
Abstract
The new species Cladocarpus salix and three species of Lytocarpia (L. brevirostris, L. delicatula and L. phyteuma) were recorded from the Bunaken National Marine Park (North Sulawesi, Indonesia). Cladocarpus salix forms large colonies reminiscent of willow trees and differs from the closely similar species C. keiensis in the shape and the size of the intrathecal septum, and in the phylactocarp structure. Moreover, the presence of grooves on the external wall of theca is unique feature of this species. The differences of the three Lytocarpia species are discussed using microscopic imaging, comparison of measurements, and macroscopic features of living specimens. The bathymetric distribution of these species, studied through vertical transect, showed that C. salix and L. delicatula are deep species occurring below 20 m and increasing their abundance from 30 to 50 m depth; L. phyteuma is more abundant from 5 to 30 m and L. brevirostris is more abundant in the shallowest and deepest part of the reef and rarer between 10 and 30 m depth. A series of surveys conducted in the same locality during a year indicated that these species are generally more abundant from November to May (wet season), except for L. delicatula which is active in July.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The aim of this work was to describe some Aglaopheniidae from the coral reef of the Bunaken Marine Park (North Sulawesi, Indonesia). In particular, data on the morphology and ecology of a new species of Cladocarpus and three species of Lytocarpia (L. brevirostris, L. delicatula and L. phyteuma) are given.
The systematics of the genus Cladocarpus is quite complex because of the high morphological variability of the gonosome and the presence of characters shared with other similar genera (e.g. Streptocaulus and Cladocarpoides). According to Bouillon et al. (2006), the genus Cladocarpus includes 65 species and it is characterised by appendages of the hydrocladia which protect the gonothecae, the so-called phylactocarps. Most species show a long, S-shaped hydrotheca and a short, median inferior nematothecae not reaching the thecal margin. One species, erroneously classified as Macrorhynchia protecta (Antsulevich 1991), should be attributed to Cladocarpus (Di Camillo et al. 2009; Vervoort and Watson 2003).
Ramil and Vervoort (1992) suggested the genus Cladocarpus contains two groups of species characterised by a different morphology of the phylactocarps. However, due to the high variability of these structures, Schuchert (2003) and Bouillon et al. (2006) suggested that the genus needs a more comprehensive and deep-going investigation before taxonomic re-arrangements can be made.
Cladocarpus species occur from boreal to tropical seas, although more than 50% of the species are concentrated in temperate and tropical waters of the Atlantic. About 20% of the species have been recorded from the Pacific Ocean and 20% from South Africa, while the species from the Indian Ocean represent less than 8%. Four Atlantic species have been also found in the Mediterranean, one near Marseille and three in the Alboran Sea.
The genus Lytocarpia includes 36 species, occurring in the Pacific Ocean (more than 50%), Atlantic Ocean (16%), Indian Ocean (20%) and South Africa (11%). One species is known from the Antarctic, one from the Arctic, and two from the Mediterranean Sea. This genus differs from the very similar Aglaophenia by the presence of a hydrotheca on the corbula ribs in at least one sex. Corbulae of Lytocarpia generally have sturdy and fused ribs, even though open ribs can also be observed. Pseudophylactocarps may occasionally occur in some species as in Lytocarpia angulosa and Lytocarpia perarmata (Billard 1913); these modified hydrocladia, which are not associated with reproductive structures, present large nematothecae and probably prevent the settlement of potential epibionts (Watson 2000).
Here we describe aspects of living colonies of Cladocarpus salix n. sp., Lytocarpia brevirostris (Busk, 1852), Lytocarpia delicatula (Busk, 1852), and Lytocarpia phyteuma (Kirchenpauer 1876). We also provide scanning electronic microscope (SEM) pictures of the main characteristics of these species.
The collection along vertical transects during a year allowed us to summarise the bathymetric and seasonal occurrences of each species.
Materials and methods
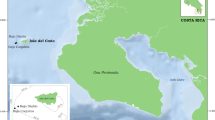
Hydroids were collected by SCUBA diving from May 2002 to October 2008 in several sites of the Bunaken Marine Park (North Sulawesi, Indonesia) from a depth of 2.5–60 m (Fig. 1). The area is characterised by a vertical coral reef with canyons and caves invested by a strong current and densely populated with benthic organisms. In particular, the reef shows an well-lit edge dominated by hardy corals and ends with a gentle slope at a depth of 50–60 m. Hydroids represent one of the richest communities from 10 to 30 m, living both on dead corals and secondary substrates.

Map of sampling sites in the Bunaken Marine Park (North Sulawesi). 1 Siladen, 2 Siladen Timur, 3 Bualo, 4 Negeri, 5 Depan Kampung, 6 Likuan, 7 Mandolin, 8 Raymond’s Point, 9 Mike’s Point, 10 Bunaken Timur, 11 Mapia
Colonies of Cladocarpus and Lytocarpia were photographed in situ before being removed and placed in plastic bags, then were examined in vivo to observe the morphology and colour of living specimens and fixed using 4% neutralised formaldehyde. The details of the colonies were drawn and photographed using light microscopy; only larger cnidocysts were measured and drawn.
Photographs were also taken under the SEM (Philips XL20). Samples were washed with distilled water, then dehydrated in a graded ethanol series, dried at the Critical Point Dryer and coated with gold-palladium in a Balzer Union evaporator.
In order to study the bathymetric and temporal distribution of Cladocarpus and Lytocarpia species, samplings were conducted bimonthly from May 2004 to February 2005 in Siladen, a small coralline island surrounded by a fringing reef. As previously described by Di Camillo et al. (2008), three vertical transects, at a distance of about 50 m from each other, were considered. In each transect, samples were collected by diving, at seven depth-ranges: lagoon, 2.5–5 m, 5–10 m, 10–20 m, 20–30 m, 30–40 m and 40–50 m. At each depth all the species found by two divers working together were collected and put in plastic bags. Hydroids were identified and an abundance value varying from 0 (absent) to 3 (very abundant) was assigned to each species.
Voucher samples of the described species were deposited at the Museum of Natural History of Genoa (MSNG).
Results
Taxonomy
Family Aglaopheniidae Agassiz, 1862
Genus Cladocarpus Allman, 1874
Cladocarpus salix n. sp.
Figures 2a, b; 3a; 4a-g; 5a-g; 6a, b.

Comparison of living specimens (underwater pictures). a Cladocarpus salix, picture of a large colony, Bualo; b close view of the same colony showing branches bearing gonophores. c Lytocarpia brevirostris, branched colony, Siladen Timur. d Lytocarpia delicatula, notice white hydrocladia, Siladen. e Lytocarpia phyteuma, several colonies on a coral, Depan Kampung; f particular of a colony, Mike’s Point

Comparison of living specimens (stereomicroscope pictures). a Cladocarpus salix, portion of a colony with the delicate and flexible hydrocladia. b Lytocarpia brevirostris, branched colony. c Lytocarpia delicatula, note the curved hydrocladia. d Lytocarpia phyteuma, fertile colony with widely spaced hydrocladia. e L. phyteuma, detail of a corbula, note the white, refringent spot in correspondence of nematothecae. f Comparison of the three Lytocarpia species, note the different hydrocladia: 1 L. brevirostris; 2 L. delicatula; 3 L. phyteuma

Cladocarpus salix: colony (a), detail of hydrocaulus and hydrocladia (b) and hydrocladium in lateral view (c); d hydrotheca in frontal view; e female phylactocarp; f male phylactocarp; g microbasic mastigophore. Scale bars a 1 cm; b 1 mm; c, d 100 μm; e, f 200 μm; g 10 μm

Cladocarpus salix (SEM pictures): hydrotheca in frontal (a) and lateral (b, c) views; note the median tooth on the thecal rim and the characteristic external grooves on the thecal wall (c); the inset shows the inner side of the thecal wall with the protruding membranes of the grooves. d–f Branch bearing hydrocladia and phylactocarps. Some hydrocladial apophyses lacking of hydrocladia (d), detail of a hydrocladium and a phylactocarp bearing one hydrotheca and two gonothecae (e). Note the nematothecae present throughout the entire length of primary and auxiliary tubes and gonothecae both on phylactocarps and on hydrocladial apophyses (f). g Detail of the first segment of the phylactocarp with the gonotheca inserted on the gonocladial apophysis. Scale bars a–c, e, g, h 200 μm; d 1 mm; f 500 μm, inset 20 μm

Optical microscope pictures of the species. a Cladocarpus salix, hydrocladia with several hydrothecae. b Higher magnification with a hydrotheca and the characteristic grooves on the wall. c Cladocarpus keiensis, (MHNG INVE 32487, schizotype, Indonesia, Kei Island, 100 m), hydrocladium and particular of internal septum and the looped ridge (inset). d-f Lytocarpia brevirostris, Lytocarpia delicatula and Lytocarpia phyteuma, hydrothecae in lateral view. Scale bars a–c 200 μm; d–f 100 μm
Material examined
Bunaken National Marine Park, North Sulawesi, (Indonesia): Bualo, 20/30 m (male sample collected in February 2004; female sample collected in November 2006). —Siladen, 20/50 m.
Type material
Holotype MSNG 55844, Bualo (Bunaken Marine Park, Indonesia), 20-30 m, formalin-preserved male colony, coll. C.G. Di Camillo (SCUBA), Feb. 2004.
Paratype MSNG 55845, Bualo (Bunaken Marine Park, Indonesia), 28 m, formalin-preserved female colony, coll. C.G. Di Camillo (SCUBA), Nov. 2006.
Other material examined
MNHG INVE 32487 Cladocarpus keiensis from Kei Islands 100 m, schizotype, two slide preparations, Natural History Museum of Geneva, material described in Schuchert (2003)
Diagnosis: large colonies with drooping hydrocladia; elongated hydrothecae with a sharp tooth on the abcauline side and with two characteristic external grooves with converging ends visible in lateral view. A very small intrathecal septum originates from the adcauline side of the theca. Phylactocarps homologous to the hydrocladia; gonothecae observed both on the lower segments of gonocladium and hydrocladial apophyses.
Description
Colonies large branching up to third order in one plane, with downward direct hydrocladia (Figs. 2a, b; 3a; 4a). Height up to 20 cm, stem and proximal parts of first-order branches deprived of hydrocladia. Stem and main branches strongly polysiphonic, composed by a bundle of straight tubes (Fig. 4b), hydrocladia only on the primary tube. Distal part of the stem and main branches with a reduced number of auxiliary tubes, terminal portion of each tube monosiphonic. Primary tubes of first-order branches originating from primary tube of the stem; primary tubes of the branches of second, third and fourth-order arising, respectively, from primary tube of first, second and third-order branches. Auxiliary and primary tubes bear nematothecae throughout their entire length. Primary tubes only occasionally or never segmented, indistinct internodes bearing an apophysis, a mamelon and two to four nematothecae, one associated to the apophysis and two to three in the lower half of the segment. Hydrocladia on primary tubes alternately arranged on anterior surface of apophyses.
Hydrocladia in one plane in the distal, younger portion of the branches and arranged at variable angles in the proximal oldest part.
Hydrocladia segmented by oblique nodes, each internode with one hydrotheca and three nematothecae: two laterals and one median inferior. Internode divided in six to seven short segments by five to six semicircular ribs (Fig. 4c).
Hydrothecae elongated, adcauline wall completely adnate, abcauline wall slightly sinuous. Margin of theca slightly oblique and with only one tooth on the abcauline side (Figs. 4d; 5a–c). The tooth is triangular and very sharp. A very short intrathecal septum originates from the adcauline side of the theca somewhat proximal of the second rib of the internode (Figs. 4c; 6a, b). Hydrothecae in lateral view show two characteristic external grooves reaching beyond the middle of the hydrotheca (Figs. 4c; 6 b), one groove arising from the adcauline wall and the other from the abcauline side near the median inferior nematotheca. Abcauline groove longer than the adcauline one (Fig. 5c). Grooves sinuous, oblique, distal ends converging in the middle of the hydrothecal wall. From each groove, very thin, short membranes project towards the lumen of the hydrotheca (Fig. 5c inset).
Lateral nematotheca tubular, lower portion slightly oblique and fused to the adcauline side of the theca; an opening is visible on the inner side of the nematotheca; upper portion straight and overtopping the thecal margin; terminal opening circular and with a deep emargination on the inner wall. Median inferior nematotheca present at the base of the hydrotheca, lower half part adnate, upper half part free and gutter-shaped.
Gonothecae borne on apophyses of both hydrocladia and phylactocarps; generally one gonotheca is present on the hydrocladial apophyses and up to three on the phylactocarp. Phylactocarps arise from the first internode of the hydrocladium near to the median inferior nematotheca; they are homologous to the hydrocladia but always shorter and composed by segments of variable length. Shorter segments with one nematotheca only; longer segments with a hydrotheca or with a gonotheca. Segments bearing hydrothecae (one or two) identical to the hydrocladial ones; those with gonothecae present one nematotheca below the gonocladial apophysis and one in the upper part of the segment directed outwards. Gonothecae ovoid in shape, male ones with a clearly distinguishable spadix, female ones with two to four eggs inside (Figs. 4e, f; 5d-g).
Colours: thicker, polysiphonic parts are brown while distal parts are yellow; male gonothecae yellow with an orange spadix, female gonothecae yellow.
Sizes (see Table 1).
Cnidocysts: Ovoidal microbasic mastigophores 17.5-20 × 6-6.5 μm present in coenosarcs and nematophores.
Etymology: The specific name refers to the colony shape resembling a willow tree for its drooping hydrocladia.
Remarks: Cladocarpus salix strongly differs from the other species belonging to the same genus in theca and phylactocarp shape; the presence of external grooves on the theca—present both on living and preserved specimens, as well as on samples analysed with electron microscope—is a unique feature of this species. The species resembling Cladocarpus salix most closely is Cladocarpus keiensis Schuchert, 2003. This author described an adcauline septum inside the hydrotheca of C. keiensis, spanning half of the hydrothecal diameter, scoop-shaped, and directed obliquely upward. Besides, the intrathecal septum appears triangular in side view as shown in pictures taken from Kei Island material (Figure 6c). Hydrothecae of Cladocarpus salix present a very small intrathecal septum, which is hardly visible and in lateral view of the hydrotheca as it is usually concealed by the grooves it originates from.
The septum described by Schuchert, continues as a looped ridge to the abcauline wall (Fig. 6c inset), while C. salix presents two external grooves with converging ends.
C. salix differs from C. keiensis also by the presence of gonothecae on the hydrocladial apophyses. Moreover, in our material the gonothecae are placed only on the lower segments of gonocladium, while in C. keiensis segments with hydrothecae are alternated with two to three segments bearing gonothecae.
Genus Lytocarpia Kirchenpauer, 1872 = Acanthocladium Allman, 1883 = Thecocarpus Nutting, 1900
Key to Lytocarpia species from North Sulawesi
-
1
Thecal margin with five teeth, hydrothecae delicate, hydrocaulus brown and hydrocladia white, with prosegment and one or two hinge-joints, corbulae open.......…...…….…........................…...…Lytocarpia delicatula
-
1
Thecal margin with seven teeth…................................…2
-
2
Hydrocaulus yellow-brown, hydrocladia yellow; hydrothecae with abcauline wall sinuous, with a deep fold just below marginal teeth; margin inclined towards below, corbulae closed and short......……Lytocarpia brevirostris
-
2
Hydrocaulus light brown, hydrocladia yellow, hydrothecae tubular, hydrocladia generally wide spaced, hydrothecal wall with an undulated and oblique intrathecal thickening, corbulae closed andelongated......................................Lytocarpia phyteuma
Lytocarpia brevirostris (Busk, 1852)
Figures 2c; 3b, f; 6d; 7a-g; 8a-k.

Lytocarpia brevirostris. Colony with long (a) and short (b) hydrocladia; c detail of an internode with apophyses and associated nematothecae and mamelons; d hydrocladium in lateral view; e hydrotheca in frontal view; f microbasic mastigophores; g corbula. Scale bars a, b 1 cm; c 200 μm; d, e 100 μm; f 10 μm; g 500 μm

SEM picture of Lytocarpia brevirostris. a Hydrocaulus and alternate hydrocladia and details (b); c higher magnification of the internodes with the hydrocladial apophyses; hydrocladium in lateral (d) and frontal (e) views; particular of a hydrotheca and associated nematothecae in lateral (f) and frontal (g) views; h corbula; i detail of hydrothecae inserted in the spaced between crests and ribs of a corbula; j coalesced corbulae on the same branch; k detail of corbula ribs with gutter-shaped nematothecae. Scale bars a, b, h 500 μm; c–e 200 μm; f, g, i 100 μm; j 1 mm; k 50 μm
Plumularia brevirostris Busk, 1852: 397.
Aglaophenia brevirostris—Kirchenpauer, 1872: 27; Bale, 1884: 169; Kirkpatrick, 1890: 611; Bale, 1913: 135, pl. 13, figs. 7, 9.
Plumularia vitana Kirchenpauer, 1872: 34, pl. 1 fig. 9, pl. 3 fig. 9.
Aglaophenia maldivensis Borradaile 1905: 843, pl 69, fig. 8; Billard, 1913: 91
Thecocarpus brevirostris—Billard, 1910: 51, fig. 24; 1913: 89, fig. 75; Briggs, 1918: 34, 45; Stechow, 1919: 137, figs. A2, B2; Bedot 1921a: 332; Bedot 1922: 157; Jarvis, 1922: 350, pl. 26, fig. 24; Pennycuik, 1959: 187; Rees and Thursfield, 1965: 186; Millard, 1968: 254, 284-285, fig. 6B; Vasseur, 1974: 158; Millard, 1975: 454, fig. 139A-C; 1978: 199; Tang, 1991: 32-33, Fig. 6.
Lytocarpia brevirostris—Ryland and Gibbons, 1991: 545-546, fig. 15.
Material examined
Bunaken National Marine Park, North Sulawesi, (Indonesia): MSNG 55846, Mandolin (Bunaken Marine Park, Indonesia), 5/10 m, formalin-preserved fertile colony, coll. C.G. Di Camillo (SCUBA), Feb. 2005. Mandolin, 25 m (fertile sample collected in February 2005). —Raymond’s Point, 16 m (fertile sample collected in January 2006). —Siladen, 2.5/50 m. —Siladen Timur, 20 m.
Description
Colonies pinnate, reaching 5 cm; stem polysiphonic, occasionally branched up to the second order (Figs. 2c; 3b, f; 7a, b); unbranched in young colonies. Polysiphonic parts composed of a superficial primary tube and some auxiliary tubes. Branches arising from primary tubes, alternately arranged and in one plane; basal portion of ramifications generally polysiphonic.
Auxiliary tubes unsegmented, primary tubes segmented in distal parts only. Internodes with a hydrocladial apophysis associated with a mamelon and two nematothecae: one inferior anterior and one axillary anterior; nematothecae large, with double opening, mamelon on the anterior surface of the apophysis (Figs. 7c; 8c).
Hydrocladia alternately arranged on the anterior surface of the hydrocaulus (Fig. 8a, b); hydrocladia of variable length and homomerously segmented by oblique nodes. Hydrothecae on hydrocladium overlapping (Figs. 6d; 7d; 8d, e).
Hydrothecae elongated, (Figs. 7e; 8f, g); adcauline wall almost straight, abcauline wall sinuous, with a deep fold just below marginal teeth; margin inclined towards below, with six lateral teeth, and a prominent abcauline tooth, adcauline tooth absent.
Hydrothecae with a short intrathecal septum in the lower half of the adcauline side.
Median inferior nematotheca adnate to the hydrotheca for about half-length and not reaching the hydrothecal margin; adnate portion tubular, free portion gutter-shaped and bent away from theca, foramen into hydrotheca; lateral nematothecae gutter-shaped and directed upwards.
Corbulae closed, replacing a hydrocladium, bearing six to eight pairs of leaf-like ribs (Figs. 7g; 8h-j); sometimes one or two ribs can be lifted. Corbula axis divided in segments by oblique nodes; first segment as the other hydrocladial internodes, with a hydrotheca and three nematothecae. Proximal portion of each rib showing a basal crest supporting one hydrotheca with lateral nematothecae and without a median inferior nematotheca; hydrotheca inserted in the space (defined as fenêtre by Billard 1913) between the crest and the rib; two nematothecae present just above the crest. Ribs with several nematothecae arranged in rows, all nematothecae gutter-shaped (Fig. 8k). Distal extremity ending with one or two hydrothecate segments.
Remarks: L. brevirostris from Bunaken occasionally produces fused corbulae on adjacent hydrocladia (Fig. 8j ).
Rarely, the ventral sides of the colony in the upper and the lower part of the colony face in opposite directions, an observation also reported by Millard, 1975.
Colour: Hydrocaulus yellow-brown, hydrocladia yellow.
Sizes (see Table 1).
Cnidocysts: Elongated microbasic mastigophores 16.25-17.50 × 2.5 μm present in coenosarcs and nematophores.
Distribution
South Pacific Islands, Northern Australia, tropical Indian Ocean from India to Africa. Type locality: off Cumberland Island, Queensland.
Lytocarpia delicatula (Busk, 1852)
Figures 2d; 3c, f; 6e; 9a-g; 10a-i.

Lytocarpia delicatula. a Colony; b detail of hydrocaulus and hydrocladia; c basal portion of the hydrocaulus; hydrotheca in frontal (d) and lateral (e) views; f corbula; g microbasic mastigophores. Scale bars a 1 cm; b, c, f 500 μm; d, e 100 μm; g 10 μm

SEM picture of Lytocarpia delicatula. a Colony; b prosegment and hinge joints on the hydrocaulus proximal portion; c internode bearing the hydrocladial apophysis and the associated nematothecae; hydrocladium in lateral (d) and in frontal (e) views; f open corbula and gonothecae; g detail of open ribs and basal hydrothecae; h distal portion of a rib bearing alternately arranged nematothecae; i detail of nematothecae on a rib. Scale bars a 2 mm; c–e, g, h 200 μm; b 500 μm, f 1 mm; i 50 μm
Plumularia delicatula Busk, 1852: 396.
Aglaophenia delicatula—Bale, 1884: 167, pl 14 fig. 4, pl 17 fig. 11; Borradale, 1905: 843, pl. 69: fig. 7; Billard, 1913: 106, fig. 95; Jäderholm 1920: 8, pl. 2 fig. 7; Jarvis, 1922: 350. Pennycuik, 1959: 185; Watson, 2000: 57, fig. 46A-E.
Thecocarpus delicatulus—Millard and Bouillon, 1973: 94, fig. 11J-K; Millard, 1975: 455, 139D-E.
Lytocarpia delicatula—Schuchert, 2003: 235-237, fig. 76.
Material examined
Bunaken National Marine Park, North Sulawesi, (Indonesia): MSNG 55847, Mapia (Bunaken Marine Park, Indonesia), 25 m, formalin-preserved unfertile colony; coll. C.G. Di Camillo (SCUBA), Jan. 2003. —Mandolin, 20/30 m (fertile sample collected in February 2005). —Mapia, 20/60 m. —Mike’s Point, 30/40 m. —Siladen, 20/50 m.
Description
Colonies pinnate, reaching 10 cm in height (Figs. 2d; 3c, f; 9a); hydrocaulus unbranched (Figs. 9b; 10a), monosiphonic. With prosegment and one or two hinge-joints in the basal part (Figs. 9c; 10b). Proximal portion of each hinge-joint with a couple of large nematothecae. Part of the stem below the hinge-joints with up to 11 segments bearing couples of nematothecae. Segments above hinge-joints separated by slightly oblique nodes and associated with an apophysis, a mamelon on its anterior surface, and three nematothecae: one inferior anterior and one flanking the apophysis (Fig. 10c).
Hydrocladia alternately arranged on the anterior surface of the hydrocaulus, divided in internodes by slightly oblique nodes and bearing up to 22 hydrothecae (Fig. 10d, e). Two weakly developed internal ribs visible in each internode. Hydrothecae on hydrocladia little spaced (Fig. 6e), vase-shaped, delicate, narrowing in the basal part; adcauline wall oblique; margin inclined towards below, with five triangular teeth, without adcauline tooth; hydrothecal walls showing an oblique intrathecal thickening spanning through the hydrotheca (Fig. 9d, e).
Median inferior nematotheca adnate to the hydrotheca for about half-length and reaching the hydrothecal margin; adnate portion narrow and tubular, free portion quite enlarged, gutter-shaped, foramen into hydrotheca; lateral nematothecae directed towards outside, gutter-shaped.
Corbulae open, short and replacing a hydrocladium (Figs. 9f; 10f-i), first segment bearing a hydrotheca, following seven to eight segments with seven ribs, six of them alternate along the corbula axis and one originating from the distal segment. First segment of a rib with a nematotheca and a hydrotheca flanked by two lateral nematothecae and lacking of the median inferior nematothecae; ribs not leaf-like, thin as a phylactocarp, replacing the median inferior nematotheca. Distal portion of the rib bearing 10-14 alternately arranged nematothecae (Fig. 10h). Up to six flattened, elongated gonothecae arising from apophyses were observed.
Remarks: colonies often ending with a tendril-like process.
Colour: Hydrocauli brown, hydrocladia white.
Sizes (see Table 1).
Cnidocysts: Elongated microbasic mastigophores 26.25-27.5 × 2.5 μm present in coenosarcs and nematophores.
Distribution
Northen Australia, Great Barrier Reef, Australia, Indonesia, Seychelles, Maldive, Moçambique. Type locality: Torres Strait.
Lytocarpia phyteuma (Kirchenpauer, 1876)
Figures 2e, f; 3d, e, f; 6f; 11a-g; 12a-l.

Lytocarpia phyteuma. a Colony; b hydrocaulus and internodes; c hydrocladium in lateral view ending with a tendril-like process; hydrotheca in frontal (d) and lateral (e) view; f microbasic mastigophores; g corbula. Scale bars a 1 cm; b, c 200 μm; d, e 100 μm; f 10 μm; g 500 μm

SEM picture of Lytocarpia phyteuma. a Detail of hydrocaulus and hydrocladia; b basal part of hydrocaulus bearing solitary nematothecae; hydrocladium in frontal (c) and lateral (d) views; hydrotheca in frontal (e) and lateral (f) views; g hydrotheca of the second type showing the swelling on the lateral wall; corbula (h) and detail of ribs (i); frontal view of leaf-like ribs (j) and detail of the proximal portion of ribs (k); l higher magnification showing the hydrothecae on the corbula. Scale bars a, k 200 μm; b, d, i, j 500 μm; e, f, g, l 100 μm; h 1 mm
Aglaophenia phyteuma Kirchenpauer, 1876: 23 no. 3a.
Aglaophenia clavicula Whitelegge, 1899: 373-374, pl. 23 figs. 4-6.
Aglaophenia elongata var. sibogae Billard, 1913: 103.
Thecocarpus phyteuma—Stechow, 1919: 139-143, figs. C2, D2; Pennycuik 1959: 187; Millard and Bouillon, 1973: 95-97, fig. 11E-F.
[Not Thecocarpus phyteuma—García Corrales et al. 1978: 66-67, fig. 32; Boero and Bouillon, 1993: 263 = Aglaophenia kirchenpaueri (Heller, 1868)].
?Thecocarpus phyteuma—Vervoort and Vasseur 1977: 86.
Thecocarpus clavicula—Bedot 1921a: 333.
Thecocarpus leopoldi Leloup, 1930a: 1-3, fig. 1; 1930b: 11-14, figs. 8, 9, pl. 2 figs. 2, 3.
Lytocarpia phyteuma—Ryland and Gibbons, 1991: 548-552, figs. 18-19; Bouillon et al. 1995: 37; Watson, 2000: 65; Vervoort and Watson, 2003: 316; Schuchert, 2003: 237, Fig. 77.
Material examined
Bunaken National Marine Park, North Sulawesi, (Indonesia): MSNG 55848, Negeri (Bunaken Marine Park, Indonesia), 15 m, coll. C.G. Di Camillo (SCUBA), Feb. 2005, formalin-preserved fertile colony. —Bualo, 12 m (fertile sample collected in February 2005). —Bunaken Timur, 10 m (fertile sample collected in October 2008). —Depan Kampung, 18 m (fertile sample collected in July 2004). —Mandolin, 5/10 m (fertile sample collected in February 2005). —Siladen, 2.5/50 m (fertile sample collected in November 2004).
Description
Colonies pinnate, about 5 cm in height, stem monosiphonic and unbranched (Figs. 2e, f; 3d-f; 11a). Basal part of hydrocaulus without hinge-joints and bearing solitary nematothecae, number variable depending on stem length (Fig. 12b). Stem divided in internodes by slightly oblique nodes (Figs. 11b; 12a). Each segment with an apophysis, a mamelon and two nematothecae: one inferior anterior and one flanking the apophysis. Hydrocladia alternately arranged on anterior surface of apophyses.
Hydrocladia generally wide spaced and regularly segmented in internodes by slightly oblique nodes. Each hydrocladial segment with one hydrotheca and three nematothecae: one median inferior and two laterals. Hydrothecae on hydrocladium quite spaced (Figs. 6f; 11c; 12c, d).
Hydrothecae vase-shaped, elongated, narrowing in the basal part (Figs. 11d, e; 12e, f). Hydrotheca with an intrathecal thickening. This undulated and oblique thickening arises from the adcauline side and reaches the abcauline wall at level where the median inferior nematotheca becomes free. Another short thickening is present in the hydrocladium in correspondence of the intrathecal one. Adcauline wall completely adnate and straight up to the intrathecal thickening, then becomes slightly oblique; abcauline wall almost straight. Margin perpendicular to the main axis or slightly inclined towards below, with seven triangular teeth, without adcauline tooth.
Median inferior nematotheca not reaching the hydrothecal margin; nematothecae adnate to the hydrotheca for about half-length, adnate portion tubular, free portion quite enlarged, gutter-shaped, foramen into hydrotheca. Lateral nematothecae directed towards outside, gutter-shaped.
Corbulae elongated, closed, sometimes the distal ribs may be open; corbula replacing a hydrocladium, first segment with hydrotheca as other hydrocladial segments; corbula axis with two alternate rows of leaf-like ribs; margin of ribs with a row of gutter-shaped nematothecae; proximal part of each rib with an elongated processes supporting a hydrotheca; processes of variable length, in general with a nematotheca on the tip; processes shorter in the proximal part of the corbula and occasionally lacking of the nematotheca (Figs. 11g; 12h-l).
Remarks: colonies often ending with a tendril-like process (Fig. 2e, f).
L. phyteuma from Bunaken may form colonies slightly different from the most common type here described. We observed another form characterised by more sturdy perisarc, less-spaced branches, shorter and closer hydrothecae (Table 1). Moreover, SEM pictures of thecae reveal a swelling on the lateral walls resembling protruding cheekbones (Fig. 12g).
Colour: Hydrocaulus light brown, hydrocladia yellow. White and refringent spots visible in correspondence of lateral and median inferior nematothecae of living specimens.
Sizes (see Table 1).
Cnidocysts: Elongated microbasic mastigophores 25-30 × 5 μm present in coenosarcs and nematophores.
Distribution
Great Barrier Reef, Northern Australia, Polynesia, Indonesia, Seychelles. Polynesia. Type locality: Tonga Islands.
Ecology
The bathymetric and temporal distributions of the four studied species on the coral reef of Siladen were obtained through vertical transects in the Siladen area, studied during an annual cycle; even if the ecological study was conducted just in Siladen, data about species composition, abundance and distribution reflect those of the entire marine park. The hydroid fauna of the coral reefs of Siladen includes 107 species belonging to 51 genera and 28 families (Di Camillo et al. 2008). The family Aglaophenidae is represented by five genera and 14 species, three of which are Lytocaropia and one Cladocarpus. In this area the number of hydroid species progressively increases from the edge of the reef to 10-20 m depth and slightly decreases until 50 m depth and the highest species diversity, abundance and fertility increased during the wet season due to elevated food availability.
The large colonies of Cladocarpus salix settle on corals in shadow habitats. Two species of Lytocarpia, L. brevirostris and L. delicatula, may settle on corals but were mainly recorded on the membranous tubes of an unidentified polychaete very abundant in the Bunaken area, while L. phyteuma was only observed on the coral substratum. In other localities the species were often recorded as epibiont of several organisms, in particular even Watson (2000 and 2002) found Lytocarpia on the tube of a worm, this belonging to the genus Eunice.
The species described in this paper show wide differences in their bathymetric ranges. Cladocarpus salix and L. delicatula occur below 20 m and their abundance increases from 30 to 50 m depth. Lytocarpia phyteuma and L. brevirostris show a contrasting bathymetric distribution. The former was recorded from the edge of the reef to 50 m but it is rare in the shallowest and deepest zones of the reef and more abundant from 5 to 30 m. Lytocarpia brevirostris has also a wide vertical distribution, but it is more abundant in the two extremities of its distribution and more rare between 10 and 30 m depth. (Fig. 13).

Bathymetric distribution of Cladocarpus salix (a), Lytocarpia brevirostris (b), Lytocarpia delicatula (c) and Lytocarpia phyteuma (d)
C. salix was abundantly recorded in November and, more rarely in May, the species was never observed in the dry season (from July to September), neither in Siladen nor in other sites, and we tentatively hypothesize it is typical of the wet season. The three species of Lytocarpia were observed all around the year but, while L. brevirostris and L. delicatula spread from May to July, L. phyteuma is mainly present during the wet season. The gonotheca maturation of C. salix and L. brevirostris occurs during the wet season. L. delicatula and L. phyteuma were, on the contrary recorded fertile all year round (Fig. 14).

Temporal distribution of the described in Siladen Island. a Cladocarpus salix; b Lytocarpia brevirostris; c Lytocarpia delicatula; d Lytocarpia phyteuma
Few data are available in literature about the bathymetric and temporal distribution and fertility period of the considered Lytocarpia species. The species are more common between the reef platform and the outer slope (Gravier-Bonnet and Bourmaud 2006a, b; Vervoort and Vasseur, 1977) and show a wide bathymetric range: L. brevirostris was collected from 2 to 70 metres depth in South Africa (Millard, 1975) while L. delicatula was reported from shallow waters in Australia (Watson 2000) and up to 500 m during the Siboga expedition (Billard, 1913); even L. phyteuma is present from the surface (Fiji Islands, Ryland and Gibbons, 1991) to deeper waters (from 60 to 130 m in New Zealand, Vervoort and Watson, 2003).
The complete list of the records of the three species of Lytocarpia (Table 2) indicate for them a typical Indo-Pacific distribution (Fig. 15).

Distribution of Lytocarpia brevirostris (circles), Lytocarpia delicatula (stars) and Lytocarpia phyteuma (squares)
References
Agassiz L (1862) Contributions to the Natural History of the United States of America. Second monograph, 4, Little, Brown & Co., Boston, p 380
Allman GJ (1874) Report on the Hydroida collected during the Expeditions of H.M.S. ‘Porcupine’. Trans Zool Soc Lond 8:469–481
Antsulevich AE (1991) Novi vid gidroidov (Cnidaria, Hydroidea) iz batiali Tikhogo Okeana. A new species of hydroids (Cnidaria, Hydroidea) from the bathyal of the Pacific Ocean. Zoolog Zh 71:134–136
Bale WM (1884) Catalogue of the Australian hydroid zoophytes. Australian Museum, Sydney pp 1-198
Bale WM (1913) Further notes on Australian hydroids. II Proc R Soc Vict 26:114–147
Bedot M (1921) Notes systématiques sur les plumularides. 1re partie. Rev Suisse Zool 28:311–356
Bedot M (1922) Les caractères sexuels secondaires des Plumularides. Rev Suisse Zool 29:147–166
Billard A (1910) Revision d’une partie de la collection des hydroïdes du British Museum Annls. Sci nat Zool 11:1–67
Billard A (1913) Les hydroïdes de l’expédition du Siboga. I. Plumulariidae. Siboga-Expeditie, Monografie 7a:1–115
Boero F, Bouillon J (1993) Fraseroscyphus sinuosus n.gen. (Cnidaria, Hydrozoa, Leptomedusae, Sertulariidae), an epiphytic hydroid with a specialised clinging organ. Can J Zool 71(5):1061–1064
Borradaile LA (1905) Hydroids. The fauna and geography of the Maldive and Laccadive Archipelagos. University Press, Cambridge 2:836–845
Bouillon J, Massin C, Kresevic R (1995) Hydromedusae de l’’Institut Royal des Sciences Naturelles de Belgique. Doc Trav I R Sc N B 78:3–106
Bouillon J, Gravili C, Pagès F, Gili JM, Boero F (2006) An introduction to the Hydrozoa. Mém Mus Nat Hist Nat 194:1–591
Briggs EA (1918) Descriptions of two new hydroids, and a revision of the hydroid-fauna of Lord Howe Island. Rec Aust Mus 12:27–47
Busk G (1852) An account of the Polyzoa and sertularian zoophytes collected in the voyage of the “Rattlesnake” on the coast of Australia and the Louisiade Archipelago. In: J. Macgillivray, Narrative of the voyage of H.M.S. Rattlesnake commanded by the late Captain O. Stanley during the years 1846-1850. 1 Appendix IV. Kessinger, Whitefish, pp 343-402
Di Camillo CG, Bavestrello G, Valisano L, Puce S (2008) Spatial and temporal distribution in a tropical hydroid assemblage. J Mar Biol Ass UK 88:1589–1599
Di Camillo CG, Puce S, Bavestrello G (2009) Macrorhynchia species (Cnidaria: Hydrozoa) from the Bunaken Marine Park (North Sulawesi, Indonesia) with a description of two new species. Ital J Zool 76:208–228
García Corrales P, Aguirre Inchaurbe A, González Mora D (1978) Contribución al conocimiento de los hidrozoos de las costas españolas. Parte I: Halecidos, Campanularidos y Plumularidos. Boln Inst Esp Oceanogr 4:5–73
Gravier-Bonnet N, Bourmaud CAF (2006a) Hydroids (Cnidaria, Hydrozoa) of coral reefs: preliminary results on community structure, species distribution and reproductive biology in the Îles Glorieuses (Southwest Indian Ocean). Proc 10th ICRS, Okinawa, Japan, pp 188-196
Gravier-Bonnet N, Bourmaud CAF (2006b) Hydroids (Cnidaria, Hydrozoa) of coral reefs: preliminary results on community structure, species distribution and reproductive biology in Juan de Nova island (Southwest Indian Ocean). West Indian Ocean J Mar Sci 5:123–132
Irving RA (1995) Near-shore bathymetry and reef biotopes of Henderson Island, Pitcairn Group. Biol J Linn Soc 56:309–324
Jäderholm E (1920) On some exotic hydroids in the Swedish Zoological State Museum. Ark Zool 13:1–11
Jarvis FE (1922) The hydroids from the Chagos, Seychelles and other islands and from the coasts of British East Africa and Zanzibar. In: Reports of the Percy Sladen Trust Expedition to the Indian Ocean in 1905, under the leadership of Mr. J. Stanley Gardiner, M.A. Trans Linn Soc London. Zoology 18:331–360
Kirchenpauer GH (1872) Ueber die Hydroidenfamilie Plumularidae, einzelne Gruppen derselben und ihre Fruchtbehälter. I. Aglaophenia, Lx. Abh Geb Naturw Hamburg 6:1–58
Kirchenpauer GH (1876) Ueber die Hydroidenfamilie Plumularidae, einzelne Gruppen derselben und ihre Fruchtbehälter II. Plumularia und Nemertesia. Abh Geb Naturw Hamburg 6:1–59
Kirkpatrick R (1890) Reports on the zoological collections made in Torres Straits by professor A.C. Haddon, 1888-1889. Hydroida and Polyzoa. Sci Proc R Dubl Soc 6:603–626
Leloup E (1930) Sur un hydropolype nouveau, Thecocarpus leopoldi nov. sp., des Indes orientales néerlandaises. Bull Mus r Hist nat Belg 6:1–3
Millard NAH, Bouillon J (1973) Hydroids from the Seychelles (Coelenterata). Annls Mus r Afr Centrale, Série in 8. Sci Zool 206:1–106
Millard NAH, Bouillon J (1974) A collection of hydroids from Moçambique, East Africa. Ann S Afr Mus 65:1–40
Millard NAH (1975) Monograph on the Hydroida of southern Africa. Ann S Afr Mus 68:1–513
Millard NAH (1968) South African hydroids from Dr. Th. Mortensen’s Java-South Africa expedition, 1929-1930. Vidensk Meddr dansk Naturh Foren 131:251–288
Pennycuik PR (1959) Faunistic records from Queensland. Part V. Marine and brackish water hydroids. Pap Dep Zool Univ Qd 1:141–210
Ramil FJ, Vervoort W (1992) Some considerations concerning the genus Cladocarpus (Cnidaria: Hydrozoa). In: Bouillon J, Boero F, Cicogna F, Gili JM, Hughes RG (eds) Aspects of hydrozoan biology. Sci Mar 56:171–176
Rees W, Thursfield S (1965) The hydroid collections of James Ritchie. P Roy Soc Edinb B 69:34–220
Ryland JS, Gibbons MJ (1991) Intertidal and shallow water hydroids from Fiji II. Plumulariidae and Aglaopheniidae. Mem Qd Mus 30:525–560
Schuchert P (2003) Hydroids (Cnidaria, Hydrozoa) of the Danish expedition to the Kei Islands. Steenstrupia 27:137–256
Stechow E (1919) Zur Kenntnis der Hydroidenfauna des Mittelmeeres, Amerikas und anderer Gebiete, nebst Angaben über einige Kirchenpauer’sche Typen von Plumulariden. Zool Jb Syst 42:1–172
Tang Z (1991) On a collection of Hydroida from the Nansha Islands, Hainan province, China. Contr St Mar Biol Nansha Isl 1:25–36
Vasseur P (1974) The overhangs, tunnels and dark reef galleries of Tuléar (Madagascar) and their sessile invertebrate communities. Proc 2nd Int Coral Reef Symp 2:143–159
Vervoort W, Vasseur W (1977) Hydroids from French Polynesia with notes on distribution and ecology. Zool Verh Leiden 159:3–98
Vervoort W, Watson JE (2003) The Marine Fauna of New Zealand: Leptothecata (Cnidaria: Hydrozoa) (Thecate Hydroids). Niwa Biodivers Mem NZ 119:1–538
Watson JE (2000) Hydroids (Hydrozoa: Leptothecatae) from the Beagle Gulf and Darwin Harbour, northern Australia. Beagle Rec Mus Art Galleries NT 16:1–82
Whitelegge T (1899) The Hydrozoa, Scyphozoa, Actinozoa, and Vermes of Funafuti. In: The atoll of Funafuti, Elliot Group; its zoology, botany, ethnology, and general structure based on collections made by Mr. Charles Hedley of the Australian Museum, Sydney, N.S.W. Mem Aust Mus 3:371–394
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Di Camillo, C.G., Puce, S. & Bavestrello, G. Lytocarpia and Cladocarpus (Cnidaria: Hydrozoa, Aglaopheniidae) from the Bunaken National Marine Park (North Sulawesi, Indonesia). Mar Biodiv 41, 517–536 (2011). https://doi.org/10.1007/s12526-011-0081-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12526-011-0081-4