
Abstract
Megachirella wachtleri Renesto et Posenato, 2003, a well preserved partial reptile skeleton from the Middle Triassic of the Dolomites (N. Italy), was originally considered a lepidosauromorph, but no phylogenetic analysis was carried out. Consequently, the taxon was overlooked in later phylogenetic analyses of the Diapsida. Here, the holotype and only known specimen of M. wachtleri is redescribed, allowing an investigation of its phylogenetic relationships. Phylogenetic analyses confirm that Megachirella is a lepidosauromorph close to the crown group lepidosaurs (Squamata + Rhynchocephalia). Megachirella enhances our knowledge of the series of morphological modifications that led to the origin of the Lepidosauria, the most diverse clade of extant reptiles.
Kurzfassung
Megachirella wachtleri Renesto et Posenato, 2003, ein gut erhaltenes Teilskelett aus der mittleren Trias der Dolomiten (Nord-Italien) wurde ursprünglich den Lepidosauromorphen zugeordnet, auch wenn keine phylogentische Analyse durchgeführt wurde. In späteren phylogenetischen Studien wurde M. wachtleri daher völlig außer Acht gelassen. Eine Neubearbeitung des Holotypus und einzigen Exemplars von M. wachtleri ermöglicht es, die phylogenetische Position der Art neu zu analysieren. Die phylogenetische Studie bestätigt, dass die Art zu den Lepidosauromorpha gehört und nah an der Krongruppe der Lepidosauren (Squamata + Rhynchocephalia) steht. Megachirella erweitert unsere Kenntnisse über den Ablauf der morphologischen Veränderungen, die zum Ursprung der Lepidosaurier geführt hat, der variabelsten Gruppe rezenter Reptilien.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Megachirella wachtleri Renesto et Posenato, 2003 is a small diapsid reptile from the Anisian (Middle Triassic) of the Dolomites (Trentino-Alto Adige/Südtirol, Italy), represented by only one incomplete, but nonetheless very well preserved, specimen. In the preliminary description, no in-depth phylogenetic investigation was carried out, but on the basis of several characters of the skull, in particular the morphology of the squamosal–quadrate complex, as well as of the pectoral girdle and fore limb (Renesto and Posenato 2003), Megachirella was originally considered a lepidosauriform, possibly a sister taxon of the Lepidosauria, nested below the dichotomy between Rhynchocephalia and Squamata.
Our scant knowledge of the early history and diversity of the stem of the Lepidosauria should have rendered Megachirella a taxon of interest for reconstructing lepidosauromorph history, but instead M. wachtleri was not included in further studies on the phylogeny of diapsids (e.g. Müller 2004), or of lepidosauromorphs (e.g. Evans and Borsuk-Bialinicka 2009). The only reference to Megachirella can be found in Evans and Jones (2010, p. 29), where it was dismissed from any phylogeny, with a statement that its affinities with other diapsids were unresolved.
In order to test previous hypotheses about the phylogenetic relationships of M. wachtleri, the holotype (and only specimen) has been restudied and available characters have been scored for phylogenetic analyses. Megachirella was included in four of the most widely accepted analyses of Diapsida and Lepidosauromorpha (see below). Results converge towards a position close to the origin of Lepidosauria.
Geological setting
The reptile was collected from a rich plant horizon that outcrops on the western slope of Monte Prà della Vacca/Kühwiesenkopf in the Braies/Prags Dolomites (Northern Dolomites, NE Italy) (Fig. 1). This area has been well known in the palaeontological literature since the nineteenth century, mainly for the Anisian (Pelsonian) brachiopods and ammonoids of the “alpiner Muschelkalk” (Loretz 1875; Mojsisovics 1879, 1882; Bittner 1890).

The deposit is located within the Dont Formation of the Braies Group (Pisa et al. 1978; Fois and Gaetani 1984; De Zanche et al. 1993; Senowbari-Daryan et al. 1993; Delfrati et al. 2000), which represents the lower part of the Anisian basinal succession. The Dont Formation in the Braies area has been interpreted as an hemipelagic carbonate-terrigenous sequence of a marginal basin environment, about 100 m deep (Senowbari-Daryan et al. 1993). In the Monte Prà della Vacca/Kühwiesenkopf section (Fig. 1), the Dont Formation is more than 200 m thick, and the fossiliferous horizon is located at about 75 m from its base (Broglio Loriga et al. 2002b), which is represented by a massive carbonate platform attributed to the Gracilis Formation (De Zanche et al. 1992; Senowbari-Daryan et al. 1993) (Fig. 1). The plant horizon, placed just above bed K 11 of Bechstädt and Brandner (1970, enclosure 2), is about 1 m thick; plants are concentrated in lenses of siltstone, marly siltstone and carbonatic siltstone layers some centimetres thick (Fig. 1) alternated with silty and marly limestone layers containing sparse terrestrial plant remains. Marine organisms (fishes, bivalves, brachiopods, ammonoids and gastropods) are present, but not abundant, throughout the horizon. The occurrence and preservation of terrestrial and marine fossils together must be related to very rapid burial events caused by gravity flows within a marine basin, connected with heavy storms in the terrestrial domain (Tintori et al. 2001; Broglio Loriga et al. 2002a, b). At the base of the deposit, a layer of carbonatic siltstone about 30 cm thick represents a marker bed, which is easily recognizable because its upper surface is rich in trunks that are roughly N–S oriented (Fig. 1). Large but rare plant remains (trunks) are also present at the top of a limestone bank situated about 2 m below (Fig. 1). This unit of nodular, massive wackestone and packstone contains a rich marine assemblage with foraminifers, byssate bivalves (Mysidioptera) and brachiopods (Punctospiriferella and Angustothyris). The Dont Formation of the Dolomites is traditionally considered Pelsonian–Illyrian in age (Delfrati et al. 2000 and references therein). The section of Monte Prà della Vacca/Kühwiesenkopf has been dated as the Pelsonian (Anisian, Middle Triassic), on the basis of brachiopod assemblages (Bechstädt and Brandner 1970) and benthic foraminifera (Broglio Loriga et al. 2002b). A detailed description of the stratigraphy of the deposit can be found in Renesto and Posenato (2003).
Materials and methods
The new anatomical study allowed coding for a number of characters. Megachirella was coded (“Appendix 1”) according to four distinct matrices used to evaluate diapsid relationships, namely Laurin (1991), from which Apisaurus witteri was pruned, since it has been shown to be a varanopid synapsid (Reisz et al. 2010; Dilkes 1998), as updated and modified according to Renesto et al. (2010), Evans and Borsuk-Bialynicka (2009) (see “Appendix 2” for details), and in a subset of the large analysis of Müller (2004) as emended by Evans and Borsuk-Bialynicka (2009). Given the focus of this study, we used Müller’s (2004) “reduced” matrix, where unstable taxa, such as turtles and ichtyopetygians, were excluded (see Müller 2004, p. 386, Fig. 3, p. 387). The new matrices were compiled and constrained (where necessary, see below) using the Mesquite system for phylogenetic computing (Maddison and Maddison 2011). Maximum parsimony analysis using PAUP* version 4.0b10 (Swofford 2003) was conducted on the datasets. Laurin (1991) was analysed using a branch-and-bound algorithm, while all other matrices were analysed using the tree search protocol proposed by Ruta et al. (2003): 10,000 random stepwise addition sequences were carried out, followed by tree bisection-reconnection (TBR) branch-swapping, holding only one tree in memory at any time. Searching on each tree with unlimited MAXTREES recovered the same island of trees. No shorter trees were found in any of the analyses by employing the iterative re-weighting strategy proposed by Quicke et al. (2001). All characters were treated as unordered and equally weighted. Node support was estimated using bootstrapping (Felsenstein 1985; Farris et al. 1996). Consensus trees were calculated using the algorithm chosen in the original paper (i.e. strict or majority rule) in order to allow direct comparison with published consensus trees.
Systematic palaeontology
Diapsida Osborn, 1903
Lepidosauromorpha Gauthier, 1984
Lepidosauriformes Gauthier, Estes et De Queiroz, 1988
Megachirella Renesto et Posenato, 2003
Megachirella wachtleri Renesto et Posenato, 2003
2003, Megachirella wachtleri Renesto and Posenato, p. 466 pl.1; p. 467 pl.2.
Holotype: PZO628 (former collection number KÜH1501) of the catalogue of Museo di Scienze Naturali dell’Alto Adige/Naturmuseum Südtirol, Bolzano/Bozen, Italy, the only known specimen of Megachirella wachtleri Renesto et Posenato, 2003 (Fig. 2).

Megachirella wachtleri PZO628, holotype and only known specimen. Scale bar = 10 mm
Age: Pelsonian (Anisian, Middle Triassic).
Horizon and locality: Plant-rich level above bed K 11 of Bechstädt and Brandner (1970) Dont Formation, Prà della Vacca/Kühwiesenkopf area Braies/Prags, Bolzano/Bozen, Alto Adige, Northeastern Italy (Fig. 1).
Diagnosis (emended from Renesto and Posenato 2003): small diapsid reptile with proportionally large skull, moderately elongate neck and stout forelimbs. Skull with large upper temporal fenestra and ventrally open lower temporal fenestra, jugal with short pointed caudal process. Squamosal with ventral cotyle for a mobile quadrate, absence of quadratojugal, quadrate wide, with deep quadrate conch. Subthecodont dentition. Vertebrae amphicoelous, non-notochordal, with subrectangular neural spines almost as long as high. First 3–4 dorsal ribs dichocephalous. Presence of paired mineralized sternal plates. Humerus distally expanded with well-developed ectepicondylar and entepicondylar foramina. Presence of an ulnar patella. Medial centrale does not reach the fourth distal carpal. Huge recurved claws in the manus.
Description
General remarks
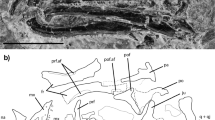
Part of the skull as well as the cervical and part of the dorsal regions of the vertebral column with associated ribs are preserved along with the pectoral girdle, the entire right forelimb, part of the left forelimb and a few elements of the gastralia (Fig. 2).
Skull
The restudy of the specimen corroborates the original description. The snout region is almost completely lost: only a fragment of the right maxilla is preserved. The median and caudal portions of the frontals are preserved, showing that the two bones were fused together, forming a wide plate ornamented by small tubercles. The parietals are narrow and deep, approximately two times longer than wide, and form the proximal margin of a wide craniocaudally elongate upper temporal fenestra. The postorbital is triradiate with a narrow squamosal process, while the postfrontal is almost entirely covered by the displaced frontals, but it enters the upper temporal fenestra. The jugal is narrow and short, ending caudally with a pointed process, and does not reach the midpoint of the upper temporal fenestra; the jugal is absent so the lower temporal bar is open. The squamosal is wide and bears a ventral articular area for the quadrate, the descending process is very small, while the process that meets the parietal is robust and forms the caudal margin of the upper temporal fenestra. The quadrate shows at its dorsal end a convex articulation for the squamosal and at its ventral end a double condyle for the articulation with the mandible. The caudal margin is deeply embayed, forming a tympanic conch (Figs. 3, 4).

Megachirella wachtleri PZO628. a The skull as preserved. b Line drawing showing the best-preserved bones. a angular, ar articular, ch ceratohyal, d dentary, fr frontal, j jugal, mx maxilla, pa parietal, pal palate bones, pf prefrontal, po postorbital, pop process of opisthotic, q quadrate, sa surangular, sq squamosal. Scale bar = 5 mm

Megachirella wachtleri, reconstruction of the preserved portion of the skull. an angular, ar articular, d dentary, fr frontal, j jugal, pa parietal, pf postfrontal, po postorbital, q quadrate, sq squamosal
Vertebral column
The entire cervical series is preserved (Fig. 2) and comprises six vertebrae, while only nine dorsal vertebrae are preserved: the first six and a further three more caudally after a gap. All vertebrae are amphicoelous and non-notochordal; reanalysis reveals the absence of accessory intervertebral articulations (zygosphene–zygantrum, contra Renesto and Posenato 2003).
Cervical vertebrae are mostly exposed in dorsal or caudal view. The neural spine is about the same height as in the dorsal vertebrae. Neural arches are fused with the centra. Pre- and post-zygapophyses have nearly horizontal articular facets.
Preserved dorsal vertebrae (Fig. 5) show a wide, squared neural arch, subrectangular neural spines approximately 1.5× longer than high; little projecting pre- and post-zygapophyses whose articular facets are only slightly inclined, and short transverse processes placed at the bases of the neural arches.

Megachirella wachtleri PZO628 dorsal region and right forelimb. Scale bar = 5 mm. cl clavicle, g gastralia, h humerus, icl posterior end of the interclavicle, ra radius, sc scapula, shr holocephalous (single-headed) rib, sp sternal plate, thr dicocephalous (two-headed) rib
Ribs and gastralia
Cervical ribs are slender, with nearly straight shafts; their articular heads cannot be detected. Dorsal ribs (Fig. 5) are long and slender; their shafts are deeply arched and hollow, as testified by the presence of calcite crystals that fill the cavity inside the rib (Renesto and Posenato 2003). The size and thickness of the dorsal ribs decreases gradually caudally. The first three to four dorsal ribs are distinctly dichocephalous, becoming holocephalous only after the fourth to fifth dorsal rib. Several thin gastralia are partially exposed on the right side of the specimen (Fig. 5).
Pectoral girdle
The clavicle is crescentic, thicker in its midportion, and with a tapering lateral tip that bears a flattened articular area that meets the anterior margin of the coracoid (Fig. 5).
The dorsal end of a high, subrectangular scapular blade emerges from the matrix, while broad flat coracoid plates can be seen below the dorsal vertebrae. Caudal to the right scapulocoracoid, a flat ossification can be identified as the right sternal plate, with suboval caudal margins. Close to this bone, the caudal end of the narrow stem of the interclavicle can be seen.
Forelimb
The right fore limb is exposed in ventral (postaxial) view (Figs. 2, 5), along with the distal portion of the left humerus and the proximal head of the left ulna, which is the only preserved part of the latter bone. The humerus has a moderately expanded proximal head which is craniocaudally flattened, while the distal head is much more expanded and dorsoventrally flattened, so that the two heads are expanded approximately at a right angle to one another. An entepicondylar foramen is present, and the ectepicondyle is perforated by an elongate foramen. The ulna is longer and stouter than the radius; its proximal head is expanded and anteroposteriorly flattened with a small olecranon. A rounded ossification lies close to the proximal head of the ulna and may represent an ulnar patella. The shaft of the ulna is straight and narrow, subcircular in section; the distal head is narrower than the proximal one, and its margin is strongly convex. The tip of the proximal head of the radius is overlapped by the distal end of the humerus; however, it can be observed that it is much narrower than that of the ulna, the shaft is narrow, straight, and somewhat thinner than that of the ulna, and the distal head is moderately expanded with a smoothly convex distal margin.
The carpus is partially obscured by pyrite crystals that make it difficult to recognise some elements close to the ulna, but its pattern can be reconstructed. Distal to the ulna, a rounded ulnare and a narrower intermedium are present; more laterally both medial and lateral centrale are present; the radiale is a small rounded bone. Four distal carpals can be recognized, the first is proximodistally compressed, while the other are rounded and their size increases from the second to the fourth, which is the largest. The first metacarpal is short and stout, the others are more cylindrical and slender; the fourth metacarpal is stouter than the third but its distal end is embedded in the matrix, so it is not possible to ascertain if its length is equal to or greater than that of the third metacarpal. Only the first digit is completely exposed, consisting of an elongate preungual phalanx and a stout, long and sharp claw, which is laterally compressed as in scansorial animals (Cartmill 1985). The apices of the large and sharp claws are the sole visible parts of digits 2–5.
Discussion
Despite the incompleteness of the specimen, restudy allowed character coding to perform a phylogenetic study. The relationships of Megachirella within diapsids as retrieved by inclusion in four previously published matrices are shown in Figs. 6 and 7. Using Laurin’s (1991) dataset, the analysis finds two most parsimonious trees (MPTs) with tree length (TL) = 110, consistency index (CI) = 0.636, retention index (RI) = 0.789 and rescaled consistency index (RC) = 0.496 (Fig. 6a). Megachirella falls within Lepidosauromorpha, although with unresolved relationships. Adding Megachirella to Dilkes’ (1998) data matrix, as updated and modified by Renesto et al. (2010), allows a more inclusive analysis of relatedness within Lepidosauromorpha (Fig. 6b). Here, Megachirella is found as a sister taxon of the early sphenodontian Gephyrosaurus + Squamata (2 MPTs, TL = 360, CI = 0.467, RI = 0.633, RC = 0.295). The whole tree loses resolution when compared to the original publication, and with the noteworthy collapse of the rhynchosaur clade. If a topologically constrained analysis is performed to investigate the effect of forcing monophyly of Rhynchosauria, the results of the search for the shortest tree are only slightly less parsimonious, finding three trees of TL = 363, three steps longer than the two MPTs with no constraints. A further analysis performed within Lepidosauromorpha on the basis of an earlier version of the character set used by Evans and Borsuk-Bialynicka (2009) for Sophineta (kindly provided by Evans, see “Appendix 2”) found Megachirella nested one node above kuehneosaurs, making it the sister taxon of the (Marmoretta (Sophineta (Lepidosauria))) clade (54 MPTs, TL = 119, CI = 0.714, RI = 0.609, RC = 0.435) (Fig. 6c).


When Megachirella was then included in the reduced matrix of Müller (2004) as emended by Evans and Borsuk-Bialynicka (2009), the strict consensus tree (Fig. 7a) showed unresolved relationships within Lepidosauromorpha, but Megachirella was consistently found as a sister taxon of the Squamata (10 MPTs, TL = 533, CI = 0.397, RI = 0.617, RC = 0.250). The 50 % majority rule consensus tree (Fig. 6b) shows a pectinate arrangement of lepidosauromorphs (Marmoretta(Sophineta(Rhynchocephalia(Squamata, Megachirella)))). This finding is less consistent than previous ones with the actual stratigraphic distribution of the taxa (Fig. 8), and with previous hypotheses on the sequence of character acquisition during the early history of lepidosauromorphs (Evans and Borsuk-Bialynicka 2009; Evans and Jones 2010).

Stratophylogenetic trees of Lepidosauromorpha, modified from Evans (2010) and timescale by Gradstein et al. (2012). a Relationships of Megachirella as suggested by its inclusion in the matrices of Laurin (1991), of Dilkes (1998) and of Evans and Borsuk-Bialynicka (2009); b relationships of Megachirella with Lepidosauria as suggested by its inclusion in the matrix of Müller (2004), as emended by Evans and Borsuk-Bialynicka (2009). Nodes are a Lepidosauromorpha, b Lepidosauria, c Rhynchocephalia. Taxon definitions according to Evans and Jones (2010)
All skeletal correlates support the nesting of Megachirella within the Lepidosauromorpha sensu Gauthier (1984), for instance the presence of paired sternal plates, of a fully enclosed ectepicondylar foramen in the humerus, and the medial centrale, which is approximately twice the size of the lateral centrale in the manus (e.g. Gauthier et al. 1988). The presence of a quadrate bowed in lateral view with a prominent lateral conch as a support to the tympanum brings Megachirella within Lepidosauriformes, designated as the clade comprising kuehenosaurids and lepidosaurs (Gauthier et al. 1988), plus Marmoretta and Sophineta (Evans and Borsuk-Białynicka 2009). Within Lepidosauria, Megachirella is constantly retrieved closer to Squamata and Rhynchocephalia than to Kueheneosauridae, and within Müller’s (2004) matrix as a sister group of the Squamata. In Müller’s dataset, Megachirella is nested within Lepidosauria because it shares an anterior process of the quadrate, a quadrate conch, the absence of the subtemporal process of the jugal and paired sternal plates (characters 20, 29, 81 and 91 of Müller 2004); then it is found to be more closely related to the Squamata than to Rhynchocephalia, mainly due to the presence of a subpleurodont dentition (character 38), of a ventrally open lower temporal fenestra and of a squamosal restricted to the dorsal region of the skull (characters 17 and 18 of Müller 2004 both scored 0 for rhynchocephalians and 2 for squamates and Megachirella).
Here, we interpret some characters previously proposed as synapomorphies for rhynchocephalians, such as the presence of gastralia (Müller 2004), as plesiomorphic features since they are widespread among diapsids. Other characters are interpreted here as homoplasies, such as the presence of an entepicondylar foramen, which is considered to have evolved independently in squamates and kueheneosaurids (contra Müller 2004).
In conclusion, Megachirella is always found nested within the Lepidosauromorpha: either nested as a sister taxon of Lepidosauria or within Lepidosauria as a sister taxon of the Squamata. Considering the incompleteness of the skeleton, the different character sets used and taxa included, as well as the still low number of known early lepidosaurs discovered, the placement of Megachirella within Lepidosauromorpha and close to the Lepidosauria seems well supported in all analyses, while more data are needed to univocally assess the position of Megachirella with respect to the Squamata.
Being Pelsonian (Anisian, Middle Triassic) in age, Megachirella is coeval with the oldest true Lepidosauria (Evans and Borsuk-Bialynicka 2009) and intermediate between the Early Triassic Sophineta and the Jurassic Marmoretta, thus filling a temporal gap in the lepidosauromorph fossil record. Furthermore, Megachirella enhances our knowledge on the series of morphological modifications that led to the origin of the Lepidosauria, the most diverse clade of extant reptiles.
References
Bechstädt, T., and R. Brandner. 1970. Das Anis zwischen St. Vigil um dem Höhlensteintal (Pragser und Olanger Dolomiten, Südtirol), 9–103. Innsbruck: Festband Geolische Institut, 300-Jahr-Feier Univ.
Bittner, A. 1890. Brachiopoden der alpinen trias. Abhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt 14: 325. (Wien).
Broglio Loriga, C., A. Fugagnoli, M. Gaetani, J. van Konijnenburg-van Cittert, E. Kustatscher, N. Mantovani, R. Posenato, S. Renesto, A. Tintori, and M. Wachtler. 2002a. Il giacimento a piante di Kühwiesenkopf/Monte Prà della Vacca (Anisico, Dolomiti di Braies): una proposta di salvaguardia. In Giornate di Paleontologia, ed. A. Tintori, 13. Riassunti: Verona-Bolca-Priabona. (6–8 June 2002).
Broglio Loriga, C., A. Fugagnoli, J. van Konijnenburg-van Cittert, E. Kustatscher, R. Posenato, and M. Wachtler. 2002b. The Anisian macroflora from the Northern Dolomites (Monte Prà della Vacca/Kühwiesenkopf, Braies/Prags): a first report. Rivista Italiana di Paleontologia e Stratigrafia 108: 381–390.
Cartmill, M. 1985. Climbing. In Functional vertebrate morphology, ed. M. Hildebrand, D. Bramble, K.F. Liem, and D.B. Wake, 73–78. Cambridge: Harvard University Press.
Delfrati, L., M. Balini, and D. Masetti. 2000. Formazione di Dont. In Carta geologica d’Italia, 1:50.000, Catalogo delle formazioni, fascicolo I, Unità validate, vol. 7 (ser. 3), ed. L. Delfrati, et al., 89–99. Italy: Quaderni del Sevizio Geologico d’Italia.
De Zanche, V., A. Franzin, P. Gianolla, P. Mietto, and C. Siorpaes. 1992. The Piz da Peres section (Valdora/Olang, Pusteria Valley, Italy): a reappraisal of the Anisian stratigraphy in the Dolomites. Eclogae Geologicae Helvetiae 85: 127–143.
De Zanche, V., P. Gianolla, P. Mietto, C. Siorpaes, and P.R. Vail. 1993. Triassic sequence stratigraphy in the Dolomites (Italy). Memorie di Scienze Geologiche 45: 1–27.
Dilkes, D.W. 1998. The Early Triassic rhynchosaur Mesosuchus browni and the interrelationships of basal archosauromorph reptiles. Philosophical Transaction of the Royal Society B 353: 501–541.
Evans, S.E., and M. Borsuk-Białynicka. 2009. A small lepidosauromorph reptile from the early Triassic of Poland. Acta Palaeontologica Polonica 65: 179–202.
Evans, S.E., and M.E.H. Jones. 2010. The origin, early history and diversification of lepidosauromorph reptiles. In New aspects of mesozoic biodiversity, vol. 27, ed. S. Bandyopadhyay, 27–44. Berlin: Springer-Verlag.
Farris, J.S., V.A. Albert, A.M. Källersjö, D. Lipscomb, and A.G. Kluge. 1996. Parsimony jackknifing outperforms neighbor-joining. Cladistics 12: 99–124.
Felsenstein, J. 1985. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39: 783–791.
Fois, E., and Gaetani, M. 1984. The recovery of reef-building communities and the role of cnidarians in carbonate sequences of the Middle Triassic (Anisian) in the Italian Dolomites. Paleontographica Americana 54: 191–200.
Gauthier, J. 1984. A cladistic analysis of the higher categories of the Diapsida (Ph.D. thesis, University of California). University Microfilms International 85-12825: 564.
Gauthier, J., R. Estes, and K. de Queiroz. 1988. A phylogenetic analysis of the Lepidosauromorpha. In Phylogenetic relationships of the lizard families, ed. R. Estes, and G. Pregill. Stanford: Stanford University Press.
Gradstein, F.M., J.G. Ogg, M. Schmitz, and G. Ogg (eds.). 2012. The geologic time scale 2012, 2nd ed. Amsterdam: Elsevier.
Laurin, M. 1991. The osteology of a Lower Permian eosuchian from Texas and a review of diapsid phylogeny. Zoological Journal of the Linnean Society 101: 59–95.
Loretz, H. 1875. Einige Petrefacten der alpinen Trias aus den Südalpen. Zeitschrift der Deutschen Gesellschaft für Geowissenschaften 27: 784–841.
Maddison, W.P., and D.R. Maddison. 2011. Mesquite: a modular system for evolutionary analysis, version 2.75. http://mesquiteproject.org.
Mojsisovics, E. 1879. Die Dolomit-Riffe von Südtirol und Venetien. In Beiträge zur Bildungsgeschichte der Alpen, 551. Wien: A. Hölder Verlag.
Mojsisovics, E. 1882. Die Cefalopoden der mediterranen Triasprovinz. Abhandlungen der Kaiserlich-Königlichen Geologischen Reichsanstalt 10: 1–320.
Müller, J. 2004. The relationships among diapsid reptiles and the influence of taxon selection. In Recent advances in the origin and early radiation of vertebrates, ed. G. Arratia, M.V.H. Wilson, and R. Cloutier, 379–408. München: Dr Friedrich Pfeil.
Osborn, H.F. 1903. The reptilian subclasses Diapsida and Synapsida and the early history of the Diaptosauria. Memoirs of the American Museum of Natural History 1: 8.
Pisa, G., E. Farabegoli, and E. Ott. 1978. Stratigrafia e paleogeografia dei terreni anisici della Conca di Agordo e dell’Alta Val di Zoldo (Dolomiti Sudorientali). Memorie della Società Geologica Italiana 18: 63–92.
Quicke, D.L.J., J. Taylor, and A. Purvis. 2001. Changing the landscape: a new strategy for estimating large phylogenies. Systematic Biology 50: 60–66.
Reisz, R.R., M. Laurin, and D. Marjanović. 2010. Apisaurus witteri from the Lower Permian of Texas: yet another small varanopid synapsid, not a diapsid. Journal of Vertebrate Paleontology 30: 1628–1631.
Renesto, S., and R. Posenato. 2003. A new lepidosauromorph reptile from the Middle Triassic of the Dolomites. Rivista Italiana di Paleontologia e Stratigrafia 109: 463–474.
Renesto, S., J.A. Spielmann, S.G. Lucas, and G. Tarditi Spagnoli. 2010. The taxonomy and paleobiology of the Late Triassic (Carnian–Norian: Adamanian–Apachean) drepanosaurs (Diapsida: Archosauromorpha: Drepanosauromorpha). New Mexico Museum of Natural History and Science Bulletin 46: 1–81.
Ruta, M., M.I. Coates, and D.L.J. Quicke. 2003. Early tetrapod relationships revisited. Biological Reviews 78: 251–345.
Senowbari-Daryan, B., R. Zühlke, T. Bechstädt, and E. Flügel. 1993. Anisian (Middle Triassic) buildups of the Northern Dolomites (Italy): the recovery of reef communities after the Perm/Triassic crisis. Facies 28: 181–256.
Swofford, D.L. 2003. PAUP*: phylogenetic analysis using parsimony (* and other methods), version 4.0b10. Sunderland: Sinauer Associates.
Tintori, A., Posenato, R., Kustatscher, E., and M. Wachtler. 2001. New Triassic fish faunas from paralic environments in the Alps. In 3rd international meeting on mesozoic fishes, Serpiano, Swizerland, 26–31 August 2001 (abstract book).
Acknowledgments
Our sincere thanks to E. Kustatscher (Museo di Scienze Naturali dell’Alto Adige/Naturmuseum Südtirol, Italy) for access to the specimen and for translation into German of the summary (together with K. Bauer, same Institution), and to S.E. Evans (University College of London, UK) for access to unpublished data and permission to reproduce it. We also thank M.J. Benton (University of Bristol, UK) for useful comments on an earlier version of the manuscript and linguistic revision. M. Caldwell (University of Alberta, Canada) and M. Laurin (Muséum National d’Histoire Naturelle, Paris, France) are thanked for their constructive reviews, and the Editor O.W.M. Rauhut (Bayerische Staatssammlung für Paläontologie und Geologie, Munich) for useful comments and corrections. This study was supported by the Promotion of Educational Policies, University and Research Department of the Autonomous Province of Bolzano-South Tyrol as part of the project “The Permian–Triassic ecological crisis in the Dolomites: extinction and recovery dynamics in Terrestrial Ecosystems”.
Author information
Authors and Affiliations
Corresponding author
Appendices
Appendix 1
Character coding for Megachirella wachtleri within the different matrices:
In Laurin (1991): 11?11 11?11 11??0 0??11 1?111 ??11? 111?? ??100 00??? ?000? 0000? ?0000 01?11 111??
In Dilkes (1998), after Renesto et al. (2010): 0001? ????? ????? 0???0 00010 1?011 ?1022 12??? ????? ????? 1?1?0 ?000? ???0? ????0 ??020 00000 10100 1???? ?11?? ??00? ????? ?000? ????? ????? ????? ????? 0??0? 0???0 ??0?
In Müller (2004), after Evans and Borsuk-Bialynicka (2009): ????? ???10 0?0?1 0221? ?1??? 11?1? ????1 ??3?1 01?10 0?0?? 1110? ???00 00020 0???? ????? ????? 10??? ???0? 2???? ????? 0001? ???0? ????? ??00? 000?? ????? 1???? ????? ????0 10??? ????0 11??? ????? ????? ???0? 100?? ????
In Evans and Borsuk-Białynicka (2009): ??0?? 10010 1??11 ?1??? ????? 100?0 ?100? 01110 10??1 101?? 10100 ????? ????? ????? ????? 0?
Appendix 2
List of characters and their character states used by Evans and Borsuk-Bialynicka (2009), which was not originally published and is reported here with permission from the author (courtesy Evans).
-
1.
Lacrimal large with extensive exposure on cheek, 0; small, confined to orbital rim, 1; absent, 2.
-
2.
Greatest width of nasals exceeds greatest width of both nares, 0; is less then the latter, 1.
-
3.
Frontoparietal suture more or less W shaped, subequal to or narrower then the nasofrontal suture in width, 0; suture more or less straight broader then naso-frontal suture, 1.
-
4.
Parietal foramen within parietal, 0; on frontoparietal border or within frontal, 1; absent, 2.
-
5.
Postparietals present, 0; absent, 1.
-
6.
Tabulars present, 0; absent, 1.
-
7.
Postorbital overlain by postfrontal, 0; postfrontal overlain by postorbital, 1.
-
8.
Squamosal extends anteriorly halfway or less over lower temporal fenestra and is widely separated from jugal by postorbital, 0; extends more than halfway over lower temporal fenestra and comes close to or contacts jugal below postorbital, 1.
-
9.
Anterior process of quadratojugal present, 0; absent, 1.
-
10.
Posterior process of jugal extends about halfway back or less below lower temporal fenestra, 0; extends more than halfway back below lower temporal fenestra, 1.
-
11.
Quadratojugal always present as a separate element at some point in ontogeny, 0; never present as a separate element, 1.
-
12.
Supratemporal present, 0; fused or lost, 1.
-
13.
Supratemporal lies superficially, 0; lies deep association with ventral face of postparietal process, 1.
-
14.
Ventral process of squamosal present, 0; absent, 1.
-
15.
Adductor chamber small, quadrate does not extend well below level of occipital condyle, 0; enlarged adductor fossa and quadrate extends well below adductor fossa, 1.
-
16.
Nares paired, 0; nares confluent, 1.
-
17.
Quadrate narrow in posterior view, 0; wide in posterior view forming lateral conch, 1.
-
18.
Quadrate foramen present, 0; absent, 1.
-
19.
Vomerine teeth numerous, 0; few or absent, 1.
-
20.
Teeth or transverse flange of pterygoid present, 0; absent, 1.
-
21.
Abducens canal absent or incomplete, 0; present and complete, 1.
-
22.
Parasphenoid teeth present, 0; absent 1.
-
23.
Paroccipital process does not extend laterally to contact quadrate, 0; process contacts quadrate, 1.
-
24.
Paroccipital process not expanded distally, 0; expanded distally, 1.
-
25.
Stapes thick and perforated for passage of stapedial artery in adults, 0; stapes thinner and imperforate in adults, 1; stapes columelliform and usually imperforate in adults, 2.
-
26.
Quadrate straight in lateral view, 0; bowed, 1.
-
27.
Postorbital contacts parietal, 0; separated from parietal, 1.
-
28.
Posterior process of postorbital does not reach to posterior end of upper temporal fenestra, 0; process extends beyond posterior margin of upper temporal fenestra, 1.
-
29.
Choanal fossa on palatine absent, 0; present, 1.
-
30.
Snout comparatively shorter and broader, 0; longer and narrower, 1.
-
31.
Premaxillae paired in full grown adults, 0; fused prior to hatching, 1.
-
32.
Parietals paired in full grown adults, 0; fused prior to hatching, 1.
-
33.
Exoccipitals sutured to opisthotic above and below metotic fissure, 0; fused only above metotic fissure with metotic fissure extended ventrally into basioccipital, 1; metotic fissure subdivided to create a dorsal vagus foramen and a ventral opening for the glossopharyngeal nerve and perilymphatic sac, 2.
-
34.
Frontals paired, 0; fused, 1.
-
35.
Splenial present, 0; absent, 1.
-
36.
Angular extends posteriorly to articular condyle, 0; does not, 1.
-
37.
Angular extends more than one-third up lateral face of mandible, 0; less than one-third, 1.
-
38.
Retroarticular process relatively small or absent, 0; large, 1.
-
39.
Outer one-third of articular condyle formed by surangular, 0; formed entirely by articular, surangular forming only outer rim, 1.
-
40.
Teeth set in shallow sockets or depressions, 0; superficially attached to medial side of jaw with labial wall higher than lingual one, but only slightly, 1; labial wall significantly higher than lingual wall (pleurodont), 2.
-
41.
Neural arches fuse to their respective centra in postembryonic development, 0; in embryo, 1.
-
42.
Zygosphenes and zygantra, absent, 0; present, 1.
-
43.
Caudal autotomy absent, 0; present, 1.
-
44.
Trunk vertebrae without accessory articulations between neural spines, 0; accessory articulations present, 1.
-
45.
Intercentra present in trunk, 0; absent, 1.
-
46.
Vertebrae amphicoelous notocordal, 0; amphicoelous solid, 1; procoelous, 2.
-
47.
Transverse processes not elongated, 0; short transverse processes, 1; long transverse processes, 2.
-
48.
One or more cervical ribs bear two distinct heads, 0; all ribs single headed, 1; one or more ribs with three heads, 2.
-
49.
No trunk vertebrae without free ribs, 0; one or more postsacrals without free ribs (lumbar), 1.
-
50.
Sacral and caudal ribs fuse to their respective centra in postembryonic development, 0; fuse in embryo, 1.
-
51.
Rib-bearing portion of sternum a single rod, 0; this region of sternum enlarged and formed from two plates; they remain paired until nearly maximum size, 1; paired sternal plates fuse in embryo, 2.
-
52.
Anterior margin of scapulocoracoid without fenestrae, 0; fenestrate, 1.
-
53.
Interclavicle robust, 0; gracile, 1.
-
54.
Entepicondylar foramen of humerus, present, 0; absent, 1.
-
55.
Prominent posteromedial process of distal epiphyses of radius absent, 0; present, 1.
-
56.
Fourth metacarpal longer than third, 0; third and fourth metacarpal subequal, 1; fourth metacarpal shorter than third, 2.
-
57.
Ilium includes 80–85 % of acetabulum, 0; 60–65 % acetabulum, 1.
-
58.
Pubic flange on ilium absent, 0; present, 1.
-
59.
Iliac blade long, 0; short, 1.
-
60.
Anteromedial portion of pubis not out-turned dorsally, 0; out-turned dorsally, 1.
-
61.
Dorsal edge of ilium essentially horizontal, 0; more steeply inclined, 1.
-
62.
Pelvis a solid plate, 0; small thyroid fenestra with broad pubic symphysis, 1; large fenestra with small pubic symphysis, 2.
-
63.
Ischial tubera short, 0; long, 1.
-
64.
In fully grown individuals, pelvic elements separate, 0; fused, 1.
-
65.
Fibula and femur articulate end to end, distal femoral condyles symmetrical, 0; fibula sits in a recess on lateral margin of distal end of femur, distal femoral condyles markedly asymmetrical, 1.
-
66.
Articular surface of fibula for astragalocalcaneum restricted to a small portion of the distal end, 0; covers most of distal end of fibula, 1.
-
67.
Astragalus and calcaneum separate throughout ontogeny, 0; fused prior to fusion of scapulocoracoid, 1.
-
68.
Lateral centrale of pes discrete throughout ontogeny, 0; fused to astragalus in embryo, 1.
-
69.
Distal tarsal 1 present, 0; absent 1.
-
70.
Distal tarsal 2 present, 0; absent 1.
-
71.
Distal tarsal 5 present at some point in postembryonic ontogeny, 0; absent or fused in embryo, 1.
-
72.
Metatarsal 5 straight, 0; inflected, somewhat hooked but without enlarged plantar tubercles, 1; hooked, inflected angulated proximally and with enlarged medial and lateral plantar tubercles, 2.
-
73.
Ridge on distal extremity of tibia for astragalar articulation, 0; no ridge present, 1.
-
74.
No tongue and groove articulation between distal tarsal 4 and astragalocalcaneum, 0; process on distal tarsal 4 extends under astragalocalcaneum to form partial tongue and groove articulation, 1; full tongue and groove articulation, 2.
-
75.
Premaxilla without posterolateral process excluding maxilla form narial rim, 0; with posterolateral process, 1.
-
76.
Cervical vertebrae not elongated, 0; elongated, 1.
-
77.
Astragalus and calcaneums without specialised joint between them, 0; with specialised joint, 1.
Rights and permissions
About this article
Cite this article
Renesto, S., Bernardi, M. Redescription and phylogenetic relationships of Megachirella wachtleri Renesto et Posenato, 2003 (Reptilia, Diapsida). Paläontol Z 88, 197–210 (2014). https://doi.org/10.1007/s12542-013-0194-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-013-0194-0