
Abstract
The mollusc fauna of the early Middle Miocene (Langhian) intramontane Alpine Lake Groisenbach is described for the first time. The shells derive from the Feistring Formation in the Aflenz Basin in Austria, which was covered by Lake Groisenbach. The assemblage is moderately diverse with 12 gastropod and 2 bivalve species, suggesting shallow lacustrine and fluvial settings. Among the gastropods, only Theodoxus crenulatus (Klein, 1853) is known from other Miocene localities, whilst all other species are documented so far only from Lake Groisenbach. None of the Early and Middle Miocene lake systems of the Alpine-Carpathian Foredeep and the Balkan Peninsula displays any faunistic resemblance with this new fauna. Even coeval lake faunas from the close-by Graz Basin have no species in common with Lake Groisenbach. This pattern points to a surprising endemicity and biogeographic fragmentation in the Central European freshwater systems during the Early and Middle Miocene. The uniqueness of the newly described fauna is also indicated by the completely erratic occurrence of the otherwise African-Mediterranean genus Bulinus, which is unknown from all other central European Miocene freshwater systems. Emmericia roetzeli Harzhauser and Neubauer nov. sp., Nematurella zuschini Neubauer and Harzhauser nov. sp., Romania fastigata Neubauer and Harzhauser nov. sp., Odontohydrobia groisenbachensis Neubauer and Harzhauser nov. sp., Odontohydrobia pompatica Neubauer and Harzhauser nov. sp., Odontohydrobia styriaca Harzhauser and Neubauer nov. sp., Planorbis austroalpinus Harzhauser and Neubauer nov. sp., Gyraulus sachsenhoferi Harzhauser and Neubauer nov. sp., Bulinus corici Harzhauser and Neubauer nov. sp., Ferrissia crenellata Harzhauser and Neubauer nov. sp. and Stagnicola reinholdkunzi Harzhauser and Neubauer nov. sp. are introduced as new species.
Kurzfassung
Erstmals wird die Molluskenfauna des intramontanen alpinen Groisenbach-Sees aus dem frühen Mittel-Miozän (Langhium) beschrieben. Die Schalen stammen aus der Feistring-Formation des Aflenzer Beckens, in dem sich im mittleren Miozän der Groisenbach-See entwickelte. Die Vergesellschaftung ist mit 12 Gastropoden und 2 Bivalven Arten moderat divers und deutet auf seicht lakustrine und fluviatile Ablagerungsbedingungen. Unter den Gastropoden ist nur Theodoxus crenulatus (Klein, 1853) auch aus anderen miozänen Fundstellen bekannt, während alle anderen Arten bisher nur aus dem Groisenbach-See bekannt sind. Keines der früh- und mittelmiozänen Seen-Systeme der Alpen-Karpaten-Vortiefe und des Balkans zeigt faunistische Beziehungen zu der neuen Fauna. Selbst die gleichaltrigen Faunen des geographisch nahen Grazer Beckens weisen keine gemeinsamen Arten auf. Diese Muster deuten auf einen überraschend hohen Endemismus sowie auf eine starke biogeographische Fragmentierung der mitteleuropäischen Süßwassersysteme während des frühen und mittleren Miozäns. Die Einzigartigkeit der Fauna zeigt sich auch in dem erratischen Auftreten der ansonsten afrikanisch-mediterranen Gattung Bulinus, die aus keinem anderen miozänen See Mitteleuropas bekannt ist. Emmericia roetzeli Harzhauser und Neubauer nov. sp., Nematurella zuschini Neubauer und Harzhauser nov. sp., Romania fastigata Neubauer und Harzhauser nov. sp., Odontohydrobia groisenbachensis Neubauer und Harzhauser nov. sp., Odontohydrobia pompatica Neubauer und Harzhauser nov. sp., Odontohydrobia styriaca Harzhauser und Neubauer nov. sp., Planorbis austroalpinus Harzhauser und Neubauer nov. sp., Gyraulus sachsenhoferi Harzhauser und Neubauer nov. sp., Bulinus corici Harzhauser und Neubauer nov. sp., Ferrissia crenellata Harzhauser und Neubauer nov. sp., und Stagnicola reinholdkunzi Harzhauser und Neubauer nov. sp. werden als neue Arten eingeführt.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Modern European freshwater faunas display little biogeographic differentiation (Glöer 2002). This pattern is contrasted by an extremely high endemicity during the Miocene (Harzhauser and Mandic 2008). This endemicity, however, may be overemphasised by the simple fact that only few stratigraphic coeval lake faunas are well studied. For example, the huge lake systems in the Alpine-Carpathian Foredeep and on the Balkan Peninsula, described as Rzehakia Lake System (RLS) and Dinaride Lake System (DLS) by Harzhauser and Mandic (2008), have little stratigraphic overlap. Other lake systems, such as Lake Steinheim, Lake Skopje and the many small lakes fringing the Paratethys Sea, are also not strictly coeval. Moreover, the data on Miocene freshwater faunas are strongly biased towards long-lived lakes with high subsidence rates, huge piles of sediments and a good fossil record. In contrast, small and geologically short-lived lakes and their faunas are virtually unknown. This lack of data is especially obvious for the many tectonically induced lakes that formed during the Early and Middle Miocene in the young Eastern Alps. Despite the considerable basin fills of several hundred metres, no mollusc faunas have been described from these lakes so far. Only along the margins of the Eastern Alps do the early Middle Miocene (Langhian) Rein and Graz basins yield comparable faunas (Gobanz 1854; Unger 1858).
Now, the discovery of a new early Middle Miocene lake fauna from the Aflenz Basin in Austria allows a direct comparison with the coeval faunas of the Rein and Graz basins and that of the Sinj Basin in Croatia (De Leeuw et al. 2010), which was part of the Dinaride Lake System. This enables the proposed endemicity to be tested at least for these lake faunas.
Geological setting
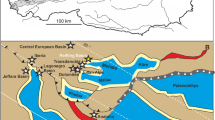
Lateral eastward extrusion of crustal blocks along strike slip faults in the Eastern Alps resulted in the formation of several tectonically controlled intramontane basins during the Early and Middle Miocene (Ratschbacher et al. 1991). The most important of these basins are the Lavant, Fohnsdorf, Seckau, Trofaiach, Leoben, Parschlug and Aflenz basins (Fig. 1). These gave rise to the development of a chain of lakes and deltaic systems which were fringed by swampy wetlands. Some of the lakes, such as Lake Ingering in the Fohnsdorf Basin, were moderately long-lived and existed up to 2 Ma (Sachsenhofer et al. 2003).

The herein studied assemblage derives exclusively from the Aflenz Basin, which was covered by the rather small and short-lived Lake Groisenbach (Sachsenhofer et al. 2003). This basin is a c. 13 km long and c. 2 km wide composite pull-apart basin (Reischenbacher and Sachsenhofer 2002; Sachsenhofer et al. 2003). Its margins are formed by the Upper Austroalpine Greywacke Unit and the Northern Calcareous Alps in the north and by Middle Austroalpine units. The Miocene basin fill consists of the coarse-grained Feistring Formation and the overlying Göriach Formation. The Feistring Fm. comprises up to 300 m of prograding fluvial deposits, whilst the Göriach Fm. is composed of up to 200 m of limnic pelites in the base and fluvial deposits in the top (Sachsenhofer et al. 2003). Both formations are separated by several coal seams and diatomites. The latter were used by Hajos (1972) to date the succession into the early Middle Miocene (=Langhian, early Badenian).
All studied samples derive from the Feistring Fm. The samples were collected during geological mapping by the Geological Survey of Austria. Samples 1 and 2 derive from the Jauring section (47°33′13.05″N, 15°15′39.01″E) and sample 4 was collected at the Pletschgraben section (47°33′46.22″N, 15°20′05.27″E).
Systematic palaeontology
The systematic arrangement follows largely Bouchet and Rocroi (2005, 2010), whilst Albrecht et al. (2007) and Jörger et al. (2010) are considered for pulmonates and Glöer (2002) for the Emmericiidae. All specimens are stored in the collection of the Geological-Palaeontological Department of the Natural History Museum in Vienna.
-
Class Gastropoda Cuvier, 1797
-
Subclass Orthogastropoda Ponder and Lindberg, 1997
-
Superorder Neritaemorphi Koken, 1896
-
Order Neritopsina Cox and Knight, 1960
-
Superfamily Neritoidea Lamarck, 1809
-
Family Neritidae Lamarck, 1809
-
Genus Theodoxus Montfort, 1810
-
Type species: Theodoxus lutetianus Montfort, 1810 (=Nerita fluviatilis Linnaeus, 1758). Recent, Europe

Neritidae, Bulininae and Emmericiidae from the Jauring section, Aflenz Basin. a–d Theodoxus crenulatus (Klein, 1853), NHMW 2011/0021/0035. e–h Bulinus corici Harzhauser and Neubauer nov. sp. e NHMW 2011/0021/0008 f NHMW 2011/0021/0009, g Holotype, NHMW 2011/0021/0006, h Paratype, NHMW 2011/0021/0007. i–l Emmericia roetzeli Harzhauser and Neubauer nov. sp., i Holotype, NHMW 2011/0021/0031, j Paratype 1, NHMW 2011/0021/0032, k Paratype 2, NHMW 2011/0021/0033, l NHMW 2011/0021/0034. Scale bar = 1 mm
- *1853:
-
Neritina crenulata Klein: 221, pl. 5, fig. 18
- 1875:
-
Neritina crenulata Klein.—Sandberger: 571, pl. 28, fig. 13
-
Material: c. 150 specimens, Jauring, Pletschgraben, NHMW 2011/0021/0035
-
Measurements: maximum diameter 11 mm
Remarks
Here we present the first SEM pictures of this species, showing strong growth lines on the early teleoconch (Fig. 2). This medium-sized species was very common in all samples and displays a wide range of variability concerning colour patterns. Interestingly, this variability decreases distinctly in the strongly fluvial-influenced samples where only specimens with large triangular speckles occur. The coeval Theodoxus sinjanus (Brusina, 1876) differs in its more globular outline and the regularly increasing diameter of the last whorl, whereas the last whorl of Theodoxus crenulatus has an allometric growth. Theodoxus semidentatus (Sandberger, 1870) has a similar sculpture of the callus pad but is distinctly higher and tends to develop a slight concavity within the adapical third of the whorls.
Distribution
Theodoxus crenulatus settled most freshwater systems of the intramontane basins of the early Alps during the Middle Miocene and is recorded also from the Fohnsdorf and Lavant basins (Hölzel and Wagreich 2004; Wank 1991). Originally, it was described from the Middle Miocene of Zwiefaltendorf in southern Germany (Klein 1853). It is widespread in the fluvial deposits fringing the Paratethys Sea throughout the Early and Middle Miocene (Schlickum 1976; Harzhauser 2002; Harzhauser and Mandic 2008). Oligocene occurrences, mentioned by Báldi (1973), represent another species based on their more spherical outline and the sharply demarcated callus pad.
-
Superorder Caenogastropoda Cox, 1960
-
Order Littorinimorpha Golikov and Starobogatov, 1975
-
Superfamily Rissooidea Gray, 1847
-
Family Emmericiidae Brusina, 1870
-
Genus Emmericia Brusina, 1870
-
Type species: Paludina patula Brumati, 1838. Recent, Dalmatia
-
Emmericia roetzeli Harzhauser and Neubauer nov. sp. (Fig. 2i–l)
-
Material: >100 specimens
-
Holotype: NHMW 2011/0021/0031, height: 6.9 mm, diameter: 4.8 mm, Fig. 2i
-
Paratype 1: NHMW 2011/0021/0032, height: 7.3 mm, diameter: 5.2 mm, Fig. 2j
-
Paratype 2: NHMW 2011/0021/0033, height: 6.2 mm, diameter: 4.8 mm, Fig. 2k
-
Stratum typicum: lacustrine sand within gravel of the Feistring Formation
-
Type locality: Jauring section, Aflenz Basin; 47°33′13.05″N, 15°15′39.01″E
-
Age: Early Middle Miocene (Langhian; Badenian)
-
Name: in honour of Reinhard Roetzel, geologist at the Geological Survey of Vienna
Description
Stout glossy shell with short conical spire consisting of three weakly convex, smooth whorls with narrow, thread-like sutures. Protoconch measuring c. 600 μm in diameter, consisting of less than one smooth depressed whorl, causing a flattened apex. Its demarcation from the teleoconch is indistinct. The last whorl attains about one-half of the total height, consisting of a weakly convex adapical third and a convex base. The height of the last whorl and its convexity are variable. Growth lines may form a faint axial sculpture which becomes most pronounced on the base. On fully grown specimens these axial threads are crossed by numerous very delicate spiral grooves, which are rarely preserved. Aperture oblique, drop-shaped, terminating in a thickened continuous lip. Columellar lip adherent but distinct; outer lip slightly thickened, expanded in a narrow collar-like structure. A narrow siphonal notch is developed in the angulated adapical part of the aperture. A shallow internal furrow appears close behind the aperture without corresponding swelling on the external shell surface. No umbilicus is developed.
Remarks
This extant genus has its roots in the Miocene. The oldest records are the species from the Aflenz Basin and the coeval Emmericia canaliculata Brusina, 1870 from the early Middle Miocene of the Drniš and Sinj basins in Croatia. The latter is clearly distinguished from the Austrian one by its convex spire whorls. E. subpatula Kókay, 1966, from the Badenian of Herend in Hungary, if an Emmericia at all, is smaller, lacks the broad collar-like aperture and strongly convex whorls with incised sutures. After a major stratigraphic gap, the next two representatives are known from the Late Miocene of Lake Pannon (Harzhauser and Mandic 2008). The more frequent Pannonian species—erroneously identified as Emmericia canaliculata by Harzhauser and Binder (2004)—differs in its convex whorls, the higher spire, the thicker lip and the relatively smaller aperture. The second Late Miocene to Pliocene species, Emmericia schulzeriana Brusina, 1882, develops a very high spire, an elongate aperture and a very narrow collar. E. botici Brusina, 1902, E. krizanici Brusina, 1902 and E. zivkovici Brusina, 1902, from the Pliocene of Slavonia and Serbia, are all conspicuously more slender and develop smaller last whorls. The sunken protoconch corresponds fully to late Miocene and Pliocene representatives of the genus as described by Harzhauser and Binder (2004) and Esu et al. (2001). A relation with Staliopsis Rzehak, 1893 is excluded based on the large and immersed protoconch of the new species, whilst Staliopsis has protoconchs of only 300 μm diameter (Kowalke and Reichenbacher 2005).
Distribution
Only known from the early Middle Miocene Lake Groisenbach.
-
Family Hydrobiidae Stimpson, 1865
-
Subfamily Hydrobiinae Stimpson, 1865
-
Genus Nematurella Sandberger, 1870
-
Type species: Nematurella flexilabris Sandberger, 1870. Middle Miocene, Tramelan, Switzerland
-
Nematurella zuschini Neubauer and Harzhauser nov. sp. (Fig. 3a–c, m)
Fig. 3 
Hydrobiidae from the Jauring section, Aflenz Basin. a–c, m Nematurella zuschini Neubauer and Harzhauser nov. sp. a Paratype 1, NHMW 2011/0021/0028, b holotype, NHMW 2011/0021/0027, c paratype 2, NHMW 2011/0021/0029, m protoconch, NHMW 2011/0021/0030. d–f, n Romania fastigata Neubauer and Harzhauser nov. sp. d Paratype 2, NHMW 2011/0021/0025, e holotype, NHMW 2011/0021/0023, f paratype 1, NHMW 2011/0021/0024, n protoconch, NHMW 2011/0021/0026. g–h Odontohydrobia groisenbachensis Neubauer and Harzhauser nov. sp. g Paratype, NHMW 2011/0021/0022, h holotype, NHMW 2011/0021/0021. i–j Odontohydrobia styriaca Harzhauser and Neubauer nov. sp. i Holotype, NHMW 2011/0021/0017, j paratype, NHMW 2011/0021/0018. k–l, o Odontohydrobia pompatica Neubauer and Harzhauser nov. sp. k, o Holotype, NHMW 2011/0021/0019, l paratype, NHMW 2011/0021/0020. Scale bar = 1 mm
-
Material: c. 550 specimens
-
Holotype: NHMW 2011/0021/0027, height: 3.0 mm, diameter: 1.6 mm, Fig. 3b
-
Paratype 1: NHMW 2011/0021/0028, height: 3.3 mm, diameter: 1.7 mm, Fig. 3a
-
Paratype 2: NHMW 2011/0021/0029, height: 3.2 mm, diameter: 1.6 mm, Fig. 3c
-
Stratum typicum: lacustrine sand within gravel of the Feistring Formation
-
Type locality: Jauring section, Aflenz Basin; 47°33′13.05″N, 15°15′39.01″E
-
Age: Early Middle Miocene (Langhian; Badenian)
-
Name: in honour of Martin Zuschin, palaeontologist at the University of Vienna

Description
Protoconch smooth, consisting of 1.25 broad and convex whorls; 280–290 μm in diameter. The transition to the teleoconch is marked by a weak axial thread and by the onset of the early teleoconch sculpture, consisting of straight prosocline growth lines. Teleoconch elongate conical with a spire angle of 40–45°, comprising 4–4.5 whorls. The first whorl is low and evenly convex, passing into successively less convex to nearly straight-sided whorls. The last two whorls may even develop a gentle concavity close to the upper suture causing an indistinct subsutural band. The last whorl attains about 60% of the total height and displays a distinct but variable angulation between flank and the slightly convex base. This angulation changes from pronounced in apertural view to less distinct and convex in lateral view. The weak growth lines are crossed by very faint spiral ribs in exceptionally well-preserved specimens. Aperture broad ovoid to slightly elongate ovoid with convex inner lip and well rounded outer lip. In lateral view, the aperture is parallel to the axis with a posterior indentation at the outer lip and a slightly protruding inner lip; columella smooth. The terminal part of the last whorl is detached from the base and grows at an angle of c. 35° relative to the suture. This part of the whorl bears prominent, densely spaced growth ridges. Umbilicus narrow but well visible, grading into the slit-like interspace between base and terminal part of the last whorl.
Remarks
The species differs from all Nematurella species, as revised by Schlickum (1960), in its less convex teleoconch whorls and the basal angulation.
Distribution
Only known from the early Middle Miocene Lake Groisenbach.
-
Genus Romania Cossmann, 1913
Type species
Juliania expansa Roman, 1910. Oligocene, France.
-
Romania fastigata Neubauer and Harzhauser nov. sp. (Fig. 3d–f, n)
-
Material: c. 25 specimens
-
Holotype: NHMW 2011/0021/0023, height: 3.1 mm, diameter: 1.5 mm, Fig. 3e
-
Paratype 1: NHMW 2011/0021/0024, height: 3.1 mm, diameter: 1.5 mm, Fig. 3f
-
Paratype 2: NHMW 2011/0021/0025, height: 3.1 mm, diameter: 1.6 mm, Fig. 3d
-
Stratum typicum: lacustrine sand within gravel of the Feistring Formation
-
Type locality: Jauring section, Aflenz Basin; 47°33′13.05″N, 15°15′39.01″E
-
Age: Early Middle Miocene (Langhian; Badenian)
-
Name: fastigata—Latin for oblique and sloped, referring to the posterior termination of the last whorl
Description
Protoconch ranging around 300 μm in diameter, poorly preserved in all available specimens, smooth and consists of about 1–1.5 strongly convex whorls with incised sutures. Teleoconch consisting of five strongly and regularly convex whorls with the maximum convexity in the middle or slightly below the middle of the whorls; sutures incised. Apical angle ranging between 40 and 45°. The height of the whorls is regularly increasing; their convexity is only decreasing on the base, causing a slightly elongate last whorl, which attains about 60% of the total height. Surface covered with delicate, straight prosocline growth lines. Well-preserved shells exhibit a very weak spiral sculpture of faint grooves. Aperture thick-shelled, oblique semilunar with slightly concave inner lip, well rounded outer lip and a weak posterior angulation. Inner lip separated from base by a narrow furrow; umbilicus slit-like. In lateral view, the aperture is strongly oblique forming an angle of c. 20° relative to the shell-axis; the aperture margins are straight without notch. A characteristic axial groove occurs close to the aperture inside the shell, being reflected by a slight bulge on the outer shell surface. The position of the groove coincides with a change in growth direction of the upper part of the whorl. This terminal part becomes detached from the base and grows in the anterior direction at an angle of 45° relative to the suture.
Remarks
The generic affiliation with Romania is based on the aperture, the shape of the detached last whorl and the overall outline. The stratigraphic gap between the Oligocene Romania expansa (Roman, 1910) and the Middle Miocene species, however, may raise doubts about the generic identification. Romania fastigata differs from the type species only in its more convex spire whorls and the less elongate shape. A closely related species, with comparable morphology, is illustrated as “Nematurella? Nikolajevići” by Brusina, 1902 from the Middle Miocene of Zvezdan in Serbia. This DLS species has a rather short last whorl, convex spire whorls, a detached last whorl with straight but oblique margin and identical umbilicus features as Romania fastigata. Differences between both species are the wider aperture and the less steep angle of the sutural-margin of the terminal part of the last whorl in Romania nikolajevici (Brusina, 1902). The general morphology is reminiscent of a Nematurella Sandberger 1870. The straight-sided margins of the aperture and its oblique angle relative to the axis, however, contradict a generic affiliation with Nematurella, which has a sigmoidal outer lip. Moreover, the anterior part of the aperture in Nematurella usually protrudes in lateral view (Schlickum 1971).
The second genus with comparable aperture is the Plio-Pleistocene Tournouerina Schlickum, 1971. The type species Tournouerina lugdunensis (Tournouër, 1879) differs in its higher and broader last whorl and the low convexity of the spire whorls. Similarly, most species of this genus, as described by Schlickum (1978), differ in their broad and high last whorl and the drop-shaped outline from the Miocene species. Only the Plio-Pleistocene Tournouerina belnensis (Delafond and Depéret, 1893) is reminiscent of the Miocene species in its rather convex spire whorls and a comparatively short last whorl.
Distribution
Only known from the early Middle Miocene Lake Groisenbach.
-
Subfamily Pyrgulinae Brusina, 1881
-
Genus Odontohydrobia Pavlović, 1928
-
Type species: Odontohydrobia ranojevici Pavlović, 1928, by subsequent designation (Wenz 1939). Late Miocene, Serbia
-
Odontohydrobia groisenbachensis Neubauer and Harzhauser nov. sp. (Fig. 3g, h)
-
Material: c. 35 specimens
-
Holotype: NHMW 2011/0021/0021, height: 3.3 mm, diameter: 1.6 mm, Fig. 3h
-
Paratype: NHMW 2011/0021/0022, height: 3.5 mm, diameter: 1.7 mm, Fig. 3g
-
Stratum typicum: lacustrine sand within gravel of the Feistring Formation
-
Type locality: Jauring section, Aflenz Basin; 47°33′13.05″N, 15°15′39.01″E
-
Age: Early Middle Miocene (Langhian; Badenian)
-
Name: referring to Lake Groisenbach
Description
Protoconch unknown. The teleoconch ranges around 3–4 mm in height and consists of three to four whorls; the first teleoconch whorl is strongly convex and low, resulting in an obtuse apex. Later whorls are moderately high and only weakly convex with the maximum convexity in the lower third of the whorl. The last whorl is high, barrel-shaped, nearly straight sided and attains about two-thirds of the total height. Shell surface covered by delicate and densely spaced growth lines, which are most prominent close to the incised sutures and on the base. This sigmoidal-prosocline axial sculpture is crossed on the last whorl by very delicate spiral threads, which are difficult to detect even in SEM pictures. The aperture is ovate to semilunar with a slightly convex inner and strongly convex outer lip; both are equally thickened. A weak indentation occurs at the anterior part of the outer lip. Inner lip well demarcated from the base, covering a narrow, slit-like umbilicus. A characteristic columellar fold occurs in the middle of the columella; it is most distinct in the inner part of the shell and fades out towards the aperture.
Remarks
Odontohydrobia is defined mainly based on its conspicuous columellar fold. Up to now, most representatives were described from the Late Miocene of Lake Pannon (Serbia: Pavlović 1928; Romania: Jekelius 1944; Vienna Basin: Papp 1953). The youngest representatives are from the Pliocene of Romania and Croatia (Jekelius 1932; Jurišić-Polšak et al. 1997). Thus, Odontohydrobia groisenbachensis and its congeners from Lake Groisenbach are the oldest known representatives of the genus. It differs from the slender Lake Pannon species O. cryptodonta Jekelius, 1944, O. clessini Pavlović, 1928 and O. ranojevici Pavlović, 1928 in its stout barrel-shaped outline and the lower number of teleoconch whorls. O. wagneri Pavlović, 1928, O. clessini Pavlović, 1928 and O. leobersdorfensis Papp, 1953 develop strongly convex whorls; O. bathyomphaloides Pavlović, 1928 is much larger, distinctly conical and has a wider umbilicus. Odontohydrobia croatica Jurišić-Polšak, Sokač, and Poje, 1997 develops five teleoconch whorls and is slender conical, and O. dacica Jekelius, 1932 differs from O. groisenbachensis also in its conical shape and convex whorls.
Distribution
So far only known from the early Middle Miocene Lake Groisenbach.
-
Odontohydrobia pompatica Neubauer and Harzhauser nov. sp. (Fig. 3k, l, o)
-
Material: 12 specimens
-
Holotype: NHMW 2011/0021/0019, height: 3.1 mm, diameter: 1.5 mm, Fig. 3k, o
-
Paratype: NHMW 2011/0021/0020, height: 3.2 mm, diameter: 1.4 mm, Fig. 3l
-
Stratum typicum: lacustrine sand within gravel of the Feistring Formation
-
Type locality: Jauring section, Aflenz Basin; 47°33′13.05″N, 15°15′39.01″E
-
Age: Early Middle Miocene (Langhian; Badenian)
-
Name: pompatica—Latin for gorgeous
Description
Protoconch measures c. 250–300 μm and consists of c. one smooth and strongly convex whorl, passing without marked boundary into the teleoconch, which is indicated by the onset of faint axial threads. The teleoconch consists of five whorls; the first two whorls are strongly convex, whilst later whorls develop nearly straight-sided flanks. The spire angle of the first four teleoconch whorls ranges around 50°. This rather regularly conical spire is contrasted by a high and barrel-shaped last whorl, which attains about one half of the total height. It has a narrower diameter than the penultimate whorl resulting in a constricted outline. The sutures are moderately incised but accentuated by the convexity of the whorls. Surface covered by delicate but distinct, feebly sigmoidal growth lines, which are crossed by indistinct and shallow spiral furrows. Both sculpture elements cause a weakly cancellated pattern. Aperture slender ovate with thin and slightly convex outer lip and almost straight inner lip, which nearly covers the slit-like umbilicus. The posterior part of the inner lip is slightly protruding in lateral view, whilst the outer lip has a weak indentation. The columella bears a broad but weak fold that becomes more prominent inside the shell.
Remarks
The characteristic last whorl and bulge-like penultimate whorl distinguish this species from all other Odontohydrobia species. No morphologic overlap with the two other congeneric species from Lake Groisenbach can be stated.
Distribution
Only known from the early Middle Miocene Lake Groisenbach.
-
Odontohydrobia styriaca Harzhauser and Neubauer nov. sp. (Fig. 3i, j)
-
Material: c. 40 specimens
-
Holotype: NHMW 2011/0021/0017, height: 3.3 mm, diameter: 1.9 mm, Fig. 3i
-
Paratype: NHMW 2011/0021/0018, height: 2.9 mm, diameter: 1.7 mm, Fig. 3j
-
Stratum typicum: lacustrine sand within gravel of the Feistring Formation
-
Type locality: Jauring section, Aflenz Basin; 47°33′13.05″N, 15°15′39.01″E
-
Age: Early Middle Miocene (Langhian; Badenian)
-
Name: Referring to the province of Styria in Austria
Description
Stout ovoid shell consisting of three teleoconch whorls. The low protoconch is poorly preserved. The first teleoconch whorl is low and only weakly convex; the second whorl increases in height and is nearly straight sided aside from the weak convexity at the upper and lower sutures. The morphology of the last whorl is rather variable ranging from globular with strong median convexity to stout with slight convexity. Shell surface covered by a dense pattern of growth lines, which are most prominent on the base. Aperture ovoid with slight posterior angulation and strongly thickened edges. The suture between aperture and base is deeply incised and results in a slightly detached position of the termination of the last whorl. Thick inner lip, well demarcated from the base, forming a moderately narrow and deep umbilicus. A central fold divides the oblique columella into two slightly concave parts. The prominent swelling of the fold fades out towards the aperture.
Remarks
This species differs from its syntopic congener Odontohydrobia groisenbachensis nov. sp. in its globose outline and the shorter last whorl and wider umbilicus. Its columella is oblique and bears two concavities close to the columellar fold. Moreover, it develops one whorl less at the same size. The stout ovoid shape and the low number of whorls distinguish O. styriaca from all Late Miocene to Pliocene Odontohydrobia species, as described by Pavlović (1928), Jekelius (1932, 1944), Papp (1953) and Jurišić-Polšak et al. (1997). These species develop at least 4 or more teleoconch whorls.
Distribution
Only known from the early Middle Miocene Lake Groisenbach.
-
Clade Panpulmonata Jörger et al., 2010
-
Order Hygrophila Férussac, 1822
-
Suborder Branchiopulmonata Morton, 1955
-
Superfamily Planorboidea Gray, 1840
-
Family Planorbidae Gray, 1840
-
Subfamily Planorbinae Rafinesque, 1815
-
Genus Planorbis Müller, 1773
-
Type species: Helix planorbis Linnaeus, 1758. Recent, Europe
-
Planorbis austroalpinus Harzhauser and Neubauer nov. sp. (Fig. 4a–d)
Fig. 4 
Planorbidae from the Jauring section, Aflenz Basin. a–d Planorbis austroalpinus Harzhauser and Neubauer nov. sp. a Paratype 1, NHMW 2011/0021/0014, b paratype 2, NHMW 2011/0021/0015, c holotype, NHMW 2011/0021/0013, d NHMW 2011/0021/0016. e–g Gyraulus sachsenhoferi Harzhauser and Neubauer nov. sp. e Holotype, NHMW 2011/0021/0010, f paratype 1, NHMW 2011/0021/0011, g paratype 2, NHMW 2011/0021/0012. Scale bar = 1 mm
-
Material: >40 shells
-
Holotype: NHMW 2011/0021/0013, height: 1.3 mm, diameter: 2.5 mm, Fig. 4c
-
Paratype 1: NHMW 2011/0021/0014, height: 1.1 mm, diameter: 2.1 mm, Fig. 4a
-
Paratype 2: NHMW 2011/0021/0015, height: 1.0 mm, diameter: 1.4 mm, Fig. 4b
-
Stratum typicum: lacustrine sand within gravel of the Feistring Formation
-
Type locality: Jauring section, Aflenz Basin; 47°33′13.05″N, 15°15′39.01″E
-
Age: Early Middle Miocene (Langhian; Badenian)
-
Name: referring to the setting of the locality in the Austrian Alps

Description
Small sized, strongly involute shells of 2.5–3 very high, bulbous and strongly convex whorls. Protoconch deeply sunken in spire consisting of 0.9 moderately convex whorls with characteristic striae. These are six to seven widely spaced, slightly wrinkled spiral ribs, which may be dissolved in two or three individual spiral threads. Spire deeply sunken and nearly covered by the last whorl. Umbilicus very deep and narrow. A wide and convex angulation separates the upper and lower half of the shells causing a bulky triangular cross section of the whorls in adult shells. Few specimens display faint spiral threads close to the angulation. This sculpture is missing on all juvenile shells. Additional threads may be formed by the growth lines close to the umbilicus. The growth lines are strongly sigmoidal with prosocyrt orientation on the top and base and opisthocyrt orientation on the flank. Distinct growth lines and even indistinct ribs appear close to the aperture, which is slightly expanding in diameter in fully grown shells.
Remarks
The specimens might be considered as juveniles of a larger species. The densely spaced growth lines close to the aperture as seen in the specimen illustrated as Fig. 4c, however, suggest that the shells represent fully grown specimens. Among the coeval fauna of the Dinaride Lake System, only Gyraulus oncostomus (Brusina, 1902) is slightly reminiscent of the species from the Aflenz Basin. It develops similar growth lines along the aperture and has high and strongly convex whorls but is distinctly less involute. Bathyomphalus microstatus (Bourguignat, 1880) from the Middle Miocene of Sansan in France is similar in size and also rather involute but differs in its distinctly less globose shell (Fischer 2000).
Distribution
Only known from the early Middle Miocene Lake Groisenbach.
-
Genus Gyraulus Charpentier, 1837
-
Type species: Planorbis albus Müller, 1774. Recent, Europe
-
Gyraulus sachsenhoferi Harzhauser and Neubauer nov. sp. (Fig. 4e–g)
-
Material: 14 specimens
-
Holotype: NHMW 2011/0021/0010, height: 1.0 mm, diameter: 3.7 mm, Fig. 4e
-
Paratype 1: NHMW 2011/0021/0011, height: 0.9 mm, diameter: 2.6 mm, Fig. 4f
-
Paratype 2: NHMW 2011/0021/0012, height: 1.0 mm, diameter: 2.4 mm, Fig. 4g
-
Stratum typicum: lacustrine sand within gravel of the Feistring Formation
-
Type locality: Jauring section, Aflenz Basin; 47°33′13.05″N, 15°15′39.01″E
-
Age: Early Middle Miocene (Langhian; Badenian)
-
Name: in honour of Reinhard Sachsenhofer, geologist at the University of Leoben (Austria)
Description
Planispiral, rather evolute shells of 2.5–3 teleoconch whorls; upper side moderately deepened; lower side with wide and deep umbilicus. Protoconch consisting of 0.75 whorls with smooth surface aside from faint wrinkles close to the initial cap and few spiral striae close to the upper suture. These striae fade out within the first 20% of the protoconch. The onset of the teleoconch is very indistinct and mainly indicated by the appearance of weak growth lines. Cross-section of the teleoconch whorls sub-triangular due to a marked keel that appears between the lower third and the upper two-thirds of the whorl during ontogeny. The keel is demarcated from the upper part of the whorl by a weak concavity but distinctly separated by a pronounced concavity along the lower part. This results in a slightly asymmetric dip of the keel towards the base. Upper part of the whorls moderately convex and nearly flat close to the keel; basal part flat after the concave area close to the keel and weakly convex close to the umbilicus. Entire shell covered by a dense pattern of prominent growth lines, which are prosocline on the upper shell and opisthocyrt on the base.
Remarks
Gyraulus goussardianus (Noulet, 1854) from the Middle Miocene of Sansan in France lacks the peripheral concavity of the base and the marked keel; its whorls are convex and less triangular in cross-section (Fischer 2000). Gyraulus matraensis Kókay in Gál et al., 1998 from the Middle Miocene of Mátraszölös in Hungary develops an even coarser sculpture, lacks the keel and is more involute. Shells from Turnau in the Aflenz Basin, referred to as “Planorbis applanatus” by Stur (1871) may represent this newly described species.
Distribution
Only known from the early Middle Miocene Lake Groisenbach.
-
Subfamily Bulininae Fischer and Crosse, 1880
-
Genus Bulinus Müller, 1781
-
Type species: Bulinus senegalensis Müller, 1781. Recent, Africa
-
Bulinus corici Harzhauser and Neubauer nov. sp. (Fig. 2e–h)
- 1889:
-
Lanistes noricus Tausch: 157 (nomen nudum)
- 1889:
-
Physa norica Tausch: 158 (nomen nudum)
-
Material: Five shells and c. 65 spire fragments from Jauring and Pletschgraben
-
Holotype: NHMW 2011/0021/0006, height: 10.0 mm, diameter: 8.0 mm, Fig. 2g
-
Paratype: NHMW 2011/0021/0007, height: 10.5 mm, diameter: 9.5 mm, Fig. 2h
-
Stratum typicum: lacustrine sand within gravel of the Feistring Formation
-
Type locality: Jauring section, Aflenz Basin; 47°33′13.05″N, 15°15′39.01″E
-
Age: Early Middle Miocene (Langhian; Badenian)
-
Name: in honour of Stjepan Ćorić, geologist at the Geological Survey of Vienna
Description
Moderately sized ovoid to subspherical sinistral shell consisting of five teleoconch whorls. Bulbous protoconch starting with a downward bent initial cap and 1.25 strongly convex raised whorls. Protoconch surface smooth except for few wrinkled spiral striae along the upper suture. Growth lines, formed by elongate wrinkles, appear close to the termination of the protoconch. These fade out on the first teleoconch whorl and are replaced by distinct spiral threads, which are most prominent in the middle part of the sutural shelf. The spiral sculpture fades out within the second teleoconch whorl, and the following whorls are smooth. Most specimens develop a stepped spire formed by convex early teleoconch whorls with marked shoulder and keel-like angulation. This feature is often covered by the following whorl resulting in a very variable shape of the spire. The angulation is replaced by a well-rounded convexity on the last, bulbous whorl. The suture is quickly shifted from the angulation down even below the middle of the whorl causing a rather irregular shape. Within the last half of the last whorl, the point of maximum convexity is moving towards the base, and the whorl develops a flat adapical part. Aperture drop shaped with wide and convex basal sinus and angulated adapical part. Columella with deep concavity and slightly convex parietal area. Outer lip thin; inner lip forming a thin and expanding sheet covering parts of the base and forming a moderately wide, semicircular umbilicus.
Remarks
The conspicuous sinistral coiling and sub-spherical outline are unique within the European Neogene freshwater gastropods. Therefore, the species was detected in the Aflenz Basin already in the 19th century during geological mapping. It was introduced as nomen nudum by Tausch (1889) as Lanistes noricus and Physa norica. The distinctly sheet-like inner lip and thickened columella exclude a classification as Lanistes Montfort, 1810. Tausch (1889) discussed the similarity with the extant African Bulinus nyassanus (Smith, 1877) but did not provide any description or illustration. The species is also mentioned from the Middle Miocene of Göriach and the Fohnsdorf Basin (Hofmann 1893; Hölzel and Wagreich 2004). The Miocene Bulinus trojanus (Neumayr, 1883) from Turkey differs in its broader shell and the strongly angulated shoulder. Lanistes beseneckeri Schütt in Schütt and Besenecker, 1973, from the Miocene of Chios, is stout, globular with deeply incised sutures and very wide last whorl.
Distribution
Bulinus corici is known so far only from the early Middle Miocene, when it settled the intramontane lakes Ingering and Groisenbach.
-
Subfamily Ancylinae Rafinesque, 1815
-
Genus Ferrissia Walker, 1903
-
Type species: Ancylus rivularis Say, 1817. Recent, eastern North America
-
Ferrissia crenellata Harzhauser and Neubauer nov. sp. (Fig. 5d)
Fig. 5 
Lymnaeidae, Ancylinae and Sphaeriidae from the Jauring section, Aflenz Basin. a–c Stagnicola reinholdkunzi Harzhauser and Neubauer nov. sp. a Holotype, NHMW 2011/0021/0002, b paratype, NHMW 2011/0021/0003, c NHMW 2011/0021/0004. d Ferrissia crenellata Harzhauser and Neubauer nov. sp., holotype, NHMW 2011/0021/0005. e Pisidium cf. casertanum, NHMW 2011/0021/0001. Scale bar = 1 mm

-
Material: Four specimens
-
Holotype: NHMW 2011/0021/0005, height: 3.0 mm, diameter: 1.9 mm, convexity: 1.1 mm, Fig. 5d
-
Paratype: NHMW 2011/0021/0036, height: c. 2.5 mm, diameter: c. 1.5 mm, convexity: c. 0.8 mm
-
Stratum typicum: lacustrine sand within gravel of the Feistring Formation
-
Type locality: Jauring section, Aflenz Basin; 47°33′13.05″N, 15°15′39.01″E
-
Age: Early Middle Miocene (Langhian; Badenian)
-
Name: referring to the sculpture, which is reminiscent of crenellation in architecture
Description
Cap-like protoconch, measuring about 350 μm, slightly inclined to the left. Its topmost part is smooth with a small pit in its centre, surrounded by numerous thin radial threads. These increase in number by splitting and decrease in strength. They fade out on the early teleoconch and are replaced by a dense pattern of growth lines. Radial threads reappear in the middle part of the teleoconch, causing an adaperturally oriented pinnacle-like structure at the intersections with the growth lines. Towards the apertural margin, the radial sculpture disappears. The shell is cap-shaped elliptical with its broadest diameter in the anterior half.
Remarks
The Early Miocene Ferrissia wittmanni (Schlickum, 1964), as described by Kowalke and Reichenbacher (2005) from the North Alpine Foreland Basin, develops radial threads on juvenile shells without pinnacle sculpture; adult shells are smooth. Moreover, its protoconch is larger than in F. crenellata. The same features allow a separation from the Early/Middle Miocene DLS species F. illyrica (Neumayr, 1880; Mandic et al. 2011). F. deperdita (Desmarest, 1814) from the Middle Miocene of southern Germany and the Eisenstadt-Sopron Basin in Austria (Schlickum 1976; own data) has a smooth teleoconch which lacks the characteristic radial sculpture of Ferrissia crenellata. F. truci Wautier, 1975, from the Late Miocene of France, differs in its slender shape, its strongly elongated apical region and the near-marginal position of the apex (Wautier 1975).
Distribution
Only known from the early Middle Miocene Lake Groisenbach.
-
Superfamily Lymnaeoidea Rafinesque, 1815
-
Family Lymnaeidae Rafinesque, 1815
-
Genus Stagnicola Jeffreys, 1830
-
Type species: Limneus communis Jeffreys, 1830 (=Buccinum palustre Müller, 1774). Recent, Europe
-
Stagnicola reinholdkunzi Harzhauser and Neubauer nov. sp. (Fig. 5a–c)
- 1889:
-
Limnaeus Hofmanni Tausch: 158 (nomen nudum)
-
Material: Nine shells
-
Holotype: NHMW 2011/0021/0002, height: 14.7 mm, width: 7.8 mm, Fig. 5a
-
Paratype: NHMW 2011/0021/0003, height: 17.2 mm, width: 9.2 mm, Fig. 5b
-
Stratum typicum: lacustrine sand within gravel of the Feistring Formation
-
Type locality: Jauring section, Aflenz Basin; 47°33′13.05″N, 15°15′39.01″E
-
Age: Early Middle Miocene (Langhian; Badenian)
-
Name: in honour of Reinhold Kunz, an enthusiastic fossil collector in Vienna
Description
A medium sized, drop-shaped lymnaeid of four to five teleoconch whorls consisting of moderately convex spire whorls, forming a spire angle of c. 60–70°, and a comparatively broad and convex last whorl. The convexity of the spire whorls increases during ontogeny and displays considerable intraspecific variability. The maximum convexity and diameter are usually situated close to the lower suture but may be shifted slightly upwards. Surface covered by narrow growth lines, which are most prominent on the base. Aperture moderately wide with maximum width in the lower third; attaining about three-fifths of the total shell height. Basal lip wide and slightly everted. Inner lip forming a narrow, glossy cover on the base; abapically slightly everted. Columella strongly twisted with fold-like swelling in the middle which grades into the basal lip. A shallow groove appears at the junction between the glossy cover on the base and the onset of the inner lip. A second, much weaker groove appears slightly below that point on the upper part of the inner lip. The columella and basal part of the inner lip form a straight line parallel to the shell’s axis.
Remarks
This species was already mentioned by Tausch (1889) as Limnaeus Hofmanni without providing any description or illustration. Two species from the coeval deposits of the DLS are reminiscent of the new species. Stagnicola hyaloleuca (Brusina, 1902) from the Middle Miocene of the Gacko Basin in Bosnia differs from Stagnicola reinholdkunzi in its strongly convex spire whorls and the much higher last whorl. Stagnicola korlevici (Brusina, 1884) from Miočić in Croatia is larger, develops a narrower, less convex last whorl and has an oblique inner lip and columella whilst this part of the shell is parallel to the axis in S. reinholdkunzi.
The late Middle Miocene Stagnicola armaniacensis (Noulet, 1857) sensu Schlickum (1976) has a shorter spire and the inner and basal lips are less prominent. Moreover, its aperture is more elongate. All other specimens referred to this species, as illustrated in Sandberger (1870) and Kókay (2006), are considerably more slender and have a higher spire. In the same way, the high spires of Lymnaea turrita (Klein, 1853) and Stagnicola praebouilleti Schlickum, 1970 allow a clear separation from Stagnicola reinholdkunzi. Radix socialis sensu Sandberger, 1870, which is slightly reminiscent concerning its broad outline, lacks the columellar angulation between base and inner lip. The widespread Miocene Radix dilatata (Noulet, 1854) sensu Schlickum (1966, 1976) and Fischer (2000) differs clearly in its short but comparably narrow spire and the large ovoid last whorl. The straight basal part of the inner lip and the angulation between base and inner lip distinguish Stagnicola reinholdkunzi also from the various lymnaeids described by Klein (1847) from the Middle Miocene of Germany. Stagnicola media (Reuss, 1852), from the Early Miocene of Tuchořice in Czech Republic, which is reminiscent of S. reinholdkunzi concerning the aperture, is much more slender and attains about double the height [note that Stagnicola media is identified as L. concinnus Rss. on the plate but described as L. medius m. in Reuss (1852)].
There are also several fragments of a Radix sp. which are not treated herein due to the insufficient material.
Distribution
Only known from the early Middle Miocene Lake Groisenbach.
-
Class Bivalvia Linnaeus, 1758
-
Superorder Heterodonta Neumayr, 1883
-
Order Venerida Gray, 1854
-
Superfamily Sphaerioidea Deshayes, 1855
-
Family Sphaeriidae Deshayes, 1855
-
Subfamily Pisidiinae Gray, 1857
-
Genus Pisidium Pfeiffer, 1821
-
Type species: Tellina amnica Müller, 1774. Recent, Northern Hemisphere
- 2006:
-
Pisidium pseudosphaerium Schlesch, 1947 var.—Kókay: 97, pl. 38, figs. 6–8, pl. 39, figs. 1–4, 6
-
Material: two specimens from Jauring section, Aflenz Basin; 47°33′13.05″N, 15°15′39.01″E
-
Measurements: specimen 1: length: 2.4 mm, height: 2.0 mm, convexity: c. 0.5 mm, NHMW 2011/0021/0001, Fig. 5e; specimen 2: length: 2.5 mm, height: 2.2 mm, convexity: c. 0.6 mm
Description
Only two right valves are preserved. The shell is roughly ovate and anteriorly extending. The umbo is broad, low and well rounded. The hinge plate is delicate and narrow. It bears a prominent but thin cardinal tooth (C3), which is sigmoidal shaped, where its oblique part points to the posterior end of the hinge plate. Furthermore, it is surrounded by two slight cavities (for the cardinal teeth C2 and C4 on the left valve) and produces a weak convexity on the ventral side of the hinge plate. The ligamental pit is long and slender and delimited ventrally by a thin fold, which also demarcates it from the cavity for the C4. The two posterior lateral teeth are slightly weaker than the anterior ones. Beneath the lateral teeth, traces of muscle scars appear. The outer surface is covered with growth lines, which are rather thin and equally spaced. In late ontogenetic phases faint and irregular axial threads appear close to the margin within a narrow band of c. 150 μm.
Remarks
We refrain from describing the specimens as new species due to the low number of available material. Pisidium casertanum Poli, 1791 is surprisingly similar in morphology, and we cannot separate the Miocene shells from the extant species based on the material available. This extant species is also described from the Late Miocene/Pliocene of Turkey (Becker-Platen and Kuiper 1979) and the Pliocene of West Germany (Kuiper 1972).
Some specimens from the Sarmatian of Várpalota in Hungary might be conspecific. Kókay (2006) treated these as variations of the extant Pisidium pseudosphaerium Favre, 1927. Again, this identification is rather doubtful in respect to the huge stratigraphic gap between the Miocene and extant populations. Moreover, the Miocene shells differ from the extant species in their higher umbo and the conspicuously extended anterior part. The species from Jauring differs strongly from the Early/Middle Miocene DLS species P. bellardii Brusina, 1884, which is regularly rounded and has an elongated cardinal tooth and a very long and thin ligamental pit. The Late Miocene P. krambergeri Brusina, 1884, from the Zagreb area, can be distinguished by its larger size and its less elongate outline. Regarding general shape and dentition a quite similar species is extant P. hydaspicola Theobald, 1878 from Kashmir region. It corresponds well in shape and proportions of the lateral teeth and the ligamental pit (Prashad 1925). However, the cardinal tooth is not sigmoidal but strongly angled. Also the ventral convexity is missing.
Distribution
Known from the early Middle Miocene Lake Groisenbach and maybe also from the late Middle Miocene (Sarmatian) freshwater systems of Hungary.
Discussion and conclusions
Twelve gastropod and two bivalve taxa are documented from the lower Middle Miocene Feistring Formation in the Aflenz Basin. Of these, 11 gastropod species represent new species, whilst only 1 species has also been recorded from other Miocene sections. Bivalves are recorded only by rare fragments of unidentifiable unionids and by two shells of Pisidium cf. casertanum Poli, 1791. The latter might also be present in other Middle Miocene lakes fringing the Paratethys Sea as suggested by specimens from the Sarmatian of Hungary.
Ecology
All taxa are aquatic; terrestrial molluscs are missing in the assemblages. The two samples from the Jauring section are dominated in specimen numbers by hydrobiids. The rissoid genera of Lake Groisenbach are largely extinct except for Emmericia, which is bound to springs, lakes, rivers and even caves and subterraneous rivulets (Schütt 2000; Glöer 2002). All representatives of Nematurella, Romania and Odontohydrobia are also described so far from freshwater deposits or from strongly freshwater-influenced brackish settings (Jekelius 1944; Schlickum 1971; Kowalke and Reichenbacher 2005; Harzhauser and Mandic 2008). Pulmonates, such as Stagnicola, Gyraulus, Planorbis, Ferrissia and Bulinus are less frequent. All extant representatives of these genera are freshwater dwellers (Glöer 2002). The preference of lymnaeids and Gyraulus for lentic waterbodies or only slowly flowing water suggests lacustrine conditions. Hence, lentic nearshore environments were already established in the Aflenz Basin during the formation of the Feistring Fm., which was considered as mainly fluvial influenced by Sachsenhofer et al. (2003) based on sedimentological features.
In contrast, the sample from Pletschgraben displays few of these elements and comprises mainly Theodoxus and Unio along with less abundant Bulinus. Theodoxus is known from freshwater or brackish environments and signals increased water energy (Pfleger 1984; Glöer 2002; Plaziat and Younis 2005). This suggests agitated depositional settings of a river or rivulet for the sample from Pletschgraben. All data point to pure freshwater conditions. The influence of the Paratethys Sea, which caused brackish conditions in many intramontane basins from the Lavant Basin to the Fohnsdorf Basin (Sachsenhofer et al. 2003), did not reach Lake Groisenbach at that time.
Palaeobiogeography
Our knowledge on Early and Middle Miocene European freshwater systems and their faunas is largely restricted to long-lived lake systems (Harzhauser and Mandic 2008). The most important and largest systems were the Early Miocene Rzehakia Lake System (RLS) in the Alpine-Carpathian Foredeep, the Early-Middle Miocene Dinaride Lake System (DLS) in Croatia and Bosnia-Herzegovina and Lake Skopje (LSK) in Macedonia (see Harzhauser and Mandic 2008 for geographic and palaeontological data). The species-level similarity of the assemblage from Lake Groisenbach with these coeval or near-coeval freshwater assemblages, however, is zero. Smaller and short-lived but geographically much closer coeval lakes developed in the Graz Basin and its satellites. Their aquatic mollusc fauna is homogenous and dominated by planorbids and lymnaeids (Gobanz 1854; Unger 1858). Even these coeval lakes, which were situated only c. 50 km away, have no species in common with Lake Groisenbach.
The poor faunistic relation among all these lakes is also evident at the generic level. In its overall composition, the assemblage of Lake Groisenbach is slightly reminiscent of that of the older RLS, which comprises 39 species. Both are dominated by Rissooidea along with few planorbids and lymnaeids. The RLS Hydrobiidae are represented by Nematurella Sandberger, 1870, Staliopsis Rzehak, 1893, Hydrobia Hartmann, 1821 and Ctyrokia Schlickum, 1965. Of these, only Nematurella is also present in Lake Groisenbach. Moreover, Bulinus, Odontohydrobia, Romania and Emmericia are completely missing in the RLS. The more diverse DLS fauna, with c. 110 species, is strongly dominated by Hydrobiidae and Melanopsidae. Numerous species of Melanopsis Férussac, 1807, Prososthenia Neumayr, 1869 and Fossarulus Neumayr, 1869 characterise the assemblages but are completely missing from the intramontane Lake Groisenbach. This low faunistic relation is also surprising with respect to the fact that the DLS and Lake Groisenbach existed at the same time, and exchange, e.g. via birds, would have been easily possible. Similarly, the typical Pyrgulidae and Melanopsidae genera of the LKS are absent in the studied fauna. This pattern points to a high rate of endemism within the various intramontane lakes of the early Alps and to a strong biogeographic fragmentation of European freshwater systems. Only Theodoxus crenulatus (Klein, 1853), probably a fluvial species, is also recorded from Early and Middle Miocene lakes, rivers and estuaries of Central Europe.
Odontohydrobia was suggested so far to have evolved in Lake Pannon during the Late Miocene. It is represented in Lake Groisenbach with at least three species and predates its Late Miocene congeners by c. 5 Ma. The stratigraphic gap is similar to that observed for the planorbid Orygoceras Brusina, 1882, which has its first occurrence in the DLS during the Middle Miocene and re-appears 5 Ma later in Lake Pannon (Harzhauser and Mandic 2008). Similarly, Romania was known so far only from Oligocene deposits. This lack of records indicates that several taxa persisted throughout the Miocene in still undetected lake systems.
The most outstanding taxon of Lake Groisenbach is Bulinus. Bulinus was unknown from the Central European Neogene so far. A late Middle Miocene record is only mentioned by Reichenbacher and Kowalke (2009) from Spain. In the east, the genus is represented during the Late Miocene by Bulinus trojanus (Neumayr, 1883), which is recorded from Behramkale in western Turkey. Today, Bulinus occurs from south-western Europe via the southern Mediterranean islands to Arabia and the Middle East but has its centre of diversity in Africa, where it appeared at least 12 Ma ago (Brown 1994; Morgan et al. 2002; Nalugwa et al. 2010). Its occurrence in the Aflenz Basin might thus be the oldest record of this genus. Its punctual occurrence in an intramontane Alpine lake during the Middle Miocene remains enigmatic and might be triggered by the coincidence with the Middle Miocene Climatic Optimum.
References
Albrecht, C., K. Kuhn, and B. Streit. 2007. A molecular phylogeny of Planorboidea (Gastropoda, Pulmonata): insights from enhanced taxon sampling. Zoologica Scripta 36: 27–39.
Báldi, T. 1973. Mollusc Fauna of Hungarian Upper Oligocene (Egerian), 1–511. Budapest: Akademia Kiado.
Becker-Platen, J.D., and J.G.J. Kuiper. 1979. Sphaeriiden (Mollusca, Lamellibranchia) aus dem Känozoikum der Türkei. Geologisches Jahrbuch B 33: 159–185.
Bouchet, P., and J.-P. Rocroi. 2005. Classification and nomenclator of gastropod families. Malacologia 47: 1–397.
Bouchet, P., and J.-P. Rocroi. 2010. Nomenclator of bivalve families; with a classification of bivalve families by R. Bieler, J.G. Carter, and E.V. Coan. Malacologia 52: 1–184.
Bourguignat, J.R. 1880. Etude sur les fossiles tertiaires et quaternaires de la vallée de la Cettina en Dalmatie. 1–55. Saint Germain.
Brown, D.S. 1994. Freshwater snails of Africa and their medical importance, 2nd ed, 1–609. London: Taylor & Francis.
Brumati, L. 1838. Catalogo sistematico delle conchiglie terrestri e fluviatili osservate nel territorio di Monfalcone. 1–56. Gorizia: Paternolli.
Brusina, S. 1870. Monographie der Gattungen Emmericia und Fossarulus. Verhandlungen der kaiserlichen und königlichen zoologisch-botanischen Gesellschaft in Wien 20: 925–938.
Brusina, S. 1876. Description d’especes nouvelles, provenant des terrains tertiaires de Dalmatie. Journal of Conchyliologie 24: 109–116.
Brusina, S. 1881. Le Pyrgulinae dell’Europa orientale. Bollettino della Società Malacologica Italiana 7(13–19): 229–292.
Brusina, S. 1882. Orygoceras eine neue Gasteropodengattung der Melanopsiden-Mergel Dalmatiens. Beiträge zur Paläontologie Österreich-Ungarns und des Orients 2: 33–46.
Brusina, S. 1884. Die Neritodonta Dalmatiens und Slavoniens nebst allerlei malakologischen Bemerkungen. Jahrbücher der Deutschen Malakozoologischen Gesellschaft 11: 17–120.
Brusina, S. 1902. Iconographia molluscorum fossilium in tellure tertiaria Hungariae, Croatiae, Slavoniae, Dalmatiae, Bosniae, Herzegovinae, Serbiae et Bulgariae inventorum. 30 plates. Agram: Officina Soc. Typographicae.
Cossmann, M. 1913. Revue de: F. Roman 1912, Monographie de la faune lacustre du Sannoisien du Gard. Revue critique de Paléozoologie 17: 107–108.
Cox, L.R. 1960. Thoughts on the classification of the Gastropoda. Proceedings of the Zoological Society of London 33: 239–261.
Cox, L.R., and J.B. Knight. 1960. Suborders of Archaeogastropoda. Proceedings of the Malacological Society of London 33: 262–264.
Cuvier, G. 1797. Tableau élémentaire de l’histoire naturelle des animaux. 1–710. Paris.
de Charpentier, J. 1837. Catalogue des mollusque terrestres et fluviatiles de la Suisse. In Neue Denkschriften der Allgemeinen Schweizerischen Gesellschaft für die Gesammten Naturwissenschaften 1. l–28.
Delafond, F., and C. Depéret. 1893. Les terrains tertiaires de la Bresse et leurs gites de lignite et de minerais de fer. In Etudes des Sites minéraux de France. 1–332. Paris: Imprimerie Nationale.
De Leeuw, A., O. Mandic, A. Vranjković, D. Pavelić, M. Harzhauser, W. Krijgsman, and K.F. Kuiper. 2010. Chronology and integrated stratigraphy of the Miocene Sinj Basin (Dinaride Lake System, Croatia). Palaeogeography, Palaeoclimatology, Palaeoecology 292: 155–167.
Deshayes, G.P. 1855. Catalogue of the Conchifera or bivalve shells in the collection of the British Museum. Part 2, Petricoladae (concluded); Corbiculadae, 217–292. London: British Museum.
Desmarest, A.G. 1814. Description des coquilles univalves du genre. Rissoa Bulletin de la Société Philomatique de Paris 3: 7–9.
Esu, D., O. Girotti, and G. Truc. 2001. New data on fossil Emmericiinae from Italy and France. Archiv für Molluskenkunde 129: 123–143.
Favre, J. 1927. Les mollusques post-glaciaires et actuels du bassin de Genève. Mémoires de la Société de Physique et d’Histoire Naturelle de Genève 40(3): 171–434.
de Férussac, A.E.J.P.J.F. d’A. 1807. Essai d’une méthode conchyliologique appliquée aux mollusques fluviatiles et terrestres d’après la considération de l’animal et de son test. Nouvelle édition augmentée d’une synonymie des espèces les plus remarquables, d’une table de concordance systématique de celles qui ont été décrites par Géoffroy, Poiret et Draparnaud, avec Müller et Linné, et terminée par un catalogue d’espèces observées en divers lieux de la France. xvi + 1–142. Paris: Delance.
de Férussac, A.E.J.P.J.F.d’A. 1822. Tableaux systématiques des animaux mollusques classés en familles naturelles, dans lesquels on a établi la concordance de tous les systèmes; suivis d’un prodrome général pour tous les mollusques terrestres ou fluviatiles, vivants ou fossiles, 1–110. Paris, Londres: Bertrand Sowerby.
Fischer, J.-C. 2000. La malacofauna de Sansan. In La faune miocéne de Sansan et son environment, ed. L. Ginsburg. Mémoires du Muséum national d’Histoire naturelle 183: 129–154.
Fischer, P., and H. Crosse. 1880. Études sur les mollusques terrestres et fluviatiles du Mexique et du Guatemala. Mission Scientifique au Mexique et dans l’Amérique Centrale 2: 1–713.
Flügel, H.W., and F. Neubauer. 1984. Steiermark. Geologie der österreichischen Bundesländer in kurzgefaßten Einzeldarstellungen, 1 Karte 1 : 200.000. Wien: Geologische Bundesanstalt.
Gál, E., J. Hír, E. Kessler, J. Kókay, L. Mészáros, and M. Vencel. 1998. Középsõ-miocén õsmaradványok, a Mátraszõlõs, Rákóczi-kápolna alatti útbevágásból I. A Mátraszõlõs 1. lelõhely. Folia Historico Naturalia Musei Matraensis 23: 33–78.
Glöer, P. 2002. Die Tierwelt Deutschlands, 73. Teil: Die Süßwassergastropoden Nord- und Mitteleuropas. Bestimmungsschlüssel, Lebensweise, Verbreitung, 1–327. Hackenheim: ConchBooks.
Gobanz, J. 1854. Die fossilen Land- und Süßwasser-Mollusken des Beckens von Rein in Steiermark. Sitzungsberichte der kaiserlichen Akademie der Wissenschaften, mathematisch-naturwissenschaftliche Classis 13: 180–201.
Golikov, A.N., and Y.I. Starobogatov. 1975. Systematics of prosobranch gastropods. Malacologia 15: 185–232.
Gray, J.E. 1840. Shells of molluscous animals. In Synopsis of the contents of the British Museum, 42nd ed, 105–152. London: G. Woodfall.
Gray, J.E. 1847. A list of the genera of recent Mollusca, their synonyma and types. Proceedings of the Zoological Society of London 15: 129–242.
Gray, J.E. 1854. A revision of the arrangement of the families of bivalve shells (Conchifera). Annals and Magazine of Natural History (series 2) 13(77): 408–418.
Gray, J.E. 1857. In A manual of the land and fresh-water shells of the British Islands, ed. W. Turton. i–xvi + 1–335. London: Longman, Brown, Green, Longmans, and Robert.
Hajos, M. 1972. Kieselgurvorkommen im Tertiärbecken von Aflenz (Steiermark). Mitteilungen der geologischen Gesellschaft in Wien 63: 149–159.
Hartmann, W. 1821. System der Erd- und Flußschnecken der Schweiz. Mit vergleichender Aufzählung aller auch in den benachbarten Ländern, Deutschland, Frankreich und Italien sich vorfindenden Arten. Neue Alpina 1: 194–268.
Harzhauser, M. 2002. Marine und brachyhaline Gastropoden aus dem Karpatium des Korneuburger Beckens und der Kreuzstettener Bucht (Österreich, Untermiozän). Beiträge zur Paläontologie 27: 61–159.
Harzhauser, M., and H. Binder. 2004. Synopsis of the late Miocene mollusc fauna of the classical sections Richardhof and Eichkogel in the Vienna Basin (Austria, Pannonian, MN9–MN11). Archiv für Molluskenkunde 133: 1–57.
Harzhauser, M., and O. Mandic. 2008. Neogene lake systems of Central and South-Eastern Europe: faunal diversity, gradients and interrelations. Palaeogeography, Palaeoclimatology, Palaeoecology 260: 417–434.
Hofmann, A. 1893. Die Fauna von Göriach. Abhandlungen der kaiserlichen und königlichen geologischen Reichsanstalt 15: 1–67.
Hölzel, M., and M. Wagreich. 2004. Sedimentology of a Miocene delta complex: the type section of the Ingering Formation (Fohnsdorf Basin, Austria). Austrian Journal of Earth Science 95(96): 80–86.
Jeffreys, J.G. 1830. A synopsis on the testaceous pneumonobranchous Mollusca of Great Britain. Transactions of the Linnean Society of London 16: 323–392.
Jekelius, E. 1932. Fauna Neogenă a României. Die Molluskenfauna der dazischen Stufe des Beckens von Braşov. Memoriile Institutului Geologic al României 2: 1–118.
Jekelius, E. 1944. Sarmat und Pont von Soceni (Banat). Memoriile Institutului Geologic al României 5: 1–167.
Jörger, K.M., I. Stöger, Y. Kano, H. Fukuda, T. Knebelsberger, and M. Schrödl. 2010. On the origin of Acochlidia and other enigmatic euthyneuran gastropods, with implications for the systematics of Heterobranchia. BMC Evolutionary Biology 10: 323. doi:10.1186/1471-2148-10-323.
Jurišić-Polšak, Z., K. Sakač, and M. Poje. 1997. New pliopleistocene gastropods from Lika, Croatia. Natura Croatica 6: 91–111.
Klein, R. 1847. Conchylien der Süßwasserkalkformation Wüttembergs. Jahreshefte des Vereins für Vaterländische Naturkunde in Württemberg 2: 60–116.
Klein, R. 1853. Conchylien der Süßwasserkalkformation Wüttembergs. Jahreshefte des Vereins für vaterländische Naturkunde in Württemberg 9: 203–223.
Kókay, J. 1966. Geologische und paläontologische Untersuchungen des Braunkohlengebietes von Herend-Márkó (Bakony-Gebirge, Ungarn). Geologica Hungarica, Series Palaeontologica 36: 1–147.
Kókay, J. 2006. Nonmarine mollusc fauna from the Lower and Middle Miocene, Bakony Mts., W Hungary. Geologica Hungarica, Series Palaeontologica 56: 1–196.
Koken, E. 1896. Die Leitfossilien; ein Handbuch für den Unterricht und das Bestimmen von Versteinerungen, 1–184. Leipzig: Ch.H. Tauchnitz.
Kowalke, T., and B. Reichenbacher. 2005. Early Miocene (Ottnangian) Mollusca of the Western Paratethys ontogenetic strategies and palaeo-environments. Geobios 38: 609–635.
Kuiper, J.G.J. 1972. Sphaeriidenfunde in den pliozänen Deckschichten der rheinischen Braunkohle. Archiv für Molluskenkunde 102(1/3): 125–130.
Lamarck, J.-B.P.A. de M. de. 1809. Philosophie Zoologique, ou exposition des considérations relatives à l’histoire naturelle des Animaux; à la diversité de leur organisation et des facultés qu’ils en obtiennent; aux causes physiques qui maintiennent en eux la vie et donnent lieu aux mouvemens qu’ils exécutant; enfin à celles qui produisent les unes le sentiment, et les autres l’intelligence de ceux qui en sont doués. 1: 1–428, 2: 1–475. Paris.
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decima, reformata 1, 1–824. Holmiae: Impensis Direct. Laurentii Salvii.
Mandic, O., A. De Leeuw, B. Vuković, W. Krijgsman, M. Harzhauser, and K.F. Kuiper. 2011. Palaeoenvironmental evolution of Lake Gacko (NE Bosnia and Herzegovina): impact of the Middle Miocene climatic optimum on the dinaride lake system. Palaeogeography, Palaeoclimatology, Palaeoecology 299: 475–492.
Montfort, D. de. 1810. Conchyliologie systématique, et classification méthodique des coquilles; offrant leurs figures, leur arrangement générique, leurs descriptions caractéristiques, leurs noms; ainsi que leur synonymie en plusieurs langues. Ouvrage destiné à faciliter l’étude des coquilles, ainsi que leur disposition dans les cabinets d’histoire naturelle. Coquilles univalves, non cloisonnées. 2: 1–676. Paris: Schœll.
Morgan, J.A.T., R.J. DeJong, Y. Jung, K. Khallaayoune, S. Kock, G.M. Mkoji, and E.S. Loker. 2002. A phylogeny of planorbid snails, with implications for the evolution of Schistosoma parasites. Molecular Phylogenetics and Evolution 25: 477–488.
Morton, J.E. 1955. The functional morphology of the British Ellobiidae (Gastropoda Pulmonata) with special reference to the digestive and reproductive system. Philosophical Transactions of the Royal Society of London B 239: 89–160.
Müller, O.F. 1773–1774. Vermium terrestrium et fluviatilium historia, seu animalium Infusoriorum, Helminthicorum et Testaceorum non marinorum succincta historia. 1: 1–135, 2: 1–214. Havniæ, Lipsiæ: Heineck & Faber.
Müller, O.F. 1781. Geschichte der Perlen-Blasen. Der Naturforscher 15: 1–20.
Nalugwa, A., A. Jørgensen, S. Nyakaana, and T.K. Kristensen. 2010. Molecular phylogeny of Bulinus (Gastropoda: Planorbidae) reveals the presence of three species complexes in the Albertine Rift freshwater bodies. International Journal of Genetics and Molecular Biology 2: 130–139.
Neumayr, M. 1869. II. Beiträge zur Kenntniss fossiler Binnenfaunen. Jahrbuch der kaiserlichen und königlichen geologischen Reichsanstalt 19: 355–382.
Neumayr, M. 1880. V. Tertiäre Binnenmollusken aus Bosnien und der Hercegovina. Jahrbuch der kaiserlichen und königlichen geologischen Reichsanstalt 30: 463–486.
Neumayr, M. 1883. Zur Morphologie des Bivalvenschlosses. Sitzungsberichte der Akademie der Wissenschaften Wien 88: 385–419.
Noulet, J.-B. 1854. Mémoires sur les coquilles fossiles des terrains d’eau douce du Sud-Ouest de la France, 1–127. Paris: Editions Masson.
Noulet, J.-B. 1857. Coquilles fossiles nouvelles des terrains d’eau douce du Sud-Ouest de la France, 1–24. Paris: Editions Masson.
Papp, A. 1953. Die Molluskenfauna des Pannon des Wiener Beckens. Mitteilungen der geologischen Gesellschaft in Wien 44: 85–222.
Pavlović, P.S. 1928. Les Mollusques du Pontien Inférieur des environs de Beograd. Annales Géologiques de la Péninsule Balkanique 9: 1–14.
Pfeiffer, C. 1821. Naturgeschichte deutscher Land- und Süsswasser-Mollusken. Erste Abtheilung. i–x + 1–134 (1–2). Weimar: Landes-Industrie-Comptoir.
Pfleger, V. 1984. Schnecken und Muscheln Europas. Land- und Süßwasserarten. 1–192. Stuttgart: Kosmos, Gesellschaft der Naturfreunde, Franckh’sche Verlagshandlung.
Plaziat, J.-C., and W.R. Younis. 2005. The modern environments of Molluscs in southern Mesopotamia, Iraq: a guide to paleogeographical reconstructions of quaternary fluvial, palustrine and marine deposits. Carnets de Géologie, article 2005/01 (CG2005_A01).
Poli, J.X. 1791. Testacea utriusque Siciliae eorumque historia et anatome tabulis aeneis illustrata. Tomus primus, 1–74. Parmae: Regio Typographeio.
Ponder, W.F., and D.R. Lindberg. 1997. Towards a phylogeny of gastropod molluscs; an analysis using morphological characters. Zoological Journal of the Linnean Society 119: 83–265.
Prashad, B. 1925. Notes on the lamellibranchs in the Indian Museum. 6. Indian species of the genus Pisidium. Records of the Indian Museum 27: 405–422.
Rafinesque, C.S. 1815. Analyse de la nature ou tableau de l’univers et des corps organisés. Le nature es mon guide, et Linnéus mon maître. 1–224. Palermo, Italy (privately published).
Ratschbacher, L., W. Frisch, H.-G. Linzer, and O. Merle. 1991. Lateral extrusion in the Eastern Alps. 2. Structural analysis. Tectonics 10: 257–271.
Reischenbacher, D., and R.F. Sachsenhofer. 2002. Das Miozän des Aflenzer Beckens. PanGeo Austria, Programm and Kurzfassungen, 144–145. Salzburg: Institut für Geologie und Paläontologie.
Reuss, A.E. 1852. Beschreibung der fossilen Ostracoden und Mollusken der tertiären Süsswasserschichten des nördlichen Böhmens. Palaeontographica 2: 16–42.
Roman, F. 1910. Faune saumâtre du Sannoisien du Gard. Bulletin de la Société Géologique de France 4(2/10): 927–955.
Rzehak, A. 1893. Die Fauna der Oncophora Schichten Mährens. Verhandlungen des naturforschenden Vereins in Brünn 31: 142–192.
Sachsenhofer, R.F., A. Bechtel, D. Reischenbacher, and A. Weiss. 2003. Evolution of lacustrine systems along the Miocene Mur-Mürz fault system (Eastern Alps, Austria) and implications on source rocks in pull-apart basin. Marine and Petroleum Geology 20: 83–110.
Sandberger, F. 1870–1875. Die Land- und Süßwasser-Conchylien der Vorwelt. 353–616 (1874), 617–1000 (1875). Wiesbaden: Kreidel.
Say, T. 1817. Descriptions of seven species of American fresh water and land shells, not noticed in the systems. Journal of the Academy of Natural Sciences of Philadelphia 1: 13–18.
Schlickum, W.R. 1960. Die Gattung Nematurella Sandberger. Archiv für Molluskenkunde 89: 203–214.
Schlickum, W.R. 1964. Die Molluskenfauna der Süßbrackwassermolasse Niederbayerns. Archiv für Molluskenkunde 93: 1–68.
Schlickum, W.R. 1965. Zur Gattung Euchilus Sandberger. Archiv für Molluskenkunde 94: 99–104.
Schlickum, W.R. 1966. Die Molluskenfauna der kirchberger Schichten des Jungholzes bei Leipheim/Donau. Archiv für Molluskenkunde 95: 321–335.
Schlickum, W.R. 1970. Zur Molluskenfauna der Brackwassermolasse Niederbayerns, 3. Was ist Lymnaea bouilleti Michaud? Archiv für Molluskenkunde 100: 89–94.
Schlickum, W.R. 1971. Zur Systematik fossiler Hyroboiiden. Archiv für Molluskenkunde 101: 159–165.
Schlickum, W.R. 1976. Die in der pleistozänen Gemeindekiesgrube von Zwiefaltendorf a. d. Donau abgelagerte Molluskenfauna der Silvanaschichten. Archiv für Molluskenkunde 107: 1–31.
Schlickum, W.R. 1978. Die Gattung Tournouerina Schlickum. Archiv für Molluskenkunde 108: 237–243.
Schütt, H. 2000. Die Höhlenmollusken der Ombla-Quelle. (The subterranean molluscs of the Ombla spring). Natura Croatica 9: 203–215.
Schütt, H., and H. Besenecker. 1973. Eine Molluskenfauna aus dem Neogen von Chios (Agäis) Archiv für Molluskenkunde 103: 1–29.
Smith, E.A. 1877. On the shells of Lake Nyasa, and on a few marine species from Mozambique. Proceedings of the Zoological Society of London 1877: 712–722.
Stimpson, W. 1865. Researches upon the Hydrobiinae and allied forms: chiefly made from materials in the Museum of the Smithsonian Institution. Smithsonian Miscellaneous Collections 7: 1–59.
Stur, D. 1871. Geologie der Steiermark. Erläuterungen zur geologischen Uebersichtskarte des Herzogthumes Steiermark, 1–654. Graz: Geognostisch-montanistischer Verein für Steiermark.
Tausch, L.v. 1889. Ueber einige nichtmarine Conchylien der Kreide und des steirischen Miocäns und ihre geographische Verbreitung. Verhandlungen der k.k. geologischen Reichsanstalt 23: 157–158.
Theobald, W. 1878. Notes on the land and freshwater shells of Kashmir, more particularly of the Jhelum valley below Srinagar and the hills north of Jammu. Journal of Asiatic Society of Bengal 47: 141–149.
Tournouër, R. 1879. Conchyliorum fluviatilium fossilium, quæ in stratis tertiariis superioribus Rumaniæ Dr. Gregorio Stefanesco collegit, novæ species. Journal de Conchyliologie 27: 261–264.
Unger, F. 1858. Über fossile Pflanzen des Süsswasser-Kalkes und Quarzes. Denkschriften der k. Akademie der Wissenschaften 14: 1–38.
Walker, B. 1903. Notes on eastern American Ancyli. The Nautilus 17(13–19): 25–31.
Wank, M. 1991. Die Fossilien von Schönweg bei St. Andrä im Lavanttal (Kärnten). Carinthia II 181: 301–313.
Wautier, J. 1975. Présence d’espèces du genre Ferrissia Walker, 1903 (Gastropoda, Basommatophora) dans le Néogène du Bassin Rhodanien (France). Geobios 8: 423–433.
Wenz, W. 1938–1944. Teil 1: allgemeiner Teil und Prosobranchia. In Handbuch der Paläozoologie, Band 6, Gastropoda, ed. O.H. Schindewolf, 1–1639. Berlin: Verlag Gebrüder Bornträger.
Acknowledgments
We thank Ulrich Bößneck (University of Giessen), Daniela Esu and Odoardo Girotti (both University of Rome), Olga Anistratenko (Institute of Geological Sciences of NAS, Ukraine) and Vitaly Anistratenko (Schmalhausen Institute of Zoology of NAS, Ukraine) for fruitful discussions. We are greatly indebted to Dietrich Kadolsky (Geoconsultants Limited, 66 Heathhurst Road, Sanderstead, Surrey, GB) for his help in identifying the hydrobiid Romania.The study is part of the FWF-grant P18519-B17 “Mollusc Evolution of the Neogene Dinaride Lake System” and contributes to the FWF-grant P21414-B16 and to the NECLIME project.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Harzhauser, M., Neubauer, T.A., Mandic, O. et al. A Middle Miocene endemic freshwater mollusc assemblage from an intramontane Alpine lake (Aflenz Basin, Eastern Alps, Austria). Paläontol Z 86, 23–41 (2012). https://doi.org/10.1007/s12542-011-0117-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-011-0117-x