GN (2008b) The Turolian proboscideans (Mammalia) of Europe: preliminary observations. Hist Nat Bulg 19:153–178
McComb K, Baker L, Moss C (2006) African elephants show high levels of interest in the skulls and ivory of their own species. Biol Lett 2(1):26–28. doi:10.1098/rsbl.2005.0400
Merceron G, Costeur L, Maridet O, Ramdarshan A, Göhlich UB (2012) Multi-proxy approach detects heterogeneous habitats for primates during the Miocene climatic optimum in Central Europe. J Hum Evol 63(1):150–161. doi:10.1016/j.jhevol.2012.04.006
Mottl M (1969) Die Säugetierfunde von St. Oswald bei Gratwein, westlich von Graz in der Steiermark. Festschrift des Landesmuseums Joanneum Graz:299-320
Mottl M (1970) Die jungtertiären Säugetierfaunen der Steiermark, Südost-Österreichs. Mitt Mus Bergbau Geol Tech 31:3–92
Musil R (1997) A Dinotherium Skeleton from Česká Třebová. Acta Mus Moraviae Sci Geol 82:105–122
Peters KF (1871) Über Reste von Dinotherium aus der obersten Miozänstufe der südlichen Steiermark. Mitt Naturwiss Ver Steiermark 2:367–399
Pickford M, Pourabrishami Z (2013) Deciphering Dinotheriensande deinotheriid diversity. Palaeobio Palaeoenv 93(2):121–150. doi:10.1007/s12549-013-0115-y
Poole JH (1996) The African elephant. In: Kangwana K (ed) Studying elephants. African Wildlife Foundation Technical Handbook Series vol 7. African Wildlife Foundation, Nairobi, pp 1–8
Stefanescu G (1894) Dinotherium gigantissimum. Annu Mus Geologia Paleontol 1894:126–199
Svistun VI (1974) Dinotheriums of Ukraine [in Russian]. Naukovka dumka, Kiev
Tarabukin BA (1968) Excavation of a deinothere skeleton in the Rezesh-Area of the Moldavian Republic [in Russian]. Proc Acad Sci Mold SSR, Biol Chem Sci 3:37–42
Tassy P (1996) Dental homologies and nomenclature in the Proboscidea. In: Shoshani J, Tassy P (eds) The Proboscidea—Evolution and palaeoecology of elephants and their relatives. Oxford University Press, Oxford, pp 21–25
Thenius E (1948) Über die Entwicklung des Hornzapfens von Miotragocerus. Oesterr Akad Wiss Math-Natwiss Kl Sitzungsber 157:203–221
Tobien H (1988) Contributions a l’étude du gisement miocène supérieur de Montredon (Herault). Les grands Mammifères. 7—Les proboscidiens Deinotheriidae. Palaeovertebrata, Mémoire Extraordinaire 1988:135-175
Vendl M (1930) Die Geologie der Umgebung von Sopron II. Erdeszeti Kiserletek 32
Vergiev S, Markov GN (2010) A mandible of Deinotherium (Mammalia: Proboscidea) from Aksakovo near Varna, Northeast Bulgaria. Palaeodiversity 3:241–247
Zázvorka V (1940) Deinotherium levius Jourdan a jeho stratigrafický význam (in Czech and English). Acta Mus Nat Pragae IIB 7:191–214
Acknowledgements
The authors are indebted to P. Havlik (GPIT) for fruitful discussions and correction of the manuscript. G. Rössner (SNSB-BSPG), G. Scharfe (IGM), R. Ziegler (SMNS), H. Lutz (NHMM), T. Engel (NHMM), H. Stapf (SSN), N. Spassov (NMNHS), G. Markov (NMNHS), L. Hristova (NMNHS), M. Ivanov (PMSU) and P. Havlik (GPIT) are thanked for the help and access to comparison material. D. Vasilyan (University Tübingen) is thanked for the help with the Russian literature. W. Gerber (University Tübingen) and A. Schumacher (NHMW) are thanked for taking pictures. N. Winkler (UMJGP) is thanked for preparation of the material. Many thanks go to the reviewers, G. Markov (NMNHS) and M. Pickford (MNHN), who greatly supported the improvement of the manuscript with their careful comments and correction of the English. For improvement of the English we want to thank as well A. Beckmann (University Tübingen). And last but not least the authors want to thank the students and volunteers from Graz, Munich and Tübingen for the good work in the excavations from 2005 to 2013.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article is a contribution to the special issue “The Sarmatian vertebrate locality Gratkorn, Styrian Basin.”
Rights and permissions
About this article
Cite this article
Aiglstorfer, M., Göhlich, U.B., Böhme, M. et al. A partial skeleton of Deinotherium (Proboscidea, Mammalia) from the late Middle Miocene Gratkorn locality (Austria). Palaeobio Palaeoenv 94, 49–70 (2014). https://doi.org/10.1007/s12549-013-0140-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-013-0140-x