
Abstract
Dinosaur tracks occur at three vertebrate tracksites in north-western Germany, in the acanthicum/mutabilis ammonoid biozone of the basal Upper Kimmeridgian (Upper Jurassic, KIM 3-4 cycle, 152.70-152.10 Ma). The trackbeds are mud-cracked, siliciclastic, tidal sand flat biolaminates, overlain by paleosol beds. Channels contain rare fossils of sauropod, ornithopod and pterosaur bones as well as shark and plant remains. Large sauropod tracks of the Elephantopoides type, which have been found in an intertidal megatracksite environment to the north of the Rhenish Massif, are reviewed herein, together with a camptosaurid track type (?Iguanodontipus), the theropod track Megalosauropus and possible dryosaurid Grallator tracks. These large dinosaur tracks have been reviewed and compared to all other known European localities. At the Barkhausen tracksite with its important ichnoholotypes, trackways of a possible sauropod herd consisting of ten small to medium-sized individuals and one large individual have been exposed, revealing different speeds of travel as well as important social behaviour in these large herbivorous dinosaurs. Two theropods also left their imprints on the same track horizon, one travelling towards the south in a direction contrary to the movement of the herd, and the second travelling towards the north-west, cutting across the other trackways. Five different types of Upper Jurassic dinosaur tracks have now been recorded from coastal environments scattered around Europe, with the best footprint records forming extensive megatracksites in biolaminates between Jurassic islands in central Europe. These intertidal flats formed periodic bridges between the islands, allowing dinosaur interchanges and migrations between America and Eurasia, which may help to explain the much broader palaeobiogeographic distributions of dinosaur species during the Late Jurassic.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In 1921, vertebrate tracks were discovered by Prof. Dr. W. Klüpfel in north-western Germany at the Barkhausen tracksite in the Wiehengebirge mountain range (cf. Ballerstedt 1921c; Bödige 1921; Malz 1971). The tracks were described, with incomplete mapping of the trackways, by Kaever and de Lapparent (1974) and Friese and Klassen (1979), with their stratigraphic age given as “Lower Kimmeridgian” or “Kimmeridgian III” by Lockley and Meyer (2000). This paper reviews these descriptions and includes new trackway maps and new dating data (cf. Diedrich 2009a) as well as records of new track discoveries. The results for these Megalosauropus and Elephantopoides ichnogenera trackways in Germany are important because they are the oldest described sauropod and theropod tracks from the European Upper Jurassic and will be of significance for a future global revision of all large Jurassic dinosaur footprints.
Since the discovery of new dinosaur tracks in carbonates of the Upper Kimmeridgian (Upper Jurassic) in northern Switzerland, more tracksites and track beds have been found in the Jura Mountains (Lockley and Meyer 2000; Marty 2008). The discovery of tracks in carbonates and especially in mud-cracked biolaminates has led researchers to focus on the palaeogeography and bathymetry of these sites as well as on the megatracksite (Lockley and Pittman 1989) and ichnofacies concepts (Lockley et al. 1994a). In contrast to the megatracksite discoveries in the carbonate tidal flats of northern Switzerland (Marty 2008), we now have, for the first time, megatracksites in Upper Jurassic siliciclastic environments at new tracksite localities in north-western Germany. These discoveries are the result of studies on track horizons and track beds as well as detailed ammonoid biostratigraphy (Diedrich 2009a).
Previously undescribed discoveries of a few dinosaur bones from the Nettelstedt locality, such as a femur fragment, a claw from a giant sauropod (?Brachiosaurus sp.), a dryosaurid ulna (?Dryosaurus sp.), and pterosaur (?Comodactylus) bones, are the first recorded dinosaur/pterosaur remains from this region (coll. Breitkreutz, casts in the Dobergmuseum/Geologisches Museum Ostwestfalen-Lippe in Bünde). Another larger sauropod bone fragment has been found at Kalkriese (cast in the Naturkundemuseum am Schölerberg, Osnabrück).
Materials and methods
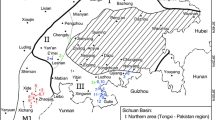
During the summer of 2002 new tracks were discovered in the Bruns quarry, Nettelstedt (52°17′34.40″N, 8°42′32.71″E) and in the Störmer quarry in Bergkirchen, Wallücke (Wiehengebirge, north-western Germany, 52°16′19.02″N, 8°46′1.02″E), some kilometres east of the known tracksite at Barkhausen (52°16′42.28″N, 8°24′49.73″E; Fig. 1). Tracks in sandstone blocks were discovered by chance in the Wallücke quarry during quarrying activities. A systematic programme was therefore started at several quarries in 2002, aimed at studing the stratigraphy and fossil content of these sites for a new exhibition at the Geologisches Museum Ostwestfalen-Lippe in Bünde, where all of the invertebrate fossils collected are stored, together with the footprints described herein. The invertebrates in particular were collected from fossil-rich layers with the aim of recovering rare ammonites and enabling both facies interpretation from the palaeocommunities and determination of their stratigraphic position within the Kimmeridgian section (see Diedrich 2009a). Also important was the identification of paleosols (cf. Bailly et al. 2000; Diedrich and Bailly 2003) and marine-to-terrestrial sedimentary sequences. Only the Barkhausen tracks are still visible, within a protected UNESCO–Geopark natural monument site, at a small quarry near Bad Essen. The Nettelstedt tracks are present in the wall of an inactive quarry. Casts of the original bones from this locality are displayed in the new permanent exhibition at the Geologisches Museum Ostwestfalen-Lippe in Bünde. The new single footprint records exposed by quarrying activities at the Barkhausen tracksite were put into stratigraphic context, and the complete quarry wall in which the trackways were found has been photographed and plans drawn up. Measurements were taken from the ten sauropod and two theropod trackways (Table 1); casts had already been made many years ago by the “Museum für Natur und Umwelt” in Osnabrück, where the casts of the holotypes are also stored. The measurements have allowed further analyses to be carried out on the dinosaur trackways (e.g. Thulborn 1990).

Geographical position of the tracksites at Bergkirchen, Nettelstedt and Barkhausen (Wiehengebrige, north-western Germany). Nettelstedt has yielded a few bones of different dinosaurs, such as sauropods, theropods and ornithopods, as well as of pterosaurs, crocodile and turtle remains and some plants. Only a single sauropod bone has been discovered in Kalkriese and Nettelstedt
Geology, palaeoenvironment and dating
As a result of tectonic uplift that occurred mainly in late Tertiary times, starting in the Lower Kimmeridgian (Upper Jurassic), the sediments in these tracksites have been deformed and dip towards the north-east at between 30° 60° in all outcrops. As a result, the trackways exposed at the Barkhausen natural monument tracksite are visible in the quarry wall, as are some at the Nettelstedt site.
With the new stratigraphic interpretation and international subdivision of the north-western German Kimmeridgian (Diedrich 2009a), track member I (Fig. 2) and overlying dinosaur bone-bearing paleosol (sensu Bailly et al. 2000) are dated into the KIM 2-3 cycle of the Lower Kimmeridgian (153.54-152.70 Ma after Hardenbol et al. 1998). The dinosaur track beds of the main track bed II at Barkhausen, Nettelstedt and Bergkirchen fall into the KIM 3-4 cycle (152.70-152.10 Ma after Hardenbol et al. 1998), in the acanthicum/mutabilis ammonoid biozone. The overlying eudoxus ammonoid biozone is marked in all outcrops by the “triple events” in the Stollenbank Member represented by the Eocallista/Thracia beds, the Nanogyra bed [in which Orthaspidoceras schilleri (Oppel) occurs at the Nettelstedt and Bergkirchen outcrops; cf. also Schweigert 1999] and the Sellithyris bed (Diedrich 2009a).

Stratigraphy and correlation of the Lower to Upper Kimmeridgian sections in the Barkhausen, Nettelstedt and Bergkirchen quarries (Wiehengebirge, north-western Germany). The described dinosaur tracks are from a megatracksite in the Upper Kimmeridgian that is marked in red. Only reptile scratch marks were found in the lower track bed of the Lower Kimmeridgian. Paleovertisol beds (brown) are not widely distributed in this region
The Lower to Upper Kimmeridgian sediments of this region are characterised by cyclic changes between marine carbonate and terrestrial siliciclastic deposits. Three of these sedimentary cycles (KIM 2-4) are exposed in the north-western German Kimmeridgian sections at Nettelstedt and Bergkirchen (Diedrich 2009a). The dominance of mud-cracked biolaminates in the terrestrial deposits, the thinning of terrestrial sequences towards the east, the absence of coal beds and the greater development of marine sequences combine to indicate a more marine setting for the Bergkirchen section to the east and a more terrestrial setting for the Nettelstedt section to the west. The Barkhausen section is the only section to include fluvial deposits, indicated by strongly cross-bedded haematite-rich sandstones, muscovite-rich claystones to siltstones, and clay-pebble beds.
The tracks were found in tidal flat sediments, especially in sand flats, where the terrestrial deposits are a result of the input of terrestrial siliciclastic sediments intercalating with the marine carbonate facies (Diedrich 2009a). The siliciclastic biolaminates from the desiccated track-bearing deposits have been formed by cyanobacterial and algal mats that covered tidal flat surfaces, in environments that are comparable to recent sand flat environments in the North Sea (cf. Krumbein et al. 1994; Kock 1998). Only early diagenesis by biofilms drying under high summer temperatures, combined with rapid sedimentation caused by storm events, could have preserved these tide-influenced, mud-cracked, track-bearing surfaces.
The Bergkirchen section at Wallücke exposes the Middle Callovian (Ornatenton Formation) through to the middle Upper Kimmeridgian; the tracks were found in the Sand-Tonkomplex Member (Fig. 2). The change from shallow marine to tidal flat deposits is marked by polygonally cracked biolaminates of 1- to 10-cm-thick sandstone beds which are thinner and comprised of siliciclastic biolaminate tidal flat and sand flat/sand bar facies types (Diedrich 2009a). The track of the Grallator sp. type (epirelief preservation) on a sandstone block came from a 50-cm-thick massive sandstone bed in the upper part of a thick sandstone series (cf. Fig. 2). The underside of this block is covered by desiccation cracks which are only found in a single layer of the Sand-Tonkomplex-Member, and hence the track bed can be accurately located at the top of the sandstone series, representing a second track horizon (Fig. 2). Possible sauropod tracks were also found here preserved as hyporeliefs on the cracked surfaces, and finally, a first “iguanodontid-like” footprint discovery was made at this site, occurring as a solitary epirelief imprint. In contrast to Nettelstedt and Barkhausen, another paleosol bed (in this case an initial paleosol sensu Bailly et al. 2000) is preserved at the top of the Bergkirchen sedimentary sequence (cf. Diedrich 2009a).
At the second track locality, the Nettelstedt section is a large outcrop in an old quarry that starts in the Middle Callovian (Ornatenton Formation) and extends to the middle Upper Kimmeridgian (Stollenbank Member, Fig. 2). Only at this location does the sequence contain dinosaur bones in the Lower Kimmeridgian (Unterbank Member) and possible dinosaur tracks in the Upper Kimmeridgian (Sand-Tonkomplex Member). The nearly 3-m-thick, massive sandstone of the Unterbank Member, at Nettelstedt, comprises a series of sandstone beds varying from 20 to 40 cm in thickness and is followed by the first terrestrial paleosol bed (= paleovertisol, Diedrich and Bailly 2003) of the Kimmeridgian in which vertebrate remains have accumulated in sandy channel deposits. A femur fragment and a claw phalanx from a huge sauropod, two different sized and shaped theropod teeth, two large sauropod teeth and some pterosaur bones are the first known non-marine reptile remains from the Kimmeridgian in this region. Osteoderms from the marine crocodile Machimosaurus and a nearly complete fin spine and teeth from the shark Hybodus indicate a mixing of coastal faunas, as does the presence of the marine turtles Eurysternum wagleri Meyer, 1839, Idiochelys fitzingeri Meyer, 1839 and Tropidemys langii Rütimeyer, 1873 (cf. Karl 1997). The presence of Hybodus together with marine turtles proves a near-shore location for the paleovertisol bed, while the presence of huge terrestrial sauropods and theropods indicates a close relationship between these dinosaurs and the tidal flats in which they left their trackways. The lower parts of these sandy channels are filled with pebbles, vertebrate remains, the remains of plant fragments and even complete tree trunks, and they therefore appear to be the products of storm events during which the paleosols in these coastal regions were subjected to rapid flooding (cf. Bailly et al. 2000). During these short-duration flood events, terrestrial dinosaur and marine vertebrate bones were winnowed out and became concentrated at the base of sandy channels, together with trees and other plant material. These channels were once some tens of metres in width but have now been completely destroyed by the Nettelstedt quarrying activities (Diedrich 2009a). In the Lower Kimmeridgian paleosol bed, lenses of plant remains with well-preserved leaves and silicified wood have been found in a “coal bed”. This Lower Kimmeridgian flora is dominated by cycadeans, bennetideans and conifers, described by Schultka (1991) (see the slab in Fig. 3). With these tidal flat deposits of the Unterbank Member a second terrestrial sequence starts. The sand flat is represented by a massive sandstone of about 5 m in thickness. Possible sauropod tracks were found as hyporeliefs in the quarry wall, in the centre of the sandstone bed of the 8-m-thick Sand-Tonkomplex Member, and these fall into the KIM 3-4 cycle of the acanthicum/mutabilis ammonoid biozone. Desiccation cracks and north–south trending drag marks (which are mutually exclusive) on the underside of the surfaces are typically found next to these dinosaur tracks. The sedimentary structures indicate a dissection of the tidal flat facies into narrow channels and ponds. The grooves represent negatives of plant remains washed offshore, leaving characteristic drag marks in the sand.

Plant remains from the Lower Kimmeridgian channel infill (see position in Fig. 2) at Nettelstedt (Dobergmuseum Bünde collection)
At Barkhausen, Europe's best known dinosaur track location, only the sequence from the Unterbank to the Stollenbank Members is exposed in the small quarry. At the base of the terrestrial sequence is a red-violet to brown sandstone paleosol bed with haematite-rich red claystones, which is a characteristic feature of this tracksite (Diedrich and Bailly 2003). This sequence is overlain by yellowish to greenish-greyish paleovertisols and finally by mud-cracked silty biolaminates in which tracks occur, forming the first track horizon (Horizon I in Fig. 2). Three, or possibly four, track horizons (Fig. 2, Barkhausen section) are known within the 13-m-high Sand-Tonkomplex Member. These track layers form a so-called “megatracksite” along the Wiehengebirgs coast, in which the layers can split into such megatracksites within different facies types, as demonstrated herein (Fig. 2). This is a “coastal megatracksite” type, and not a “terrestrial megatracksite”: the latter types are mostly found along lakes or rivers and are much more difficult to date accurately (cf. Lockley and Meyer 2000). In coastal megatracksites, the presence of underlying and overlying marine cephalopod-bearing sequences allows precise dating of the track horizons in northern Germany (Fig. 2). All of these track horizons contain sauropod tracks as epireliefs, but Horizon I also has theropod tracks. Only Horizon I contains well-preserved trackways, the other track beds, such as track Horizon II (cf. Fig. 2), appear to be trampled “dinoturbated” horizons. The lowermost of the terrestrial paleosols that are intercalated into the track horizons is a paleovertisol (Diedrich and Bailly 2003). A series of various sandstones, such as red sandstones or arkoses, overlies the trackbeds; ripple marks and mud cracks are present in these reddish terrestrial sandstone complexes as are fine plant remains within a muscovite mica-rich bed (cf. Fig. 2).
Systematic palaeontology
Two vertebrate track types have been described to date from the Kimmeridgian of north-western Germany. Two giant dinosaur track types, one sauropod and one theropod, were reported from the first Upper Jurassic tracksite discovered in Europe (e.g. Friese 1960; Kaever and de Lapparent 1974). A third small tridactyl track has subsequently been briefly mentioned (Diedrich 2004a; Diedrich and Strauß 2004) and is discussed here in detail. A previously unpublished fourth track type, which was discovered some years ago, can now be added to the list. Five different vertebrate tracks and traces can therefore now be described from the Kimmeridgian of north-western Germany, four of which are dinosaur tracks and the fifth of which is a scratch mark. The known ichnospecies from Barkhausen (cf. Kaever and de Lapparent 1974) have also been studied and are now redescribed and discussed below as valid ichnogenera and ichnospecies.
Ichnogenus Elephantopoides Kaever and Lapparent, 1974
Elephantopoides barkhausenensis Kaever and Lapparent, 1974, Figs. 4–9
1974 Elephantopoides barkhausenensis. Kaever and Lapparent: 524, figs. 1, 3–6.
Ichnoholotype: Tracks herein designated as Trackway 1 (a fast-moving individual) and Trackway 2 (a slow-moving individual), and the manus/pes set of Trackway 1 (6th manus/pes set of the trackway, see Figs. 7, 8b and 17–Trackway 1 and Tab. 1). Original in the quarry, cast in the Naturkundemuseum, Osnabrück, coll. No. Barkhausen, track 1.
Ichnoparatypes: Trackways 3–10 (questionable: Trackway 7, see Fig. 17).
Type locality: Barkhausen, north-western Germany.
Type horizon:Sand-Tonkomplex Member, acanthicum/mutabilis ammonoid biozone, KIM 3-4 cycle, middle Upper Kimmeridgian, Upper Jurassic.
Material: Nine (possibly ten) trackways and some single imprints from Barkhausen on mud-cracked biolaminated siltstones/sandstones are positives (still in place, Fig. 17); two possible pes and one manus negatives on a sandstone slab from Bergkirchen destroyed during quarrying activities (Fig. 4), and one questionable manus/pes set at the base of a sandstone bed from Nettelstedt (still in situ, Figs. 5 and 6).

Possible tracks of the type Elephantopoides isp. (negatives, destroyed) on an isolated track slab from track bed 2 at the Bergkirchen tracksite. The bed consists of 50-cm-thick sandstone exposing desiccation marks as hyporeliefs in between the possible tracks and was found in the acanthicum/mutabilis ammonite zone (KIM 3-4 cycles, Upper Kimmeridgian)

Possible manus/pes set of the track type Elephantopoides isp. (negatives) from the acanthicum/mutabilis ammonite zone (Upper Kimmeridgian) in the quarry wall at the Nettelstedt tracksite. a in downside view, b redrawing, c lateral view

Middle section of Trackway 10: Elephantopoides barkhausenensis Kaever and Lapparent, 1974 (cf. Fig. 9d)
Diagnosis: Quadruped mainly wide-gauge trackway with strong heteropody. The pes trace is oval to triangular in outline (30–50 cm in diameter), sometimes with preserved claw marks from the three largest digits (see Figs. 7 and 8b). The step measurements depend on the speed and size of the trackmaker and are here between 74 and 114 cm, with a stride range of about 110–164 mm and with an internal trackway width measured from inside the pes of between 15 and 30 cm. The manus trace (20–35 cm in diameter) is crescentic and mainly has a typical hoof-shaped sauropod-like impression.


The heteropod track Elephantopoides barkhausenensis Kaever and Lapparent, 1974 and its possible trackmaker. a Brachiosaurus (skeleton redrawn and modified from Czerkas and Czerkas 1991), b Brachiosaurus pedal anatomy (redrawn after Thulborn 1990) projected onto a manus pes set track (cf. Fig. 7). c Part of the Barkhausen Trackway 1, showing partial overprinting of the hind foot on the fore foot imprint on the left hand side, whereas on the right hand side the imprints are of characteristic outward rotated manus/pes sets (cf. Fig. 9e)
Discussion: E. barkhausensis occupies a special place in the history of sauropod track research in Europe because this was the first sauropod track to be named and described. The description of the ichnoholotype Elephantopoides barkhausenensis was lacking the differentiation of manus and pes imprints, as illustrated by Friese (1960) and had confusing and contradictory interpretations of the sense of movement of the trackmaker (cf. Friese 1960; Kaever and de Lapparent 1974). Kaever and de Lapparent (1974) did not designate a single trackway or manus/pes set as the holotype. In Lockley and Meyer (2000), the opposite direction of movement was corrected, as they distinguished the manus/pes footprints by their heteropody. Herein, Trackway 1 and the well-preserved manus/pes set of Fig. 7 (6th manus/pes set of the trackway, see Fig. 17, Trackway 1) is therefore designated as the holotype trackway. Manus and pes imprints are clearly represented and are distinctly different in their outlines. The characteristic outward rotation is present in the manus and pes imprints, although overstepping of the hind foot onto the forefoot print has sometimes resulted in a single, oval shaped “fused double print”. The manus/pes distance and stride are characteristics that should not be used in defining an ichnospecies as they vary with the speed or age of the trackmaker (see Fig. 9).

Trackways of Elephantopoides barkhausenensis Kaever and Lapparent, 1974 from the Barkhausen site (cf. Figs. 6–8 and 17) showing the variability of the speed and individual size of the trackmakers. Six manus/pes sets were selected from each of the four important trackways (cf. Fig. 17). a This trackway has a small trackway width, a long stride and short manus pes distance, representing a fast-moving, small-to-medium-sized individual. b This is a very small individual, moving slowly, as indicated by the small trackway width and short stride and manus pes distances. c This medium-sized animal has a slightly larger trackway width and longer stride than Trackway 8. d Manus and pes, and the trackway width are the widest and were left by the largest animal from this tracksite
The description by Kaever and de Lapparent (1974) has been amended, especially with regard to reversing the direction of travel, made possible here by the clear differentiation of the hoof-shaped manus and oval pes prints. This new understanding of the pedal anatomy and heteropody has enabled a revision of the earlier interpretations (see Friese 1960)—and even of more recent preliminary interpretations (Diedrich 2004a)—of the travel direction of the possible sauropod maker of tracks 1–10 as being towards the north (see Fig. 17, and also Lockley and Meyer 2000), rather than towards the south, as described by Kaever and de Lapparent (1974).
The ichnogenera Rotundichnus from the Lower Cretaceous Berriassian of Münchehagen in north-western Germany (cf. Fischer 1998) or the Jurassic–Cretaceous Brontopodus, Parabrontopodus and Brevirparopus created by Lockley et al. (1994b, c) from North American or Portuguese sauropod trackways, seem to be, at least in some cases, nothing more than tracks of the Elephanotopoides ichnogenus track-type. Similar heteropod-shaped and wide-gauged trackways also described from the Upper Cretaceous (Cenomanian) of Croatia (Adriatic Platform: Dalla Vecchia 2008) have recently been assigned to Elephantopoides (Diedrich 2010a). Rotundichnus trackways from Münchehagen and Bückeburg have similar heteropod shapes (oval pes and u-shaped smaller manus imprints), although in several cases the identification and comparison of shapes is difficult due to overstepping of the hind limb imprint onto the fore limb imprint, or incompletely preserved and overwashed tracks (cf. Ballerstedt 1921a; Fischer 1998). The separation of the manus/pes and the gauge of the trackway are also variable at the Lower Cretaceous Münchehagen track site, where different trackway sizes also indicate possible herding, with individuals of different ages (cf. Fischer 1998). In trackways 1–4 of Münchehagen (cf. Fischer 1998), the trackways appear to lack the manus imprint, as is the case in several of the Barkhausen trackways, which may possibly be only a result of overstepping. Only in trackways 6 and 7 is the manus similarly close in front of the pes imprint (cf. Fischer 1998). Rotundichnus was created later as an ichnogenus without a critical analysis or a good comparison with the Barkhausen trackways. Only narrow-gauged tracks, such as those recently described from Croatia (Diedrich 2010a) and other sites in the Upper Cretaceous, are significantly different and must be differently named as Brontopodus, whereas the trackmakers are titanosaurids (e.g. Lockley 1991) which also leave heteropod tracks, but the manus and pes are mostly more widely spaced at any speed (e.g. Lockley 1991; Diedrich 2010a). The manus in Brontopodus is also slightly different in its hoof-shape, and the pes is generally larger and more rounded. Parabrontopodus is similar to Brontopodus (cf. Lockley et al. 1994b).
The gauge of the trackway is not always a reliable way to distinguish these sauropode trackways since, as is demonstrated herein, the gauge also depends on the trackmaker's size and speed (cf. Fig. 9). Similar variations (due to individual size and speed) are well documented at the Münchehagen trackways (cf. Fischer 1998). The Elephantopoides ichnogenus is the older and therefore the only valid ichnogenus in some cases for Upper Jurassic (up to basal Upper Cretaceous; cf. Diedrich 2010a) sauropod tracks, and is herein described anew. This description is of considerable importance because, even in recent studies (Marty 2008), these tracks from Barkhausen have not been compared to recent Upper Jurassic discoveries in Switzerland of similarly aged and completely identical manus/pes sets and narrow gauged trackways (cf. Marty 2008). The trackways of the young individuals at Barkhausen are approximately the same size as those from small individuals at the Cabo Espichel tracksite in Portugal (cf. Lockley et al. 1994c) or recently published tracks from Switzerland which were referred (possibly) to “Brontopodus” (cf. Marty 2008). Some of the Brontopodus tracks are even unclear because they often represent undertracks, which give false measurements. Most of these Swiss discoveries can herein also be referred to the same E. barhhausensis ichnospecies, which are of similar sizes, heteropod and within the range of the individual size- and speed-dependant trackways from the Barkhausen site. The redescription in this study of the Barkhausen tracks, together with new discoveries in north-western Germany, has served to confirm their importance and the validity of the older systematic names. This will necessitate a revision of the Upper Jurassic “Brontopodus” or “Brevirparopus” and other sauropod trackways of the world, which are, in several cases, most probably simply Elephantopoides trackways, as discussed here for the Lower Cretaceous Münchehagen locality or some basal Upper Cretaceous Croatian localities as well as for the Upper Jurassic Swiss sites. The Elephantopoides trackway type can be reported to range from the (?Early) Late Jurassic, to Early and even basal Late Cretaceous (Cenomanian–Turonian; Diedrich 2010a), and appears to be distributed globally.
That the manus–pes distance and stride depend on the age and speed of the trackmaker (see Fig. 9, Table 1) has also been demonstrated at the Swiss Combe Ronde tracksite (cf. Marty 2008). It is therefore not valid to create ichnospecies on the basis of differences in stride or trackway width, as has often been the done in the past. Even the track gauge is partly dependent on the speed, as can be demonstrated in the Barkhausen trackways (cf. Fig. 9, Table 1): the faster the animal runs, the narrower the track gauge (cf. Fig. 9a, c). Marty (2008) also described similar variations in track gauge in Kimmeridgian trackways from Switzerland.
Four main groupings of speed and age can be seen for the makers of Barkhausen Trackways 1–2 and 7–8 (cf. Fig. 9). For a comparison of the stride, gauge or trackway width, six manus-pes sets were selected from each trackway. The differences are so obvious from the illustrations that the presentation of precise metric data (see Table 1) is not considered to be necessary in this instance, but it could be used in another study for comparison with other trackways.
Two different speeds of young individuals are shown in Trackways 1 and 8 (cf. Fig. 9, Table 1), and a similar situation is also present from Switzerland (cf. Marty 2008). Trackway 1 represents a fast-moving adolescent individual, resulting in a long stride, the overlapping of the manus and pes imprints and a very narrow trackway width. Trackway 8 was also left by a younger individual, but one moving at a slower speed, as can be seen from the shorter stride, mostly non-overlapping manus/pes imprints and the greater trackway width. In Trackways 2 and 7, the speeds are similar, but the stride and manus–pes distances as well as the track size and trackway width increase with the body size. The producer of Trackway 2 is therefore of medium-size, while the producer of Trackway 7 is the largest individual in the group. Note that these interpretations will only be correct if the trackmakers are all from a single dinosaur species, which is believed to be the case in this instance.
Possible trackmaker: There are a number of arguments in favour of the interpretation of a single sauropod trackmaker. The general shapes of the manus/pes sets are all the same in all trackways, with nearly oval pes and hoof-like manus imprints, and the trackways are all similar narrow gauged ones (cf. Fig. 9), supporting this interpretation. The shape of tracks depends on the sediment moisture (cf. Fichter 1982; Brand 1996; Marty 2008), the trackmaker's age and its speed (e.g. Thulborn 1990) of travel (Fig. 9): all of these factors are variable and, hence, criteria such as stride, trackway width and manus–pes distance are not helpful in the systematic ichnotaxonomic definition of new track types. The three sites described at Purgatoire Valley (USA), Cabo Espichel (Portugal) and Combe de Ronde (Switzerland), together with the Barkhausen site in Germany, all appear to show more or less parallel trackways from mobile social groups of sauropods, of which the most probable trackmaker is believed to be Brachiosaurus (cf. Lockley et al. 1994c). Other possible large trackmakers have to be considered as well, such as Apatosaurus, Camarasaurus or Diplodocus. Giant herbivorous dinosaurs, such as Brachiosaurus, are also believed to be one of the most likely trackmakers for the Elephantopoides trackways (Fig. 8a), not only for the north German sites but also the Swiss sites (cf. Lockley et al. 1994b, c, d; Marty 2008). Recent discoveries of three bones, most probably from this dinosaur genus (two teeth, one claw phalanx, and one femur head, and one giant lumbar vertebra centrum; coll. H. Breitkreutz; casts in the Geologisches Museum OWL, Bünde, coll. No. 2002/1510-11, cf. Fig. 19) at the Nettelstedt locality support the new interpretations of the Barkhausen tracksite, where the trackways have documented individuals of different sizes and ages (Figs. 9, 17). Both the possible trackmakers and the tracks were found in close stratigraphic proximity and in similar coastal intertidal palaeogeographic situations, providing additional evidence to support the new trackmaker interpretations, although the dinosaur fauna did not consist solely of these giant sauropods (cf. Figs. 19, 20). Not all of the different sauropods have been found in the bone-preserving paleosol beds of north-western Germany; they are much better known from the nearly complete skeletons of the North American Morrison Formation sites or from England (cf. Galton 1980a, b; Dodson et al. 1980a, b; Hagood and West 1994; Holtz 2007). Other large, Upper Kimmeridgian sauropods, such as Diplodocus sp. (25 m in length), Camarasaurus sp. (18 m) or Apatosaurus sp. (26 m), with their different weights (e.g. Diplodocus 10 t, Camarasaurus 18 t, Apatosaurus 35-50 t, cf. Dodson et al. 1980a), are also possible trackmakers from the Jurassic period, but they have each different body proportions, which will require further detailed comparisons with the footprint or trackway record. Apatosaurids and camarasaurids have recently been suggested to be the most probable trackmakers of the Elephantopoides trackways in the intertidal megatracksites of Croatia (Diedrich 2010a). These track types have been found in northern America, Africa and Portugal, while Brachiosaurus and probably other sauropods, including camarosaurids and apatosaurids, also appear to have been recently identified in northern Germany from single tooth and postcranial bone remains (bones from Nettelstedt, Fig. 19, 2–4). All of these sauropods have well-developed claws on the three lateral digits (digits 3–5), which are responsible for the claw mark impressions, especially on the pes imprints (cf. Fig. 8b). A large Brachiosaurus claw from the Lower Kimmeridgian paleosol bed at Nettelstedt (see Fig. 19) and an incompletely prepared giant lumbar vertebra centrum (27 cm in diameter) which cannot be attributed to the smaller sauropods mentioned above and which were both compared directly with the giant Brachiosaurus skeleton from Tendanguru in Tanzania (Museum für Naturkunde, Humboldt-Universität, Berlin) support their existence here, with fully grown animals fitting into the trackway map from Barkhausen (see Trackway 7 left by a fully grown individual; cf. Figs. 10, 17).

The middle section of the ichnoholotype trackway of Megalosauripus teutonicus Kaever and Lapparent, 1974 overstepping some tracks of Elephantopoides barkhausenensis Kaever and Lapparent, 1974, which show some slipping of the trackmaker
Ichnogenus Megalosauropus Kaever and Lapparent, 1974
Megalosauropus teutonicus Kaever and Lapparent, 1974, Figs. 9–11
1974 Megalosauropus teutonicus. Kaever and Lapparent: 524, figs. 2 and 3.
1998 Megalosauripus teutonicus Kaever and Lapparent (1974) Amended. Lockley et al.: 326–327.
Ichnoholotype: Trackway herein designated as Trackway A and the pes of Figs. 10 and 11. 2nd pes of the trackway, see Fig. 17 and Tab. 1. Original in the quarry, cast in the Naturkundemuseum, Osnabrück coll. No. Barkhausen-Track 2.

Ichnoparatypes: Trackways A, B of Fig. 17.
Type locality: Barkhausen, north-western Germany.
Type horizon: Sand-Tonkomplex Member, acanthicum/mutabilis ammonoid biozone, KIM 3-4 cycle, middle Upper Kimmeridgian, Upper Jurassic.
Material: Two trackways (Trackways A and B) from Barkhausen on mud-cracked biolaminated siltstones/sandstones as positives (still in situ).
Diagnosis: Tridactyl bipedal of a large size with claw impressions but no digital pad preservation and digitigrade progression. The longest toe is in the middle, whereas the angles between digits II and III vary by about 25°, and between digits III and IV by about 20°. Digit IV is slightly longer then digit II. The long stride, in this case 140–160 cm, is not diagnostic as it depends on the individual’s size and speed. The central digit curves slightly outwards.
Discussion: Tridactyl theropod tracks have been found at many localities around the world in different stages of the Jurassic and Cretaceous; they were discussed anew by Lockley et al. (1994d). This ichnogenus is present in the Kimmeridgian of northern Germany, northern Switzerland, southern England and western Portugal (e.g. Lockley 1991; Lockley and Meyer 2000). Different Megalosauropus species names, described in the past, might be a result of different preservation types (cf. Beckles 1852, 1854; Haubold 1984; Delair and Sarjeant 1985; Thulborn 1990; Delair 1991; Lockley et al. 1994d; Dalla Vecchia 1999; Lockley and Meyer 2000). The suggestion that the confusion resulting from the ichnogenus Megalosauropus, thought to have been produced by the dinosaur genus Megalosaurus, would be overcome by changing the genus name to Megalosauripus (cf. Lockley et al. 1994d) does not comply with the International Code of Zoological Nomenclature, but the latter is nevertheless still being used (cf. Marty 2008). Kaever and de Lapparent (1974) were first to describe the ichnogenus “Megalosauropus” on the tridactyl tracks and trackways of Barkhausen; only the best tracks from this locality can therefore be chosen as holotypes, and ichno-paratypes from other localities cannot be declared to be “new holotypes” without violating of the International Code of Zoological Nomenclature. The Barkhausen tracks are still present and preserved without damage and can therefore not easily be declared as nomen nudum, since they are reasonably well preserved and there is even a trackway as the ichnoholotype. Furthermore, their preservation is not worse than most tridactyl tracks, which lack digital details; even the angle of the digits can be measured and compared. The original description and valid nomenclature should therefore be adhered to. There must certainly be many different “ichnospecies” described as Megalosauropus that, in many cases, represent nothing more than variations in preservation type, or in the individual size or speed of the trackmakers, as has been demonstrated at a Swiss tracksite (cf. Marty 2008). The same Megalosauropus teutonicus tracks are also present at this site, although not identified as such (“Therangospodus” in Marty 2008). In contrast to, for example, large tridactyl theropod tracks from the Mediterranean (cf. Dalla Vecchia 1999) or German Lower Cretaceous at Bückeburg and Obernkirchen (cf. Struckmann 1880; Grabbe 1881; Ballerstedt 1921b; Diedrich 2004b) or England (e.g. Targart 1846) which show strong variability in track morphology or digit and digital pad presence depending on the moisture content of the sandy substrate (cf. Diedrich 2004b), other dinosaur tracks in the Kimmeridgian, such as theropod trackways in Switzerland, show a consistent lack of detail (cf. Marty 2008) that, together with the limited number of known trackways, prevents them from being studied in any detail. The Megalosauropus trackways of Barkhausen are represented in Trackway A by deeply impressed tracks in sand flat biolaminate deposits. Neither this trackway nor Trackway B reveal digital pads or clear claw marks, as a result of the relatively soft and moist sandy/silty substrate. In comparison, the recently discovered tracks from the Lower Cretaceous of Obernkirchen, in north-western Germany (Diedrich 2004b), with their shallow digital pad and claw mark preservation, fit well into the outlines of the Upper Kimmeridgian tracks of Barkhausen, but a valid comparison of the two (including two different theropod dinosaurs) is not possible as the pedal pad details are missing from Jurassic footprints.
Possible trackmaker: The trackmaker for the Megalosauropus trackways was a large carnosaurian (e.g. Lockley et al. 1994d). At the Nettelstedt tracksite, three large theropod tooth fragments were found in the Lower Kimmeridgian paleosol bed (Fig. 19, 5). In the Upper Kimmeridgian, Allosaurus sp. or Megalosaurus sp. might be the most likely trackmakers (Lockley et al. 1994d; Diedrich 2004b, Fig. 11a), which are also possibly represented in the dinosaur fauna of Nettelstedt, where one large, strongly recurved tooth (Fig. 19, 5) may belong to a large carnosaur (?Megalosaurus), and a second smaller, and straighter tooth with well-preserved serrulation on both margins may belong to another carnosaurian (?Allosaurus) (cf. Fig. 19, 6).
Ichnogenus ?Iguanodontipus Sarjeant, Delair and Lockley, 1998
?Iguanodontipus sp., Figs. 12 and 13

?Iguanodontipus isp. (positive; Bergbaumuseum, Kleinenbremen collection) on an isolated sandstone block from the acanthicum/mutabilis ammonite zone (KIM 3-4 cycles, Upper Kimmeridgian) at the Bergkirchen tracksite

The track ?Iguanodontipus isp. and its possible trackmaker. a Camptosaurus skeleton reconstruction (redrawn after Carpenter and Wilson 2008 and modified for quadrupedal locomotion), b the pedal skeleton projected into the footprint from Bergkirchen, c idealised reconstructed trackway based on Iguanodontipus trackways (cf. Diedrich 2004b)
Material: A single imprint from Bergkirchen (Original in the Bergbaumuseum Kleinenbremen, No. coll. No. Bergkirchen-1).
Horizon: Sand-Tonkomplex Member, acanthicum/mutabilis ammonoid biozone, KIM 3-4 cycle, middle Upper Kimmeridgian, Upper Jurassic.
Diagnosis: The tridactyl and large pes impressions show hoof marks and therefore rounded toe tips in contrast to theropod tridactyl trackways, with the middle digit being the longest. Digits II and III are wider than digit IV, which seems to have had no hoof-like ending and is therefore more pointed. The heel impression is not convex, as in tridactyl theropod footprints, but on the contrary is U-shaped. The angle between digits II and III is about 15° and between digits III and IV is less at about 12°, but can only be determined for a single track.
Discussion: Sarjeant et al. (1998) have discussed the “Iguanodon” footprints and created the ichnogenus Iguanodontipus with the ichnospecies Iguanodontipus burreyi Sarjeant, Delair and Lockley, 1998 on a trackway of a specimen with a “bipedal” motion, with pes imprints but no manus imprints. This is problematic because one of the more complete trackways of iguanodontids with quadrupedal motion should instead have been designated as an ichnoholotype, such as the trackways described in Spain (cf. Moratalla et al. 1994a, b) or the newly discovered imprints from Obernkirchen (Diedrich 2004b). Following the descriptions and revisions of the Lower Cretaceous (Berriassian) dinosaur footprints of Iguanodontipus sp. (cf. Diedrich 2004b) from Obernkirchen and other north-western German sites, the single imprint from the Upper Jurassic (Kimmeridgian) presented herein can be seen to have typical characteristics of iguanodontid footprints, such as the blunt-ended, oval-shaped, distal toe pads. In the German Jurassic specimen only digits II and III are present. Digit IV has no hoof, is more slender and also shorter (see skeleton anatomy in Fig. 13), and hence the footprints differ from typical Iguanodon footprints as a result of this more slender and non-hoofed digit IV. In general, however, the specimen has a tridactyl outline similar to iguanodontid footprints (cf. also Bachhofen-Echt 1926; Ballerstedt 1921a, b, c; Thulborn 1990; Moratalla et al. 1994a, b; Sarjeant 1974; Sarjeant et al. 1998; Diedrich 2004a). This footprint is therefore only attributed to this ichnogenus in a preliminary way and would at least represent a different ichnospecies from the Lower Cretaceous iguanodontid footprints. The problem with defining a new genus or species is the worldwide scarcity of these footprints; a similar unnamed footprint has been described from the Kimmeridgian, and a solitary trackway has been described from the Upper Kimmeridgian Swiss site at Combe Ronde (cf. Marty 2008), which seems to be similar to the footprint described herein from northern Germany.
Possible trackmaker: Following the reconstruction of the pedal anatomy, it appears that the trackmakers of these large footprints must have been dinosaurs of the ornithopod hadrosaurid/camptosaurid lineage and that they have possibly been left by the large camptosaurid Camptosaurus (C. dispar, C. amplus, C. depressus) known from North America and a few European sites (C. pretswichii, near Oxford; up to 8 m long and 800 kg in weight: Gilmore 1909; Foster 2007). All of these possibly represent the same species, which could be the most likely trackmaker, having an appropriate pedal pes skeleton anatomy and body size (Fig. 13), thus providing evidence of a faunal interchange (Galton 1980a, b; Holtz 2007). These camptosaurids had two larger middle toes and two larger hoof claws on digits II and III, while the lateral digits were shortened (Fig. 13b), which is also characteristic of the iguanodontids. As is known for Iguanodon, those dinosaurs were not primarily bipedal, as illustrated, for example, by Carpenter and Wilson (2008). New skeletal reconstructions with a quadrupedal locomotion are presented herein (Fig. 13a), in agreement with other similar quadrupedal reconstructions of Iguanodon, with its body shape similar to Camptosaurus (cf. Diedrich 2004b for the lower Cretaceous Iguanodon). The possible ?Camptosaurus tooth from the Lower Kimmeridgian paleosol bed at Nettelstedt (see Fig. 19, 1) supports this suggestion.
Ichnogenus Grallator Hitchcock, 1858
Grallator isp., Figs. 14–15
Material: A track slab with one well-preserved pes imprint (Fig. 14) and another poorly preserved footprint as positives from a partial trackway in the Sand-Tonkomplex Member, acanthicum/mutabilis zone, KIM 3-4 cycle, Upper Kimmeridgian, Upper Jurassic, of the quarry at Störmer at Bergkirchen (Wallücke), on desiccated sandstones (original in the collection of the Dobergmuseum/Geologisches Museum OWL in Bünde; Bergkirchen-2).

The track Grallator isp. (positive; Dobergmuseum, Bünde collection) on an isolated sandstone block. Two imprints from one trackway (of which the better preserved one is illustrated here) were found in the acanthicum/mutabilis ammonite zone (KIM 3-4 cycles, Upper Kimmeridgian) at the Bergkirchen tracksite
Diagnosis: Footprints of about 15 cm in length, medium-sized, bipedal tridactyl and digitigrad imprints, often with claw impressions although details can be absent depending on the preservation type. The phalanx formula resulting from the interphalangeal articulation impressions is: 3 (digit IV), 4 (digit III), 5 (digit II) (Fig. 15b). Digit II is the longest, and digit IV the shortest. The angle between digits II and III is about 22°, and between digits III and IV about 15°.

The track Grallator isp. and a possible trackmaker. a Dryosaurus (redrawn after an original skeleton from the Upper Kimmeridgian of Tendanguru in Tanzania, Africa, exhibited in the Museum für Naturkunde der Humboldt-Universität, Berlin), b pes anatomy (from the same skeleton) projected onto the Bergkirchen track, c idealised trackway of Grallator isp. from the two Bergkirchen imprints
Discussion: In these two pes impressions the pes–pes distance measures 58 cm and falls within the range for this ichnogenus. A stride cannot be given here, with only two footprints. The comparison with the tracks of the Grallator ichnotype (see discussion in Rainforth 2002) from tracksites in Portugal (Santos et al. 2000a, b, c; Unnamed 2002), or northern America (Lockley 1991) shows similarities to other tracks, depending on the substrate. The small tridactyl footprints in Switzerland always lack one digital pad on all digits IV (cf. Marty 2008), and are therefore different; they also have a longer middle digit III.
Possible trackmaker: Grallator isp. tracks appear to have been produced by 2-to 3-m-long medium-sized bipedal dinosaurs. A lightweight bone construction similar to that of pterosaurs is characteristic of the Kimmeridgian ornithopod Dryosaurus sp. (Fig. 15a), which has a maximum body weight of 100 kg (cf. Dodson et al. 1980a) and a worldwide distribution (cf. Galton 1980a, b). Examples of other tridactyl dinosaurs from the Kimmeridgian that have left their footprints are medium-sized coelurosaurids, such as Ornitholestes (cf. Galton 1980a,b; Carpenter et al. 2005). The footprints from the Swiss Combe Ronde site (cf. Marty 2008) are possibly from this trackmaker, although they are a little different from the tridactyl tracks of northern Germany and have only three digital pads on digit IV as opposed to four on the well-preserved tracks from Obernkirchen. A humerus fragment was also found in the Lower Kimmeridgian paleosol bed at Nettelstedt which appears to belong to Dryosaurus (see Fig. 19).
Scratch marks (Fig. 16)

Scratch marks (negatives) from the Lower Kimmeridgian at the Bergkirchen tracksite
Material: One track slab (Fig. 16) from the Grenzsandstein Member, KIM 2-3, Lower Kimmeridgian, Upper Jurassic of Bergkirchen, in mud-cracked biolaminates with scratch marks consisting of three parallel sinuous scratches as negatives.
Diagnosis: These scratch marks have not been determined ichnotaxonomically. Scratch marks found here consist of two or three parallel, sinuous or convex grooves of different lengths (5–15 cm), sometimes interrupted, and curving convex to the midline.
Discussion: Scratch marks of this size have been described from various strata (e.g. Thulborn 1990; Diedrich 2005), and similar ?crocodilian scratches have been identified in the Kimmeridgian Morrison Formation of Wyoming (cf. Foster and Lockley 1997). The producer could have been, for example, a lacertian or a crocodilian, but other producers are also possible. Even limulid crustaceans have been demonstrated to create similar two to four parallel scratches with their pusher appendices in intertidal beach zones (cf. Diedrich 2009c, 2010b).
Ichnotaxonomic problems and intertidal site comparisons
The recently described Swiss tracksite at Combe Ronde, which has not been compared to Barkhausen, revealed similar sauropod trackways, possibly also from Elephantopoides (= Brontopodus after Marty 2008), showing several individuals, also of different ages and travelling at different speeds, in various different track layers. There are also tracks of ?Iguanodontipus (one trackway) that were not referred by Marty (2008) to any ichnogenus or trackmaker. Theropod trackways are also preserved, but again the holotype Megalosauropodus was not used here for the track ichnotaxonomy. Finally, there are many ornithopod trackways (Carmelopodus) which were referred to smaller ornithopods. Dryosaurid trackways of Grallator type seem to either be absent from the Swiss site or could not be easily identified.
Megatracksite concept
In the Kimmeridgian, different ichnofacies types represent activity zones or even facies areas, in which specialised dinosaurs lived. These areas are similar to those that have recently been recorded for much earlier archosaurs and other reptiles in the Lower Triassic Buntsandstein terrestrial facies (e.g. Haubold 1984) or Middle Triassic coastal intertidal environments of Central Europe (Diedrich 2001, 2008, 2009b), or to the later dinosaur faunas in the Upper Jurassic intertidals of Europe (Meyer 1993; Dalla Vecchia 1994; Meyer et al. 1994; Meyer and Mezga 2000; Diedrich 2004a; Marty 2008) and the coastal swamps of the Lower Cretaceous of Central Europe (e.g. Struckmann 1880; Ballerstedt 1921a, b; Dietrich 1926; Lehmann 1978; Diedrich 2004b), such as the carbonate intertidal flats of the Lower Cretaceous Mediterranean Platform margin (Gianolla et al. 2000; Dalla Vecchia 2008) and Upper Cretaceous intertidal flats of the Mediterranean Carbonate Platform and Northern Africa (e.g. Dalla Vecchia 2008; Diedrich 2010a). At the Barkhausen tracksite, only two tracks, from Elephantopoides and Megalosauropus, made up a sauropod-theropod ichnocoenosis in tidal flat biolaminate deposits. Such track associations are also known from northern America and northern Switzerland (cf. Lockley 1991; Marty 2008). Ornithopod trackways are less common in the German tracksites and also in most of the Swiss tracksites, but they are abundant in Kimmeridgian tracksites found in the carbonate tidal flat deposits of Portugal (cf. Santos et al. 1998, 2000a, b, c) and at the Swiss Combe Ronde site (cf. Marty 2008).
The first descriptions of vertebrate tracks in the Upper Kimmeridgian sandstones at the Barkhausen locality did not mention exact biostratigraphic positions of the track beds and horizons due to the lack of ammonoids in the intercalating marine deposits (cf. Klassen 1968, 1984, 2000; Kaever and de Lapparent 1974). The description by Lockley and Meyer (2000) dated the Barkhausen tracks as being of Lower Kimmeridgian age. Their dating has recently been revised through new ammonite finds, the correlation of events and marker bed stratigraphy to be Upper Kimmeridgian (see Diedrich 2009a). The dating of the Sand-Tonkomplex Member track horizon places it into the acanthicum ammonite biozone (sub-Mediterranean subdivision) of the Upper Kimmeridgian or in the mutabilis-biozone (French subdivision: cf. Zeiss 1991; Schweigert 1999).
With the discovery of new dinosaur tracks and sites between the Barkhausen and Bergkirchen tracksites, a megatracksite has for the first time been identified in north-western Germany spanning a distance of approximately 20 km. Elephantopoides, Megalosauropus, Grallator and ?Iguanodontipus tracks have been found in the Sand-Tonkomplex Member of the basal Upper Kimmeridgian (Fig. 2). Correlations are therefore now possible with northern Switzerland, where newly described dinosaur trackways are of similar Upper Kimmeridgian age (cf. Marty 2008).
During the break-up of Pangea, Kimmeridgean megatracksites were distributed across the world in intramontane biotopes of northern America (Lockley et al. 1986; Lockley and Prince 1988; Lockley 1991) and central Asia (Lockley et al. 1996) and especially in the coastal zones of Germany (Diedrich 2004a), Switzerland (Marty 2008), Portugal (Santos et al. 1998, 2000a, b, c), England (Ensom 1989, 1994, 1995a, b) and Morocco (Dutuit 1980). Megatracksites of Upper Kimmeridgian age are also known from numerous locations in the Jura Mountains near Cerin, in south-eastern France (Lockley and Meyer 2000) and at additional sites, such as Lommiswil, Moutier or Combe Ronde in northern Switzerland (Meyer 1993; Marty 2008), as well as in the Morrison Formation of Kansas, South Dakota and Arizona in the USA (Lockley 1991). Other Kimmeridgian dinosaur tracksites have also been found near Arroyo Cerezo in Spain (Lockley 1991).
In contrast to the Swiss megatracksites (cf. Marty 2008) and also the French and Portuguese megatracksites, all of which are carbonate intertidal flat palaeoenvironments (Fig. 18), as well as the extensive Middle Triassic carbonate intertidal megatracksites of Central Europe (Diedrich 2009b), the north German locations are all found within siliciclastic tidal flats. The aforementioned five different dinosaur track ichnogenera have now been recognised at all European megatracksites, with a dominance of sauropod tracks and less common theropod and ornithopod or coelurosaurid trackways. The problem encountered in a detailed discussion about dinosaur assemblages and their footprints is the absence of a representative fossil record. Many more track types may be present, but these are yet to be discovered, and there may possibly also be faunal differences that are difficult to recognise from single footprint records, such as some of those from northern Germany. There are, however, clear similarities in footprints and ichnogenera, such as can be seen for the Elephantopoides and Megalosauropus trackways that are distributed over much of central Europe (Fig. 18). These similarities seems to be the result of specialised sauropods, theropods, ornithopods and other dinosaurs living either primarily, or most probably periodically, along coastal zones (or using them as seasonal migration highways), particularly in carbonate or siliciclastic tidal flat regions surrounding old massifs. In northern Switzerland, a carbonate platform seems to have formed a bridge between the ancient Rhenish and Central Massifs during a lowstand period (cf. Marty 2008). During such periods, extensive tidal flat regions would have allowed different dinosaurs to move between the ancient massifs. This fact is very important for the understanding of dinosaur and megatracksite distributions during the Kimmeridgean, at a time when the Upper Jurassic world was experiencing the breaking up of geotectonic plates. The Swiss megatracksites are different because they are not “coastal beach-zone tracksites” such as the ones in Portugal, England and Germany (Fig. 11). The megatracksites in North America are different again from those in Europe in that they are intramontane megatracksites (Lockley 1991).

New trackway interpretation of the Barkhausen tracksite (see Fig. 2 for legend of section). In track horizon I there are ten nearly parallel trackways of Elephantopoides with a travel direction towards the north–north-east. The trackway on the left (Trackway 1) is of a very fast moving sauropod. One trackway (Trackway 7) is of a large individual flanked on the right by a very small animal (Trackway 8), and another trackway (Trackway 3) is irregular with sliding impressions. Trackway 7 may belong to a different dinosaur species from the other sauropod trackways, or it may belong to the male or female head of the herd and be of the same species. Some solitary undertracks do not create trackways. The well-preserved first trackway of Megalosauropus (Trackway a) shows a contrary direction of travel, towards the south, the second (Trackway b) is a poorly preserved theropod trackway, with travel towards the north-west. On track horizon II a trampling bed shows “dinoturbation”, and it is difficult to get a clear impression of these trackways from a different time period. Arrows indicate the track ichnoholotypes manus/pes imprint set from Elephantopoides (Trackway 1) and pes imprint of Megalosauropus (Trackway a)

Geographical position and paleogeography of the megatracksites, and distribution of dinosaurs in north-west Germany, northern Switzerland, southern England, and western Portugal during the Kimmeridgian. Megatracksites from 1–2. northern Germany in siliciclastic tidal flat facies (this paper), 3. northern Switzerland in carbonate tidal flat facies, after Marty (2008), 4 southern England in carbonate tidal flat facies (cf. Ensom 1994, 1995a,b), western Portugal in carbonate tidal flat facies (Lockley et al. 1994c; Santos et al. 1998, 2000a–c)
Dinosaur ecology and herding theory
The dinosaur trackway map for the Barkhausen tracksite and its new interpretation provide an insight into the palaeoecology of sauropod behaviour. A hypothesis is presented herein that at least ten sauropods travelled, possibly at the same time, in more or less parallel (partly subparallel/diagonal) directions towards the north (Fig. 17) and that the trackmakers were all of the same sauropod species. As such the map provides new information about the social behaviour of these giants. In this herd, only one large individual (Trackway 7) was present, being close to the centre, flanked on the left by six and on the right by three juvenile and adolescent individuals. One of the smallest trackmakers (Trackway 8) stayed close to the sauropod of Trackway 7. This huge sauropod could have been either the main bull or the female head of this herd, accompanied by a very juvenile individual. There may also have been much larger animals travelling with such a herd, but that has not been documented in this case. Around the exposed trackways, additional trackways might be revealed, adjacent to the ten that have already been exposed. The impression we get from the exposed trackways provides a glimpse into a possible giant sauropod herd consisting of 10–12 animals, in which each individual traversed the same surface. It is not certain that these ten dinosaurs really moved isochronously through this tidal flat zone, but a similar footprint record of several different aged dinosaurs travelling along more or less “parallel” tracks has also been reported recently from Switzerland (Marty 2008). Supporting evidence for this interpretation of sauropod herds in Barkhausen derives from the similar orientation and preservation of the tracks, and the almost complete absence of crossing trackways (even Trackway 4). However, what we can definitely obtain from this scenario is a view into a partial or complete herd of sauropods of dissimilar ages travelling at different speeds (cf. Fig. 17), with one individual slipping (Figs. 10 and 17, Trackway 3) and a large male or female closely flanked by the youngest individual (Fig. 17, Trackways 7, 8).
There are differences in behaviour visible in some of the trackways. The outer left trackway (Trackway 1) was produced by a juvenile to adolescent animal, moving very quickly in comparison with the other trackmakers. The very irregular Trackway 3 may be the result of a young sauropod slipping on the biolaminate substrate, as can be seen from the two elongated imprints in the upper part of the trackway. After this slip, the trackway becomes irregular in subsequent imprints; recent systematic documentation has finally revealed the “normal” gait for this individual. Experiments with elephants in Osnabrück by Friese and Klassen (1979) have clearly shown similar slide marks, as have “human footprint experiments” carried out with on intertidal carbonate flats by Marty (2008). The sauropod of Trackway 3 may possibly have slid into the tracks of the animal from Trackway 4, which are no longer visible due to subsequent overtrampling. This phenomenon ties in with the interpretation of cyanobacterial and algae mats covering the tidal flat surfaces. Such microbial biofilms have been found in the tidal sand and mud flats of the coastal zones of the southern North Sea Basin (cf. Reineck 1972; Reineck and Singh 1973; Kock 1998). In Trackway 5, a limping dinosaur may have left tracks with alternating long and short steps. Such limping dinosaurs have also been described from sauropod and theropod trackways at other dinosaur tracksites (cf. Dantas et al. 1994).
Five nearly parallel sauropod trackways have been illustrated from the Morrison Formation (Salt Wash Member, Upper Kimmeridgian) of Purgatoire Valley in Colorado, USA (cf. Lockley et al. 1986; Lockley and Prince 1988; Lockley 1991). More than 100 individuals were investigated from 1,300 traces, of which 40% were sauropods and 60% tridactyl theropod and ornithopod dinosaurs (mostly theropods). One hundred and thirty individuals were identified in the Morrison Formation, from more than 20 locations, forming a megatracksite in a medium-sized intramontane lacustrine basin in North America. The large numbers of trackways at these tracksites do not allow an exact interpretation of the number of individuals walking in a herd at any one time, but they do show the same behaviour as sauropods in many of the other sites mentioned above, walking together with either a few or many other individuals at similar distances from each other, along parallel trends all heading in the same direction.
The tracksite most similar to the Barkhausen trackways is the Cabo Espichel locality, at Lagosteiros Bay in Portugal (cf. Lockley et al. 1994c). This site contains various different track-bearing levels with abundant sauropod, but also some theropod trackways. The one-track horizon bears the tracks from a herd of at least seven small sauropods that travelled towards the south-east. All trackmakers had similar foot sizes and wide-gauge trackways, indicating a similar age group of a single species, all travelling at a similar speed. These juvenile sauropods were accompanied by at least three larger, slow-moving individuals, also with wide-gauge trackways, that were moving in nearly the same direction (cf. Lockley et al. 1994c). This site was the first convincing example of sauropod herd behaviour to be reported from a European tracksite. This locality, and now also the Barkhausen tracksite in northern Germany, provides possible examples of herding amongst juvenile to adolescent sauropods. The Cabo Espichel trackway evidence for gregarious herding behaviour in sauropods (cf. Lockley et al. 1994c) is comparable to that from the Barkhausen tracksite.
At another tracksite, in the Kimmeridgian Morrison Formation of eastern Utah (USA), three sauropod trackways have been described. They were produced by individuals of approximately the same size walking towards the west, probably as a social group (Barnes and Lockley 1994).
Carnivorous theropod dinosaurs hunted sauropods, especially their juveniles, but the scenario of a carnivorous theropod hunting a herd of sauropods (Kaever and de Lapparent 1974) described for the Barkhausen locality does not agree at all with the trackway map (cf. Fig. 17). On the contrary, the theropod of Trackway a (Fig. 17) moved in the opposite direction to the sauropods, and the theropod Trackway b (Fig. 17) cuts across the sauropod tracks. A hunting scenario following a sauropod herd cannot therefore be inferred from these trackways, although the theropods were certainly the largest predators of the time in this region.
Dinosaur fauna and palaeoecology of the coasts of northern Germany
In northern Germany's Kimmeridgian forests, diversified flora extended to the coastal zones (cf. Schultka 1991) and provided potential food for snake-necked dinosaurs, such as Brachiosaurus, Camptosaurus or Dryosaurus walking along the coastlines, for example, of the Rhenish Massif at Barkhausen, Nettelstedt and Bergkirchen. On the beaches, after storm events, which also created channel systems in the “marsh” coastal environment with vertebrate and plant enrichments, such as at Nettelstedt, carcasses of turtle-crushing Machimosaurus, fish-hunting gavial-like Steneosaurus crocodiles (cf. Seebach 1864; Selenka 1867; Struckmann 1878; Diedrich 2009b), different turtles, such as Craspedochelys, Tropidemys, Plesiochelys and Hylaeochelys (cf. Karl 1997; Karl et al. 2007), different fish, such as Macromesodon, Lepidotes, Proscinetes, sharks, such as the large Asteracanthus or Hybodus (cf. Duffin and Thies 1997; Diedrich 2009b) and even pterosaurs were possible food sources for smaller or medium-sized carnivorous dinosaurs. In the shallow marine environments along the beach, the biodiversity of the marine invertebrate fauna was supplemented by freshwater influences (e.g. in the Barkhausen region) or fully-haline conditions, with coral patch reefs (cf. Bertling 1990; Diedrich 2009b).
Finally, the Wiehengebirge megatracksite is a key area for understanding the dinosaur fauna of Oker, in the Harz Mountains, where a juvenile brachiosaur specimen was misidentified by its bone structure as being a “dwarf island species”, for which a new dinosaur ichnogenus and species “Europasaurus holgeri” was established from bone histological studies by Sander et al. (2006). This was subsequently revised as a result of studies by Karl (2006) who showed that it was a juvenile brachiosaur. The large theropod bones, including jaws and teeth from the brachiosaur-bearing layers of the Upper Kimmeridgian of the Wiehengebirge and Oker, also do not support the “dwarf theory”. In contrast, the bone structures and trackways of Barkhausen and other sites of similar age show that a normal, full-sized dinosaur fauna existed during the Upper Kimmeridgian right along the northern German coasts, and not only at the Wiehengebirge Mountain chain (Fig. 20). Intertidal zones were present possibly with periodical sand flats between the coasts of the Wiehengebirge and the Harz Mountains, so that dinosaurs of normal size could easily have interchanged over these “intertidal highways” and possibly even to Bohemia (cf. Fig. 18). The juvenile brachiosaurs of Oker therefore seem more likely to be the trackmakers of the juvenile trackways at Barkhausen. The other dinosaur fauna of northern Germany is, however, still under investigation.
In the Wiehengebirge at least, a few remains of Brachiosaurus (lumbar vertebra, tooth, phalanx), Camptosaurus (?tooth) and Dryosaurus (femur) (Fig. 19), all of normal dinosaur size, fit well into the foot print record; hence the scenario depicted in Fig. 20 of a north German Kimmeridgian beach with a variety of normal-sized dinosaurs and plants in a coastal marsh landscape, with palaeovertisols dissected by storm channels, similar to the Oker dinosaur site.

Upper Kimmeridgian dinosaur remains from Nettelstedt in the Wiehengebirge close to the track site Barkhausen (see Fig. 1). 1 Large sauropod (?Camarosaurus) tooth cusp (Breitkreutz coll.; cast in the Geologisches Museum OWL, Bünde, coll. No. 2002/1514), a lingual, b lateral. 2 Large sauropod (?Brachiosaurus) tooth cusp (Breitkreutz coll.), lingual. 3 Large sauropod (?Brachiosaurus or Camarasaurus) claw digit I, phalanx II (Breitkreutz coll.; cast in the Geologisches Museum OWL, Bünde, coll. No. 2002/1510), a dorsal, b lateral inner view. 4 Large sauropod distal femur fragment (Breitkreutz coll.; cast in the Geologisches Museum OWL, Bünde, coll. No. 2002/1510), a cranial, b joint view. 5 Incomplete theropod (?Megalosaurus) tooth (Breitkreutz coll.), lateral. 6 Serrulated theropod (?Allosaurus) tooth (Breitkreutz coll.), lateral. 7 Medium-sized ornithopod dryosaurid (?Dryosaurus) radius (Breitkreutz coll.; cast in the Geologisches Museum OWL, Bünde, coll. No. 2002/1512), lateral outer view. 8 Distal metapodial joint of a large pterosaur (?Comodactylus) (Breitkreutz coll.; cast in the Geologisches Museum OWL, Bünde, coll. No. 2002/1513), a lateral outer view, b joint view

Dinosaurs such as Apatosaurus (upper left), Brachiosaurus (upper right), Camptosaurus (lower left), Dryosaurus (lower centre), and the large theropod Allosaurus/Megalosaurus (lower right) in the siliciclastic beach environment north of Wiehengebirge, northern Germany (illustration by G. “Rinaldino” Teichmann)
References
Bachhofen-Echt A (1926) Iguanodon-Fährten auf Brioni. Paläont Z 7:172–173
Bailly F, Felix-Henningsen P, Klassen H, Stephan S (2000) Synsedimentäre Paläoböden im Oberjura des westlichen Wiehengebirges. Osn Naturw Mitt 26:15–46
Ballerstedt M (1921a) Dinosaurierfährten im Wealdensandstein des Harrl bei Bückeburg und einer zur Zeit freiliegenden Spur eines „vierfüßigen“ plumpen Dinosauriers. Z Dt Geol Ges 72:231–233
Ballerstedt M (1921b) Zwei große, zweizehige Fährten hochbeiniger Bipeden aus dem Wealdensandstein bei Bückeburg. Z dt Geol Ges Mb 73 B:76–91
Ballerstedt M (1921c) Über Schreckensaurier und ihre Fußspuren. Kosmos 19:77–80
Barnes FA, Lockley MG (1994) Trackway evidence for social sauropods from the Morrison Formation, Eastern Utah (USA). Gaia 10:37–42
Beckles SH (1852) On the Ornithoidichnites of the Wealden. Q J Geol Soc London 8:396–397
Beckles SH (1854) On the Ornithoidichnites of the Wealden. Q J Geol Soc London 10:456–464
Bertling M (1990) Sedimentologische Kontrolle von Flachschelf-Faunen im Oberen Jura Nordwestdeutschlands. N Jb Geol Paläont Abh 181:211–224
Bödige N (1921) Saurierfährten bei Barkhausen. Osnabrück und seine Berge. Wegweiser durch den Teutoburger Wald und das Wiehengebirge 15:3
Brand LR (1996) Variations in Salamander trackways resulting from substrate differences. J Paleont 70(6):1004–1010
Carpenter K, Wilson Y (2008) A new species of Camptosaurus (Ornithopoda: Dinosauria) from the Morrison Formation (Upper Jurassic) of Dinosaur National Monument, Utah, and a biomechanical analysis of its forelimb. Ann Carnegie Mus 76:227–263
Carpenter K, Miles C, Ostrom J, Cloward K (2005) Redescription of the small maniraptoran theropods Ornitholestes and Coelurus from the Upper Jurassic Morrison Formation of Wyoming. In: Carpenter K (ed) The Carnivorous dinosaurs. Indiana University Press, Bloomington, pp 49–71
Czerkas SJ, Czerkas SA (1991) Dinosaurs—a global view. Mallard Press, New York
Dalla Vecchia FM (1994) Jurassic and Cretaceous sauropod evidence in the Mesozoic carbonate platforms of Southern Alps and Dinarids. Gaia 10:65–73
Dalla Vecchia FM (1999) A sauropod footprint in a limestone block from the Lower Cretaceous of northeastern Italy. Ichnos 6(4):269–275
Dalla Vecchia FM (2008) The impact of dinosaur palaeoichnology in palaeoenvironmental and palaeogeographic reconstructions: the case of the Periadriatic carbonate platforms. Oryctos 8:89–106
Dantas P, Santos VF, Lockley MG, Meyer CA (1994) Footprint evidence for limping dinosaurs from the Upper Jurassic of Portugal. Gaia 10:43–48
Delair JB (1991) A history of Dinosaur footprint discoveries in the British Wealden. In: Gilette DD, Lockley MG (eds) Dinosaur tracks and traces. Cambridge University Press, Cambridge, pp 19–25
Delair JB, Sarjeant WAS (1985) History and bibliography of the study of fossil vertebrate footprints from the British Isles: Supplement 1973-1983. Palaeogeogr Palaeoclimatol Palaeoecol 49:123–160
Diedrich C (2001) Megatracksites in Triassic carbonate tidal flats of the Germanic Basin in Central Europe and their consequences onto the palaeogeography and sequence stratigraphy. Schr Dt Geol Ges 14:41–42
Diedrich C (2004a) Dinosaurier im Wiehengebirge. Fossilien 2004(3):180–185
Diedrich C (2004b) New important iguanodontid and theropod trackways of the tracksite Obernkirchen in the Berriassian of NW Germany and the megatracksite concept of Central Europe. Ichnos 11(3–4):215–228
Diedrich C (2005) Actuopalaeontological trackway experiments with Iguana on intertidal flat carbonates of the Arabian Gulf - a comparison to fossil Rhynchosauroides tracks of Triassic carbonate tidal flat megatracksites in the European Germanic Basin. Senck marit 35(2):203–220
Diedrich C (2008) Millions of reptile tracks - Early to Middle Triassic carbonate tidal flat migration bridges of Central Europe. Palaeogeogr Palaeoclimat Palaeoec 259:410–423
Diedrich C (2009a) Stratigraphy, fauna, palaeoenvironment and palaeocology of the Stollenbank Member (Süntel Formation, mutabilis/eudoxus boundary, KIM 4, Upper Kimmeridgian) of north-western Germany. N Jb Geol Paläont 252(3):327–359
Diedrich C (2009b) Palaeogeographic evolution of the marine Middle Triassic marine Germanic Basin changements – with emphasis on the carbonate tidal flat and shallow marine habitats of reptiles in Central Pangaea. Glob Planet Change 65(2009):27–55
Diedrich C (2009c) Die Saurierspuren-Grabung im basalen Mittleren Muschelkalk (Anis, Mitteltrias) von Bernburg (Sachsen-Anhalt). Archäologie in Sachsen-Anhalt, Sonderband 2009:1–63
Diedrich C (2010a) Dinosaur megatracksites in carbonate intertidal flats and their possible producers in the Cenomanian/Turonian of the Northern Tethys—coastal migration zones between Afrika and Europe. Bull Tethys Geol Soc (in press)
Diedrich C (2010b) Eine Wattwanderung im Mittelgebirge. Silbersee-Hüggel-Exkursionsführer, Osnabrück
Diedrich C, Bailly F (2003) Stratigraphy and genesis of paleosols in marine and terrestrial environments in the Kimmeridgian (Upper Jurassic) of Northwest Germany. Terra Nostra 03(3):23
Diedrich C, Strauß M (2004) Eine neue Dinosaurierspur aus dem “Jurassic-Park von Norddeutschland”. GMit 13:27
Dietrich OW (1926) Über Fährten ornithopodider Saurier im Obernkirchner Sandstein. Z Dt Geol Ges 78:614–621
Dodson P, Behrensmeyer AK, Bakker RT, McIntosh JS (1980a) Tapphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison Formation. Paleobiology 6(2):208–232
Dodson P, Behrensmeyer AK, Bakker RT, McIntosh JS (1980b) Taphonomy of the Morrison Formation (Kimmeridgian-Portlandian) and Cloverly Formation (Aptian-Albian) of the western United States. Mem Soc Geol Fr NS 139:87–93
Duffin CJ, Thies D (1997) Hybodont shark teeth from the Kimmeridgian (Late Jurassic) of northwest Germany. Geol Palaeont 31:235–256
Dutuit JM (1980) Découverte d`une piste de dinosaur sauropode sur le site d`empreintes de Demnat (Haut-Atlas Marocain). Mem Soc Geol Fr NS 139:95–102
Ensom PC (1989) Sunnydown Farm sauropod footprint site. Proc Dorset Nat Hist Archaeol Soc 110:167–168
Ensom PC (1994) A new vertebrate trackway from the Intermarine Member, Purbeck Limestone Formation, Dorset. Proc Dorset Nat Hist Archaeol Soc 115:183–184
Ensom PC (1995a) Dinosaur footprints in the Purbeck Limestone Group (?Upper Jurassic–Lower Cretaceous) of southern England. Proc Dorset Nat Hist Archaeol Soc 116:77–104
Ensom PC (1995b) Dinosaur footprint records for the Purbeck Limestone Group, Dorset, since 1981. Proc Dorset Nat Hist Archaeol Soc 116:151–152
Fichter J (1982) Aktuopaläontologische Untersuchungen an den Fährten einheimischer Urodelen und Lacertilier. Teil I: Die Morphologie der Fährten in Abhängigkeit von der Sedimentbeschaffenheit. Mainz naturwiss Arch 20:91–129
Fischer R (1998) Die Saurierfährten im Naturdenkmal Münchehagen. Mitt geol Inst Univ Hannover 37:3–59
Foster J (2007) “Camptosaurus dispar”—Jurassic West, the Dinosaurs of the Morrison Formation and Their World. Indiana University Press, Bloomington
Foster JR, Lockley MG (1997) Probable crocodilian tracks and traces from the Morrison Formation (Upper Jurassic) of eastern Utah. Ichnos 5:121–129
Friese H (1960) Das Geheimnis der Saurierfährten von Barkhausen. Hann Allg Z, 23/24:7
Friese H, Klassen H (1979) Die Saurierfährten von Barkhausen im Wiehengebirge. Veröff Landkr Osnabrück 1:1–36
Galton PM (1980a) Dryosaurus and Camptosaurus, intercontinental genera of Upper Jurassic ornithopod dinosaurs. Mém Soc Geol Fr NS 139:103–108
Galton PM (1980b) Dryosaurus, a hypsilophodont dinosaur from the Upper Jurassic of North America and Africa. Postcranial skeleton. Paläont Z 55(3/4):271–312
Gianolla P, Morsilli MD, Dalla Vecchia FM, Bosellini A, Russo A (2000) Impronte di dinosauri in facies di piattaforma interna nel Cretaceo inferiore del Gargano (Puglia, Italia meridionale) (abstract no. 80). Riunione Estiva S.G.I. Trieste, pp 265–266
Gilmore CW (1909) Osteology of the Jurassic reptile Camptosaurus, with a revision of the species of the genus, and descriptions of two new species. Proc US Nat Mus 36:197–302
Grabbe H (1881) Neue Funde von Saurierfährten im Wealdensandstein des Bückeberges. Verh naturhist Ver preuß Rheinl Westf 38(2):161–164
Hagood A, West L (1994) Dinosaur. The Story behind the scenery. KC Publications, Las Vegas
Haubold H (1984) Saurierfährten. Wittenberg Lutherstadt, Neue Brehm Bücherei
Hardenbol J, Thierry J, Farley MB, Jaquin T, De Graciansky P-C, Vail PR (1998) Mesozoic and Cenozoic sequence stratigraphic framework of European basins. SEMP Spec Publ 60:3–13
Holtz RT (2007) Dinosaurs. Random House, New York
Kaever M, de Lapparent AF (1974) Les traces des pas le Dinosaures du Jurassique des Barkhausen (Basse Saxe, Allemagne). Bull Soc Geol Fr 16:516–525
Karl H-V (1997) Schildkröten aus dem Kimmeridge von Nettelstedt und Wallücke/Deutschland (Reptilia, Testudines). Mauritiana 16:289–298
Karl H-V (2006) Über fossile Osteoblasten in Sauropodenknochen – der Einsatz der Mikroskopie in der Wirbeltierpaläontologie. Mikrokosmos 4:199–202
Karl H-V, Staesche U, Tichy G, Lehmann J, Peitz S (2007) Systematik der Schildkröten (Anapsida: Chelonii) aus dem Oberjura und Unterkreide von Norddeutschland. Geol Jb B 98:5–89
Klassen H (1968) Stratigraphie und Fazies des tieferen Malm im Wiehengebirge und Teutoburger Wald. Veröff naturwiss Ver Osnabr 32:39–96
Klassen H (1984) Malm. In: Klassen H (ed) Geologie des Osnabrücker Berglandes. Osnabrück, pp 387–422
Klassen H (2000) Steinbruchgelände westlich Wallücke (Fa. Störmer). Callovium, Oxford, Unter Kimmeridge, Mittelkimmeridge I-IV. In: Ebel R, Klassen H, Metz M (eds) DUGW-Stratigraphische Kommission Subkommission für Jura-Stratigraphie. Jahrestagung in Bünde-Randringhausen. Exkursionsführer, pp 77–78 (unpublished)
Kock K (1998) Das Watt. Lebensraum auf den zweiten Blick. Boyens, Heide
Krumbein WE, Paterson DM, Stal LJ (1994) Biostabilization of Sediments. BIS-Verlag, Oldenburg
Lehmann U (1978) Eine Platte mit Fährten von Iguanodon aus dem Obernkirchner Sandstein (Wealden). Mitt geol-paläont Inst Univ Hamburg 48:101–114
Lockley MG (1991) Tracking dinosaurs. Cambridge University Press, Cambridge
Lockley MG, Meyer CA (2000) Dinosaur tracks and other fossil footprints of Europe. Columbia University Press, New York
Lockley MG, Pittman J (1989) The megatracksite Phenomenon. Soc vert Paleont 9:29–30A
Lockley MG, Prince NK (1988) The Purgatoire Valley Dinosaur Tracksite Region, Southeast Colorado. Geol Soc Am Field Guideb Prof Cont Gold Colorado School Mines 1988:275–278
Lockley MG, Houck K, Prince NK (1986) North America’s largest dinosaur tracksite: Implications for the Morrison Formation Paleoecology. Bull Geol Soc Am 97:1163
Lockley MG, Meyer C, Santos VF (1994a) Megalosauripus, Megalosauropus and the concept of megalosaur footprints. In: Morales, M. (ed) Continental Jurassic Symposium, Museum Northern Arizona, 113–118
Lockley MG, Farlow JO, Meyer CA (1994b) Brontopodus and Parabrontopodus ichnogen. nov. and the significance of wide- and narrow-gauge sauropod trackways. Gaia Rev Geoci Mus Nac Hist Nat Lisbon 10:135–146
Lockley MG, Meyer C, Santos VF (1994c) Trackway evidence for a herd of juvenile sauropods from the Late Jurassic of Portugal. Gaia Rev Geoci Mus Nac Hist Nat Lisbon 10:27–36
Lockley MG, Meyer CA, Hunt AP, Lucas SG (1994d) The distribution of sauropod tracks and trackmakers. Gaia 10:233–248
Lockley MG, Meyer CA, Schultz-Pittman R, Fornay G (1996) Late Jurassic dinosaur tracksites from Central Asia: a preliminary report on the world’s longest trackways. In: Morales, M (ed) Continental Jurassic Symposium Volume, Museum Northern Arizona, Flagstaff, pp 271−273
Malz H (1971) Ein fossiler “Wildwechsel” im Wiehengebirge. Nat Mus 101:431–436
Marty D (2008) Sedimentology, taphonomy, and ichnology of Late Jurassic dinosaur tracks from the Jura carbonate platform (Chevenez-Combe Ronde tracksite, NW Switzerland): insights into the tidal-flat palaeoenvironment and dinosaur diversity, locomotion, and palaeoecology. Geofocus 2:1–278
Meyer CA (1993) A sauropod megatracksite from the Late Jurassic of Northern Switzerland. Ichnos 2:1–10
Meyer CA, Mezga A (2000) A new late Jurassic megatracksite from Northern Switzerland. In: Sachs S, Windolf R (eds) First Symposium on European Dinosaurs. Düsseldorf, pp 19−20
Meyer CA, Lockley MG, Robinson JW, Santos VF (1994) A comparison of well-preserved sauropod tracks from the late Jurassic of Portugal and the Western United States: Evidence and implications. Gaia 10:57–64
Moratalla JJ, García-Mondéjar J, Dos-Santos VF, Lockley MG, Sanz JL, Jiménez S (1994a) Sauropod trackways from the Lower Cretaceous of Spain. Gaia 10:75–83
Moratalla JJ, Sanz JL, Jimenez S (1994b) Dinosaur tracks from the Lower Cretaceous of Regumiel de la Sierra (province of Burgos, Spain): inferences on a new quadrupedal ornithopod trackway. Ichnos 3:89–97
Rainforth EC (2002) Ornithichnology: the ichnotaxonomic legacy of Edward Hitchcock and the status of the grallatorid ichnogenera Grallator, Anchisauripus and Eubrontes. Abstr Geol Soc Am 32(1):31–32
Reineck H-E (1972) Tidal flats. In: Rigby JK, Hamblin WK (eds) Recognition of ancient sedimentary environments. Soc Econ Palaeont Mineral Spec Publ 16:146−159
Reineck H-E, Singh IB (1973) Depositional sedimentary environments. Springer, Berlin Heidelberg New York
Sander PM, Mateus O, Laven T, Knötschke N (2006) Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur. Nature 441:39–741
Santos VF, Lockley MG, Moratalla JJ, Meyer CA, Galopim de Carvalho AM (1998) Dinosaurs tracksites in Portugal: the Jurassic-Cretaceous record. I Encontro Internacional sobre Paleobiologia dos Dinossáurios Lisboa, pp 7–16
Santos VF, Dantas P, Moratalla JJ, Araújo MJ, Galopim de Carvalho AM (2000a) Pegadas de Terópodes em Alcanede, Portugal. I Congresso Ibérico de Paleontologia 16. Jornadas de la Sociedad Española de Paleontología, pp 17
Santos VF, Dantas P, Moratalla JJ, Galopim de Carvalho AM (2000b) Pegadas de Terópodes e de Ornitópodes do Jurássico Superior da Lourinhã, Portugal. I Congresso Ibérico de Paleontologia 16. Jornadas de la Sociedad Española de Paleontología, pp 15−16
Santos VF, Dantas P, Moratalla JJ, Moreira J, Galopim de Carvalho AM (2000c) Pistas de dinossáurio do Kimeridgiano da Serra do Bouro, Portugal. I Congresso Ibérico de Paleontologia 16. Jornadas de la Sociedad Española de Paleontología, pp 18−19
Sarjeant WAS (1974) A history of the study of fossil vertebrate footprints in the British Isles. Palaeogeogr Palaeoclimatol Palaeoecol 16:265–378
Sarjeant WAS, Delair JB, Lockley MG (1998) The footprints of Iguanodon: a history and taxonomic study. Ichnos 6:183–202
Schultka S (1991) Beiträge zur oberjurassischen Flora des Wiehengebirges. Geol Paläont Westf 19:55–93
Schweigert G (1999) Neue biostratigraphische Grundlagen zur Datierung des nordwestdeutschen höheren Malm. Osnab naturw Mitt 25:25–40
von Seebach K (1864) Der Hannoversche Jura. Verlag Wilhelm Hertz, Berlin
Selenka E (1867) Die fossilen Krokodilinen des Kimmeridge von Hannover. Palaeontographica 16:137–144
Struckmann C (1878) Der Obere Jura der Umgebung von Hannover. Eine paläontologisch-geognostisch-statistische Darstellung. Han’ sche Buchhandlung, Hannover
Struckmann C (1880) Vorläufige Nachricht über das Vorkommen grosser vogelähnlicher Thierfährten (Ornithoidichnites) im Hastingssandsteine von Bad Rehburg bei Hannover. N Jb Min Geol Paläont 1880:125–128
Targart E (1846) On markings in the Hastings sands near Hastings, supposed to be footprints of birds. Quart J Geol Soc London 2:267
Thulborn RA (1990) Dinosaur tracks. Chapman and Hall, Melbourne
Unnamed (2002) Ichnologia de Dinossáurios. jazidas. Available at: www.mnhnul.pt/dinos/public_html/Jazidas/index.html
Zeiss A (1991) Ein neuer Aspidoceras-Fund aus dem Oberen Jura Norddeutschlands und seine Bedeutung für die Biosratigraphie des norddeutschen Kimmeridge. Osnabr naturw Mitt 17:87–94
Acknowledgements
My sincere thanks go to Messrs. Störmer (junior and senior) for providing access to the Bergkirchen-Wallücke quarry and for their kind and dedicated support in rescuing track slabs from the mounds in the mining area. They also made possible the integration of the discoveries into the permanent inventory of the Dobergmuseums federation. Studies on the Barkhausen tracksite were made possible by the support of the city Bad Essen (Mayor G. Harmeyer) and the UNESCO-Geopark-Management (H. Escher). H. Breitkreutz discovered the important vertebrate remains, such as dinosaur and pterosaur bones, shark spines, and turtle and crocodile remains, in the Lower Kimmeridgian outcrop at Nettelstedt; I am grateful for his kindness in obtaining casts of the most important material for the permanent exhibition in the new Dobergmuseum/Geologisches Museum Ostwestfalen-Lippe in Bünde, and for allowing me to study and report on his discoveries and to take some photos of selected bones. I am also grateful to Dr. U. Menkveldt of the Naturhistorisches Museum, Bern and Dr. E. Müller-Merz of the Naturhistorischen Museum, Solothurn for making it possible for me to study the Kimmeridgian invertebrate and vertebrate material from Swiss sites, held in their collections. The reviewer Dr. S. de Valais and an anonymous reviewer greatly improved the manuscript. I thank E. Manning for his support. Finally, G. “Rinaldino” Teichmann allowed the use of his dinosaur fauna illustration.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Diedrich, C. Upper Jurassic tidal flat megatracksites of Germany—coastal dinosaur migration highways between European islands, and a review of the dinosaur footprints. Palaeobio Palaeoenv 91, 129–155 (2011). https://doi.org/10.1007/s12549-010-0044-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-010-0044-y