
Abstract
Reconstruction of fossil teleost faunas can provide important information on palaeoenvironments, palaeogeography and evolution, and otoliths are particularly useful for that purpose. Here we present an otolith-based fish fauna from the middle Miocene of the Eastern Paratethys, i.e. the Karagaily section of the Mangyshlak Peninsula in Western Kazakhstan, and report on the accompanying nannoplankton, foraminifera, molluscs and ostracods. A total of 30 teleost species are described and figured, including ten new species: Alosa paulicrenata n.sp., Morone? bannikovi n.sp., Centracanthus pobedinae n.sp., Genyonemus? karagiensis n.sp., Trewasciaena suzini n.sp., Parablennius prokofievi n.sp., Aphia djafarovae n.sp., Neogobius udovichenkoi n.sp., Ponticola zosimovichi n.sp., Pomatoschistus bunyatovi n.sp. Nannoplankton and gastropods indicate a Konkian (late Badenian, early Serravallian) age for this fish assemblage. The dominance of Gadidae and Gobiidae, together with the composition of the nannoplankton, indicates an inner-neritic to coastal environment with high productivity. The Konkian fish fauna of the Eastern Paratethys shows a high degree of autonomy relative to approximately contemporaneous fish faunas from the Central Paratethys and other European basins. This confirms that the Konkian was a time of limited faunal exchange between the Central and Eastern Paratethys, while a marine connection may have persisted between the Central Paratethys and the northern Mediterranean. We conclude that the fish fauna reported here records an early endemic development in the Eastern Paratethys during the middle Miocene (Konkian). The disappearance of Bregmacerotidae and Gonostomatidae (Bonapartia) during the preceding stage of the Karaganian and the first appearance of Palimphemus minusculoides in the Konkian are important biostratigraphical markers.
Kurzfassung
Die Rekonstruktion fossiler Teleostei Faunen kann wichtige Erkenntnisse zur Paläoumwelt, Paläogeographie und Evolution liefern und Otolithen spielen hierfür eine besonders wichtige Rolle. In dieser Arbeit wird eine auf Otolithen basierende Fischfauna aus dem Mittel-Miozän der Östlichen Paratethys (Karagaily-Aufschluss, Mangyshlak Halbinsel, West-Kasachstan) vorgestellt und eine Übersicht über das begleitende Nannoplankton, Foraminiferen, Mollusken und Ostrakoden gegeben. 30 Teleostei-Arten werden beschrieben und abgebildet, darunter 10 neue Arten: Alosa paulicrenata n.sp., Morone? bannikovi n.sp., Centracanthus pobedinae n.sp., Genyonemus? karagiensis n.sp., Trewasciaena suzini n.sp., Parablennius prokofievi n.sp., Aphia djafarovae n.sp., Neogobius udovichenkoi n.sp., Ponticola zosimovichi n.sp., Pomatoschistus bunyatovi n.sp.. Auf der Basis des Nannoplankton und der Gastropoden ist die Fischgemeinschaft zeitlich in die Stufe des Konkium (spätes Badenium, frühes Serravallium) einzuordnen. Die dominierenden Gadiden und Gobiiden sowie die Zusammensetzung der Nannoplankton-Gemeinschaft lassen auf ein inner-neritisches bis küstennahes, nährstoffreiches und produktives Paläomilieu schließen. Insgesamt zeigt die Fischfauna des Konkium der Östlichen Paratethys einen hohen Anteil endemischer Arten im Vergleich zu gleichaltrigen Fischfaunen der Zentralen Paratethys und anderer Europäischer Becken. Dies bestätigt die Annahme eines begrenzten Faunenaustauschs zwischen der Östlichen und der Zentralen Paratethys zur Zeit des Mittel-Miozäns, während eine marine Verbindung zwischen der Zentralen Paratethys und dem nördlichen Mittelmeer offenbar weiterhin existierte. Die hier bearbeitete Fischfauna lässt weiterhin darauf schließen, dass die endemische Entwicklung der Östlichen Paratethys schon zu einem relativ frühen Zeitpunkt des Mittel-Miozäns (Konkium) begann. Das Verschwinden der Bregmacerotidae und Gonostomatidae (Bonapartia) in der vorherigen Stufe des Karaganium und das erste Auftreten von Palimphemus minusculoides im Konkium sind wichtige biostratigraphische Kriterien.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Eastern Paratethys was a marine-to-brackish inland water body that extended from Bulgaria and Romania across the Black Sea and the lowlands north of the Caucasus to the Caspian Sea during the Neogene (Rögl and Steininger 1983; Rögl 1998; Popov et al. 2006). It is well known for the endemic evolution of its biota that was initiated when the Paratethys was separated from the Tethys during the middle Miocene (e.g. Rögl 1998; Harzhauser et al. 2002, 2003). Nowadays, an endemic teleost fauna consisting primarily of species of the Clupeidae and Gobiidae exists in the Caspian Sea and a much more impoverished endemic fauna occurs in the brackish water and freshwater tributaries of the Black Sea (Hoestlandt 1991; Miller 2003, 2004). Previous studies have shown that reconstruction of fossil fish faunas from the Eastern Paratethys can provide important information on the evolution of endemics during the middle and late Miocene (Carnevale et al. 2006; Schwarzhans 2014b). In this context, otoliths are particularly useful for reconstructing fossil fish faunas because they are highly informative and much more abundant than fossil skeletons in deposits laid down since the late Cretaceous.
This study presents the first otolith-based description of a fish fauna from the Konkian (lower Serravallian) deposits of Mangyshlak, Kazakhstan, which was part of the Eastern Paratethys at that time. We provide new stratigraphical data, mainly derived from nannoplankton and other accompanying fossils, and discuss their palaeoenvironmental and palaeogeographic implications based on the fish fauna from the studied section. The Konkian interval is especially interesting because it precedes the severe changes in water chemistry and the consequent transformation of local ecosystems during the succeeding Sarmatian s. l. (Pisera 1996; Kowalke and Harzhauser 2004; Carnevale et al. 2006), and very little is known about its fish faunas.
Fossil otoliths from Neogene strata of the Eastern Paratethys have been studied since the 1940s. Material from Romania has been described by Pana (1965, 1977, 1982, 1995), samples from Bulgaria by Strashimirov (1972, 1980, 1981a, b, 1982, 1984, 1985a, b), specimens from Moldavia by Paghida (1962), Paghida-Trelea et al. (1967) and Paghida-Trelea (1969), and examples from Azerbaijan by Chalilov (1946), Pobedina (1954), Pobedina et al. (1956), Klein (1960) and Djafarova (2006). Works by Weiler (1943, 1949, 1950) and Rado (1965, 1968, 1969, 1971, 1981) were based on otoliths obtained from the eastern part of the Central Paratethys in Romania, and are also relevant to the current study. The works of Pobedina, Strashimirov and Djafarova account for the bulk of the otolith-based species from the Eastern Paratethys. Another, apparently very substantial, collection from the Crimea and the Russian northern Caucasus was studied by Suzin but was unfortunately never published; only citations of an internal report dated to 1939 in the holdings of the Azerbaijanian Oil Institute (AzNIPINeft) are now available. In 1968, Suzin (in Zhizhchenko) published figures depicting 46 new species. However, no descriptions were given, so that, with the exception of a few which have been validated through descriptions in later publications (Klein 1960; Strashimirov 1972, 1980, 1981a, b, 1984), these species names are not available in the sense of the ICZN (Article 13.1.1). Moreover, this impressive list of publications dealing with fossil otoliths from the Neogene of the Eastern Paratethys remains problematic in several respects, such as limited access to relevant literature at the time the reports appeared, a lack of Recent comparative material, or a predominance of juveniles among the specimens studied. A thorough revision of many previously described species is therefore required.
Fossil fish skeletons from Neogene strata of the Eastern Paratethys have been described by Bogatshov from Azerbaijan (1929, 1933, 1938), southern Russia (1942) and eastern Ukraine (1955). Further studies on fossil skeletons from the Neogene of the Eastern Paratethys include those reported by Menner (1948), Svichenskaya (1957, 1959, 1973), Fedotov (1976), Daniltchenko (1986), Bannikov (1993, 2006, 2008, 2009, 2010), Baykina (2013), Carnevale and Harzhauser (2013), Carnevale et al. (2006, 2012), Prokofiev (2002, 2004, 2005, 2007), and Sytchevskaya and Prokofiev (2007, 2010), all of which served as a basis for the correlation of otolith and skeleton data in this study. However, data on fish skeletons from the Konkian of the Eastern Paratethys is comparatively sparse (see Daniltchenko 1986). Clupeonella sp., Protonymus gontsharovae Sytchevskya and Prokofiev, 2007 and Mugil finitimus Svichenskaya, 1973 are the only skeleton records from the Konkian deposits of the Taman Peninsula and the northern Caucasus. The latter was also found in the Konkian of Mangyshlak (Kaundy Depression) (Svichenskaya 1973; Bannikov 2010).
The otoliths from the Konkian (lower Serravallian) of Karagaily studied here are exceptionally species-rich, well-preserved and large-sized when compared with previously published data. The otolith collection contains 30 species, ten of which are new to science and six remain in open nomenclature. With the help of Djafarova’sFootnote 1 monographic review, it was possible to collate the new findings with older descriptions, arrive at a better understanding of many previously described and stratigraphically contemporaneous species, and allow for a proper redescription and definition of the recognised species.
Institutional abbreviations used are: AMS, Australian Museum, Sydney, New South Wales, Australia; BMNH, Natural History Museum, London, United Kingdom; NMNH, National Museum of Natural History of the National Academy of Sciences of Ukraine, Kyiv; SMF, Senckenberg Museum, Frankfurt/Main, Germany; ULUZ and KUZU Luhansk Taras Shevchenko National University, Ukraine; WAM, Western Australian Museum, Perth, West Australia, Australia; ZMH, Zoological Museum, Hamburg, Germany; ZMUC, Zoological Museum, Copenhagen, Denmark.
Regional geology
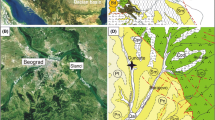
Previous work on the stratigraphy and palaeontology of the Neogene deposits from Mangyshlak includes the studies of Eichwald (1834), Barbot-de-Marni (1889), Andrusov (1915, 1917), Bajarunas (1917), Kolesnikov et al. (1940), Merklin (1953, 1954), Merklin and Nevesskaia (1955), Merklin et al. (1960), Ilyina and Utkin (1963) and others. One of the most comprehensive recent contributions was published by Liverovskaya (1960). She noted that the Miocene deposits of Mangyshlak consistently begin with the middle Miocene (commonly Karaganian and Konkian, partially Sarmatian) and that these beds are often transgressive and unconformably overlie Oligocene, Eocene or Cretaceous sediments. In addition, she presented several detailed lithological columns of the Paleogene and Neogene deposits of South Mangyshlak, including the outcrops along the northern rim of the Karagie Depression (Elyktay, Burliu, Aksenger), which are located close to the Karagaily Hill outcrop sampled here (Fig. 1). Accordingly, the Konkian sediments have an average thickness of about 10 m and a maximum thickness of 25 m, and encompass grey clays and marls with intercalations of soft coquina and stromatolite concretions in places. Directly above are lower Sarmatian sediments 5–10 m in thickness, comprising alternations of clays, marls, coquina and sands. Clays with intercalated layers of sands, marls and occasionally also limestones represent the middle Sarmatian deposits; their thicknesses vary between 20 and 40 m. The upper Sarmatian largely consists of limestones and is up to 15 m thick (for details see Liverovskaya 1960).

Location map of the Karagaily Hill, Karagie Depression, Mangyshlak Peninsula, Western Kazakhstan
Study site
The following is a detailed description of the sedimentary section of the northern cliffs of the Karagaily outcrop (43°50′33.96″N, 51°35′26.61″E.). The outcrop cliffs form a steep ravine, which opens northwards to the road to the water-pump station Kuyulus. The lithological succession is described from top to bottom (Figs. 2, 3):

Overview of the Karagaily section

1 Overview of Karagaily Hill. 2, 3 Composite photograph of the upper part of the Karagaily section. 4 Sampling of productive levels by N. Udovichenko
Level 1 Conglomerate, brown-yellow, discordant on underlying strata and probably of Quaternary age (visible thickness 2 m).
Level 2 Clay, olive-green, with some thin (to 0.5 m) yellow interlayers; Sarmatian; thickness 10 m.
Level 3 Shelly limestone, with dissolved shells, brownish yellow; Sarmatian; thickness 0.2–0.3 m.
Level 4 Clay, green with thin (0.1 m) grey interlayer in the upper part; Sarmatian; thickness 0.6 m.
Level 5 Marl, light grey, solid, firm, with imprints of bivalves, lower boundary uneven; Sarmatian; thickness 0.5 m.
Level 6 Clay, light yellow grey; probably Sarmatian; thickness 0.4 m.
Level 7 Shelly limestone, rusty brown; probably Sarmatian; thickness 0.1 m.
Level 8 Clay, brown, laminated, with bivalve shells on the bedding surface; Konkian; thickness 1 m.
Level 9 Marl, mustard-yellow, with thin (0.03–0.07 m) white interlayer of coquina, light grey, friable; Konkian; thickness 0.2–0.3 m. (sample no. 1).
Level 10 Clay, light grey, with imprints of bivalves. Distinct fossil rich lens about 0.2 m below, light grey, friable; Konkian; thickness 2.5 m (sample no. 2).
Level 11 Marl, light grey, solid, firm; Konkian; thickness 0.5 m.
Level 12 Clay, light grey, with imprints of bivalves; Konkian; thickness 2 m.
Level 13 Marl, light grey, with imprints of bivalves; Konkian; thickness 0.3 m.
Level 14. Clay, light grey, with imprints of bivalves; Konkian; thickness 0.5 m.
Level 15 Limestone, light grey, with reddish-brown spots, mollusc detritus and shells, bioturbation with loaf-like structure; Konkian; thickness 0.5 m.
Level 16 Clay, light grey; Konkian; thickness 1.5 m.
Level 17 Marl, greenish-grey, solid, firm; Konkian; thickness 0.3 m.
Level 18 Clay, green, with shells of bivalves. Interlayer (up to 0.3 m) of light grey clay in the middle part of section; Konkian; thickness 5 m.
Level 19 Marl, light grey, with nodular very firm concretions up to 0.5 m length; Konkian; thickness 0.6 m.
Level 20 Clay, dark green, with shells of bivalves. Two thin (up to 0.2 m), light yellow and soot-black (?sulphidic) interlayers about 1 and 2.5 m below top. There are siderite concretions lower in this level. The lower contact is not exposed; it is probably Karaganian according to a microfossil sample taken near the top of the section; visible thickness 20 m.
Materials and methods
The otolith-bearing samples were collected during two field-trips in 2013 to the Northern Karagie Depression (Fig. 3) by the senior author and N. Udovichenko. Presence of gastropods in the sediments was used as criterion for otolith prospection. Two particularly prospective coquina layers from Levels 9 and 10 were sampled with 15 and 55 kg of bulk sediment, respectively. The sediment was sieved to the 0.5-mm fraction in the field. The derived concentrate was picked at the laboratory and yielded foraminifera, ostracods, molluscs, remains of crabs, shark tooth (probably belonging to the Alopiidae, oral comm. by N. Udovichenko), bones and a total of about 2000 otoliths, of which 1788 specimens were identified to the species level.
The terminology for the morphological description of the otoliths follows Koken (1891), Weiler (1942) and Schwarzhans (1978). The morphometric measurements follow Schwarzhans (2013). The following abbreviations are used: otolith length = OL; otolith height = OH; otolith thickness = OT; ostium length = OsL; cauda length = CaL; ostial colliculum length = OCL; caudal colliculum length = CCL; sulcus length = SuL. The caudal curvature index is measured as a ratio of the horizontal stretch of the cauda against the length of the inclined portion. The rostrum length is measured from the tip of the rostrum to the level of the deepest point of incision of the excisura, or, in the absence of an excisura, to the point where the dorsal margin of the ostium meets the otolith rim, and is calculated as a percentage of OL. For otoliths of Gobiidae, the curvature index of the inner face is calculated as a percentage of OL.
Photos of otoliths were taken with a Leica M 165 FC stereomicroscope in the Department for Earth and Environmental Sciences, Palaeontology and Geobiology, Ludwig-Maximilians-Universität München. SEM pictures of some very small otoliths were produced (SEM of the Zoological State Collection, Munich) (Fig. 9-1A, -2, -5, -6A, -9). For optimal comparison, all figures show otoliths from the right side, except for those of the Pleuronectiformes, which may depict lateral dimorphism. Photos of left otoliths were therefore mirrored and are marked accordingly in the figure captions.
The otoliths are deposited in the collection of the National Museum of Natural History of the National Academy of Sciences of Ukraine, Kyiv, NMNH, in the Department of Monographic Collection under the collection registrations NMNH 2532/1 to 2532/176.
Nannoplankton samples were prepared using the standard preparation method (Perch-Nielsen 1985) and examined using a Leica DMLP microscope under 1000× magnification. The standard zonation of Martini (1971) was applied.
Systematic palaeontology
Remarks
Generic allocation of fossil otoliths depends very much on comparison with Recent material, except for those rare instances of fossil skeletons described with otoliths in situ. As a consequence, the accuracy of attribution of fossil otolith-based species to (extant) genera decreases with geological age and has led specialists in the field of otolith research to propose alternative open generic nomenclature schemes in order to avoid erecting many otolith-based fossil genera. Names were then built by applying genitive plural forms of the family or a higher taxonomic unit into which the fossil otolith-based species could be placed with the acceptance of the author, for example Otolithus (Albulidarum) circularis or “genus Albulidarum” circularis for a given species considered to be an albulid of unknown generic relationship. It has long been known that this practice is not compliant with the regulations of the ICZN (see Schwarzhans 2012 for a detailed discussion). Recently, Nolf (2013) reviewed this practice and proposed an alternative scheme—“Albulida” circularis, meaning “an albulid” of unknown generic relationship. In a book review of Nolf’s work from 2013, Tracey (2014) concluded that this new practice would have to be considered “zoological formulae” according to article 1.3.7 of the ICZN and thus, in the usage proposed by Nolf, would not be compliant with the ICZN. We have therefore refrained from any of these practices in cases of unresolved generic allocations, which are, however, rather uncommon for Miocene otoliths. Instead, we follow Janssen (2012) by using an unambiguous genus name, i.e. the name of the type genus of the family, followed by a question mark, indicating that this species might as well belong to any other known or unknown genus in the family.
The classification of the systematic part follows Nelson (2006).
Class Osteichthyes Huxley, 1880
Subclass Actinopterygii Klein, 1885
Order Albuliformes Jordan, 1923
Suborder Albuloidei Jordan, 1923
Family Pterothrissidae Gill, 1893
Genus Pterothrissus Hilgendorf, 1877
Type species. Pterothrissus gissu Hilgendorf, 1877, type by monotypy, Japan seas; Recent.
Pterothrissus sp.
Figure 4-1

1 Pterothrissus sp., NMNH 2532/070. Figure is mirrored. 2, 3 Alosa paulicrenata n.sp. Bratishko, Schwarzhans and Reichenbacher. 2: Holotype, NMNH 2532/104, ventral view (2B). 3: Paratype, NMNH 2532/105. Figure 3 is mirrored. 4, 5 Sardina? sp. 4: NMNH 2532/106. 5: NMNH 2532/168. Figure 4 is mirrored. 6–12 Onogadus simplicissimus (Schubert, 1906). 6: NMNH 2532/011, ventral view (6B). 7: NMNH 2532/013, ventral view (7B). 8: NMNH 2532/010, ventral view (8B). 9: NMNH 2532/014. 10: NMNH 2532/016. 11: NMNH 2532/012. 12: NMNH 2532/019. Figures 6, 8, 10–12 are mirrored. 13 Onogadus argentatus (Reinhardt, 1837), AFORO Database, fish ID 9994. Figure is mirrored. 14–21 Palimphemus minusculoides (Schubert, 1912). 14: NMNH 2532/001, ventral view (14B). 15: NMNH 2532/003, ventral view (15B). 16: NMNH 2532/007, ventral view (16B). 17: NMNH 2532/149. 18: NMNH 2532/152, outer view (18B). 19: NMNH 2532/158. 20: NMNH 2532/009. 21: NMNH 2532/151. Figures 18, 19, 21 are mirrored
Material One juvenile specimen NMNH 2532/070, Karagaily outcrop, Level 10.
Discussion The single small specimen of about 2 mm in length cannot be identified at the species level. The rather short ostium, the slightly bent cauda and blunt, nearly vertical anterior rim appear to be characteristic. A similar small otolith has been described as P. umbonatus (Koken 1884) by Djafarova (2006) from the Maeotian of Azerbaijan. It differs from the specimen presented here in the less blunt shape of the anterior rim. Nevertheless, these two finds indicate the persistence of the genus Pterothrissus during the middle and late Miocene in the Eastern Paratethys. Otoliths of P. umbonatus, one of the most long-ranging otolith-based species known from various European basins from the middle Eocene to the middle Miocene, differ from Pterothrissus sp. in the almost equal length of the ostium and cauda (OsL:CaL = 0.5 vs. 0.8–0.9). Juvenile otoliths of P. umbonatus differ also in the rich ornamentation of rims, outer face and part of the inner face (Schwarzhans 2010). Pterothrissus caspiensis Bratishko and Udovichenko, 2013 from the Rupelian of Mangyshlak represents a different lineage within this genus, and is characterised by a distinctly convex inner and a flat to concave outer face (vs nearly flat inner and convex outer faces in Pterothrissus sp.).
Order Clupeiformes Bleeker, 1859
Suborder Clupeioidei Bleeker, 1859
Family Clupeidae Rafinesque, 1810
Genus Alosa Linck, 1790
Type species. Clupea alosa Linnaeus, 1758, no type species initially mentioned, type species added by Cuvier, 1829, apparently by subsequent absolute tautonymy, European Ocean; Recent.
Alosa paulicrenata n.sp. Bratishko, Schwarzhans and Reichenbacher
Figure 4-2, -3
Etymology From Latin crenatus, crenate, incised, combined with paulum indicating a small excisura.
Type material Holotype: NMNH 2532/104 (Fig. 4-2). Paratype: 1 specimen NMNH 2532/105 (Fig. 4-3)
Type locality Karagaily outcrop, Level 10; Mangyshlak, Kazakhstan.
Age Konkian, early Serravallian, middle Miocene.
Diagnosis Elongate, thin otoliths; OL:OH = 1.9–2.0. Rostrum moderately long, 35–42 % of OL, blunt, with small projecting tip situated just below the ostium. No or minute excisura. Strong postdorsal projection. Ventral rim with few strong denticles in the middle. Ostium about 1.6–2.0 times the length of cauda; anterior margin of ostium not reaching tip of rostrum. Cauda terminating at some distance from posterior rim.
Description (n = 2). Elongate, thin otoliths reaching about 2 mm of length; OL:OT about 6.5. Rostrum 35–42 % of OL. Posterior rim rounded, postdorsally projected (Fig. 4-3). Dorsal rim short, shallow, slightly curved; antirostrum positioned high on dorsal rim. Ventral rim long, shallow, curved upward along posterior third, undulating, with 2–3 massive denticles at mid-section. Excisura absent (Fig. 4-3) or very small (Fig. 4-2), resulting in a regularly inclined predorsal rim along the opening of the sulcus.
Inner face almost flat with centrally positioned deep and wide sulcus. Ostium wide, opened anteriorly, long, its upper rim short, upward directed at an angle of 30–45° to meet antirostrum. Ostium ventrally underpinned by distinct crista inferior. Cauda short, deep, separated from ostium at collum by a ventral incision from ventral and a shallow dorsal notch on dorsal sulcus rims. Small dorsal depression above central portion of dorsal sulcus rim; no ventral furrow. Outer face flat with few and indistinct radial furrows.
Comparison Otoliths of A. paulicrenata resemble the Recent European species A. alosa (Linnaeus, 1758) and A. fallax (Lacepède, 1803) (see Lombarte et al. 2006; Tuset et al. 2008) in the blunt rostrum with the convex curvature of the anterior upper rim of the ostium and small inferior tip, and also in the postdorsal projection. It differs from the Recent species in the robust denticles on the mid-ventral rim and the lack of a deep notch just below the posterior tip. Another characteristic trait in comparison with many other otoliths of Recent and fossil clupeid otoliths species is the relatively small excisura (see Smale et al. 1995; Nolf 2013)
Genus Sardina Antipa, 1906
Type species. Sardina dobrogica Antipa, 1906 (synonym of Clupea pilchardus Walbaum, 1792), type by monotypy, Black Sea, Romania; Recent.
Sardina? sp.
Figure 4-4, -5
Material 12 specimens (NMNH 2532/106, 107, 168), Karagaily outcrop, Level 10.
Discussion None of the specimens are fully preserved, with the rostrum always broken off to some extent. Characteristic traits of these otoliths are the well-developed, large antirostrum that is directed anteriorly and underpinned by a deep excisura, and the convex shape of the ventral rim with anteroventral crenulations. The otoliths resemble those of the Recent Sardina pilchardus (Walbaum, 1792) (see Lombarte et al. 2006) except for the deep excisura and the convex shape of the ventral rim. Similar otoliths have been described from the Chokrakian (early Badenian) of Bulgaria as Gobius latirostratus Strashimirov, 1980 and G. latirostratus ellipticus Strashimirov, 1980. However, the latter was based on a non-diagnostic juvenile otolith, and the former displays a much shallower excisura and also a shorter antirostrum than the species presented here.
Order Gadiformes Goodrich, 1909
Family Gaidropsaridae Jordan and Evermann, 1898
Genus Onogadus de Buen, 1934
Type species. Onogadus ensis Reinhardt, 1837, type by monotypy, Greenland; Recent.
Onogadus simplicissimus (Schubert, 1906)
Figure 4-6 to -12
1906 Otolithus (Crenilabrus) simplicissimus; Schubert 1906: pl. 18, figs. 43, 44.
1966 Crenilabrus simplicissimus Schubert, 1906; Smigielska 1966: pl. 18, figs. 5, 6.
1974 Bodianus cf. simplicissimus (Schubert, 1906); Brzobohaty and Stancu 1974: pl. 1, fig. 4.
1981 Crenilabrus simplicissimus Schubert, 1906; Nolf 1981 [species rejected by Nolf]
?1984 Bodianus josephinae; Strashimirov 1984: pl. 4, figs. 44, 45.
Material 75 specimens (NMNH 2532/010–019), Karagaily outcrop, Level 10.
Remark A detailed diagnosis and re-description of this species is given here, because Schubert’s documentation is not optimal for species recognition.
Diagnosis Elongate otoliths with marked dorsal angle at about middle of otolith. OL:OH ranging from 2.1 to 2.4, increasing up to 2.8 in the largest specimens. Anterior-ventral tip often projecting rostrum-like; posterior tip broadly rounded. Inner face slightly convex and very slightly twisted posteriorly. Ostial and caudal colliculum long, caudal colliculum slightly wider (particularly ventrally) than ostial colliculum, slightly inclined.
Description (n = 10). Elongate otoliths reaching about 4 mm in length. Outline triangular with broad but distinct mediodorsal angle, pointed and often projecting anterior-ventral tip and broadly rounded posterior tip (not rounded in specimen NMNH 2532/012, see Fig. 4-11, which is only tentatively referred to O. simplicissimus). Ventral rim only slightly curved. All rims smooth or very slightly undulating.
Inner face slightly convex and very slightly twisted posteriorly (the latter particularly well seen in ventral views). Sulcus straight, with slightly inclined median position, divided into well-marked ostium and cauda of about equal lengths, each filled with a slightly deepened colliculum. CCL:OCL = 1.15–1.4. Ostium anteriorly widened, ostial colliculum not opening towards anterior rim, tapering towards narrow and short collum. Cauda slightly inclined, clearly terminating prior to posterior tip of otolith, slightly widened posteriorly and ventrally, tapering towards collum. Short triangular dorsal depression only above central part of sulcus. Ventral furrow moderately strong, nearly straight inward of mid-section of ventral rim of otolith. Outer face flat to slightly concave except for small umbo at about level of collum on inner face, smooth.
Variability and ontogeny The otoliths of O. simplicissimus exhibit a considerable degree of variability in the expression of the anterior tip of the otolith and the length to height ratio (OL:OH). The latter is also strongly affected by ontogenetic variation. Otoliths ≤2 mm in length show OL:OH ratios of 2.1–2.3, whereas larger specimens (up to nearly 4 mm in length) show OL:OH ratios of 2.3–2.6, and in rare instances even up to 2.8 (Fig. 4-8). Also, the size of the anterior-ventral rostrum-like projection increases considerably with the otolith length.
Discussion When Nolf (1981) reviewed the type specimens of Schubert, he noted Crenilabrus simplicissimus as follows (translated in senso lato from French): “The specimen of fig. 43 of Schubert is an otolith of Gaidropsarus Rafinesque, 1810, of which the inner face is strongly eroded. The one of fig. 14 [probably erroneous for fig. 44] is a very small specimen, difficult to interpret, but possibly representing another species of Gaidropsarus than fig. 43. In order to avoid any future confusion the specimen GBW 1906/01/19a (fig. 43 of Schubert) is selected as lectotype.” He then goes on to reject the species (Nolf 1981, 1985). However, the new otolith material from Mangyshlak suggests that the two otoliths figured in Schubert (1906: pl. 18, figs. 43–44) represent a single species of the family Gaidropsaridae, which shows considerable variability and ontogenetic allometry. This species existed in the Serravallian of the Paratethys (possibly from the middle Badenian—Karsy and Rybnica localities, see Smigielska, 1966) and the original name introduced by Schubert can be applied. However, selection of a neotype might be advisable. Radwanska (1992) described similar otoliths from the lower Badenian of Poland as Ciliata sp. and Gaidropsarus acuticaudatus Gaemers, 1973, which are similar to Onogadus simplicissimus but show a wider sulcus according to her photographs (pl. 8, figs. 1–5). She does not mention whether she observed a twist along the horizontal axis of the inner face of the otoliths, nor did she show the intensity of the curvature of inner and outer faces, as observed in specimens from the contemporaneous Hemmoorian of the North Sea Basin, from where Gaidropsarus acuticaudatus was originally described (see Schwarzhans 2010 for figures). Until Radwanska’s specimens have been reviewed, we believe that they should not be considered to represent O. simplicissimus as re-defined here.
Within Gaidropsaridae, two main patterns of otolith morphology are observed: one with thin otoliths with convex inner and concave outer faces, an elongate outline, a low and rounded mediodorsal rim, a wide cauda and a notable twist along the horizontal axis, found in the genus Gaidropsarus and in Ciliata mustella; the other with thick otoliths with a flat or slightly convex inner and a flat or slightly convex outer face, a moderately elongate to compressed outline with a distinct mediodorsal angle, a rather narrow cauda and no or only a slight twist along the horizontal axis. This second group comprises the genera Enchelyopus and Onogadus as well as Ciliata septentrionalis. Note that we follow Svetovidov (1986a, in Whitehead et al.) here in the recognition of Onogadus as a separate genus based on the differences between otoliths of Gaidropsarus and Onogadus described above. This is in contrast to Svetovidov (1986b), who regarded Onogadus as a junior synonym of Gaidropsarus in his review of the genus Gaidropsarus, and also in contrast to Eschmeyer and Fong (2013), who follow the interpretation of Svetovidov (1986b). However, a recent molecular-based phylogenetic analysis showed that Gaidropsarus ensis and G. argentatus, which represent species of Onogadus in the sense of Svetovidov (1986a), form a separate clade among the species of Gaidropsarus analysed (Francisco et al. 2014). This reinforces our assumption tthat Onogadus should be recognised as a separate genus.
Otoliths of Recent specimens of Onogadus argentatus (as Gaidropsarus argentatus), O. ensis (as Gaidropsarus ensis), Ciliata mustella, Enchelyopus cimbrius, Gaidropsarus biscayensis, G. mediterraneus and G. vulgaris are figured in Lombarte et al. (2006). Among them, the otoliths of O. argentatus (Reinhardt, 1837) (see fig. 4.13) are more similar to the fossil species studied here than those of O. ensis, but differ in a more compressed shape (OL:OH = 1.9–2.2 vs 2.1–2.8) and a relatively short sulcus (<70 % of otolith length vs >80 %).
Similar fossil otolith-based species of the Gaidropsaridae have been described from the middle and late Miocene of the North Sea Basin: Ciliata crimmeni Schwarzhans, 2010 from the Tortonian is related to the Recent C. septentrionalis (recently tentatively and erroneously synonymised with C. mustela by Nolf 2013) and differs from O. simplicissimus in the symmetrically developed anterior and posterior tips and the more downward-inclined cauda; Onogadus asper (Gaemers and Schwarzhans, 1973) differs in the anterior-ventral projection being considerably shorter than the posterior-ventral projection and the sulcus terminating further apart from the anterior and posterior rims than in O. simplicissimus; Enchelyopus gaemersi Schwarzhans, 2010 from the Langhian is recognised by the much more forward-positioned dorsal angle, the more compressed shape (OL:OH = 1.8–2.1 vs 2.1–2.8) and the relatively long cauda (CCL:OCL = 1.45–1.65 vs 1.15–1.4).
Recently, the skeleton-based species Gaidropsarus pilleri Carnevale and Harzhauser, 2013 has been described from the early Badenian (Langhian) of Austria. This fossil species appears to be related to the Recent G. biscayensis and G. capensis based on meristic characters. Otoliths of both Recent species represent the first otolith pattern as described above, and hence we consider G. pilleri to be unrelated to any of the fossil otolith-based species mentioned above.
Bodianus josephinae Strashimirov, 1984 was described from the early Sarmatian from Bulgaria based on a unique small otolith 1.35 mm in length, and is tentatively placed in synonymy with O. simplicissimus since it shares the rostrum-like projection of the anterior-ventral tip but is slightly more compressed than the specimens studied from Mangyshlak (OL:OH = 2.0 vs 2.1–2.8). Onogadus simplicissimus is now recorded from the Serravallian (late Badenian/Konkian to early Sarmatian s.l.) of the Central and Eastern Paratethys.
Family Gadidae Rafinesque, 1810
Genus Palimphemus Kner, 1862
Type species. Palimphemus anceps Kner, 1862, type by monotypy, St. Margarethen, Austria; Leitha Limestone, early Badenian, Langhian, middle Miocene.
Remark The association of certain fossil otolith-based species with this fossil skeleton-based genus has been concluded from the study of otoliths found in situ in Palimphemus anceps (see Schwarzhans 2014a).
Palimphemus minusculoides (Schubert, 1912)
Figure 4-14 to -21
1906 Otolithus (Gadidarum) minusculus; Schubert 1906 (part): pl. 19, figs. 48, 49, ?50 (non 51, 52).
1912 Otolithus (Gadus) minusculoides; Schubert 1912: fig. 16.
1943 Gadus? minusculoides Schubert, 1912; Weiler 1943: pl. 1, figs. 43, 47.
1949 Gadus? minusculoides Schubert, 1912; Weiler 1949: pl. 5, figs. 43, 47.
1968 Otolithus (Gadidarum) angustus; Suzin 1968 (in Zhizhchenko): pl. 18, fig. 29; [name not available: ICZN article 13.1.1].
?1969 Macruridarum minusculus Schubert, 1906; Stancu 1969: pl. 3, figs. 2, 3.
?1974 Otolithus (Macruridarum?) minusculus (Schubert, 1906); Brzobohaty and Stancu 1974: pl. 2, figs. 11, 12.
1981 Otolithus (Gadidarum) minusculus Schubert, 1906 (part, see above); Nolf 1981 [species rejected by Nolf].
1981 Gadus minusculoides Schubert, 1912; Nolf 1981 [species rejected by Nolf]
?2006 Micromesistius sp.; Carnevale et al., 2006: fig. 6 (otolith in situ).
2006 Palaeogadus emarginatus (Koken, 1884); Djafarova 2006: pl. 8, fig. 4, pl. 10, figs. 2, 3.
2013 Micromesistius arcuatus Radwanska, 1992; Schultz 2013: pl. 78, fig. 3 (refigured holotype of Gadus minusculoides).
Material 1253 specimens (NMNH 2532/001–009, 147–165), Karagaily, Level 10; 5 specimens, Karagaily, Level 9.
Remark A detailed diagnosis and re-description of this species is given here because Schubert’s documentation is not optimal for species recognition.
Diagnosis Elongate, thin otoliths with a nearly flat to slightly bent inner face. Anterior tip slightly pointed or rounded, posterior tip tapering and pointed or rounded. Dorsal rim slightly convex rounded anteriorly, nearly horizontal in the median part, with a slight postdorsal angle close to posterior tip. Ostial colliculum anteriorly reduced. Collum moderately wide with convex lower margin and very weak or no pseudocolliculum.
Description (n = 28) Elongate, thin otoliths of moderate size for a species of Palimphemus, reaching about 8 mm in length. OL:OH = 2.2–2.7, increasing with size; OL:OT = 6–8, increasing with size. Dorsal rim anteriorly rounded, medio-dorsal part nearly horizontal, straight to slightly depressed and with broad postdorsal angle close to posterior tip, often slightly undulating but sometimes smooth; ventral rim slightly curving, deepest slightly before the middle, slightly undulating or smooth, often with indentation close to its middle in small specimens. Anterior tip slightly pointed with median projection or rounded, then with dorsal accentuation; posterior tip tapering and short, rarely pointed, mostly rounded, particularly in small specimens.
Inner face flat to slightly bent along horizontal axis. Sulcus long, moderately wide, median to slightly supramedian and slightly bent with deepest point at collum. CaL:OsL = 1.1–1.6, increasing with size. Ostial colliculum terminating rather distant from anterior tip of otolith; caudal colliculum reaching closer to posterior tip of otolith, particularly in large specimens. Consequently, ratio CCL:OCL increasing from about 1.3 to 2.2 with ontogeny. Collum moderately wide, with convex lower margin, with weak pseudocolliculum or pseudocolliculum missing. Dorsal field with narrow, indistinct depression above mid-section of sulcus and with common marginal furrows positioned perpendicular to dorsal rim of otolith. Ventral furrow usually well developed, shallow, running at some distance from ventral rim of otolith, particularly at its middle section. Fewer radial furrows on ventral rim than dorsal rim. Outer face slightly convex to slightly concave, mostly smooth except marginal regions with short radial furrows of variable intensity and number.
Variability and ontogeny Otoliths of P. minusculoides show a moderate variability mainly expressed in the intensity of ornamentation along the margins of the inner and outer faces, the expression of the anterior and posterior tips and some proportions, for instance relating to colliculi size. However, the remarkable morphological plasticity is associated with an intense ontogenetic allometric growth. The ontogenetic alterations seem to take place mostly at sizes between 3 and 4.5 mm in length and concern the following aspects (juveniles first, adults second, n = 15 and 13): OL:OH = 2.2–2.45 in juveniles vs 2.4–2.7 in adults; OL:OT = 6–7 vs 7–8; CaL:OsL = 1.1–1.3 vs 1.3–1.6; CCL:OSL = 1.3–1.6 vs 1.6–2.2; inner face flat vs flat to slightly convex; posterior tip blunt vs rounded or slightly pointed; ventral rim smooth except common indentation at its middle vs crenulated and regularly curved without indentation.
Discussion Considering its high degree of ontogenetic variation, it is not surprising that P. minusculoides was described as two different species by Schubert (1906, 1912) based on otoliths of different sizes, and that the subsequent literature contains many problematical citations referring to Gadus minusculus or Gadus minusculoides (and its variations), often based on very small otoliths less than 2 mm in size. Such small otoliths cannot be identified reliably and hence are omitted from our list of synonyms.
Nolf (1981) reviewed the type specimens of Otolithus (Gadidarum) minusculus Schubert, 1906. He stated that the five syntypes of O. (G.) minusculus represent juveniles of different gadid species, and selected the specimen of Schubert’s fig. 52 as lectotype (GBW 1906/01/46e according to Schultz 2013). This specimen, and probably also the one figured by Schubert on his fig. 51, likely represent juvenile specimens of P. anceps Kner, 1862. It is therefore clear that O. (G.) minusculus is not a valid species, which is consistent with the view of Nolf (1981), who rejected this species.
The second species reported by Schubert (1912), and also rejected by Nolf (1981) is Gadus minusculoides Schubert, 1912. Gadus minusculoides was based on a single, somewhat eroded large otolith of about 7 mm length. This specimen clearly shows the diagnostic characters of the fossil species from Mangyshlak presented here and is consequently re-validated.
The abundance and range of sizes of the specimens from Mangyshlak allow P. minusculoides to be redefined and for it to be distinguished from P. anceps Kner, 1862 (see Schwarzhans 2014a). Palimphemus minusculoides differs from P. anceps in being thinner (OL:OT = 6–8 vs 4–5), presenting a nearly flat inner face (vs distinctly convex), a slightly pointed anterior tip (vs rounded), a nearly horizontal mediodorsal rim (vs inclined backwards), a far backward positioned postdorsal angle (vs an indistinct angle located about 2/3 from anterior tip of otolith) and a wider collum and anteriorly reduced ostial colliculum.
Records of P. minusculoides are thus far confined to the Paratethys, where it first occurs in the late Badenian/Konkian and ranges at least until the early Sarmatian s.l. It may represent an endemic species or lineage of the Central and Eastern Paratethys. Palimphemus anceps, on the other hand, had a wide geographic distribution because it had been reported from the late Oligocene to the early Pliocene of Europe. This species apparently became extinct in the Paratethys during the early Serravallian, with a brief overlap with P. minusculoides during that time in the Central Paratethys (see discussion above).
Carnevale et al. (2006) have figured a small otolith (2.5 mm in length) found in situ in a skeleton described as Micromesistius sp. in the lower Sarmatian s.l. (Volhynian, late Serravallian) of the northern Caucasus. Based on the drawing given for this otolith, it resembles P. minusculoides but displays a narrower collum, which is not ventrally widened. This specimen may represent a hitherto unrecognised small species of Palimphemus (see Schwarzhans 2014a for details), and we have placed this record only tentatively in the list of synonymies of P. minusculoides.
Order Mugiliformes Regan, 1909
Family Mugilidae Risso, 1827
Chelon? sp.
Figure 5-1, -2

1–2 Chelon? sp. 1: NMNH 2532/020. 2: NMNH 2532/021, ventral view (2B). Figure 1 is mirrored. 3. Atherina gidjakensis (Pobedina, 1956), ESMGS, Simeonovo B-7, Bulgaria, lower Chersonian. 4, 5, 7, 8 Atherina gidjakensis (Pobedina, 1956). 4: NMNH 2532/030, ventral view (4B). 5: NMNH 2532/031, ventral view (5B). 7: NMNH 2532/034. 8: NMNH 2532/033. Figures 5, 7 are mirrored. 6 Atherina aff. gidjakensis (Pobedina, 1956), NMNH 2532/032. Figure is mirrored
Material 6 specimens (NMNH 2532/020, 021), Karagaily, Level 10.
Remarks Juvenile otoliths that resemble otoliths of the recent Chelon labrosus (Risso, 1826) in the presence of a prominent bulge of the anterior dorsal rim, but are considerably more compressed than recent Chelon otoliths (see Chaine 1938, plate 25, as “Mugil chelo Cuv.”). The compressed shape differentiates these otoliths from the two previously described fossil species of Chelon, i.e. C. gibbosus Reichenbacher, 1992 (in Reichenbacher and Weidmann 1992) from the late Oligocene and C. reichenbacherae Schwarzhans and Wienrich, 2009 from the early to middle Miocene of Western Europe, respectively. Mugil finitimus Switchenska 1973 is a skeleton-based representative of the Mugilidae from the Konkian of Mangyshlak (Bannikov, 2010).
Order Atheriniformes Rosen, 1964
Family Atherinidae Risso, 1827
Genus Atherina Linnaeus, 1758
Type species. Atherina hepsetus Linnaeus, 1758, type by monotypy, Mediterranean Sea; Recent.
Atherina gidjakensis (Pobedina, 1956)
Figure 5-4, -5, -7, -8
1956 Ot. (Clupea) gidjakensis; Pobedina 1956: pl. 27, fig. 4.
1968 Rhombus karaganensis; Suzin 1968 (in Zhizhchenko): pl. 18, fig. 15; [name not available: ICZN article 13.1.1]
? 1981b Atherina austriaca longa; Strashimirov 1981: pl. 2, figs. 11, 12.
1992 genus Atherinidarum sp.; Brzobohaty 1992: pl. 1, fig. 1.
1993 Atherina kalinoraensis; Rückert-Ülkümen and Kaya 1993: pl. 1, fig. 14, ?pl. 2, figs. 4, 7 (eroded spec.), pl. 3, fig. 11.
1996 Atherina kalinoraensis Rückert-Ülkümen, 1993; Rückert-Ülkümen 1996: pl. 3, figs. 1–3, 5, 6 (non fig. 4).
? 2006 Myctophum cf. splendidum (Prochazka, 1893); Djafarova 2006: pl. 4, figs. 5?, 6.
2006 Serranus noetlingi (Koken, 1891); Djafarova 2006: pl. 13, fig. 1.
? 2006 Trachinus mutabilis Koken 1891; Djafarova 2006: pl. 23, fig. 10 (non fig. 9).
? 2006 Otolithus (inc. sed.) albitus; Djafarova 2006: pl. 28, figs. 1–4 (non fig. 7).
Material 6 specimens (NMNH 2532/030, 031, 033–035), Karagaily, Level 10 and 1 specimen from Karagaily, Level 9. An additional specimen (NMNH 2532/032) shown in Fig. 5-6 is only tentatively assigned.
Description (n = 6) Small, thin and rather elongate otoliths up to 1.8 mm in length. OL:OH = 1.45–1.6; OL:OT = 5–6. Ventral rim smooth and slightly bent, somewhat asymmetrical with the deepest curvature in the posterior part; dorsal rim with a high, broadly undulating bulge, but without clearly defined angles. Rostrum moderately long and pointed; no or feeble excisura and antirostrum. Posterior tip mostly rounded, sometimes blunt or tapering.
Inner face slightly convex with narrow, slightly supramedian and moderately deep sulcus. Cauda narrow, nearly straight, just very slightly bent at termination, which is at moderate distance from posterior tip of otolith. Ostium only slightly wider than cauda and short; CaL:OsL = 1.85–2.3. Dorsal depression long, ventrally marked by well-developed crista superior, dorsal margin indistinct; ventral furrow mostly indistinct, at moderate distance from ventral rim of otolith. Outer face slightly concave, smooth.
Discussion Atherina gidjakensis differs from the widespread A. austriaca from the Langhian and Serravallian of the Tethys and the Badenian of the Central Paratethys in the more elongate shape (OL:OH = 1.45–1.6 vs 1.3–1.4), the less curved ventral rim, the high dorsal bulge, the longer rostrum and the lack of a finely crenulated dorsal rim. Atherina mutila Rückert-Ülkümen, 1996 from the Sarmation to Pontian of the Thrace Basin in Turkey differs in the even more elongate shape (OL:OH = 1.8–1.9 vs 1.45–1.6) and the long cauda reaching close to the posterior tip of the otolith.
Atherina gidjakensis was first described by Pobedina (1956) as Clupea gidjakensis from the Maeotian of Azerbaijan based on a single small specimen about 0.7 mm in length. Her drawing shows a typical otolith of the genus Atherina characterised by a rather elongate shape (OL:OH about 1.6) and a gently curving dorsal rim. Some of the studied specimens from Mangyshlak are similar in shape and proportions (Fig. 5-4, -8), while others display a higher dorsal rim (Fig. 5-5, -7). However, the dorsal rim curvature apparently represents intraspecific variation.
Suzin 1968 (in Zhizhchenko) figured a similar otolith as Rhombus karaganensis (species name not available according to ICZN article 13.1.1) from the Karaganian of the northern Caucasus and the Kerch Peninsula. Strashimirov (1981b) described Atherina austriaca longa from the Konkian of Bulgaria based on a single somewhat eroded otolith with a slightly indented posterior tip, which is similar to one of our specimens (Fig. 5-4). A specimen recently retrieved from undescribed material of the late B. Strashimirov from the upper Sarmatian s.l. (Fig. 5-3) corresponds more to the forms with a rounded posterior rim. Atherina kalinoraensis Rückert-Ülkümen and Kaya, 1993 from the Sarmatian to Pannonian of the Thrace Basin of the south-eastern Paratethys represents the same species and is particularly well documented in Rückert-Ülkümen (1996). Furthermore, Djafarova (2006) described otoliths that may perhaps represent A. gidjakensis under several identities from the Karaganian to Maeotian of Azerbaijan.
Atherina gidjakensis thus appears to be a common and widespread species throughout the Karaganian to Maeotian of the Eastern Paratethys including the Thrace Basin of Turkey and to be sporadic in the uppermost Pannonian of the Vienna Basin. It has been described under numerous different names, and there could be more in the works of Suzin and Djafarova, which remain unresolved unless their original material becomes located and reviewed. Bannikov (2010) lists four skeleton-based species of Atherina from the Karaganian to the late Sarmatian in the Eastern Paratethys (no otoliths in situ known).
Atherina aff. gidjakensis (Pobedina, 1956)
Figure 5-6
Discussion A singular specimen from Karagaily differs from the others in the overall shape and the deepened ostium and is therefore referred to as A. aff. gidjakensis.
Order Scorpaeniformes Garman, 1899
Suborder Scorpaenoidei Garman, 1899
Family Scorpaenidae Risso, 1827
Genus Pontinus Poey, 1860
Type species. Pontinus castor Poey 1860, type by monotypy, Havanna, Cuba; Recent.
Pontinus? aff. obrotchishtensis (Strashimirov, 1981)
Figure 6-1

1 Pontinus? aff. obrotchishtensis Strashimirov, 1981, NMNH 2532/092, ventral view (1B). 2 Pontinus? obrotchishtensis Strashimirov, 1981, ESMGS, Goren Bliznak C-2, 158 m, Bulgaria, probably Tarchanian. Figure is mirrored. 3 Morone? ionkoi (Bannikov, 1993), otolith in situ, PIN, Karpov Yar, Moldavia, lower Sarmatian. 4 Morone sp., NMNH 2532/043, ventral view (4B). Figure is mirrored. 5–9 Morone? bannikovi n.sp. Bratishko, Schwarzhans and Reichenbacher. 5: Paratype, NMNH 2532/045, ventral view (5B). 6: Paratype, NMNH 2532/046, ventral view (6B). 7: Holotype, NMNH 2532/048. 8: Paratype, NMNH 2532/049. 9: Paratype, NMNH 2532/047. 10 Trachurus sp., NMNH 2532/053, ventral view (10B). 11–12 Pagellus sp. 11: NMNH 2532/036, ventral view (11B), 12: NMNH 2532/037, ventral view (12B). Figure 12 is mirrored. 13–16 Centracanthus pobedinae n.sp. Bratishko, Schwarzhans and Reichenbacher. 13: Paratype, NMNH 2532/038, ventral view (13B). 14: Paratype, NMNH 2532/040, ventral view (14B). 15: Holotype, NMNH 2532/039, ventral view (15B). 16: Paratype, NMNH 2532/041, ventral view (16B). Figure 13 is mirrored
?1981 Perca obrotchishtensis; Strashimirov 1981b: pl. 2, figs. 3, 4.
?2006 Centropristis integer Schubert, 1906; Djafarova 2006: pl. 13, fig. 5.
Material 1 specimen (NMNH 2532/092), Karagaily, Level 10.
Description A single well-preserved otolith about 1.7 mm in length. OL:OH = 1.7; OL:OT about 6. Ventral rim moderately deep, with rounded medioventral angle slightly in front of middle of otolith; dorsal rim almost symmetrically formed to ventral rim, but with sharper mediodorsal angle and feebly rounded predorsal and angular postdorsal angles. Rostrum and posterior tip almost symmetrically developed as pointed projections, the former slightly below median axis of otolith, the latter slightly above. Sulcus with thin forward projection at anterior margin.
Inner face slightly convex with moderately deep and nearly medianly positioned sulcus. Ostium and cauda of about equal length, but ostium about twice as wide as cauda. Cauda almost straight, very slightly inclined, terminating at considerable distance from posterior tip of otolith. Dorsal depression short, but well marked by crista superior toward sulcus; ventral furrow feeble, best visible only along middle section of ventral rim. Outer face flat, almost smooth.
Discussion Our knowledge of Recent otoliths of the family Scorpaenidae is still limited and therefore this species is only tentatively allocated to Pontinus, particularly because of the small size of the fossil species (see Nolf 1985, Lombarte et al. 2006 for figures of Recent otoliths). The single otolith bears some resemblance to the specimens figured by Strashimirov (1981b) and Djafarova (2006) except for its considerably shorter rostrum and a forward expansion of the sulcus opening, which is lacking in Strashimirov’s and Djafarova’s specimens. A very small specimen recently received from Strashimirov’s collection of unpublished otoliths from the well Gren Bliznak C-2 (158 m) in NW Bulgaria and possibly of Tarchanian age seems to represent the same species and is figured here for comparison (Fig. 6-2).
Order Perciformes Bleeker, 1859
Suborder Percoidei Bleeker, 1859
Family Moronidae Jordan and Evermann, 1896
Genus Morone Mitchill, 1814
Type species. Morone rufa Mitchill 1814 (synonym of Perca americana Gmelin, 1789), type by subsequent designation, New York, U.S.A., Atlantic; Recent.
Morone sp.
Figure 6-4
Material 1 specimen (NMNH 2532/043), Karagaily, Level 10.
Description A single otolith 3.1 mm in length. OL:OH = 1.77; OL:OT about 5. Ventral rim moderately deep, regularly curved, smooth; dorsal rim shallower, slightly undulating, with rounded mediodorsal angle and marked postdorsal angle located far backward on dorsal rim. Rostrum short, its tip somewhat abraded by erosion; posterior tip blunt. No excisura or antirostrum.
Inner face markedly convex with moderately deep, slightly supramedian positioned sulcus. Ostium short, about twice as wide as cauda, particularly dorsally widened. CaL:OsL about 1.5. Cauda long, narrow, straight except for a slight downward flexure at its tip, slightly inclined, terminating at moderate distance from posterior tip of otolith. Dorsal depression narrow, well marked by crista superior toward sulcus; ventral furrow indistinct. Outer face slightly concave, smooth.
Discussion Morone moravica Weiler, 1966 (see Reichenbacher 1993, for comprehensive figures) has been recorded from the Eggenburgian and Ottnangian (Burdigalian) of the Central Paratethys. The single specimen described here from the Konkian (early Serravallian) of Mangyshlak differs from this species because of its compressed shape (vs. elliptical in M. moravica), blunt posterior rim (vs. pointed) and narrow, almost straight cauda (vs. broad, terminally bent).
Morone? bannikovi n.sp. Bratishko, Schwarzhans and Reichenbacher
Figure 6-5 to -9
Etymology In honor of A. Bannikov in recognition of his important contributions to the knowledge of fossil fishes from the Eastern Paratethys.
Type material Holotype: NMNH 2532/046 (Fig. 6-6), Paratypes: 4 specimens NMNH 2532/045, 047–049 (Figs. 6-5, -7 to -9). Additional specimens: 5 otoliths from Karagaily Level 10 and 1 specimen from Karagaily Level 9.
Type locality Karagaily, Mangyshlak, Kazakhstan.
Age Konkian, early Serravallian, middle Miocene.
Diagnosis Moderately elongate otoliths; OL:OH = 1.55–1.7. Ventral rim regularly curved; dorsal rim irregularly undulating. Rostrum long, 25–30 % OL, massive, with horizontal dorsal margin; excisura rectangular.
Description (n = 8) Moderately elongate and thin otoliths up to about 3.5 mm in length. OL:OT = 6–7. Ventral rim shallow, regularly curved, smooth; dorsal rim short, with broad mediodorsal angle and depressed, weak postdorsal angle located above posterior part of cauda. Rostrum long, its tip broadly rounded, its dorsal margin straight, with broad, nearly rectangular excisura but no anteriorly projecting antirostrum; posterior tip broadly rounded.
Inner face slightly convex with moderately deep, slightly supramedian positioned sulcus. Ostium short, spacious, more than twice as wide as cauda, its dorsal margin at equal level with dorsal margin of cauda, open to anterior rim for more than 2/3 of its length. CaL:OsL about 1.5. Cauda long, narrow, straight except for a slight downward flexure at its end, terminating relatively close to posterior tip of otolith. Dorsal depression wide, well marked by crista superior; no ventral furrow. Outer face flat to slightly convex, smooth.
Discussion These otoliths display some similarity to “Morone ionkoi Bannikov 1993”, of which two otoliths in situ are known (one figured in Bannikov 1993, the other illustrated here in Fig. 6-3). However, the ventral rim of the specimens from Karagaily is much deeper, which results in a different OL/OH value (1.55–1.7 vs. 1.9). Bannikov placed this species in the genus Morone and the meristics of his description support allocation to the Moronidae. The relatively long pectoral fin reaching near to the base of the second dorsal and containing only 10 rays (13–19 in Recent Morone species) may, however, indicate a different genus from Morone, possibly a fossil genus.
Similar fossil otolith-based species have been tentatively placed in the acropomatid genus Parascombrops, i.e. P.? postgeron Schwarzhans, 2010 from the Burdigalian of the North Sea Basin and P.? geron (Koken, 1891) from the late Oligocene of the North Sea Basin. They both differ by a more depressed postdorsal rim and a deeper ventral rim.
Family Carangidae Rafinesque, 1815
Genus Trachurus Rafinesque, 1810
Type species. Trachurus saurus Rafinesque, 1810 (unnecessary new name for Scomber trachurus Linnaeus 1758), type by subsequent designation, Mediterranean Sea; Recent.
Trachurus sp.
Figure 6-10
Material 1 juvenile specimen NMNH 2532/053, Karagaily, Level 10.
Remarks A well-preserved juvenile specimen about 2.5 mm in length considered to be diagnostically immature, exhibiting a low postdorsal rim and resembling T. elegans Jonet, 1973 in the elongate shape.
Family Sparidae Rafinesque, 1810
Genus Pagellus Valenciennes, 1830
Type species. Sparus erythrinus Linnaeus 1758, type by subsequent designation, Mediterranean Sea; Recent.
Pagellus sp.
Figure 6-11, -12
Material 3 specimens NMNH 2532/036, 037, 042, Karagaily, Level 10.
Remarks None of the specimens available are either diagnostically mature or complete. The generic allocation is based on correlation with the Recent species of Pagellus such as P. bogaraveo (Brünnich, 1768) and P. erythrinus (L. 1758), figured in Tuset et al. (2008) and Nolf et al. (2009). Bannikov (2010) lists three skeleton-based species of the related genus Sparus from the Sarmatian and Maeotian of the Eastern Paratethys (no otoliths in situ known). An otolith specimen from the Badenien of the Central Paratethys has been recorded as P. erythrinus by Brzobohaty et al. (2007).
Family Centracanthidae Gill, 1891
Genus Centracanthus Rafinesque, 1810
Type species. Centracanthus cirrus Rafinesque 1810, type by monotypy, Sicily, Mediterranean Sea; Recent.
Centracanthus pobedinae n.sp. Bratishko, Schwarzhans and Reichenbacher
Figure 6-13 to -16
Etymology In honor of the late V. Pobedina, who was one of the first researchers to investigate fossil otoliths from the Caspian Basin.
Type material Holotype: NMNH 2532/039 (Fig. 6-15). Paratypes: 3 specimens NMNH 2532/038, 041, 070 (Fig. 6-13, -14, -16). Additional specimens: 2 otoliths NMNH 2532/075, 076, Karagaily, Level 10
Type locality Karagaily, Mangyshlak, Kazakhstan.
Age Konkian, early Serravallian, middle Miocene.
Diagnosis Otoliths moderately elongate, OL:OH = 1.6–1.75. Dorsal rim low with broad mediodorsal and moderately developed postdorsal angles. Rostrum short; no or minute excisura and antirostrum. Ostium moderately wide with parallel dorsal and ventral margins. Cauda slightly bent, with tapering tip.
Description (n = 4) Moderately elongate otoliths up to 5 mm in length. OL:OH = 1.6–1.75; OL:OT = 4.5–6. Ventral rim moderately deep and regularly curved, smooth; dorsal rim low, slightly crenulated, with broad mediodorsal angle and moderately developed postdorsal angle positioned at or behind tip of cauda. Rostrum moderately long, with broad tip; no or minute excisura and antirostrum; posterior tip oblique, blunt.
Inner face convex with slightly supramedian positioned sulcus. Ostium comparatively long, narrow, its dorsal and ventral margins parallel, about twice as wide as cauda. CaL:OsL = 1.4–1.5. Cauda long, narrow, straight except for slight downward flexure of its tapering tip, terminating relatively close to posterior-ventral rim of otolith. Dorsal field with few short radial furrows, dorsal depression wide, long, well marked by crista superior; no ventral furrow. Outer face flat to slightly concave, rather smooth.
Comparison The combination of the low dorsal rim with the broad mediodorsal and the moderate postdorsal angle as well as the form of the ostium and the only slightly bent, tapering cauda distinguish this species from centracanthid and sparid species otherwise known from the European Oligocene and Miocene. The late Oligocene C. varians (Koken, 1884) (figured in Nolf 2013: Pl. 265) differs from the new species because of its deeper ventral rim and shorter cauda. Miocene species of Centracanthus are not yet known because Centracanthus sp., described from the Miocene of the North Sea in Schwarzhans (2010, pl. 93, figs. 7–8), has been re-interpreted as Spondyliosoma in Schwarzhans (2014b). Otoliths of Centracanthus cirrus Rafinesque, 1810, the only Recent species of the genus, show a similar shape of the sulcus including the ostium with nearly parallel margins but exhibit a more pronounced postdorsal region and a deeper ventral rim (see Nolf 2013; Plate 264 for figures). Bannikov (2010) reports a single skeleton-based centracanthid from the early Sarmatian of the Eastern Paratethys belonging to the fossil genus Nasclavcea Bannikov, 2006.
Family Sciaenidae Cuvier, 1828
Remark. See Béarez and Schwarzhans (2013) for terminology and details on otolith measurements.
Genus Pogonias Lacepède, 1801
Type species. Pogonias fasciatus Lacepède 1801 (synonym of Labrus chromis Linnaeus, 1766), type by monotypy, location not stated; Recent.
Pogonias? sp.
Figure 7-1

1 Pogonias? sp., NMNH 2532/052, ventral view (1B). Figure is mirrored. 3–7 Genyonemus? karagiensis n.sp. Bratishko, Schwarzhans and Reichenbacher. 3: Paratype, NMNH 2532/055, ventral view (3B). 4: Paratype, NMNH 2532/056, ventral view (4B). 5: Holotype, NMNH 2532/058, ventral view (5B). 6: Paratype, NMNH 2532/059. 7: Paratype, NMNH 2532/060. Figure 3 is mirrored. 2, 8 Genyonemus? aff. karagiensis n.sp. Bratishko, Schwarzhans and Reichenbacher. 2: NMNH 2532/054, ventral view (2B). 8: NMNH 2532/057. 9–14 Trewasciaena suzini n.sp. Bratishko, Schwarzhans and Reichenbacher. 9: Paratype, NMNH 2532/064, ventral view (9B). 10: Paratype, NMNH 2532/066. 11: Paratype, NMNH 2532/065, ventral view (11B). 12: Paratype, NMNH 2532/061, ventral view (12B). 13: Paratype, NMNH 2532/062, ventral view (13B). 14: Holotype, NMNH 2532/063, ventral view (14B). Figures 9, 13, 14 are mirrored
Material 1 specimen NMNH 2532/52, Karagaily, Level 10.
Discussion Pogonias chromis (Linnaeus, 1766), the sole Recent species of the genus, lives along the subtropical and temperate shores of North and South America. Pogonias otoliths are recognised by their compact shape, flat outer face, moderately sized ostium and the rather short cauda (Schwarzhans 1993). In the fossil record, the genus has been identified from the Miocene and Pliocene of NE America (Müller 1999) and also from the late Miocene of the North Sea Basin (Schwarzhans, 2010). Somewhat similar compressed sciaenid otoliths have been reported from the middle Miocene of the Central Paratethys by Schubert (1902) as “Otolithus (Sciaenidarum) kittli” and by Weinfurter (1952) as “Otolithus (Sciaenidarum) styriacus”. These two taxa might represent a single species, but its validity is, however, currently considered doubtful due to the poor preservation of the otoliths (Nolf and Brzobohaty 2009; Schultz 2013). The fragmentary specimen from Mangyshlak differs in the straight dorsal rim and the less broad posterior rim. It rather resembles the specimens from the North Sea Basin (see Schwarzhans 2010). Due to its incomplete preservation, its generic attribution remains tentative.
Genus Genyonemus Gill, 1861
Type species. Leiostomus lineatus Ayres, 1855, type by original designation, San Francisco Bay, California; Recent.
Genyonemus? karagiensis n.sp. Bratishko, Schwarzhans and Reichenbacher
Figure 7-3 to -7
Etymology Named after the Karagie Depression, Mangyshlak, Kazakhstan, where the otoliths have been collected.
Type material Holotype: NMNH 2532/058 (Fig. 7-5). Paratypes: 5 specimens NMNH 2532/055, 056, 059, 060, 067, (Fig. 7-3, -4, -6, -7). Additional specimens: 7 otoliths from Karagaily, Level 10 and 1 specimen from Level 9.
Type locality Karagaily, Mangyshlak, Kazakhstan.
Age Konkian, early Serravallian, middle Miocene.
Diagnosis Otoliths moderately large, reaching up to 6 mm in length, very elongate, OL:OH = 2.1–2.4. Dorsal and ventral rims nearly straight; posterior rim rounded. Ostium long, moderately wide, with weak postostial lobe. Horizontal part of cauda long, expressing a high caudal curvature index of 2.4–2.6. Outer face flat or with very shallow postcentral umbo.
Description (n = 6) Very elongate, moderately thick otoliths up to 6 mm in length. OL:OH = 2.1–2.4; OL:OT = 4.8–5.5. Dorsal and ventral rims nearly straight, ventral rim deepest at about level of collum, smooth, dorsal rim highest at level of collum or mid-dorsally, smooth, sometimes undulating. Anterior tip broadly rounded; posterior tip rounded, usually slightly projecting dorsally.
Inner face only slightly convex along horizontal axis, not bent in perpendicular direction. Sulcus occupying large portion of inner face. Ostium long, wide, flat, with horizontal ventral and slightly anteriorly inclined dorsal margins, with feeble postostial lobe. Cauda long, narrow, somewhat deepened, anterior part long, horizontal, straight, downturned posteriorly at almost 90° angle, terminating close to posterior-ventral rim of otolith. CaL:OsL = 0.9–1.0; caudal curvature index = 2.4–2.6. Dorsal field very narrow, sometimes with indication of dorsal depression; ventral field smooth. Outer face nearly flat to slightly convex, smooth, sometimes with broad, very shallow postcentral umbo.
Comparison Genyonemus? karagiensis resembles the Recent G. lineatus (Ayres, 1855) in the moderately expanded ostium with a very weak postostial lobe, the long horizontal stretch of the cauda expressed in a high caudal curvature index, coupled with a short length for the downturned portion of the cauda, the rather smooth inner face bent only in the horizontal direction and finally the broad, flat umbo on the outer face (see Schwarzhans 1993: figs. 153–155). It differs from the Recent species in the more slender, elongate shape and the nearly straight dorsal and ventral rims. Similar otoliths have been described by Müller (1999) from the middle Miocene of NE America as G. pertenuis Müller, 1999 and Cynoscion fitchi Müller, 1999. Genyonemus pertenuis is thinner and also mostly more elongate than G.? karagiensis (OL:OT = 7.7–8.2 vs 4.8–5.5 and OL:OH = 2.25–2.6 vs 2.1–2.4). Cynoscion fitchi likewise is thinner than G? karagiensis (OL:OT about 6.0 vs 4.8–5.5) and is characterised by a very low dorsal rim, which is almost concave and a more regularly curved ventral rim.
In the Recent, Genyonemus is monospecific and restricted to the waters off California. The fossil record has already extended its distribution to the North American east coast. The record from the Eastern Paratethys extends the palaeogeographic distribution even further, but in the light of the large distance the generic allocation remains tentative. It is also possible that G.? karagiensis represents an extinct sciaenid genus. In this respect it is worth mentioning that a fossil skeleton-based sciaenid genus—Caucasciaena Bannikov, Carnevale and Landini, 2009—has been described from the early Miocene of the Eastern Paratethys (no otoliths in situ known).
Genyonemus? aff. karagiensis Bratishko, Schwarzhans and Reichenbacher (this study)
Figure 7-2, -8
Description Two specimens NMNH 2532/054, 057 (Fig. 7-2, -8) show a distance between the posterior end of the ostium to the downturned part of the cauda which is shorter than seen in Genyonemus? karagiensis (CaL:OsL = 2.1–2.2 vs 2.4–2.6). In addition they are slightly more compressed (OL:OH = 2.0–2.2 vs 2.1–2.4).
Genus Trewasciaena Schwarzhans, 1993
Type species. Sciaena moguntina Weiler, 1942, type by original designation, Alsheim near Mainz, Germany; otolith-based fossil genus, early Miocene.
Trewasciaena suzini n.sp. Bratishko, Schwarzhans and Reichenbacher
Figure 7-9–-14
Etymology In honor of the late A. Suzin, one of the first researchers of study fossil otoliths from the Eastern Paratethys.
Type material Holotype: NMNH 2532/063 (Fig. 7-14). Paratypes: 5 specimens NMNH 2532/061, 062, 064–066 (Fig. 7-9–-13). Additional specimens: 2 otoliths from Karagaily, Level 10.
Type locality Karagaily, Mangyshlak, Kazakhstan.
Age Konkian, early Serravallian, middle Miocene.
Diagnosis Otoliths moderately large, reaching up to 5 mm in length, elongate, OL:OH = 1.75–1.95. Dorsal rim shallow, nearly straight and horizontal, ventral rim slightly convex; posterior rim with dorsal projection. Ostium long, moderately wide, with moderate postostial lobe. Caudal curvature index high = 1.35–1.85. Outer face slightly thickened postcentrally, but without distinct umbo.
Description (n = 6) Elongate, moderately thin otoliths up to 5 mm in length. OL:OH = 1.75–1.95; OL:OT = 4.8–5.5. Dorsal rim shallow, nearly straight and horizontal without angles, ventral rim somewhat deeper, deepest anterior of middle below posterior part of ostium, both smooth. Anterior tip broadly rounded; posterior tip with pronounced dorsal projection.
Inner face only slightly convex along horizontal axis, not bent in perpendicular direction. Sulcus occupying large portion of inner face. Ostium long, wide, flat, with horizontal ventral and dorsal margins, dorsal margin slightly concave at its middle, with moderately deep postostial lobe. Cauda long, narrow, somewhat deepened, straight anteriorly, downturned posteriorly at almost 90° angle, terminating close to posterior-ventral rim of otolith. CaL:OsL = 0.95–1.05; caudal curvature index = 1.35–1.85. Dorsal field with indistinct depression, ventral fields smooth. Outer face anteriorly concave, posteriorly slightly convex, smooth.
Comparison Trewasciaena suzini resembles T. kokeni (Schubert, 1902) (see Schwarzhans 1993 for figures), which is known from the Pannonian of the Paratethys and differs mainly in being less elongate (OL:OH = 1.75–1.95 vs 1.6–1.75 for T. kokeni). Trewasciaena suzini and T. kokeni appear to represent parts of an endemic lineage in the Eastern Paratethys leading further to T. moguntiniformis (Pana, 1977) in the Dacian and an undescribed species in the late Pliocene and early Pleistocene identified as Sciaena aff. excissus Schubert, 1902 in Djafaraova (2006), before becoming extinct. Trewasciaena suzini differs from the contemporaneous Genyonemus? karagiensis in the more compressed shape, the lower caudal curvature index and the shape of the posterior tip.
Family Mullidae Rafinesque, 1815
Genus Mullus Linnaeus, 1758
Type species. Mullus barbatus Linaeus, 1758, type by subsequent designation, Mediterranean Sea; Recent.
Mullus bifurcatus (Strashimirov, 1972)
Figure 8-1 to -7

1–7 Mullus bifurcatus (Strashimirov, 1972). 1: NMNH 2532/022, ventral view (1B). 2: NMNH 2532/023, ventral view (2B). 3: NMNH 2532/024, ventral view (3B). 4: NMNH 2532/026. 5: NMNH 2532/025. 6: NMNH 2532/028. 7: NMNH 2532/068. Figures 1, 3, 4, 5 are mirrored. 8. Mullus bifurcatus (Strashimirov, 1972), HT ESMGS XI9 Goren Bliznak C-55, 180.3 m. Tarchanian. 9–10 Polydactylus cf. gaemersi Steurbaut, 1984. 9: NMNH 2532/108. 10: NMNH 2532/069, ventral view (9B). 11 Trachinus sp., NMNH 2532/086, ventral view (11B). Figure is mirrored
1956 Otolithus (Sparidarum) sp.; Pobedina 1956: pl 8, fig. 8.
1966 Mullus aff. fuscatus Rafinesque, 1810; Smigielska 1966: pl. 18, fig. 1.
1966 Trigla sp.; Smigielska 1966: pl. 19, fig. 5
1972 Otolithus (inc. sed.) bifurcatus; Strashimirov 1972: pl. 2, figs. 9, 10
Material 11 specimens NMNH 2532/022 –029, 068, Karagaily, Level 10
Remark The specimens described by Strashimirov (1972) have been recorded from the Tarkhanian (lowermost middle Miocene) of NE Bulgaria. A detailed diagnosis and re-description of this species is given here because Strashimirov’s documentation is not optimal for species recognition. Mullus bifurcatus is the only species described by Strashimirov for which the holotype could be retrieved (see Fig. 8-8).
Diagnosis Small, moderately elongate otoliths, OL:OH = 1.5. Dorsal rim anteriorly depressed with backward-positioned postdorsal angle. Ventral rim shallow. Cauda with long horizontal stretch.
Description (n = 7) Moderately elongate, thin otoliths up to 1.5 mm in length. OL:OT = 6–7. Ventral rim shallow, regularly curved, slightly to moderately strongly undulating; dorsal rim anteriorly depressed, inclined, posteriorly with distinct but rather low postdorsal angle, slightly undulating, particularly the postdorsal region, sometimes with separated mediodorsal angle just in front. Rostrum moderately long, with massive tip, length about 15 % of OL; excisura distinct, deep, mostly sharp; antirostrum variable, long, mostly sharp; posterior tip blunt, with obtuse angle at level of caudal tip.
Inner face markedly convex, slightly twisted along horizontal axis. Sulcus rather deep, slightly supramedianly positioned. Ostium short, only slightly (and primarily ventrally) widened, and widening commencing slightly in front of collum. Cauda typical for mullids, with long horizontal section and drop-like widened and slightly downturned posterior section, terminating close to posterior rim of otolith. CaL:OsL = 1.9–2.3; caudal curvature index = 3.0–4.2. Dorsal depression narrow, long, well marked by crista superior toward sulcus; ventral furrow short, positioned approximately halfway between cauda and ventral rim of otolith. Outer face concave, with some radial furrows.
Comparison These otoliths seem to represent a rather small species since all available specimens are small, but they show good diagnostic features. Mullus bifurcatus is not as elongate as M. elongatus Steurbaut, 1984 from the Burdigalian of SW-France (OL:OH = 1.5 vs 1.6–1.7), but it is more elongate than the small, unidentified Mullus specimens occasionally recorded from the Langhian of the Central Paratethys (Radwanska 1992) and the Serravallian of Turkey (Schwarzhans 2014) (OL:OH = 1.5 vs 1.4) or the two Recent European species M. barbatus (OL:OH = 1.3–1.4) and M. surmuletus (OL:OH = 1.3–1.45) (see Lombarte et al. 2006). Both Recent species also show a wider caudal tip and a smaller caudal curvature index below 3.0 (mostly 1.5–2.5). There is a single skeleton-based mullid species recorded by Bannikov (2010) from the early Sarmatian of the Eastern Paratethys (no otoliths in situ known).
Family Polynemidae Rafinesque, 1815
Genus Polydactylus Lacepède, 1803
Type species. Polyadactylus plumierii Lacepède, 1803 (snynonym of Polynemus vriginicus Linnaeus, 1758), type by monotypy, no locality stated; Recent.
Polydactylus cf. gaemersi Steurbaut, 1984
Figure 8-9 to -10
1984 genus Polynemidarum gaemersi; Steurbaut 1984: pl. 30, figs. 8–10.
Material 2 specimen NMNH 2532/069, 108, Karagaily, Level 10.
Discussion The few specimens found in Mangyshlak all lack the rostrum. Nevertheless, they are readily recognised as Polynemidae by the peculiarly shaped, widened and deepened caudal tip, the narrow ostium and the broad mediodorsal region set off anteriorly and posteriorly by a distinct concave stretch of the dorsal rim. The elongate shape resembles recent otoliths of the genus Polydactylus (see Smale et al. 1995; Lombarte et al. 2006 for figures). Likewise, the elongate shape and specific shape of the dorsal rim are in good agreement with P. gaemersi, so far only recorded from the Burdigalian of SW France. Bannikov (2010) recorded a single skeleton-based species of the genus Polydactylus as P. frivolus Bannikov, 1989 from the early Sarmatian of the Eastern Paratethys.
Suborder Trachinoidei Bertin and Arambourg, 1958
Family Trachinidae Rafinesque, 1810
Genus Trachinus Linnaeus, 1758
Type species. Trachinus draco Linnaeus, 1758, type by subsequent designation, Northern Ocean; Recent.
Trachinus sp.
Figure 8-11
Material 1 specimen NMNH 2532/086, Karagaily, Level 10.
Remarks The single specimen about 3.5 mm in length is well preserved but lacks a rostrum, so no specific identification is possible.
Suborder Callionymoidei Berg, 1937
Family Callionymidae Bonaparte, 1831
Remarks Fossil callionymid otoliths are generally rare, but have been recorded since Eocene times. Otoliths of Recent species are poorly studied and hence all fossil species have been assigned to the genus Callionymus, probably mostly due to convenience (C. lerenardi Nolf and Lapierre, 1979 from the middle Eocene, C. schuermanni Schwarzhans, 1973 from the late Eocene, C. pachyotus Gaemers, 1985 from the early Oligocene, C. primus Weiler, 1943 from the middle Miocene of Romania). The exception is the Miocene of the Eastern Paratethys, where callionymid otoliths are more common and species-rich, as documented below. In 2007, Sytchevskaya and Prokofiev described the fossil genus Protonymus with its type species (P. gontsharovae) based on a skeleton with an otolith in situ. They discussed in detail the possible relationships of previously described fossil otolith-based species and concluded that none of them can be placed in Protonymus. Following the new finds from Mangyshalk, a review of Weiler (1943, 1949), Pobedina (1954, 1956), Suzin (1968), Strashimirov (1981) and Djafarova (2006) and a re-examination of the specimens figured in Weiler, we consider it possible that the previously described callionymid species from the Eastern and Central Paratethys can be assigned to Protonymus Sytchevskaya and Prokofiev, 2007. Moreover, we suppose that P. gontsharovae may perhaps be a junior synonym of the species described here as P.? primus. A detailed re-investigation of the otolith of P. gontsharovae, together with a thorough examination of Recent species of Callionymus, was not possible for this study, so we keep the suggested tentative synonymies.
Genus Protonymus Sytchevskaya and Prokofiev, 2007
Type species. Protonymus gontsharovae Sytchevskaya and Prokofiev, 2007, type by monotypy, Taman, Krasnodar district, Russia; middle Miocene, Konkian.
Protonymus? primus (Weiler, 1943)
Figure 9-1, -2

1–2 Protonymus? primus (Weiler, 1943). 1: NMNH 2532/140, ventral view (1B). 2: NMNH 2532/141. 3 Protonymus? primus (Weiler, 1943), holotype, SMF P 2655a, Melicesti, Buglovian. Figure is mirrored. 4 Protonymus? aff. primus (Weiler, 1943), paratype of Callionymus primus, SMF P 2655f, Melicesti, Buglovian. 5–6 Protonymus? aff. primus (Weiler, 1943). 5: NMNH 2532/142. 6: NMNH 2532/143. Figure 5 is mirrored. 7–9 Protonymus? aff. miocenicus (Pobedina, 1954). 7: NMNH 2532/139, 8: NMNH 2532/138, ventral view (8B). 9: NMNH 2532/145. 10–14 Parablennius prokofievi n.sp. Bratishko, Schwarzhans and Reichenbacher. 10: Paratype, NMNH 2532/087, anterior view (10B). 11: Paratype, NMNH 2532/088, anterior view (11B). 12: Holotype, NMNH 2532/089, anterior view (12B). 13: Paratype, NMNH 2532/090, anterior view (13B). 14: Paratype, NMNH 2532/091, anterior view (14B)
1943 Callionymus primus; Weiler 1943: pl. 1, figs. 31–35, non 36.
1949 Callionymus primus Weiler, 1943; Weiler 1949: pl. 4, figs. 32, 34?, 35, non 36.
? 2007 Protonymus gontsharovae; Sytchevskaya and Prokofiev 2007: fig. 2b.
Material 4 specimens NMNH 2532/142, 168, Karagaily, Level 9 and 3 specimens NMNH 2532/140, 141, 144, Karagaily, Level 10.
Remark A detailed diagnosis and re-description of this species is given here because Weiler’s documentation is not optimal for species recognition.
Diagnosis OL:OH = 1.6–1.9. Middorsal angle broad, rounded; ventral rim flat. Rostrum short, not projecting much beyond antirostrum. OL:SuL = 1.45–1.65; OsL:CaL = 1.5–1.6.
Description (n = 3) Small otoliths less than 1.5 mm in length, roughly triangular in outline with flat ventral rim and broadly rounded, rather low mediodorsal angle. OL:OH = 1.6–1.9; OL:OT = 3.5–4. Rostrum close to ventral rim, short, not or only slightly projecting beyond small antirostrum; excisura variable, minute to marked. Posterior tip close to ventral rim, moderately pointed to rounded. All rims smooth.
Inner face almost flat, with typical callionymid upward-oriented sulcus. Ostium and cauda narrow, ostium more deepened than cauda. Ostium about twice as long as cauda, not widened, anteriorly open. Cauda straight, terminating at about 1/3 or more of otolith length from posterior tip of otolith. OL:SuL = 1.45–1.65; OsL:CaL = 1.5–1.6. Dorsal depression distinct, very narrow; ventral furrow moderately developed, close to ventral rim of otolith at central part. Outer face slightly convex, flat at its middle, smooth.
Discussion Weiler’s holotype of P.? primus is refigured here for comparison (Fig. 9-4). Protonymus? primus resembles the figures given for P.? konkensis in Pobedina (1954, 1956) as well as in Djafarova (2006), differing mainly in the smaller OL:OH ratio (1.6–1.9 vs 1.9–2.1) and a straight to slightly concave inclined postdorsal rim. However, the mentioned iconographies are based on the same specimens of P.? konkensis that have been published three times; in our view, these figures are not adequate for diagnostic identification without a review of the type specimens. Until this is done, we consider P.? konkensis to be a nominally valid but doubtful species.
Our otoliths from Karagaily are similar to that figured from an in situ find by Sytchevskaya and Prokofiev (2007), which is about 2 mm long. As a difference from Weiler’s C. primus, the authors mentioned a longer, less inclined sulcus. However, without a re-investigation of P. gontsharovae, it is difficult to work out whether they represent the same species or not. Our specimens are generally smaller, 0.5–1 mm in length, and the smaller ones may represent juvenile specimens, which may not have developed all pertinent diagnostic characters. Furthermore, Sytchevskaya and Prokofiev also report a second callionymid skeleton-based species, Callionymus macrocephalus Gorjanovic-Kramberger, 1882, from the early Sarmatian of the Paratethys, supporting the presence of several callionymid species in the Miocene of the Paratethys.
Protonymus? aff. primus (Weiler, 1943)
Figure 9-5, -6
1943 Callionymus primus; Weiler 1943: pl. 1, fig. 36, non figs. 31–35
1949 Callionymus primus Weiler, 1943; Weiler 1949: pl. 4, fig. 36, non figs. 32, 34, 35.
Material 2 specimens NMNH 2532/142, 143, Karagaily, Level 10.
Discussion Four specimens from Karagaily and one of Weiler’s paratypes from Romania (Fig. 9-3) differ from P.? primus as described above in the lack of a clear rostrum and antirostrum and in being more slender (OL:OH = 1.9–2.1 vs 1.6–1.9) and displaying a narrower ostium. However, all involved specimens are smaller than 1 mm in length and are probably from juveniles. We have also refrained from formally separating the specimens into separate species because there is still considerable uncertainty regarding ontogenetic changes as well as the level of intraspecific variation in P.? primus and the nature of P.? konkensis.
Protonymus? aff. miocenicus (Pobedina, 1954)
Figure 9-7 to -9
? 1950 Callionymus primus Weiler, 1943; Weiler 1950: pl. 7, fig. 52.
? 1954 Otolithus (Trigla) miocenicus; Pobedina 1954: pl. 1, figs. 5, 6.
? 1956 Otolithus (Trigla) miocenicus Pobedina, 1954; Pobedina 1956: pl. 10, figs. 3, 4.
1966 Callionymus primus Weiler, 1943; Smigielska 1966: pl. 18, fig. 3.
1981 Trigla miocenicus triangularis; Strashimirov 1981b: pl. 2, figs. 7, 8.
? 2006 Callionymus miocenicus (Pobedina, 1954); Djafarova 2006: pl. 17, fig. 2 (non fig. 1, non pl. 16, fig. 9).
Material 3 specimens NMNH 2532/138, 139, 145, Karagaily, Levels 9 and 10.
Diagnosis OL:OH = 1.35–1.55. Middorsal angle high, rounded; ventral rim slightly convex. Rostrum projecting, except short in small specimens < 0.4 mm in length. OL:SuL = 1.5–1.6.
Description (n = 3) Only one of the three specimens from Karagaily placed in P.? aff. miocenicus is about 1 mm long (Fig. 9-8) and exhibits a morphology which might be considered diagnostically mature. The two other specimens are certainly from juveniles and show a shorter rostrum and a shallow excisura. All otoliths are roughly triangular in outline with low but gently curved ventral rim and prominent, high mediodorsal angle. OL:OH = 1.35–1.55; OL:OT = 2.5–3, decreasing with size. Rostrum close to ventral rim, usually well developed, moderately long, pointed, short in specimens smaller than 0.4 mm in length; antirostrum and excisura well developed in large specimen (Fig. 9-8), feeble in small specimens (Fig. 9-7). Posterior tip moderately close to ventral rim, slightly projecting, rounded. All rims smooth.
Inner face almost flat, with typical callionymid upward-oriented sulcus. Ostium and cauda narrow, ostium more deepened than cauda. Ostium about twice as long as cauda, slightly widened posteriorly towards joint with cauda, anteriorly open. Cauda straight, terminating at less than 1/3 of otolith length from posterior tip of otolith. OL:SuL = 1.5–1.6; OsL:CaL variable, 1.3–1.9. Dorsal depression distinct, short; ventral furrow well developed, moderately close to ventral rim of otolith. Outer face convex, smooth.
Discussion Otoliths of Protonymus? aff. miocenicus are the most compressed of the otolith-based callionymid species observed in the Konkian of Mangyshlak. Their allocation to the genus Protonymus is therefore tentative. Pobedina’s figures in 1954 (reposted in Pobedina 1956) show otoliths with an apparently deeper curved ventral rim and a rather narrow, poorly structured sulcus. We assume that part of the discrepancy with our specimens from Mangyshlak is caused by the orientation of the figure and the somewhat schematic drawing, which may have resulted from the small size of the otoliths and difficulties in finding pertinent diagnostic features to draw. The figures by Strashimirov show the same orientation, while Smigielska and Djafarova show their otoliths with the typical callionymid orientation. The photos from Smigielska, identified as Callionymus primus, show the best agreement with our largest specimen. Nevertheless, some degree of uncertainty will remain until Pobedina’s type material can be revisited, if it is still preserved.
Suborder Blennioidei Bleeker, 1859
Family Blenniidae Rafinesque, 1810
Genus Parablennius Miranda Ribeiro, 1915
Type species. Blennius pilicornis Cuvier 1829, type by monotypy, Brazil; Recent.
Parablennius prokofievi n.sp. Bratishko, Schwarzhans and Reichenbacher
Figure 9-10 to -14
Etymology In honor of A. Prokofiev, Moscow, for his contributions to the knowledge of Recent and fossil fishes of Russia.
Type material Holotype: NMNH 2532/089 (Fig. 9-12). Paratypes: 5 specimens NMNH 2532/087, 088, 090, 091,146 (Fig. 9-10, -11, -13, -14). Additional specimens: 5 otoliths from Karagaily, Level 10.
Type locality Karagaily, Mangyshlak, Kazakhstan.
Age Konkian, early Serravallian, middle Miocene.
Diagnosis Small, compact, thick otoliths, OL:OH = 1.05–1.3; OL:OT = 2.5–3.0. Dorsal rim high, broadly rounded. Ventral rim shallow. Rostrum blunt, short, but sometimes long, with rounded tip. Posterior rim broadly rounded. Posterior half of cauda widened and slightly deepened, anterior half narrower and shallower. Dorsal field low; ventral field bulged.
Description (n = 5) Small, high-bodied otoliths less than 1.5 mm in length. OL:OH = 1.05–1.3; OL:OT = 2.5–3. Dorsal rim high, without prominent angles; ventral rim shallow, almost straight and horizontal. Posterior tip rounded; anterior tip with variably developed rostrum ranging from nearly absent (Fig. 9-10, -13) to mostly short and to distinct and rather prominent with rounded tip (Fig. 9-14); excisura and antirostrum minute. All rims smooth.
Inner face slightly convex, with slightly supramedianly positioned long sulcus. Ostium short, anteriorly open, deepened and somewhat widened, often with clearly marked outline of ostial colliculum. Cauda slightly bent upwards, anterior half narrower and shallower than widened and deepened posterior half. Cauda terminating close to posterior tip of otolith; posterior portion of cauda usually with well-marked colliculum; no colliculum seen in anterior portion of cauda. CaL:OsL = 1.45–1.65. Dorsal field generally low, depressed; ventral field between well-marked ventral furrow and sulcus bulged, inflated. Outer face flat to slightly convex, smooth.
Discussion Otoliths of Parablennius prokofievi are very typical, with a sulcus resembling that found in several genera of extant blenniids. The compressed shape and the cauda being “subdivided” into two parts of near-equal length are found in Recent otoliths of Omobranchus Valenciennes, 1836 and Parablennius (see Smale et al. 1995 for figures), with those of Parablennius showing the same kind of restricted collicular filling in the cauda.
Bannikov (2010) lists two monotypic fossil skeleton-based blenniid genera from the early and middle Miocene, Mioblennius Bannikov, 1998 and Bestiolablennius Prokofiev, 2001, both predating our otolith finds from Mangyshlak, and a fossil representative of the related family Clinidae from the early Sarmatian, Clinitrachoides gratus (Bannikov, 1989) with an otolith in situ, which however is readily distinguished by its more elongate shape, long rostrum and narrow sulcus.
Suborder Gobioidei Jordan and Evermann, 1896
Family Gobiidae Cuvier, 1816
Genus Aphia Risso, 1827
Type species. Aphia meridionalis Risso 1827 (synonym of Atherina minuta Risso 1810), type by monotypy, Mediterranean Sea; Recent.
Aphia djafarovae n.sp. Bratishko, Schwarzhans and Reichenbacher
Figure 10-1 to -6

1–6 Aphia djafarovae n.sp. Bratishko, Schwarzhans and Reichenbacher. 1: Paratype NMNH 2532/015, ventral view (1B). 2: Paratype, NMNH 2532/116, ventral view (2B). 3: Paratype, NMNH 2532/117, ventral view (3B). 4: Holotype, NMNH 2532/018, ventral view (4B). 5: Paratype, NMNH 2532/119. 6: Paratype, NMNH 2532/120. 7–12 Knipowitschia suavis Schwarzhans, 2014. 7: NMNH 2532/097, dorsal view (7B). 8: NMNH 2532/103, dorsal view (8B). 9: NMNH 2532/098, dorsal view (9B). 10: NMNH 2532/100, dorsal view (10B). 11: NMNH 2532/101. 12: NMNH 2532/102. Figures 7, 8, 11, 12 are mirrored. 13–17 Pomatoschistus bunyatovi n.sp. Bratishko, Schwarzhans and Reichenbacher. 13: Paratype, NMNH 2532/122. 14: Holotype, NMNH 2532/123, dorsal view (14B). 15: NMNH 2532/121, dorsal view (15B). 16: NMNH 2532/124. 17: NMNH 2532/125. Figures 13, 14, 15, 16, 17 are mirrored
Etymology In honor of the late J. Djafarova, for her contribution to the knowledge of fossil otoliths from Azerbaijan.
Type material Holotype: NMNH 2532/118 (Fig. 10-4). Paratypes: 5 specimens NMNH 2532/115–117, 119, 120 (Fig. 10-1–-3, -5, -6). Additional specimens: 24 otoliths from Karagaily, Level 10.
Type locality Karagaily, Mangyshlak, Kazakhstan.
Age Konkian, early Serravallian, middle Miocene.
Diagnosis Small size up to 1 mm. OL:OH = 0.9–1.0. Outline round with slightly flattened posterior rim. Inner face completely flat; outer face convex. Sulcus shifted towards anterior part of inner face; ostium much larger and deeper than cauda. No subcaudal iugum.
Description (n = 6) High-bodied, moderately massive otoliths; OL:OH = 0.9–1.0; OL:OT = 2.7–3.2. Outline regularly rounded with (usually) a somewhat flattened, blunt posterior rim; less commonly with a flattened middle section of ventral rim too. All rims smooth.
Inner face completely flat. Sulcus eccentrically positioned and relatively close to anterior rim, with typical gobiid sole shape. Sulcus inclination 10–20°. Ostium inclined, much deeper, wider and longer than cauda. OsL:CaL = 1.9–2.5. Dorsal depression wide, with indistinct margins; ventral furrow broad, indistinct at some distance from ventral margin of otolith. Outer face convex, smooth.
Discussion Otoliths from the dwarf genera Aphia and Crystallogobius can be distinguished from juvenile otoliths of other, larger gobiid species mainly by the peculiar eccentrically positioned sulcus and the deepened ostium showing a clear margin towards the shallower cauda (see Härkönen 1986 and Lombarte et al. 2006 for figures of Recent otoliths). Many small gobiid otoliths have been described in the literature from the Eastern Paratethys (see below), but they all show a sulcus centrally positioned on the inner face and thus may, to a certain extent, represent juveniles of other gobiid species. Exceptions are Trimma triangularis (Weiler, 1943) (see Schwarzhans 2010 for discussion) and Gobius atropatanus Djafarova, 2006 from the middle Sarmatian s.l. (Bessarabian) of Azerbaijan. The latter probably also represents the genus Aphia and is characterised by an extremely compressed outline with an OL:OH of 0.7–0.75. Another fossil species of the genus Aphia has recently been described from the Tortonian equivalent of the North Sea Basin—A. weinbrechti Schwarzhans, 2010. Aphia djafarovae differs in the slightly less compressed outline (OL:OH = 0.9–1.0 vs 0.85–0.95) and the large ostium (OsL:CaL = 1.9–2.5 vs 1.1–1.5).
The following nominal species, including nonvalid species figured by Suzin (1968, names not available according to ICZN article 13.1.1) from the Neogene of the Eastern Paratethys, should not be used until the original material has been reviewed because they are suspected to represent juvenile specimens: Gobius captiosus Pobedina, 1954; G. imperfectus Pobedina, 1954; G. latus Suzin, 1968; G. lawis Suzin, 1968; G. rotundus Pobedina, 1954; G. rotundus tarchanicus Pobedina, 1954; G. rotundus tchokrakensis Strashimirov, 1980; G. sarmatus Suzin, 1968; G. tenuis Suzin, 1968 (probably a synonym of Trimma triangularis); Hymenocephalus quadratus Strashimirov, 1981a (a Gobiidae). The species G. crenulatus Weiler, 1943; G. dorsolobatus Weiler, 1943 have been reviewed and are considered to represent juveniles of unknown gobiids; although nominally valid, it is suggested that they should not be used.
Genus Knipowitschia Iljin, 1927
Type species. Gobius longecaudatus Kessler 1877, type by monotypy, southern and middle Caspian Sea; Recent.
Knipowitschia suavis Schwarzhans, 2014
Figure 10-7 to -12
2014b Knipowitschia suavis; Schwarzhans 2014b: pl. 10, figs. 7–14.
Material 125 specimens NMNH 2532/097–103, Karagaily, Level 10.
Description OL:OH = 0.85–0.9; OH:OT = 3.0–3.5. Curvature index of inner face 5–13 % of OL. Outline with flat ventral rim, marked pre- and postventral angles, high mediodorsal and low predorsal angles and postdorsal projection weak or absent. Sulcus inclination <10°. Ostial lobe low; subcaudal iugum very long, extending below entire cauda and, at times, beyond.
Discussion This species is best recognised by the very long subcaudal iugum (for specific gobiid otolith terminology see Schwarzhans 2014b). The specimens from Mangyshlak seem to grow to slightly larger sizes (about 1.5 mm) than those described from the Karaman Basin, SE Turkey (up to 1.2 mm), but otherwise correspond very well. This is the only species to co-occur in the middle Miocene of the SE Mediterranean and the Eastern Paratethys. A similar otolith has been described from the Pleistocene of Azerbaijan as Gobius gansi Suzin in Klein (1960) but, according to Klein’s excellent drawings, is lacking a distinct subcaudal iugum.
Genus Neogobius Iljin, 1927
Type species. Gobius fluviatilis Pallas, 1814, type by monotypy, Black Sea and Caspian estuaries; Recent.
Neogobius udovichenkoi n.sp. Bratishko, Schwarzhans and Reichenbacher
Figure 11-1–-8

1–8 Neogobius udovichenkoi n.sp. Bratishko, Schwarzhans and Reichenbacher. 1: Paratype, NMNH 2532/071, dorsal view (1B). 2: Paratype, NMNH 2532/166. 3: Holotype, NMNH 2532/075, dorsal view (3B). 4: Paratype, NMNH 2532/073, dorsal view (4B). 5: Paratype, NMNH 2532/072. 6: Paratype, NMNH 2532/169, dorsal view (6B). 7: Paratype, NMNH 2532/170. 8: Paratype, NMNH 2532/171. Figures 1, 2, 6 are mirrored. 9–21 Ponticola zosimovichi n.sp. Bratishko, Schwarzhans and Reichenbacher. 9: Paratype, NMNH 2532/077, dorsal view (9B). 10: Paratype, NMNH 2532/078, dorsal view (10B). 11: Paratype, NMNH 2532/114, dorsal view (11B). 12: Holotype, NMNH 2532/076, dorsal view (12B). 13: Paratype, NMNH 2532/079, dorsal view (13B). 14: Paratype, NMNH 2532/082. 15: Paratype, NMNH 2532/080. 16: Paratype, NMNH 2532/085, dorsal view (16B). 17: Paratype, NMNH 2532/084, dorsal view (17B). 18: Paratype, NMNH 2532/113, dorsal view (18B). 19: NMNH 2532/074, dorsal view (19B). 20: NMNH 2532/174. 21: NMNH 2532/173. Figures 10, 12–14, 17, 19 are mirrored
Etymology In honor of N. Udovichenko (Luhansk) for his contribution to research of the fossil ichthyofauna of Western Eurasia and his support as mentor of the senior author.
Type material Holotype: NMNH 2532/075 (Fig. 11-3). Paratypes: 10 specimens NMNH 2532/071–074, 166, 167, 169–172 (Fig. 11-1, -2, -4–-8). Additional specimens: 16 otoliths from Karagaily, Level 10.
Type locality Karagaily, Mangyshlak, Kazakhstan.
Age Konkian, early Serravallian, middle Miocene.
Diagnosis OL:OH = 1.2–1.4; OL:OT about 5. Curvature index of inner face 15–20 % of OL. Preventral projection short or absent; postdorsal projection massive, long, strongly bent outwards. Predorsal angle high. Dorsal rim highest at about middle of rim, undulating, sometimes coarsely crenulated. Sulcus inclination about 10°. Ostial lobe moderate to low; no subcaudal iugum.
Description (n = 11) Moderately elongate, compact otoliths up to about 3.3 mm in length. Outline rectangular, with weak preventral projection or well marked angle, well-developed predorsal angle, positioned high at anterior rim, angular or rounded postventral angle and massive and long postdorsal projection. Ventral rim flat, smooth, horizontal; dorsal rim moderately curved, somewhat irregularly undulating or coarsely, sometimes deeply crenulated; anterior rim nearly vertical; posterior rim vertical, with slight indentation at level of sulcus.
Inner face moderately convex, but postdorsal projection strongly bent outwards. Sulcus centrally positioned, anteriorly inclined at about 10°, rather shallow and with typical sole-shaped although weakly developed ostial lobe. Subcaudal iugum absent. Dorsal depression wide, with indistinct margins; ventral furrow distinct, broad, at some distance from ventral rim, anteriorly and posteriorly curving upward to at least sulcus level. Outer face slightly concave with little ornamentation or smooth.
Discussion In a recent phylogenetic review of the Ponto-Caspian Gobiidae, Medvedeva et al. (2013) presented results based on cytochrome b extraction that showed a clear separation of the two genera Neogobius and Ponticola. These observations are corroborated by otolith analysis (ongoing research), which show a compressed otolith shape in the species of the genus Neogobius and an elongate, parallelogram-like shape with prominent preventral and postdorsal projections in species of the genus Ponticola. Both genera are already presented in the Konkian of Mangyshlak with typical forms.
Neogobius udovichenkoi resembles the Recent N. melanostomus (Pallas, 1814), which shows a very similar outline except for a more pronounced preventral projection and the presence of a subcaudal iugum (see Jacobs and Hoedemakers 2013 for figures of Recent otoliths). A similar fossil otolith is “Gobius” pricaspicus Pobedina, 1956 from the middle Sarmatian s.l. of Azerbaijan, although this differs in the higher dorsal rim, shorter postdorsal projection and a rather small sulcus. This species may perhaps be related to the genus Zosterisessor Whitley, 1935 because of the small sulcus.
Genus Pomatoschistus Gill, 1864
Type species. Gobius minutus Pallas, 1770, type by original designation, Belgian Sea; Recent.
Pomatoschistus bunyatovi n.sp. Bratishko, Schwarzhans and Reichenbacher
Figure 10-13 to -17
1966 Gobius praeclarus Prochazka, 1893; Smigielska 1966: pl. 19, figs. 2–3.
2006 Pomatoschistus laevis Weiler, 1942; Djafarova 2006: pl. 19, figs. 1–3 (non pl. 18, fig. 4)
Etymology In honor of Y. Bunyatov (Aktay) for his invaluable support during the field work in Mangyshlak.
Type material Holotype: NMNH 2532/123 (Fig. 10-14). Paratypes: 4 specimens NMNH 2532/121, 122, 124, 125 (Fig. 10-13, -15–-17) from Karagaily, Level 10.
Type locality Karagaily, Mangyshlak, Kazakhstan.
Age Konkian, early Serravallian, middle Miocene.
Diagnosis OL:OH = 0.95–1.05; OH:OT = 2.7–3.2. Inner face flat to slightly convex. Outline regularly rectangular with rounded angles, dorsally wider than ventrally. Sulcus narrow, deepened, inclined at about 15°. Ostial lobe very low; no subcaudal iugum.
Description (n = 5) Small, thick otoliths with regular rounded to rectangular outline up to about 1 mm in length. All rims smooth. Dorsal rim gently curving without prominent angles, highest at about the middle. Ventral rim straight, horizontal, with broadly rounded pre- and postventral angles. Anterior and posterior rims inclined, resulting in location of longest axis of otolith distinctly above centre, mostly above sulcus.
Inner face flat to slightly convex and smooth. Sulcus median to slightly supramedian, narrow, somewhat deepened, moderately inclined at about 15°, with typical shoe-sole shape, but very low, feeble ostial lobe and very narrow caudal tip. No subcaudal iugum. Dorsal depression broad, shallow; ventral furrow distinct, wide, running at some distance from ventral rim, anteriorly and posteriorly curving upward above sulcus level. Outer face markedly convex, smooth.
Discussion Pomatoschistus bunyatovi is a typical species of the genus Pomatoschistus characterised by a regular outline and a narrow sulcus with a low ostial lobe. It differs from P. rueckertae Schwarzhans, 2014 from the Serravallian of SE Turkey in the flat inner face, the shape of the sulcus with its low ostial lobe and its deepening, and the distinct dorsal shift of the longest axis of the otolith. The latter character and the narrow sulcus distinguishes P. bunyatovi from otoliths of all known Recent species of the genus (Härkönen 1986; Lombarte et al. 2006) except for P. marmoratus (Risso, 1810), which shows a shallow and wider sulcus and a more convex inner face. Pomatoschistus bunyatovi is one of the few species also recorded from contemporaneous strata in the Outer Carpathian Foredeep in Poland.
Genus Ponticola Iljin, 1927
Type species. Gobius ratan Nordmann, 1840, type by subsequent designation, Odessa, Ukraine; Recent.
Ponticola zosimovichi n.sp. Bratishko, Schwarzhans and Reichenbacher
Figure 11-9 to -21
Etymology In honor of V. Zosimovich (Kyiv) for his contribution to the Palaeogene and Neogene stratigraphy of the Eastern Paratethys and his support as mentor of the senior author.
Type material Holotype: NMNH 2532/076 (Fig. 11-12). Paratypes: 11 specimens NMNH 2532/077–085, 113, 114. (Figure 11-9 to -11, -13 to -18). Additional specimens: 70 otoliths from Karagaily, Level 10 and 1 specimen from Level 9. Tentatively assigned specimens: 10 otoliths NMNH 2532/74, 173, 174 (Fig. 11-19 to -21) from Level 10.
Type locality Karagaily, Mangyshlak, Kazakhstan.
Age Konkian, early Serravallian, middle Miocene.
Diagnosis OL:OH = 1.5–1.7; OL:OT = 4.5–6.5. Curvature index of inner face 14–17 % of OL. Preventral projection very long, sharp; postdorsal projection long, moderately bent outwards. Predorsal angle high. Dorsal rim low, nearly flat to slightly curved, undulating. Sulcus inclination about 15–20°. Ostial lobe low; no or faint and very narrow subcaudal iugum.
Description (n = 12) Elongate, large otoliths reaching about 4 mm in length. Outline parallelogram-like, with long preventral projection and about equally as long postdorsal projection, predorsal angle orthogonal and positioned high at anterior rim, postventral angle orthogonal as well. Ventral rim flat, smooth, horizontal; dorsal rim flat to slightly curved without prominent angles except predorsally, irregularly undulating; anterior and posterior rims steeply inclined, latter with slight indentation at level of sulcus.
Inner face moderately convex, postdorsal projection moderately bent outwards. Sulcus centrally positioned, large, anteriorly inclined at about 15–20°, rather shallow and with typical sole-shaped although weakly developed ostial lobe. Subcaudal iugum absent or very faint and thin. Dorsal depression wide, with indistinct margins; ventral furrow distinct, broad, at some distance from ventral rim, anteriorly and posteriorly curving upward to at least sulcus level. Outer face slightly concave with little ornamentation or smooth.
Variability A number of specimens exhibit only a very weak or no preventral projection (Fig. 11-19 to -21). It is possible that they represent another species, but, due to the lack of further features that could support such differentiation, we refer to them as tentatively assigned specimens.
Discussion Ponticola zosimovichi resembles the Recent N. kessleri (Günther, 1861) (see Jacobs and Hoedemakers 2013 for figures of Recent otoliths), but these are even more elongate (OL:OH = 1.8–2.1 vs 1.5–1.7) and show a more strongly outwardly bent postdorsal projection.
Order Pleuronectiformes Bleeker, 1859
Suborder Pleuronectoidei Bleeker, 1859
Family Bothidae Regan, 1910
Genus Arnoglossus Bleeker, 1862
Type species. Pleuronectes arnoglossus Bloch and Schneider, 1801 (synonym of Pleuronectes laterna Walbaum, 1792), type by monotypy, no locality given; Recent.
Arnoglossus? tenuis (Schubert, 1906)
Figure 12-1 to -3

1–3 Arnoglossus? tenuis (Schubert, 1906). 1: NMNH 2532/093, ventral view (1B). 2: NMNH 2532/095, ventral view (2B). 3: NMNH 2532/096, ventral view (3B). 4–8 Dicologlossa patens (Bassoli, 1906). 4: NMNH 2532/109, ventral view (4B). 5: NMNH 2532/110, ventral view (5B). 6: NMNH 2532/134, ventral view (6B). 7: NMNH 2532/136, ventral view (7B). 8: NMNH 2532/132. 9–12 Solea rotunda (Priem, 1914). 9: NMNH 2532/094, ventral view (9B). 10: NMNH 2532/126, ventral view (10B). 11: NMNH 2532/128, ventral view (11B). 12: NMNH 2532/130
1906 Solea tenuis; Schubert 1906: pl. 20, fig. 9.
1954 Rhombus corius miocenicus; Pobedina 1954: pl. 1, fig. 1.
1954 Rhombus corius foliformis; Pobedina 1954: pl. 1, fig. 3.
1956 Rhombus corius miocenica Pobedina, 1954; Pobedina 1956: pl. 8, fig. 3.
1956 Rhombus corius foliformis Pobedina, 1954; Pobedina 1956: pl. 8, fig. 4.
2006 Rhombus corius foliformis Pobedina, 1954; Djafarova 2006: pl. 24, fig. 2.
Material 3 specimens NMNH 2532/093, 095, 096, Karagaily, Level 10.
Remark A detailed diagnosis and re-description of this species is given here because Schubert’s documentation is not optimal for species recognition.
Diagnosis Moderately elongate, thick otoliths with oval, well-rounded outline. Inner face flat to slightly convex. Sulcus deep, with considerably widened ostium, which is closed anteriorly. Sulcus inclination angle low, less than 10°.
Description (n = 3) Small, thick otoliths about 1 mm in length. OL:OH = 1.3–1.5; OL:OT = 2.8–4.0. Outline is a regular oval without significant angles; posterior rim sometimes somewhat expanded.
Inner face flat to slightly convex. Sulcus median, deepened, only slightly inclined. Ostial and caudal colliculi well marked, sometimes indistinctly differentiated. Ostium about 50 % wider than cauda and slightly longer. OL:SuL = 1.6–1.7; OcL:CcL = 1.2–1.4. Circumsulcal depression wide, distinct, leaving only cristae elevated around sulcus. Outer face convex, smooth.
The specimen shown in Fig. 12-1 is somewhat more elongate and thinner than the other specimens studied and is therefore only tentatively placed in A.? tenuis.
Discussion The wide, anteriorly closed ostium and the regular oval outline of these otoliths distinguish A.? tenuis from all known Recent species of the genus (Schwarzhans 1999) and may point to the presence of a separate, extinct genus. In his review of Schubert’s type material, Nolf (1981) considered the unique type a juvenile and later (2013) noted it as a doubtful species “based on a non-diagnostic juvenile otolith”. Although pleuronectiform otoliths generally bear few characters of diagnostic value, with the new material from Mangyshlak we now consider this species to be sufficiently well defined and to probably represent a small-sized fish species.
Family Soleidae Bonaparte, 1835
Genus Dicologlossa Chabanaud, 1927
Type species. Solea cuneata Moreau, 1881, type by original designation, Atlantic coast of France; Recent.
Dicologlossa patens (Bassoli, 1906)
Figure 12-4 to -8
1906 Solea patens; Bassoli 1906: pl. 2, fig. 4.
1906 Solea kokeni; Schubert 1906: pl. 20, fig. 8.
1999 Dicologlossa patens (Bassoli, 1906); Schwarzhans 1999: figs. 767–769 (with further synonymies).
2013 Dicologlossa patens (Bassoli, 1906); Schultz 2013: pl. 94, fig. 10 (refigured holotype of Solea kokeni).
Material 24 specimens NMNH 2532/109–112, 132–136, Karagaily, Level 10.
Discussion These thin, elongate soleid otoliths are easily recognised (Schwarzhans 1999). The history of its competing description by Bassoli and Schubert has been explained in the review of Schubert’s type material by Nolf (1981) and Bassoli’s type material by Nolf and Steurbaut (1983). Dicologlossa patens and Solea rotunda (see below) represent some of the few early Badenian species of the Paratethys that persist into the Konkian (early Serravallian) of the Eastern Paratethys.
Genus Solea Quensel, 1806
Type species. Solea vulgaris Quensel, 1806 (synonym of Pleuronectes solea Linnaeus, 1758), type by monotypy, Bay of Biscay, France; Recent.
Solea rotunda (Priem, 1914)
Figure 12-9 to -12
1914 Gobius rotundus; Priem 1914: fig. 66.
1984 Solea rotunda (Priem, 1914); Steurbaut 1984: pl. 36, figs. 5–8
1992 Solea solea (Linnaeus, 1758); Radwanska 1992: textfig. 162 pl. 38, figs. 10–14.
1999 Solea rotunda (Priem, 1914); Schwarzhans 1999: figs. 774–777.
Material 26 specimens NMNH 2532/94, 126–131, 137, Karagaily, Level 10.
Discussion Solea rotunda is recognised by the rather regular rounded outline, with the smoothly curved dorsal rim, the rounded anterior tip and the slightly deeper curved ventral rim, and the small postdorsal projection. The otoliths are thin, with a slightly convex inner and a flat outer face. They show increasing thickness with size, and reach sizes of slightly more than 2 mm in length. The narrow sulcus and circumsulcal depression are typical for soleid otoliths. The otoliths from Mangyshlak are on average slightly more elongate than those from the early Badenian of Poland described by Radwanska (OL:OH 1.23–1.33 vs 1.15–1.25) and also relatively thin, particularly in larger specimens. These subtle differences could indicate the presence of two separate species, but the available data from the various locations are not felt to be adequate to definitively draw such a conclusion.
Accompanying fossils and stratigraphy
Nannoplankton assemblages
Calcareous nannoplankton assemblages from Levels 8–10, 18 and 20 of the Karagaily section (Fig. 2) were investigated in order to define their precise chronostratigraphic position. The calcareous nannoplankton is generally well preserved. The following taxa were found to occur in all samples: Coccolithus pelagicus (Wallich, 1877) Schiller, 1930; Cyclicargolithus floridanus (Roth and Hay in Hay et al. 1967) Bukry, 1971; Reticulofenestra pseudoumbilicus (Gartner, 1967) Gartner, 1969; Helicosphaera carteri (Wallich, 1877) Kamptner, 1954; Reticulofenestra gelida (Geitzenauer, 1972) Backman, 1978 and Reticulofenestra minuta Roth, 1970. Found to sporadically occur: Calcidiscus leptoporus (Murray and Blackman, 1898) Loeblich and Tappan, 1978; Helicosphaera wallichii (Lohmann, 1902) Okada and McIntyre, 1977; Perforocalcinela fusiformis Bona, 1964; Rhabdosphaera sp.; Sphenolithus moriformis (Bronnimann and Stradner, 1960) Bramlette and Wilcoxon, 1967; Thoracosphaera heimii (Lohmann) Kamptner, 1944 and Thoracosphaera saxea Stradner, 1961.
The last occurrence (LO) of Sphenolithus heteromorphus Deflandre, 1953 defines the boundary between Zones NN5 and NN6 (Martini, 1971). The absence of this species in the investigated samples, together with the presence of Cyclicargolithus floridanus, thus indicates nannoplankton Zone NN6 as the maximum age. This zone can be correlated with the late Badenian (Konkian) and early Sarmatian (Volhynian/Bessarabian). The occurrence of Cyclicargolithus floridanus (which has its LO in the lower NN6) in all samples points to a late Badenian (Konkian) rather than an early Sarmatian age. In the Central Paratethys, this part of NN6 can be attributed to the Kosovian (early Serravallian; late Badenian) (Hohenegger et al. 2014).
The samples from Levels 8 to 10 contain Braarudosphaera bigelowii (Gran and Braarud, 1935) Deflandre, 1947, which is particularly abundant in Level 8. High percentages of Braarudosphaera bigelowii characterise palaeoenvironments with reduced salinity (usually caused by freshwater inflow or shallowing), high nutrient levels and cold temperatures. In addition, Levels 8–10 are characterised by high percentages of reworked nannoplankton. Reworking from Eocene/Oligocene sediments is documented by occurrences of Reticulofenestra bisecta (Hay, Mohler and Wade, 1966) Roth, 1970; Reticulofenestra dictyoda (Deflandre in Deflandre and Fert, 1954) Stradner in Stradner and Edwards, 1968; Reticulofenestra hillae Bukry and Percival, 1971; Reticulofenestra lockeri Müller, 1970; Reticulofenestra stavensis (Levin and Joerger, 1967) Varol, 1989; Zygrhablithus bijugatus (Deflandre in Deflandre and Fert, 1954) Deflandre, 1959. Reworking of upper Cretaceous sediments is attested to by Arkhangelskiella cymbiformis Vekshina, 1959; Arkhangelskiella maastrichtiana Burnett, 1998; Eiffellithus turriseiffeli (Deflandre) Reinhardt, 1965; Prediscosphaera cretacea (Arkhangelsky, 1912) Gartner, 1968; Retecapsa crenulata (Bramlette and Martini, 1964) Grün in Grün and Allemann, 1975. The abundance of B. bigelowii, coupled with strong reworking in samples L8 and L9, almost certainly indicates decreasing salinity and shallowing in this part of the studied section. It can be assumed that freshwater influx from a river contributed Eocene/Oligocene sediments from the hinterland and probably triggered the high productivity of the coastal sea.
An additional nannoplankton sample from Level 10 was examined by L. Golovina (Moscow) and yielded abundant Reticulofenestra pseudoumbilicus (>7 µm), Braarudosphaera bigelowii and rare specimens of Rhabdosphaera sicca, Cyclicargolithus floridanus and Helicosphaera carteri. This assemblage is very similar to those known from the uppermost part of the Konkian in the Taman Peninsula, and confirms the correlation with late Badenian (Kosovian) nannofossil associations from the Central Paratethys (Radionova et al. 2012).
The sample from Level 20 did not contain any calcareous nannofossils.
Foraminiferal and ostracod assemblages
The following microfossils have been identified in the samples (Table 1):
Level 8 Ostracods: A single juvenile specimen of Euxinocythere stabilis Schneider.
Level 9 (sample no. 1) Benthic foraminifera: Nonion bogdanowichi Voloshinovae, N. biporus Kracheninnikov, Porosononion martkobi (Bogdanowicz), Elphidium macellum (Ficht. et Moll), E. horridum Bogdanowicz, Ammonia ex gr. beccari (L.), Bulimina aff. leninabadensis (Z. Kuzn.).
Ostracods (identifications of V. Kovalenko): Loxoconcha turgida Stancheva, Amnicythere mironovi mironovi (Schneider), A. mironovi estranea Stancheva, Euxinocythere naviculata (Schneider), E. pseudonaviculata Stancheva, Eux. gerke Suzin).
Level 10 (Sample no. 2) Benthic Foraminifera: Obtained foraminifera represent a melange of euryhaline species (Nonion bogdanowichi, N. biporus, Porosononion martkobi, Elphidium macellum, E. angulatum, E. horridum, Ammonia ex gr. beccari) and normal-marine species (Quinqueloculina aff. gracilis, Globulina gibba, Discorbis kartvelicus, Bulimina aff. leninabadensis, Cassidulina ex gr. bulbiformis) (see Krasheninnikov 1959; Bogdanovich 1965). Some species are typical for the Konkian of the Eastern Paratethys (Quinqueloculina aff. gracilis, Discorbis kartvelicus, Nonion biporus, Globulina gibba, Cassidulina ex gr. bulbiformis) (Krasheninnikov 1959; Bogdanovich 1965; Djanelidze 1970). Bulimina aff. leninabadensis is common in the lower part of the lower Sarmatian, but already occurs, albeit rarely, in the Konkian (Bugrova et al. 2005). Elphidium horridum occurs during the late Konkian and the lower part of the early Sarmatian (Bogdanovich 1965; Djanelidze 1970). Elphidium angulatum is known from the Konkian to the middle Sarmatian, and Nonion bogdanowichi and Porosononion martkobi from the Konkian to Early Sarmatian; all of these species are usually indicative of shallow-water Konkian sediments (Krasheninnikov 1959; Bogdanovich 1965).
Ostracods (identifications of V. Kovalenko): The obtained ostracod fauna also represents a mixture of brackish-water species (Euxinocythere stabilis, E. naviculata, E. pseudonaviculata, E. gerke, Amnicythere mironovi mironovi, A. mironovi estranea, Cyprideis torosa littoralis), brackish-marine (Paracytherois tenerum) and marine species (Loxoconcha turgida, L. assimulatoformis, L. elliptica). Almost all of these species have a wide stratigraphic range (see Suzin 1956; Voroshilova 1957; Stancheva 1990; Liuliev 1967), but Amnicythere mironovi estranea, Euxinocythere pseudonaviculata, E. gerke, Loxoconcha assimulatoformis are considered to be typical of the middle Sarmatian deposits of the Eastern Paratethys (see Suzin 1956; Liuliev 1967). However, the knowledge base of middle Miocene ostracods from the Eastern Paratethys is not deep enough to allow a detailed biostratigraphic evaluation.
Level 18 Ostracods: A single specimen of Paracytherois tenerum (Brady, Grosskey et Robertson).
Level 20 Benthic foraminifera: Lobatula lobatula (Walker et Jacobs); molluscs: Spirialis sp.
Mollusc assemblages
Molluscs occur abundantly in sample no. 2 (corresponding to Level 10) (see Table 1). They comprise marine euryhaline species of Bivalvia (Musculus naviculoides (Koles.), Mytilaster volhynicus buglovensis (Gatuev), Obsoletiforma litopodolica ruthenica (Hilber), Ervilia dissita dissita (Eichw.), Abra alba scytica (Sokolov), Pholas sp., Barnea sp.) and marine to mesohaline species of Gastropoda (Tectura incognita Friedb., T. laevigata (Eichw.), Turritella pithagoraica Hilber, Hydrobia sp., Mohrensternia nitida Zhizhchenko, M. laskarevi Liverovskaya, M. inflata (Hörnes), Nassarius sp. (juv.), Chrysallida interstincta (J. Adams), Chrysallida sp., Acteocina lajonkaireana (Basterot), Retusa truncatula (Bruguiere), Retusa sp., Cylichnina sp.).
Table 1 shows the stratigraphic distribution of these species in the Miocene of the Eastern Paratethys according to previous studies (Ilyina 1993; Nevesskaya et al. 1993). Obsoletiforma litopodolica ruthenica is known from the late Konkian to early Sarmatian (chiefly in the lower part) (Nevesskaya et al. 1993), but Ali-Zadeh (1974) also reported this species from the middle Sarmatian deposits of Azerbaijan. Ervilia dissita dissita is known from lower and middle Sarmatian sediments. Small-sized shells of this species, such as those present in sample no. 2, are typical of the lower part of the lower Sarmatian in the deposits from the Eastern Paratethys (Kolesnikov 1935), but also occur in the Konkian sediments of Mangyshlak, Northern Caucasus, Konka River as well as in the upper Badenian (Buglovian) of Volhyn (West Ukraine) (Liverovskaya 1935). The species of Turritella, Tectura, Chrysallida, Nassarius, Acteocina only lived during the early middle Miocene in the Paratethys and disappeared after the Konkian (Ilyina 1993).
The species of Mohrensternia are particularly important for biostratigraphic zonations in the middle Miocene of the Eastern Paratethys (Anistratenko 2005; Ilyina 2006). Three species of Mohrensternia—M. laskarevi, M. inflata and M. nitida—were found in sample no. 2. M. laskarevi was identified by Liverovskaya (1935) from the Konkian deposits of the North Caucasus and from upper Badenian deposits (Buglovian). Ilyina (2006) considered M. laskarevi to have evolved in the late Konkian of the Eastern Paratethys. Mohrensternia nitida is known from Chokrakian and Konkian deposits (Zhizhchenko 1936), while M. inflata was reported by Sokolow (1899) from the lower part of the early Sarmatian from the Konka River (Konkian stratotype). Liverovskaya (1935), however, recorded M. inflata from the Konkian deposits of Dubrovaya (North Caucasus). In 2006, Ilyina revised the species of the genus Mohrensternia and, according to her, M. inflata first appeared in the Eastern Paratethys during the lower part of the early Sarmatian.
V. Anistratenko (Kiyv) kindly reviewed the gastropod shells from sample no. 2 (identified as M. laskarevi, M. inflata and M. nitida) and advised that, in his opinion, the specimens described as “M. inflata var.” sensu N. Sokolov (1899) by Zhizhchenko (1936) as well as “M. inflata” sensu V. Laskarev (1903) do not represent “M. inflata” sensu stricto, but instead have to be placed in M. nitida Zhizhchenko, 1936. Anistratenko further advised that, in agreement with Liverovskaya (1935), only the specimen illustrated by Laskarev (1903, Pl. 5, Fig. 31) as M. inflata (Hörnes, 1856) is genuine, while the two other figures of “M. inflata” (sensu Laskarev 1903, Pl. 5, Figs. 29–30) represent M. laskarevi Liverovskaya, 1935. Mohrensternia inflata sensu Laskarev (1903, Pl. 5, Fig. 31) corresponds with the specimens from Karagaily identified as M. inflata and also corresponds to M. inflata sensu Hörnes, 1856 (see Anistratenko, 2005). Therefore, it can be concluded that both M. inflata (Hörnes, 1856) and M. laskarevi Liverovskaya, 1935 occur in the Buglovian of Volhynia, West Ukraine (Laskarew 1903) and the Konkian of Dubrovaya (northern Caucasus) (Liverovskaya 1935). Consequently, and in conflict with the postulation of Ilyina (2006), both species may have occurred contemporaneously during Konkian times.
The molluscs of sample no. 2 include both mesohaline species, which are characteristic of normal marine middle Miocene sediments, and euryhaline species, which are typical of the closed Eastern Paratethys sea basin of the early Sarmatian. Similar observations were made by Sokolov (1899) for the upper Konkian sediments of the Konka River stratotype section, as well as by Liverovskaya (1935, 1960) from the Konkian of the northern Caucasus and Mangyshlak. Unfortunately, Liverovskaya did not specify the part of the Konkian section from which these gastropod shells were obtained. Ilyina (2000) presented data from various researchers which also highlighted a mixture of euryhaline (typical Sarmatian) and euhaline (typical Konkian) molluscs from the upper Konkian deposits of the northern Caucasus.
In 1953, Merklin identified three stages in the development of the Konkian deposits—the Kartvelian, Sartaganian and Veselyankian. He defined the late Konkian (Veselyankian) as (translated from Russian in senso lato): “desalination basin, which came into being as a result of the closure of the gateway which connected the Konkian Sea with the Mediterranean. In such a basin only the most widely adapted ecological species of molluscs survived and endemic species and subspecies arose. Some of these species became extinct at the end of the Konkian. Others gave rise to the Sarmatian mollusc fauna”. Later, the Veselyankian was also described as “layers with an impoverished (euryhaline) marine Konkian fauna” (Vereshagin and Mironova 1982; Nevesskaya et al. 1986). However, certain researchers have mentioned that layers with euryhaline faunas are often found to be interbedded with layers containing a marine fauna in different levels of the Konkian and in different areas of the Eastern Paratethys (Vernigorova 2009). Therefore, a euryhaline composition of the Konkian fauna alone cannot be maintained as a criterion for the determination of a late Konkian age.
According to unpublished data available to us and data published by Bogdanovich (1965), the uppermost parts of certain Konkian sections from the northern Black Sea shores and the Kerch Peninsula and the northern Caucasus contain (as seen in mollusc assemblages) foraminifera assemblages of marine and euryhaline species that are considered to be characteristic of lower Sarmatian sediments. It is therefore possible to assume that the occurrence of a large percentage of Sarmatian species together with assemblages of typical Konkian molluscs and foraminifers is indicative of the latest time interval of the Konkian Stage.
Results and discussion
Composition and environmental evaluation of the fish fauna
More than 2000 otolith specimens were recovered from the two levels of the Konkian strata of the Karagaily section studied here. The otolith assemblage represents a faunal composition that is typical of the shallow-shelf or nearshore environment. It is dominated by just a few species of the families Gadidae and Gobiidae, and the latter are often found to be very common in nearshore marine-to-brackish environments. The most common species is the gadid Palimphemus minusculoides—which comprises about 70 % of the entire assemblage, while the gadid Onogadus simplicissimus accounts for 4.2 % of the whole. The most common gobiid taxa are Knipowitschia suavis (7 %), Ponticola zosimovichi (5.5 %), Aphia djafarovae (1.7 %) and Neogobius udovichenkoi (1.5 %). Other accessory species (between 1 and 2 %) are a sciaenid (Genyonemus? karagiensis) and two soleids (Dicologlossa patens and Solea rotunda). The level of species diversity is moderate, with 11 species accounting for 90 % of the total assemblage.
No otoliths indicative of open marine mesopelagic or benthopelagic fishes were found. For instance, representatives of the families Myctophidae, Bregmacerotidae or Macrouridae, which are common in pelagic sediments of the Neogene of the Mediterranean or the deep-water environments of the Central Paratethys (Schubert 1906; Holec 1975; Brzobohaty 1995) were entirely absent. No clear evidence of nonmarine fishes has been observed, and brackish-water species are rare. If such forms had been present in significant numbers, one would expect to find them among the Gobiidae, given that the Recent Ponto-Caspian fauna includes a large number of euryhaline or brackish-water and freshwater species belonging to the genera Neogobius and Ponticola. Of course, it cannot be excluded that some of the gobiid species identified could tolerate brackish water or even freshwater and, in one case (Knipowitschia suavis), the occurrence in both marine and brackish environments has already been observed in the Serravallian of the Karaman Basin in SE Turkey (Schwarzhans 2014b). Other species characteristic of brackish to nonmarine environments that are known from Turkey or the Central Paratethys, such as Neogobius rhachis Rückert-Ülkümen and Kaya, 1993 or Gobius dorsorostralis Weinfurter, 1954, have not been found.
In this regard, one must also consider the rapid changes in water salinity that occurred within the Paratethys during the Neogene. Liverovskaya (1960) and Ilyina (2006) interpreted the palaeoenvironment of the late Konkian of the Eastern Paratethys as having been characterised by a slightly reduced salinity, which in Ilyina’s assessment would explain why certain rissoid gastropods, such as Mohrensternia and subgenera of Rissoa, flourished at this time. She also postulated that a more substantial decrease in salinity would have been detrimental to these rissoids, and that they therefore provide a good indicator of a slightly reduced marine salinity (without, however, specifying values). The fishes represented by otoliths, specifically the Gobiidae, Atherinidae, Mugilidae, Gaidropsaridae, Moronidae and Clupeidae (see Nelson 2006), all belong to groups that would probably have tolerated a moderate decrease in marine salinity. Other groups include forms that are known to occasionally migrate into brackish water and even freshwater—for instance, certain Sciaenidae, Polynemidae, Blenniidae (rare) or Pleuronectiformes, and some Recent taxa contain species that have adapted to nonmarine environments (Nelson 2006). Pterothrissidae, Mullidae and Callionymidae are exclusively or almost exclusively euhaline (Nelson 2006). Their presence in the Konkian of Karagaily indicates that if salinity was indeed reduced, it could only have been so to a very small degree.
We conclude that the fish fauna represented by the otoliths observed in the late Konkian of Mangyshlak is typical of a shallow marine, probably nearshore, environment in which water depths did not exceed 50 m and an open pelagic or estuarine influence was absent. Salinity was either typically marine or slightly less.
Comparison of skeleton- and otolith-based data
It has been long noted that skeleton- and otolith-based data generally do not match very well. Nolf (1985) devoted a short chapter to this phenomenon and observed “that in nearly every known fossil association, otoliths originated from fishes that were hardly 10–15 cm long”. He further stated: “In most cases, it is (also) striking that the bones of all the fishes, represented by thousands of otoliths, are totally absent in the sediments.” He went on to conclude that “it is very probable that most otoliths are derived from small fishes constituting the prey of larger predators,” whose bones are in turn well known from the corresponding fossil records. One might further argue that the conditions that favour the fossilisation of skeletons differ from those that promote the preservation of otoliths, and that otolith assemblages accumulated primarily in well-oxygenated clastic environments, whereas skeleton-based faunas were primarily deposited under anaerobic conditions or in limestones (see also Nolf 1985). All of these factors or a combination thereof may help to explain the observation that skeleton- and otolith-based fish faunas commonly complement each other.
However, many fish skeletons have been described from Karaganian, Konkian and Early Sarmatian deposits of the Eastern Paratethys by Bannikov (1989, 1993, 2006, 2008, 2010), Bannikov et al. (2009), Baykina (2013), Carnevale et al. (2006), Fedotov (1976), and Sytchevskaya and Prokofiev (2007), and they do correlate with otolith-based data to a reasonable extent (see Table 2). Clupeidae, Gadidae, Atherinidae, Mugilidae, Moronidae, Centracanthidae, Sciaenidae and Callionymidae are all represented in parallel by skeletons and otoliths. So far, three species have been recorded with otoliths in situ—Morone? ionkoi, Clinitrachoides gratus and Protonymus? primus. It is not unlikely that some skeleton- and otolith-based species would be found to be identical if adequate skeletons with otoliths in situ were identified. Bannikov (2010) mentioned in passing that he had regularly come across fish skeletons from the Sarmatian bearing otoliths in situ. [It is also worth mentioning in this respect that Schubert (1906) noted otoliths in situ in 10 skeleton-based species described by Kramberger-Gorjanovic from the Sarmatian of the Central Paratethys (Croatia). These have never been investigated (Schwarzhans 2014a), but they were excluded from our comparison as they represent rather different species from those reported in the Eastern Paratethys.]
There are, however, also discrepancies between the skeleton- and otolith-based fish records of this time interval in the Eastern Paratethys. The Clupeidae and Mugilidae are underrepresented by otoliths, while the Sciaenidae, Gobiidae and Pleuronectiformes are underrepresented by skeletons. Surprisingly, Gadidae also appear to be overrepresented by skeletons, but we suspect that a careful review of the skeleton data will either lead to a reduction in valid species and/or a disentanglement of the relative stratigraphic positions, so the apparent overrepresentation may well be superficial rather than real.
Biogeographical context
The Konkian fish fauna of the Eastern Paratethys shows a high degree of endemicity when compared with equivalent fish faunas of neighbouring European Basins in the same interval. Only nine of the 22 species recognised in Mangyshlak (excluding the eight species left in open nomenclature) are shared with the Badenian of the Central Paratethys (Table 3). This is an amazingly low percentage when one considers that the number of otolith-based species recorded so far from the Badenian of the Central Paratethys is well over 100. This faunal autonomy is further emphasised by the fact that not a single species is shared with the well-known Early Badenian—all nine shared species are first recorded in the Konkian of the Eastern Paratethys at a time that corresponds to the late Badenian of the Central Paratethys. A similar relation is observed between the skeleton-based fish data for the Karaganian to early Sarmatian s.l. of the Eastern Paratethys (Bannikov 2010; Baykina 2013; Fedotov 1976; Sytchevskaya and Prokofiev 2007) and the Sarmatian s.s. (early Sarmatian s.l.) of the Central Paratethys as described by Kramberger-Gorganovic (1883), with the caveat, however, that the latter requires a modern revision.
Previous reports on fossil otoliths from the Central Paratethys have rarely distinguished between the lower, middle and upper Badenian. The works of Smigielska (1966), Holec (1975), Brzobohaty (1980), Brzobohaty and Nolf (2000) are exceptional in this respect, although the former used the outdated term “upper Tortonian” in her descriptions. Judging from the locations cited, for instance in Schubert (1902, 1905, 1906), it is reasonable to assume that the majority of the otoliths described from the Pannonian and Vienna Basins originate from lower Badenian strata. But some of the sites annotated in Schubert (1906, 1912) represent the middle-upper Badenian (i.e. Walbersdorf = Borbolya and Theben-Neudorf = Deveny-Ujfalu = Devinska Nova Ves—in German, Hungarian and Slovak equivalents) in the sense of the Badenian stratotype (Papp et al. 1978). Recently, Hohenegger et al. (2014) modified the subdivision of the Badenian and accepted a redefinition and chronometric timing of the late Badenian substages Wielician and Kosovian as essential. The middle to late Badenian otolith-based fish associations within the Central Paratethys show a certain degree of faunal differentiation across the Pannonian Basin and exhibit a wide range of fishes from shelf to open marine environments. Walbersdorf (Austria) represents a deep marine, pelagic environment, and the otolith associations (Schubert 1906, Bachmayer and Weinfurter 1965, Brzobohaty 1995) are middle Badenian in age (Spirorutilus carinatus Zone in the sense of the regional stratigraphy, not an equivalent of the Kosovian, see Rögl et Müller 1975), while Devinska Nova Ves in Slovakia (Deveny Ujfalu) records a neritic or shelf environment. However, the literature dealing with otoliths does not permit clear differentiation or recognition of middle Badenian strata. In the Carpathian Foredeep and the Transylvanian Basin, the middle Badenian is characterised by an evaporitic event that separates the underlying lower Badenian from the uppermost Badenian—also known as the Buglovian (“Buglow” in Weiler 1943, 1949, 1950). The uppermost Badenian otolith associations described by Smigielska (1966), Brzobohaty (1980) and Weiler (1943, 1949, 1950) exhibit a less rich faunal composition with a few indigenous species not recorded from the Pannonian Basin. A few skeleton finds have been described from the middle Badenian of the Carpathian Foredeep of Poland by Jerzmanska (1962), and an otolith found in situ in one of them was recently described as Palimphemus anceps by Schwarzhans (2014a). The little that we know about a confirmed middle Badenian fish fauna thanks to these publications is more reminiscent of the underlying lower Badenian than the upper Badenian. Strashimirov (1982) described otoliths from the Badenian of the Morava Basin in the SW of Bulgaria and referred a few of them to the late Badenian.
Recently, Schwarzhans (2014b) described a shallow marine, near-shore otolith-based fish fauna from the late Serravallian (early Sarmatian s.l. equivalent) of the Karaman Basin in southeastern Turkey comprising 47 species, of which 33 were identified to the species level. The fish fauna from the Karaman Basin shows a rather close relationship to the slightly older Badenian fish fauna of the Central Paratethys (61 % shared species, see Schwarzhans 2014b), which is also clearly discernible in the very rich molluscan fauna (Landau et al. 2013). In contrast, only one species—Knipowitschia suavis—is shared between Karaman and Mangyshlak, despite the overall similarities between the two environments. Interestingly, K. suavis is one of three species in the Karaman Basin that are also recorded from a brackish environment. We therefore assume that K. suavis was a euryhaline species, much like many of the Recent members of the genus, and thus was able to penetrate into areas that would have represented barriers to the expansion of stenohaline or less adaptive species.
The origin of the Konkian fish fauna of the Eastern Paratethys
The palaeogeographic development of the Paratethys and its ever-changing pattern of seaway connections was highly complex during the relevant time window of the Langhian and Serravallian between 16 and 12 Ma, as depicted by Rögl (1999) and Popov et al. (2006) (see Figs. 13, 14). In a sequence of detailed palaeogeographic reconstructions, Rögl (1999) showed a Paratethys that was linked to the Mediterranean in the West and the Mediterranean and Indian Ocean in the East during the early Langhian (Tarkhanian, early part of early Badenian). At that time, a common fish fauna would have populated the Paratethys and Tethys. The otolith-based information available from the Central Paratethys (Schubert 1906, 1912; Radwanska 1992) and the much less well-known Tarkhanian otolith associations from the Eastern Paratethys (Pobedina 1954, 1956; Strashimirov 1972; Djafarowa 2006) are in good agreement, which supports the above hypothesis, again subject to the caveat that the Eastern Paratethys fauna requires a fundamental revision.

Konkian palaeogeography of the Paratethys with geographic provinces indicated by color coding (Indian Ocean inferred, NSB = North Sea Basin). A Possible connections between Central Paratethys and Mediterranean; B possible connection between Eastern Paratethys and Mediterranean. Palaeogeography modified after Rögl (1998, 1999) and Popov et al. (2006). Transitional color coding in the Paratethys indicates presumed fish-faunal changes as deduced from the otolith database. Asterisks denote prime locations studied for otoliths (one asterisk may denote many different locations in one region): *1: Schubert (1906, 1912); *2: Smigielska (1966), Brzobohaty (1980); *3: Weiler (1943, 1949, 1950); *4: Strashimirov (1981b); *5: Suzin in Zhizhchenko (1968); *6: Pobedina (1954, 1956) and Djafarova (2006); *7: this study; *8: Schwarzhans (2014b)

Konkian palaeogeography and distribution of the species of the gadid genus Palimphemus. Distributions were interpolated between data points. Areas that are not coloured lack data. For legend and map source, see Fig. 13
Our correlation of the Eastern Paratethys with the Central Paratethys stages and the international stages of the Langhian to Serravallian is based on Rögl (1998), Harzhauser et al. (2002), Piller et al. (2007) and Semenenko et al. (2009). During the Chokrakrian, Popov et al. (2006) show that the Central and Eastern Paratethys is already separated, with the Central Paratethys open to the West and the Eastern Paratethys to the East, as before. Our knowledge of otoliths from this time interval lacks stratigraphic resolution in the Central Paratethys, and tit is restricted and in need of profound revision for the Eastern Paratethys (Pobedina 1954, 1956; Strashimirov, 1980; Djafarova 2006). However, there does not seem to be any major differentiation between what is known from the Tarkhanian and the Chokrakrian otolith assemblages in the Eastern Paratethys (Table 4). Relatively few new entrants occur in the Chokrakrian, and the overall faunal composition had the same distinctive “Mediterranean” aspect as before. A few species occur for the first time, possibly indicating the onset of an endemic evolution in the Eastern Paratethys. These are Paratrisopterus insectus (recorded under multiple identities in (Djafarova, 2006), mostly as Macruridarum minusculus, which has not yet, however, been reported from Konkian strata in the Eastern Paratethys) and an otolith described by Djafarova as Osmeridarum crassa Djafarova, 2006, and possibly representing the clupeid genus Clupeonella.
The Karaganian/middle Badenian brought about a revolution in the configuration of the Paratethys. According to Rögl (1999), the Eastern Paratethys became isolated; in the Central Paratethys, the Pannonian Basin remained open to the Mediterranean while the Carpathian Foredeep and Transsylvanian Basin became isolated and subject to evaporite development (Kovac et al. 2007). Again, the stratigraphic resolution of otoliths described from the Pannonian Basin does not permit a detailed analysis, but this may serve as an indication that not much faunal change should be expected. The Eastern Paratethys has, however, experienced a major reshuffling of its fish fauna. Many of the Tarkhanian and Chokrakrian faunal elements of Mediterranean affinities have disappeared, for instance Bregmaceros and Bonapartia, while many new species appear for the first time (Pobedina 1954, 1956; Suzin 1968; Strashimirov 1981a; Djafarova 2006). Among the latter is the first callionymid (Protonymus? primus) and Atherina gidjakensis, which replaces the Langhian-to-Serravallian Atherina austriaca of the Pannonian and the Karaman Basins. Very similar observations are pithily summarised by Kovac et al. (2007), who states that “no Chokrakrian genus survived the Karaganian crisis”.
In the palaeogeographic reconstructions of Rögl (1999), Popov et al. (2006) and Kovac et al. (2007), the Konkian itself is depicted as a brief transgressive period in which connections with the Paratethys were re-established (Fig. 13). The degree of interconnection varies somewhat in the reconstructions presented by Rögl and Popov et al., as do the interpretations of seaway connections in the West to the Mediterranean and the East to the Indian Ocean. Rögl shows a Paratethys closed to the West and open to the East. Popov et al. depict restricted connections between Central and Eastern Paratethys and from the Eastern Paratethys to the Indian Ocean as well as potential connections of the Paratethys to the West. As mentioned above, the similarities between the Konkian fish fauna of the Eastern Paratethys and the late Badenian of the Central Paratethys are few, but they are significant in the sense that they all represent newcomers to the Central Paratethys (Onogadus simplicissimus, Palimphemus minusculoides, Paratrisopterus insectus, Callionymidae, Arnoglossus? tenuis), which are unknown from the Mediterranean while they were widespread in the Eastern Paratethys (Pobedina 1954, 1956; Suzin 1968; Strashimirov 1981b; Djafarova 2006; this study). The newcomers originating from the Eastern Paratethys are fewer in the Pannonian Basin (Onogadus simplicissimus, Palimphemus minusculoides, Arnoglossus? tenuis) than in the Pre-Carpathian Trough (Table 3). This may indeed indicate a link between the Central and Eastern Paratethys during the Konkian, chiefly with a westward-directed influx, but at a relatively low level, and very similar to the events and faunal migration directions discussed in Kovac et al. (2007). Generally, the otolith-based fish fauna of the Pannonian Basin is very diverse and rich and contains neritic associations which show an uninterrupted continuation of the early Badenian fauna. The Carpathian Foredeep and Transsylvanian Basin show a few faunal elements that are indigenous to the region and possibly represent primary endemics (Diaphus obliquus, Paratrisopterus rumanus). We conclude from these observations that the Konkian was a time of limited faunal exchange between the Central and Eastern Paratethys and that a marine connection could have persisted between the Pannonian Basin and the northern Mediterranean (Fig. 13). A good regional representation of the fish faunas of the European seas in the early Serravallian is given by the distribution patterns of the three species of the genus Palimphemus at that time (Fig. 14).
The Konkian fish fauna of the Eastern Paratethys is rich in elements that are indicative of the early onset of endemic development. In addition to the Callionymidae, Gadidae (Palimphemus minusculoides, Paratrisopterus insectus), Atherina gidjakensis and Arnoglossus? tenuis mentioned before, one may cite Alosa paulicrenata, Pontinus? aff. obrotchishtensis, Morone? bannikovi, the sciaenids Genyonemus? karagiensis and Trewasciaena suzini, Parablennius prokofievi and the first occurrence of the persistent endemic gobiid genera Neogobius and Ponticola. We have shown that the fish fauna of the Konkian (and Karaganian too) of the Eastern Paratethys shows no or only a very weak relationship to the contemporaneous faunas of the Mediterranean or Central Paratethys (Table 3). As yet, nothing is known about the fish fauna of the adjacent Indian Ocean during this phase, but it appears from the above observations that certain groups might possibly have immigrated from there. We postulate, however, that the speciation of fishes observed in the Konkian is primarily an expression of an early endemic development in the Eastern Paratethys, in particular as far as gadids, callionymids, sciaenids and gobiids are concerned. In all of these taxa, further endemic developments ensued in the Basin, as will be shown in further papers which are currently in preparation.
This continued endemic development and the lack of shared faunal elements with the Karaman Basin in SE Turkey (except for Knipowitschia suavis, see above) thus contradict the palaeogeographic reconstruction of Rögl (1999) and the discussion in Kovac et al. (2007), both of which assume that a marine connection existed between the SE Mediterranean in the vicinity of the Karaman Basin and the southern part of the Eastern Paratethys during the Konkian.
Biostratigraphical evaluation
Our knowledge of otoliths from the Tarkhanian to the Konkian of the Eastern Paratethys is still at an early stage, and much of the material described earlier requires fundamental revision. Nevertheless, it is now possible to discern the stratigraphic distributions of relevant species to some extent. Table 4 presents a summary of selected otolith-based species that are considered to be sufficiently well represented in the currently available fossil record relevant to this time interval in the Eastern Paratethys. Two events of probable stratigraphic value can be already identified. The first is the major change in faunal composition from the Chokrakrian to the Karaganian, which is best evidenced by the disappearance of the Bregmacerotidae and Gonostomatidae (Bonapartia). A second and fairly reliable marker is the first occurrence of Palimphemus minusculoides in the Konkian. Palimphemus minusculoides is a highly diagnostic species whose otoliths are easily distinguishable from the widely distributed P. anceps by outline, thickness and curvature of inner face, and collum size. Furthermore, P. minusculoides appears to be entirely restricted to the Paratethys (Fig. 14) and is one of the few species that is correlatable across the extent of the Paratethys, and probably appeared synchronously.
Conclusions and outlook
The present study covers the richest otolith collection obtained so far from any location in the Eastern Paratethys. It comes from a critical time interval in the evolution of the basin, i.e. the terminal Konkian (middle Miocene, early Serravallian), which marks the final separation of the Eastern Paratethys from the Central Paratethys and the world oceans, and sets the stage for a dynamic endemic evolution. The Konkian fish fauna of the Eastern Paratethys, as reconstructed from these otoliths, shows novel faunal elements/first occurrences in abundance, many of which are interpreted here as endemisms. However, the origin of these new faunal elements remains largely unresolved, since the correlation with temporally and environmentally equivalent faunas from the Central Paratethys is limited, with the SE Mediterranean showing a very low level of shared species that argues against a putative connection between the two during that time, while comparable data from the Indian Ocean are completely lacking.
The dominant faunal elements represented by otoliths are assigned to the families Gobiidae and Gadidae, and they are accompanied by significant accessory groups such as Atherinidae, Moronidae, Sciaenidae and Callionymidae. The Gadidae are the most common and already exhibit a strongly endemic composition unrelated to the well-known gadid faunas of northern Europe. The Gobiidae are the most species-rich and the first clear representatives of the Recent Ponto-Caspian endemics are recognisable. The Sciaenidae and Callionymidae in particular show a degree of species diversity rarely observed elsewhere in Neogene sediments in European basins, and the Sciaenidae in particular primarily represent lineages that no longer occur in European seas.
We have also attempted to revise previously described otolith-based fish faunas from the Eastern Paratethys in the course of this study, and were able to redefine some of the rarely cited species, but the taxonomic status of many others remains doubtful and will remain unresolved until the original material has been recovered and made available for review. Our current experience, however, is such that we must assume that a certain proportion of otolith type material previously recovered from salient sites can no longer be located, so the status of some “older” species may remain permanently doubtful or will require redefinition with the help of material newly collected from representative locations.
We were very fortunate that the richness of the fossil fauna collected at Karagaily enabled us to undertake a very detailed stratigraphic assessment, primarily through the identification of molluscs, foraminifera and nannoplankton. In a basin which has seen such rapid geological change as the Eastern Paratethys did, the importance of the exact stratigraphic placement of the otolith finds for the task of calibrating the data and disentangling the complex evolution of its fishes cannot be overestimated. Furthermore, a detailed analysis of the associated fauna provides valuable information for an integrated palaeoenvironmental assessment.
This study is part of a broader program of work on otoliths from the Neogene of the Eastern Paratethys which focuses on unravelling the evolutionary history of today’s Ponto-Caspian fish fauna. Drawing on collections already made, and judging from initial explorations of locations selected for future collections, we believe that a set of similarly rich and diverse otolith-based fish faunas can be retrieved for study. Emphasis will be placed on adequate stratigraphic allocation of the samples obtained, on crucial time periods in the development of the Eastern Paratethys and, wherever possible, on investigating locations representing different palaeoenvironments and different sub-basins.
Notes
Regrettably, Djafarova could not complete the final version of her monograph before she died. Fortunately, however, G. Aliev subsequently prepared her unfinished manuscript for publication.
References
Ali-Zadeh, A.A. 1974. Sarmat Azerbaidzhana (Sarmatian of Azerbaijan). Moskva: Nedra. (in Russian).
Andrusov, N.I. 1915. Materialy dlia geologii Zakaspiyskoy oblasti Mangyshlaka (Materials for the geology of the Zacaspiy region). Trudy Aralo-Kaspiyskoy ekspedicii 8: 1–456. (in Russian).
Andrusov, N.I. 1917. Konskiy gorizont (foladovie plasty) (Konkian horizon (Pholada level)). Trudy geologicheskogo i mineralogicheskogo muzeia Akademii nauk 2(6): 167–261. (in Russian).
Anistratenko, V. 2005. Morphology and taxonomy of Late Badenian to Sarmatian Mohrensternia (Gastropode: Rissooidea) of the Central Paratethys. Acta Geologica Polonica 55(4): 371–392.
Bajarunas, M.V. 1917. Bezottochnie vpadiny uzhnogo Mangyshlaka. (Inland depressions of South Mangyshlak). Izvestiya Rossiyskogo geograficheskogo obschestva 53: 1–44. (in Russian).
Bannikov, A.F. 1989. The first discovery of scale-bearing blennies (Teleostei) in the Sarmatian of Moldavia. Paleontological Journal 23(2): 64–70.
Bannikov, A.F. 1993. A new species of bass (Teleostei, Moronidae) from the Sarmatian of Moldova. Paleontological Journal 27(1): 43–50.
Bannikov, A.F. 2006. About the Miocene Centracanthidae (Teleostei, Perciformes) from the Eastern Paratethys. Paleontological Journal 40(1): 100–102.
Bannikov, A.F. 2008. Fishes of the lower Sarmatian (Volhynian) from locations near Naclavcha (northern Moldavia, Pri-Dnjestr). Bulletin National Museum prirody i etnographii Moldavy, Estestvennye nauki 8(21): 110–114.
Bannikov, A.F. 2009. On Early Sarmatian fishes from the Eastern Paratethys. Paleontological Journal 43(5): 569–573.
Bannikov, A.F. 2010. Fossil vertebrates of Russia and adjacent countries: Acanthopterygian fishes (Teleostei, Acanthopterygii). Moscow: Geos (in Russian).
Bannikov, A.F., G. Carnevale, and W. Landini. 2009. A new Early Miocene genus of the family Sciaenidae (Teleostei, Perciformes) from the eastern Paratethys. Comptes Rendus Paleoevol 8: 535–544.
Barbot-de-Marni, N.P. 1889. Cherez Mangyshlak i Ustiurt v Turkmenistan. (Through Mangyshlak and Ustiurt to Turkmenistan). Trudy Aralo-Kaspiyskoy ekspedicii 6: 1–111. (in Russian).
Baykina, E.M. 2013. A revision of Clupea doljeana Kramberger and Sarmatella vukotinovici (Kramberger) (Pisces, Clupeidae) from the Sarmatian of Croatia. Paleontological Journal 47: 523–532.
Béarez, P., and W. Schwarzhans. 2013. Robaloscion, a new genus for Sciaena wieneri Sauvage, 1883 (Teleostei, Sciaenidae) from the southeastern Pacific, with clarification of the taxonomic status of Sciaena starksi Evermann and Radcliffe, 1917. Cybium 37: 273–279.
Bogatshov, V.V. 1929. Whale and fish remains in diatom deposits of Apsheronski Peninsula. Azerbaidjanski neftyanoe khozyaistvo 77: 33–39. (in Russian).
Bogatshov, V.V. 1933. Materialy po izucheniyu tretichnoiy ikhtiofauny Kavkaza. (Materials on the study of Tertiary ichthyofauna of Caucasus). Trudy Azerbaijanskogo Neftianogo Instituta. Geologicheskoe otdelenie 15: 1–62. (in Russian).
Bogatshov, V.V. 1938. Fishes from the Pontian deposits of Kirovski region. Trudi Azerbaidjanski philosophicheski Annali SSSR 9: 36–60. (in Russian).
Bogatshov, V.V. 1942. Materialy k ikhtiofaune meoticheskogo moria. (Materials about ichthyofauna of Meotian see). Doklady Akademii Nauk SSSR 37(2): 89–92. (in Russian).
Bogatshov, V.V. 1955. Amvrosievskie nizhnesarmatskie ryby (Lower Sarmatian fishes from Amvrosievka). Doklady Akademii Nauk SSSR 105(3): 570–573. (in Russian).
Bogdanovich, A.K. 1965. Stratigraficheskoe i fatsialnoe raspredelenie foraminifer v miotsene Zapadnogo Predkavkaz’ya i voprosi ih genezisa (Stratigraphic and facies distribution of foraminifera in the Miocene of Western Ciscaucasia and questions of their genesis). Trudy Krasnodarskogo filiala Vsesoyuznogo neftegazovogo nauchno-issledovatelskogo instituta 16: 300–350. (in Russian).
Bratishko, A., and M. Udovichenko. 2013. Fish otoliths from the Early Oligocene of Mangyshlak, Kazakhstan. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 270(2): 195–208.
Brzobohaty, R. 1980. Die Otolithen-Fauna des oberen Badenien von Strahovice (Miozän, Opava-Gebiet). Casopis Slezskeho Muzea Opava (A) 29:133–144. (in Czech, German Summary).
Brzobohaty, R. 1992. Otolithen aus dem Obermiozän, Pontien, des Wiener Beckens (Götzendorf und Stixneusiedl, NÖ). Annalen des Naturhistorischen Museums in Wien, serie A 94: 1–6.
Brzobohaty, R. 1995. Macrouridae (Teleostei, Otolithen) im Oligozän und Miozän der Zentralen Paratethys und ihre paläogeographische Bedeutung. Jahrbuch der Geologischen Bundesanstalt Wien 138: 615–637.
Brzobohaty, R., and J. Stancu. 1974. Die Fischfauna des Sarmatien s. str. In Chronostratigraphie und Neostratotypen Miozän der zentralen Paratethys. VI, M 5 Sarmatien, eds. Papp, A., F. Marinescu and J. Senes, 492–515. Bratislava: Veda.
Brzobohaty, R., D. Nolf, and O. Kroupa. 2007. Fish otoliths from the Middle Miocene of Kienberg at Mikulov, Czech Republic, Vienna Basin: their paleoenvironmental and paleogeographic significance. Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre 77: 167–196.
Bugrova, E.M., V.I. Gladkov, T.V. Dmitrieva, L.S. Nevzorova, and B.C. Sokolov. 2005. Prakticheskoe rukovodstvo po mikrofaune, 8: Foraminiferi kainozoya (Guidebook of microfauna, 8: Cenozoic Foraminifera). St. Petersburg: VSEGEI. (in Russian).
Carnevale, G., A.F. Bannikov, W. Landini, and C. Sorbini. 2006. Volhynian (Early Sarmatian s.l.) fishes from Tsurevsky, North Caucasus (Russia). Journal of Paleontology 80: 684–699.
Carnevale, G., and M. Harzhauser. 2013. Middle Miocene rockling (Teleostei, Gadidae) from the Paratethys (St. Margarethen in Burgenland, Austria). Bulletin of Geosciences 88(3): 609–620.
Carnevale, G., M. Harzhauser, and O. Schultz. 2012. The Miocene gadid fish Palimphemus anceps Kner, 1862: a reappraisal. Geodiversitas 34(3): 625–643.
Chalilov, D.M. 1946. Kakraganskie i konkskie sloi severo-vostochnogo Azerbaijana (Karaganian and Konkian Beds of northeastern Azerbaijan). Doklady Akademii Nauk Azerbaijanskoy SSR 2(7): 275–277. (in Russian).
Daniltchenko, P.G. 1986. Morskie i solonovatovodnie ryby Paratetisa (Marine and brackish water fishes of the Paratethys). In Stratigrafia SSSR. Neogenovaya sistema. Vol. 2, eds. Muratov, M.V., and L.A. Nevesskaia, 302–305. Moscow: Nedra (in Russian).
Djafarova, J.D. 2006. Otolity neogena Azerbaidjana (Neogene otoliths of Azerbaijan), 167. Baku: Nafta (in Russian).
Djanelidze, O.I. 1970. Foraminiferi nizhnego i srednego miotsena Gruzii (Foraminifera from the Late and Middle Miocene of Georgia). Tbilisi: Metsniereba. (in Russian).
Eichwald, E. 1834. Reise auf dem Caspischen Meere und in den Caucasus unternommen in den Jahren 1825–1826. Stuttgart, Tubingen: Verlag der J.G. Cotta’schen Buchhandlung.
Eschmeyer, W.N., and J.D. Fong. 2013. Species of fishes by family/subfamily. http://research.calacademy.org/research/ichthyology/catalog/speciesbyfamily.asp. Accessed 31 Oct 2014.
Fedotov, V.F. 1976. Treskovye paleogen-neogenovyh otlozheniy SSSR. (Codfishes from Palaeogene-Neogene deposits of USSR). Trudy Paleontologicheskogo Instituta Akademii Nauk SSSR 157: 1–83. (in Russian).
Francisco, S.M., J.I. Robalo, S. Stefanni, A. Levy, and V.C. Almada. 2014. Gaidropsarus (Gadidae, Teleostei) of the North Atlantic Ocean: a brief phylogenetic review. Journal of Fish Biology 85: 473–487.
Gaemers, P.A.M., and W. Schwarzhans. 1973. Fisch-Otolithen aus dem Pliozän von Antwerpen (Belgien) und Ouwerkerk (Niederlande) und aus dem Plio-Pleistozän der Westerschelde (Niederlande). Leidse Geologische Mededelingen 49: 207–257.
Härkönen, T. 1986. Guide to the otoliths of the bony fishes of the northeast Atlantic. Hellerup: Danbiu ApS.
Harzhauser, M., O. Mandic, and M. Zuschin. 2003. Changes in Paratethyan marine molluscs at the Early/Middle Miocene transition: diversity, palaeogeography and palaeoclimate. Acta Geologica Polonica 53(4): 323–339.
Harzhauser, M., W.E. Piller, and F.F. Steininger. 2002. Circum-Mediterranean Oligo-Miocene biogeographic evolution—the gastropods’ point of view. Palaeogeography, Palaeoclimatology, Palaeoecology 183: 103–133.
Hoestlandt, H. 1991. The freshwater fishes of Europe, Clupeidae, Anguillidae. Wiesbaden: AULA.
Hohenegger, J., S. Ćorić, and M. Wagreich. 2014. Timing of the Middle Miocene Badenian stage of the Central Paratethys. Geologica Carpathica 65(1): 55–66.
Holec, P. 1975. Fisch-Otolithen aus dem Baden (Miozän) des nördlichen Teils des Wiener Beckens und der Donau-Tiefebene. Geologica Carpathica 26(2): 253–266.
Ilyina, L.B. 1993. Opredelitel’ morskih srednemiotsenovih gastropod Yugo-Zapadnoy Evrasii (Handbook for identification of the marine Middle Miocene gastropods of Southwestern Eurasia). Trudy Paleontologicheskogo instituta 255: 1–151. (in Russian).
Ilyina, L.B. 2000. On the regional Konkian stage (Middle Miocene) in the Eastern Paratethys. Stratigraphy and Geological Correlation 8(4): 59–64.
Ilyina, L.B. 2006. Morphogenesis of Rissoidae in the inland basins of the Eastern Paratethys. Paleontological Journal 4: 54–63.
Ilyina, A.P. and V.S. Utkin. 1963. Neogenovye otlozheniya Mangyshlaka. (Neogene deposits of Mangyshlak) Trudy Vsesouznogo nauchno-issledovatelskogo geologo-razvedochnogo instituta (VNIGRI) 218: 425–430. (in Russian).
Jacobs, P., and K. Hoedemakers. 2013. The round goby Neogobius melanostomus (Pallas, 1814) (Perciformes: Gobiidae), an invasive species in the Albert Canal (Belgium). Belgian Journal of Zoology 143(2): 148–153.
Janssen, A.W. 2012. Validation of holoplanktonic molluscan taxa from the Oligo-Miocene of the Maltese Archipelago, introduced in violation with ICZN regulations. Cainozoic Research 9: 189–191.
Jerzmanska, A. 1962. Szcatki ryb kostnoszkieletowych z Miocenu Górnego Slaska. Acta Palaeontologica Polonica 7: 235–247.
Klein, L.N. 1960. Resultaty issledovaniya otolitov ryb iz pliocenovyh i postpliocenovyh otlozheniy Prikurinskoi nizmennosti Azerbaidjana. (Results of fish otoliths research in Pliocene and Postpliocene deposits of the Prikurin lowland). Trudy AzNII no dobytche nefti 10: 101–121. (in Russian).
Koken, E. 1884. Über Fisch-Otolithen, insbesondere über diejenigen der norddeutschen Oligocän-Ablagerungen. Zeitschrift der deutschen geologischen Gesellschaft 36: 500–565.
Koken, E. 1891. Neue Untersuchungen an tertiären Fisch-Otoloithen II. Zeitschrift der deutschen geologischen Gesellschaft 43: 77–170.
Kolesnikov, V.P. 1935. Paleontologiya SSSR. Sarmatskie molluski (Palaeontology of USSR. Sarmatian molluscs), 10 (2). Moscow: Izdatelstvo Akademii Nauk SSSR (in Russian).
Kolesnikov, V.P., B.P. Zhizhchenko, and A.G. Eberzin. 1940. Stratigrafia SSSR, 12: Neogen SSSR (Stratigraphy of USSR, 12: Neogene USSR). Moscow: Izdatelstvo Akademii Nauk SSSR. (in Russian).
Kovac, M., A. Andreyeva-Grigorovich, Z. Bajraktarevic, R. Brzobohaty, S. Filipescu, L. Fodor, M. Harzhauser, A. Nagymarosy, N. Osczcypko, D. Pavelic, F. Rögl, B. Saftic, L. Sliva, and B. Studencka. 2007. Badenian evolution of the Central Paratethys Sea: paleogeography, climate and eustatic sea-level changes. Geologica Carpathica 58(6): 579–606.
Kowalke, T., and M. Harzhauser. 2004. Early ontogeny and palaeoecology of the Mid-Miocene rissoid gastropods of the Central Paratethys. Acta Palaeontologica Polonica 49: 111–134.
Kramberger-Gorganovic, D. 1883. Die jungtertiäre Fischfauna Croatiens. Beiträge zur Paläontologie Österreich-Ungarns und des Orients 3: 65–85.
Krasheninnikov, V.A. 1959. Foraminiferi (Foraminifera). In Atlas srednemiotsenovoy fauni Severnogo Kavkaza i Kryma (Atlas of the Middle Miocene fauna from North Caucasus and Crimea), ed. B.P. Zhizhchenko, 15–103. Moskva: Gostoptehizdat. (in Russian).
Landau, B.M., M. Harzhauser, Y. Islamoglu, and C. Marques da Silva. 2013. Systematics and palaeobiogeography of the gastropods of the middle Miocene (Serravallian) Karaman Basin, Turkey. Cainozoic Research 11–13: 1–584.
Laskarev, V. 1903. Fauna Buglovskih sloev Volhini (Fauna of the Buglow Beds of Volhyn). Memores du comite geologique, Nouvelle serie 5: 1–148. (in Russian).
Liverovskaya, E.V. 1935. Fauna konkskogo gorisonta gory Dubrovoy (Severniy Kavkaz) (Fauna of the Konkian horizon of Dubrowaia Mt. (North Caucasus)). Trudy neftyanogo geologo-razvedochnoro instatuta. Seria A 44: 1–35. (in Russian).
Liverovskaya, E.V. 1960. Tretichnye otlozheniia Mangyshlaka (Tertiary deposits of Mangyshlak). Trudy Vsesouznogo neftianogo nauchno-issledovatelskogo geologorazvedochnogo instituta 151: 1–144. (in Russian).
Lombarte, A., O. Chic, V. Parisi-Baradad, R. Olivella, J. Piera, and E. Garcia-Ladona. 2006. A web-based environment for shape analysis of fish otoliths. The AFORO database. Scientia Marina 70(1): 147–152.
Liuliev, Y.B. 1967. Ostracody i stratigrafia miocenovyh otlozheniy Yuzhnoy Ukrainy (Ostracods and stratigraphy of the Miocene of Southern Ukraine). MS dissertation. Kiev: Kand. Geol.-Min. Nauk (in Russian).
Martini, E. 1971. Standard Tertiary and Quaternary calcareous nannoplankton zonation. In: Proceedings of the II Planktonic Conference, ed. Farinacci, A., 739–785. Roma: Tecnoscienza.
Medvedeva, D.A., P.A. Sorokina, V.P. Vasil’eva, N.V. Chernovab, and E.D. Vasil’eva. 2013. Reconstruction of phylogenetic relations of Ponto-Caspian gobies (Gobiidae, Perciformes) based on mitochondrial genome variation and some problems of their taxonomy. Journal of Ichthyology 53(9): 687–698.
Menner, V.V. 1948. Sciena iz kimmeriyskih otlozheniy Kerchenskogo poluostrova (Sciaena from the Kimmerian deposits of the Kerch Peninsula). Voprosy teoreticheskoy i prikladnoy geologii 5: 23–31. (in Russian).
Merklin, R.L., and L.A. Nevesskaia. 1955. Opredelitel dvustvorchatyh molliuskov miocena Turkmenii i Zapadnogo Kazakhstana (Identification guide of mollusks of Turkmenia and Western Kazakhstan). Trudy Paleontologicheskogo instituta Akademii Nauk SSSR 59: 1–115. (in Russian).
Merklin, R.L. 1953. Etapy razvitia konkskogo basseina v miocene na uge SSSR (The stages of Konkian basin development in the Miocene of the southern USSR). Bulletin Moskovskogo obchsestva ispytateley prirody, otdelenie geologii 28(3): 89–91. (in Russian).
Merklin, R.L. 1954. O konkskom gorizonte severnogo poberezhia Kara-Bogaz-Gola i o prisutstvii v nem bezzamkovyh brachiopod Lingula (About the Konkian horizon of the northern shore of Kara-Bogaz-Gol and the presence of the ecardinal brachiopods Lingula). Doklady Akademii Nauk SSSR 95(1): 155–158. (in Russian).
Merklin, R.L., V.G. Morozova, and A.S. Stoliarov. 1960. O biostratigrafii maikopskih otlohzeniy Uzhnogo Mangyshlaka (About biostratigraphy of the Maikop deposits of South Mangyshlak). Doklady Akademii Nauk SSSR 133(3): 653–656. (in Russian).
Miller, P.J. 2003. The freshwater fishes of Europe, Mugilidae, Atherinidae, Atherinopsidae, Blenniidae, Odontobutidae, Gobiidae 1. Wiesbaden: AULA.
Miller, P.J. 2004. The freshwater fishes of Europe, Gobiidae 2. Wiesbaden: AULA.
Müller, A. 1999. Ichthyofaunen aus dem atlantischen Tertiär der USA. Leipziger Geowissenschaften 9–10: 1–360.
Nelson, J.S. 2006. Fishes of the world, 4th ed. Hoboken: John Wiley and Sons.
Nevesskaya, L.A., K.G. Bogdasaryan, A.K. Bogdanovich, et al. 1986. Stratigrafia SSSR. Neogenovaya sistema, 1 (Stratigraphy of the USSR. Neogene, 1). Moskva: Nedra (in Russian).
Nevesskaya, L.A., I.A. Goncharova, N.P. Paramonova, S.V. Popov, E.V. Babak, K.G. Bagdasaryan, and A.A. Voronina. 1993. Opredelitel’ miotsenovih dvustvorchatih molluskov Yugo-Zapadnoy Evrazii (Handbook for identification of the bivalvia of Southwestern Eurasia). Trudy Paleontologicheskogo instituta 247: 1–412. (in Russian).
Nolf, D. 1981. Révision des Types d’Otolithes de Poissons Fossiles décrits par R. Schubert. Verhandlungen der geologischen Bundesanstalt 2: 133–183.
Nolf, D. 1985. Handbook of paleoichthyology, Volume 10, Otolithi piscium. Munich: Dr. Friedrich Pfeil.
Nolf, D., and R. Brzobohaty. 2009. Lower Badenian fish otoliths of the Styrian and Lavanttal basins, with a revision of Weinfurter’s type material. Annalen des Naturhistorischen Museums in Wien—Serie A (Mineralogie und Petrographie, Geologie und Paläontologie, Archäozoologie, Anthropologie und Prähistorie) 111: 323–356.
Nolf, D. 2013. The diversity of fish otoliths, past and present. Brussels: Royal Belgian Institute of Natural Sciences.
Nolf, D., and E. Steurbaut. 1983. Revision des otolithes de teleosteens du Tortonien stratotypique et de Montegibbio (Miocene Superieur de l’Italie septentrionale). Mededelingen van de Werkgroep voor Tertiaire en Kwartaire Geologie 20(4): 143–197.
Paghida, N. 1962. Otolite din Buglovianul Podisului Moldovenesc. Analele Stiintifice ale Universitatii Alexandru Ioan Cuza din Iasi, Seria Geologie (Sect. 2) 8: 13–20.
Paghida-Trelea, N. 1969. Micro-si mezofauna de Apa indulcita din Bessarabianul Podisului central Moldovenesc. Analele Stiintifice ale Universitatii Alexandru Ioan Cuza din Iasi, (Sect. 2b) 8: 113–120.
Paghida-Trelea, N.T. Simionescu, and L. Olaru. 1967. Contributii la studiul micropaleontologie al Sarmatianului din imprejurimile Harlaului. Analele Stiintifice ale Universitatii Alexandru Ioan Cuza din Iasi, Seria noua (Sect. 2b) 13: 83–99.
Pana, I. 1965. Otolitele pliocene din regiunea de Curbura a Carpatilor. Studii si cercetari de geologie, geofizica, geografie. Seria Geologie 1(10): 3–14.
Pana, I. 1977. Problems arisen by the appearance of the new otoliths species and their stratigraphical implications, examples of the Dacian otoliths from S. Dobrudja. Analele Universitatii Bucuresti. Seria geologie 26: 105–124.
Pana, I. 1982. Otolithes Dacien superieur (Parascovien) d’oltenie. Analele Universitatii Bucuresti. Seria geologie 31: 33–42.
Pana, I. 1995. Otolithes daciens. In Chronostratigraphie und Neostratotypen. 9, Pliozän PL 1, Dazien, eds. Marinescu, F. and I. Papaianopol, 427–445. Bucharest: Acad. Romane.
Papp, A., I. Cicha, and J. Seneš. 1978. M4, Badenien (Moravien, Wielicien, Kosovien). Chronostratigraphie und Neostratotypen, 4:1–594. Bratislava: VEDA.
Piller, W.E., M. Harzhauser, and O. Mandic. 2007. Miocene Central Paratethys stratigraphy—current status and future directions. Stratigraphy 4(2/3): 151–168.
Perch-Nielsen, K. 1985. Cenozoic calcareous nannofossils. In Plankton stratigraphy, ed. Bolli H.M., J.B. Saunders, and K. Perch-Nielsen, 427–554. Cambridge: Cambridge University Press.
Pisera, A. 1996. Miocene reefs of the Paratethys: a review. Concepts in Sedimentology and Paleontology 5: 97–104.
Pobedina, V.M. 1954. Iskopaemye otolity ryb miocenovyh otlozheniy Azerbaijana i ih stratigraficheskoe znachenie (Fossil fish otoliths from the Miocene deposits of Azerbaijan and their stratigraphical significance). Izvestia Akademii Nauk Azerbaidjanskoy SSR 10: 23–37. (in Russian).
Pobedina, V.M., A.G. Voroshilova, O.I. Rybina, and Z.V. Kuznetsova. 1956. Spravochnik po mikrofaune sredne—i verkhnemiocenovykh otlozhenii Azerbaidjana (Handbook about microfauna Middle and Upper Miocene deposits of Azerbaijan). Baku: Azerbaidjanskoe gosudarstvennoe izdatelstvo neftyanoi nauchno-tekhnicheskoy literatury. (in Russian).
Popov, S.V., I.G. Shcherba, L.B. Ilyina, L.A. Nevesskaya, N.P. Paramonova, S.O. Khondkarian, and I. Magyar. 2006. Late Miocene to Pliocene paleogeography of the Paratethys and its relation to the Mediterranean. Palaeogeography, Palaeoclimatology, Palaeoecology 238: 91–106.
Prokofiev, A.M. 2002. New and little known Perciform fishes from the Upper Oligocene–Lower Miocene boundary deposits of the Caucasus (Osteichthyes, Perciformes). Zoosystematika Rossica 11(1): 209–217.
Prokofiev, A.M. 2004. On the attribution of Paratrisopterus avus to the genus Gadiculus (Gadiformes: Gadidae). Journal of Ichthyology 44(6): 412–417.
Prokofiev, A.M. 2005. Revision of protacanthopterygii (Osteichthyes: Euteleostei) from Cenozoic marine deposits of the Caucasus and Turkmenistan: I. The orders Argentiniformes and Salmoniformes s.str. (sensu Johnson, Patterson, 1996) in the Oligocene–Miocene of the Caucasus. Journal of Ichthyology 45(1): 1–15.
Prokofiev, A.M. 2007. Meso- and bathypelagic fishes of the orders Stomiiformes and Myctophiformes in the Paleogene-Neogene of Russia and adjacent territories. Modern Paleontology: Classical and New Methods 2007: 45–58.
Radwanska, U. 1992. Fish otoliths in the Middle Miocene (Badenian) deposits of southern Poland. Acta Geologica Polonica 42(3–4): 141–328.
Radionova, E., L. Golovina, N. Filippova, V. Trubikhin, S. Popov, I. Goncharova, Y. Vernigorova, and T. Pinchuk. 2012. Middle-Upper Miocene stratigraphy of the Taman Peninsula, Eastern Paratethys. Central European Journal of Geosciences 4(1): 188–204.
Rado, G. 1965. Otolite din depozitele tortoniene de la Costeiul de Sus. Analele Universitatii Bucuresti, Seria stiintele naturii, Geologie-geografie 14: 55–71.
Rado, G. 1968. Etude des otolithes sarmatiens de Copacel-Chijic (Bassin du Crisul Repede). Travaux du museum d’histoire naturelle “Grigore Antipa” 8(2): 581–585.
Rado, G. 1969. Grupe noi de organisme in fauna Tortoniana de la Buituri. Studii si cercetari de geologie, geofizica, geografie. Seria geologie 14(1): 189–204.
Rado, G. 1971. Otolite din deposizitele miocene din forajele de la Islaz. Buletinul Societatii de Stiinte geologice din R S. Romania 13: 180–194.
Rado, G. 1981. Otolite din depositele langhiene (Badenian inferior) de la Lapugiul de Sus. Analele Universitatii Bucuresti, Seria geologie 30: 29–46.
Reichenbacher, B. 1993. Mikrofaunen, Paläogeographie und Biostratigraphie der miozänen Brack- und Süßwassermolasse in der westlichen Paratethys unter besonderer Berücksichtigung der Fisch-Otolithen. Senckenbergiana Lethaea 73(2): 277–374.
Reichenbacher, B., and M. Weidmann. 1992. Fisch-Otolithen aus der oligo-miozänen Molasse der West-Schweiz und der Haute-Savoie (Frankreich). Stuttgarter Beiträge zur Naturkunde Serie B (Geologie und Paläontologie) 184: 1–83.
Rögl, F. 1998. Palaeogeographic considerations for Mediterranean and Paratethys seaways (Oligocene to Miocene). Annalen des Naturhistorischen Museums in Wien 99A: 279–310.
Rögl, F. 1999. Mediterranean and paratethys. Facts and hypotheses of an Oligocene to Miocene paleogeography (short overview). Geologica Carpathica 50(4): 339–349.
Rögl, F., and F.F. Steininger. 1983. Vom Zerfall der Tethys zu Mediterran und Paratethys. Annalen des Naturhistorischen Museum in Wien 85: 135–163.
Rückert-Ülkümen, N. 1996. Weitere Beiträge zur Otolithenfauna von Avcılar W Küçükçekmece See (Thrakien, Türkei). Mitteilungen der Bayerischen Staatssammlung für Paläontologie und Historische Geologie 36: 117–133.
Rückert-Ülkümen, N., and O. Kaya. 1993. Neue Beiträge zur Tertiär-Stratigraphie und Otolithenfauna der Umgebung von Istanbul (Küçükçekmece- und Büyükçekmece See), Türkei. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und Historische Geologie 33: 51–89.
Schubert, R.J. 1902. Die Fischotolithen des österr.-ungar. Tertiärs. I. Jahrbuch der kaiserlich-königlichen geologischen Reichsanstalt 51: 301–316.
Schubert, R.J. 1906. Die Fischotolithen des österr.-ungar. Tertiärs. III. Jahrbuch der kaiserlich-königlichen geologischen Reichsanstalt 56: 623–706.
Schubert, R.J. 1912. Die Fischotolithen der ungarischen Tertiärablagerungen. Mitteilungen aus dem Jahrbuch der königlichen ungarischen geologischen Reichsanstalt 20(3): 115–139.
Schultz, O. 2013. Catalogus Fossilium Austriae. Band 3 Pisces. Vienna: Verlag der Österreichischen Akademie der Wissenschaften.
Schwarzhans, W. 1978. Otolith-morphology and its usage for higher systematical units, with special reference to the Myctophiformes s.l. Mededelingen Werkgroep Tertiaire en Kwartaire Geologie 15: 167–185.
Schwarzhans, W. 1993. Piscium Catalogus. Part Otolithi Piscium. Volume 1. A comparative morphological treatise of recent and fossil otoliths of the family Sciaenidae (Perciformes). Munich: Dr. Friedrich Pfeil.
Schwarzhans, W. 1999. Piscium Catalogus. Part Otolithi Piscium. Volume 2. A comparative morphological treatise of recent and fossil otoliths of the order Pleuronectiformes. Munich: Dr. Friedrich Pfeil.
Schwarzhans, W. 2010. The otoliths from the Miocene of the North Sea Basin. Weikersheim/Leiden: Backhuys/Margraf.
Schwarzhans, W. 2012. Fish otoliths from the Paleocene of Bavaria (Kressenberg) and Austria (Kroisbach and Oiching-Graben). Palaeo Ichthyologica 12: 1–88.
Schwarzhans, W. 2013. A comparative morphological study of the Recent otoliths of the genera Diaphus, Idiolychnus and Lobianchia (Myctophidae). Palaeo Ichthyologica 13: 41–82.
Schwarzhans, W. 2014a. Synonymisation of the skeleton-based Palimphemus anceps Kner, 1862 and the otolith-based Colliolus sculptus (Koken, 1891) (Pisces, Teleostei, Gadidae). Cainozoic Research 14(1): 9–16.
Schwarzhans, W. 2014b. Otoliths from the middle Miocene (Serravallian) of the Karaman Basin, Turkey. Cainozoic Research 14(1): 35–69.
Semenenko, V.N., A.S. Andreyeva-Grigorovich, N.V. Maslun, and S.A. Luljeva. 2009. Priamaia korreliaciia neogena Vostochnogo Paratetisa s mezhdunarodnoj okeanicheskoj shkaloj po planktonnym mikrofossiliiam (The direct correlation of the Neogene of the Eastern Paratethys with the International Oceanic Scale by planktonic microfossils). Geologichniy zhurnal 4: 9–27. (in Russian).
Smale, M.J., G. Watson, and T. Hecht. 1995. Otolith atlas of southern African marine fishes. Ichthyological Monographs. Grahamstown: J.L.B. Smith Institute of Ichthyology.
Smigielska, T. 1966. Otolity Ryb z Tortonu Południowej Polski. Roczink Polskiego Towarzystwa Geologicznego—Annales de la Societé geologique de Pologne 36(3): 205–275.
Sokolow, N. 1899. Sloi s Venus konkensis na r. Konke (Venus konkensis beds at the Konka River). Memores du comite geologique 9(5): 1–96 (in Russian).
Stancheva, M. 1990. Upper Miocene ostracods from Northern Bulgaria. Sofia: Bulgarian Academy of Sciences.
Stancu, J. 1969. Otolitele sarmatiene de la Soceni (Banat-Romania). Dari de Seama ale sedintelor, Paleontologie 3: 5–24.
Strashimirov, B. 1972. Otolity ot tarkhana na Severoistochna Bulgaria. Annuaire de l’Ecole Supérieure des Mines et de Geologie, Sofia 18(2): 301–313.
Strashimirov, B. 1980. Otolithes du Tchokrakien de la Bulgarie nord-orientale. Geologica Balcanica 10(2): 61–70.
Strashimirov, B. 1981a. Otolity ot karagana na Severoistochna Bulgaria. Palaeontology, Stratigraphy and Lithology 14: 19–28.
Strashimirov, B. 1981b. Otolity ot konka na Severoistochna Bulgaria. Palaeontology, Stratigraphy and Lithology 15: 52–65.
Strashimirov, B. 1982. Otolity ot badena na Bulgaria. Palaentology, Stratigraphy and Lithology 17: 13–37.
Strashimirov, B. 1984. Otolity ot dolnia sarmat na Severnaia Bulgaria. Palaentology, Stratigraphy and Lithology 20: 15–41.
Strashimirov, B. 1985a. Otolity ot srednia sarmat na Severnaia Bulgaria. Annual of the Highest Institute of Mining and Geology Sofia 31: 7–20.
Strashimirov, B. 1985b. Otolity ot gornia sarmat na Severozapadna Bulgaria. Annual of the Highest Institute of Mining and Geology Sofia 31: 21–36.
Svetovidov, A.N. 1986a. Gadidae. In Fishes of the North-eastern Atlantic and the Mediterranean, ed. Whitehead, P.J.P., M.-L. Bauchot, J.-C. Hureau, J. Nielsen, and E. Tortonese, 680–710. Paris: UNESCO.
Svetovidov, A.N. 1986b. Review of the three-bearded rocklings of the genus Gaidropsarus Rafinesque, 1810 (Gadidae) with description of a new species. Voprosy Ikhtiologii 26(1): 3–23.
Svichenskaya, A.A. 1957. Noviy vid kefali nizhnego miocena Kavkaza (New species of Mugil from the Lower Miocene of the Caucasus). Materialy k “Osnovam paleontologii” 1: 61–62 (in Russian).
Svichenskaya, A.A. 1959. Kefal iz sarmata Moldavii (Mugil from the Sarmatian of Moldavia). Paleontologicheskiy zhurnal 1: 98–99. (in Russian).
Svichenskaya, A.A. 1973. Iskopaemye kefaleobraznie SSSR (Fossil Mugiliformes of USSR). Trudy Paleontologicheskogo Instituta Akademii Nauk SSSR 138: 1–63. (in Russian).
Suzin, A.V. 1956. Ostracodi tretichnich otlozheniy Severnogo Predkavkazia (Ostracods of the Tertiary of North Ciscaucasus). Grozniy: Groznensky neftyanoi institut Gostoptehizdat. (in Russian).
Suzin, A.V. 1968. Otoliths. In Micropaleontologicheskie metody stratigraficheskih postroeniy v neftegazonosnyh oblastiah (Micropalaeontological methods of stratigraphical research in the oil-and-gas bearing region), ed. Zhizhchenko, B.P., 74–77. Moscow: Nedra. (in Russian).
Sytchevskaya, E.K., and A.M. Prokofiev. 2007. A dragonet (Perciformes: Callionymidae) from the Middle Miocene of southern Russia. Voprosy Ikhtiologii 47(9): 750–756.
Sytchevskaya, E.K., and A.M. Prokofiev. 2010. On finding of monkeyfishes (Lophiidae) in the lower Miocene of Transcaucasia. Voprosy Ikhtiologii 50(2): 274–279.
Tracey, S. 2014. Notes on Nolf’s nomenclatural system. In: Book reviews—“The diversity of fish otoliths, past and present”, by Dirk Nolf. Cainozoic Research 14(1): 71–72.
Tuset, V.M., A. Lombarte, and C.A. Assis. 2008. Otolith atlas for the western Mediterranean, north and central eastern Atlantic. Scientia Marina 72(S1): 7–198.
Vereshagin, V.N. and L.V. Mironova. 1982. Stratigraficheskiy slovar SSSR. Paleogen, Neogen, Chetvertichnaya sistema (Stratigraphic dictionary of the USSR. Paleogene, Neogene, Quaternary). Leningrad: Nedra (in Russian).
Vernigorova, J.V. 2009. Karagansky i konksky regioyarusi Vostochnogo Paratetisa: voprosi ih obyoma i stratigraficheskoy samostoyatelnosti (The Karaganian and Konkian regional stages of the Eastern Paratethys: problems of their stratigraphic range and validity). Geologichniy zhurnal 2: 34–47. (in Russian).
Voroshilova, A.G. 1957. Stratigrafia i fauna ostrakod miotsenovih otlozheniy Kobistana (Stratigraphy and ostracod fauna of the Miocene of Kobistan). Avtoreferat. Baku: Kand. Geol.-Min. Nauk, Akademiya nauk Azerbaidzhana (in Russian).
Weiler, W. 1942. Die Otolithen des rheinischen und nordwestdeutschen Tertiärs. Abhandlungen des Reichsamts für Bodenforschung, Neue Folge 206: 5–140.
Weiler, W. 1943. Die Otolithen aus dem Jungtertiär Süd-Rumäniens; 1. Buglow und Sarmat. Senckenbergiana 26(1/3): 87–115.
Weiler, W. 1949. Die Otolithen aus dem Jung-Tertiär Süd-Rumäniens; 1b. Ergänzende Tafeln zu den Otolithen des Buglow und Sarmat. Senckenbergiana 30(4/6): 291–293.
Weiler, W. 1950. Die Otolithen aus dem Jung-Tertiär Süd-Rumäniens; 2. Mittel-Miozän, Torton, Buglow und Sarmat. Senckenbergiana 31(3): 209–258.
Zhizhchenko, B.P. 1936. Paleontologiya SSSR. Chokrakskie molluski. (Palaeontology of USSR. Tschokrakian molluscs), 10 (3). Moscow: Academy of Sciences of USSR (in Russian).
Acknowledgments
We would like to thank N. Udovichenko (LTSNU, Luhansk) for his participation in field works and the scientific support of the career of the senior author, Y. Bunyatov (Aktay) for his support with field works in Mangyshlak, E. Vasilyeva (ZMMGU, Moscow) for her support in providing access to otolith extraction for many Ponto-Caspian Gobiidae to W. Schwarzhans, which proved to be invaluable for comparisons of the fossil gobiid otoliths from Karagaily. A. Bannikov (PIN, Moscow) and A. Prokofiev (SIEE, Moscow) gave support and valuable advice about fossil skeleton-based fishes from the Miocene of the Eastern Paratethys and certain otoliths present in them in situ. S. Strashimirov (UMG, Sofia) provided undescribed otoliths collected by his father during the 1970s and 1980s to W. Schwarzhans, which were very helpful for comparison purposes. We gratefully acknowledge access to the Leica M 165 FC stereomicroscope and digital camera system in the geobiology lab, as provided by O. Voigt and G. Wörheide (both LMU, Munich). We thank C. Franz and A. Lord (both SMF, Frankfurt/Main) for preparing the SEM photographs of type specimens of Protonymus? primus, and we also thank Z. Gholami (LMU, Munich) and R. Melzer (ZSM, Munich) for the SEM photographs of our callionymids otoliths. O. Anistratenko (IGS NASU, Kiyv) and V. Anistratenko (IZ NASU, Kiev) are thanked for their support in identifying the gastropods to species, particularly those of the genus Mohrensternia in the samples from Karagaily. V. Kovalenko (IGN NASU, Kiyv) is thanked for identifying the ostracods found associated with the studied fossils, and we also thank L. Golovina (GI RAS, Moscow) for drawing important nannoplankton conclusions. Finally, we thank the reviewers R. Brzobohaty (Brno) and O. Aguilera (Niteroi) for their most valuable input regarding an earlier version of the manuscript. The research performed by A. Bratishko was funded by Luhansk Taras Shevchenko National University and the German Academic Exchange Service (DAAD).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Bratishko, A., Schwarzhans, W., Reichenbacher, B. et al. Fish otoliths from the Konkian (Miocene, early Serravallian) of Mangyshlak (Kazakhstan): testimony to an early endemic evolution in the Eastern Paratethys. Paläontol Z 89, 839–889 (2015). https://doi.org/10.1007/s12542-015-0274-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-015-0274-4