
Abstract
In recent years, reports on bone breakage at archaeological sites have become more common in the taphonomic literature. The present work tests a recently published method, based on the use of machine learning algorithms for analysing the processes involved in bone breakage, to identify the agent that broke the bones of medium-sized animals at the Mousterian Navalmaíllo Rock Shelter (Pinilla del Valle, Madrid). This is the first time this method has been used in an archaeological setting. The results show that these bones were mostly broken by anthropic action, while some were slightly ravaged by carnivores, probably hyaenas. These findings agree very well with published interpretations of the site, and show the method used to be useful in taphonomic studies of archaeological materials with poorly preserved cortical surfaces.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Studies of the faunal remains at archaeological sites have been helpful in explaining the origin of bone accumulations and the role of hominins in their appearance. The “hunting-scavenging” debate has focused the attention of taphonomists on efforts to determine whether hominins or carnivores were the main agents responsible, and how secondary agents might modify the characteristics of these remains (Bunn 1982; Bunn and Ezzo 1993; Domínguez-Rodrigo 2002; Egeland et al. 2007; Pante et al. 2012; James and Thompson 2015; Domínguez-Rodrigo 2015; Harris et al. 2017; Parkinson 2018). The examination of bone surface modifications (BSM) has been the traditional manner in which these questions have been addressed. Percussion marks made during attempts to reach the bone marrow (Blumenschine and Selvaggio 1988; Blumenschine 1995; Domínguez-Rodrigo and Barba 2006; Galán et al. 2009; Blasco et al. 2014; Yravedra et al. 2018), cut marks on the surface of bones (Abe et al. 2002; Andrews and Cook 1985; Behrensmeyer et al. 1986; Bello et al. 2009; Bello and Soligo 2008; Binford 1981; Braun et al. 2016; Bromage and Boyde 1984; Bunn 1982, 1981; Bunn et al. 1986; Courtenay et al. 2017; de Juana et al. 2010; Dewbury and Russell 2007; Domínguez-Rodrigo 1997; Domínguez-Rodrigo et al. 2009, 2005; Maté-González et al. 2019, 2018, 2015; Olsen and Shipman 1988; Palomeque-González et al. 2017; Shipman and Rose 1983; Wallduck and Bello 2018; Yravedra et al. 2017), and tooth marks left by carnivores (Selvaggio 1994; Blumenschine 1995; Blumenschine et al. 1996; Domıínguez-Rodrigo and Piqueras 2003; Domínguez-Rodrigo and Barba 2006; Njau and Blumenschine 2006; Baquedano et al. 2012a; Andrés et al. 2013; Saladié et al. 2013; Arilla et al. 2014; Aramendi et al. 2017; Arriaza et al. 2017b; Yravedra et al. 2018) have all provided clues.
BSM, however, cannot always provide the answer. Sometimes it is difficult to identify de visu what actually made an alteration due to the equifinality involved in mark interpretation and its highly subjective identification (Domínguez-Rodrigo et al. 2017). For example, when a site has been subject to trampling, the marks left on bone remains may look very similar to cut marks (Behrensmeyer et al. 1986; Olsen and Shipman 1988; Blasco et al. 2008; Domínguez-Rodrigo et al. 2009; Pineda et al. 2014; Courtenay et al. 2018). In addition, percussion marks can be confused with tooth marks if the former have no associated microstriations (Galán et al. 2009; Yravedra et al. 2018). On other occasions, the state of preservation renders the task of identification impossible, and no conclusions can be drawn regarding the agent responsible for making bone accumulations (Egeland and Domínguez-Rodrigo 2008; e.g. Domínguez-Rodrigo and Martínez-Navarro 2012; Pineda et al. 2014, 2017, 2019; Yravedra et al. 2016; Pineda and Saladié 2018). However, the debate surrounding anthropic activity at archaeological sites has tended to overlook the evidence that bone breakage (which has been documented at sites aged at least 2.6 m.a. (Domínguez-Rodrigo et al. 2005)) can provide. Bone marrow is an important source of lipids and essential fat-soluble vitamins (A, D, E, K) (Saint-Germain 2005; Malet 2007; Costamagno and Rigaud 2014) for both human groups and carnivores (Binford 1978; Metcalfe and Jones 1988; Jones and Metcalfe 1988), and both break bones to some degree in order to obtain it.
BSM may not provide information by themselves on how a bone was broken, thus requiring the use of other methods to determine how this occurred, although the distribution of percussion or tooth marks can be related with the bone breakage. One way to do these studies has been to examine the notches that appear on fracture planes. Via their classification, it has been possible to distinguish between breakages made by anthropic percussion or the action of carnivores (Capaldo and Blumenschine 1994; de Juana and Domínguez-Rodrigo 2011; Galán et al. 2009; Moclán and Domínguez-Rodrigo 2018).
Another method is to analyse the fracture planes themselves, which can have different morphologies depending on the state of the bone (i.e. green or dry) (Villa and Mahieu 1991) and angles (Alcántara García et al. 2006). The main variations in green fracture planes are owed to the fact that anthropic breakage is percussive, i.e. it involves dynamic loading which distributes the energy of a strike differently around a bone’s structure. In contrast, the static loading associated with carnivore bites means the associated energy is distributed much more evenly across the bone surface (Johnson 1985).
With this in mind, Alcántara García et al. (2006) suggested an analytical method (for use with the remains of large and small prey animals) for identifying the agent responsible for the breaking of a bone via the angles of the fracture planes. However, the results obtained with this method were never very clear when used with archaeological remains (Pickering et al. 2005)—some authors highlighted methodological problems associated with the technique (Coil et al. 2017), while others raised concerns about the size of the samples available for analysis (Moclán et al. 2019).
Based on the method of Alcántara García et al. (2006), Moclán et al. (2019) recently reported the use of machine learning (ML)-based statistical techniques for analysing the fracture planes of broken bones of medium-sized animals (50–200 kg) and thus identifying the bone-breaking agent. In that work, it was shown experimentally that it is possible to distinguish between bone accumulations made by hyaenas, wolves and hominins with more than 95% confidence. The technique involved examining 12 variables related to bone breakage, including the morphology of the fracture plane, the angle of the fracture plane and the presence and type of notches on bone fragments. The results also suggested this method might be useful for examining material with poorly preserved cortical surfaces. Other authors too have shown that ML algorithms (Arriaza and Domínguez-Rodrigo 2016; Egeland et al. 2018; Domínguez-Rodrigo and Baquedano 2018; Domínguez-Rodrigo 2018; Courtenay et al. 2019), with their enormous statistical potential, can be of use in solving taphonomic problems.
However, in the above work (Moclán et al. 2019), no material from the archaeological record was ever examined. The Navalmaíllo Rock Shelter, a Mousterian site, is a good laboratory for testing this method on such remains. The BSM on the bone remains at the site are well enough preserved to have established Homo neanderthalensis as the main agent behind their accumulation. The clear majority of the remains worked by these hominins are of very large-, large- and medium-sized animals (Huguet et al. 2010; Moclán et al. 2017, 2018a). The marks left by carnivores are scant, and generally found on the remains of small and very small animals (Moclán et al. 2017; Arriaza et al. 2017a; Moclán et al. 2018a). The aim of the present work was to test the validity of the above ML method on the remains of medium-sized animals from the Navalmaíllo Rock Shelter, to re-determine whether the bone breakages seen there are the result of intense anthropic activity (Huguet et al. 2010).
The Navalmaíllo Rock Shelter: a Middle Palaeolithic site on the Iberian Plateau
The archaeological sites at Calvero de la Higuera in Pinilla del Valle (Madrid, Spain) are located in the upper valley of the Lozoya River in the Guadarrama Mountains (Sierra de Guadarrama), 55 km north of the city of Madrid. Five different sites have been found to date: the Cueva del Camino, the Buena Pinta Cave, the Des-Cubierta Cave, the Ocelado Rock Shelter and the Navalmaíllo Rock Shelter (Fig. 1) (Alférez et al. 1982; Arsuaga et al. 2009, 2010; Baquedano et al. 2010; Pérez-González et al. 2010; Arsuaga et al. 2011; Baquedano et al. 2012b; Arsuaga et al. 2012; Álvarez-Lao et al. 2013; Laplana et al. 2013; Márquez et al. 2013; Baquedano et al. 2014; Blain et al. 2014; Karampaglidis 2015; Laplana et al. 2015; Baquedano et al. 2016; Márquez et al. 2016a; Laplana et al. 2016; Márquez et al. 2017; Moclán et al. 2018a).

Upper left: orthophotographic view of the Pinilla del Valle archaeological sites (1, the Camino Cave; 2, the Navalmaíllo Rock Shelter; 3, the Buena Pinta Cave; 4, the Des-Cubierta Cave). Upper right: general view of the excavation area of Level F from the north part of the site (note the presence of the archaeological pit for context in further figures). Lower image: orthophotograph of the Navalmaíllo Rock Shelter where Level F and other parts of the site can be seen. Orthophotograph by Alfonso Dávila Lucio/MAR
The Navalmaíllo Rock Shelter (hereinafter NV) was discovered in 2002, and has been excavated without interruption ever since. The site is a rock shelter, carved out by the Valmaíllo stream (Pérez-González et al. 2010), with a minimum surface area of ~ 250 m2 (Análisis y Gestión del Subsuelo S.L. [AGS] 2006). The stratigraphic sequence (Fig. 2) from top to bottom consists of an Ap horizon (10YR 5/2) some 0.20–0.40 m thick and at least two colluvium stages of dolomitic clasts within a silt-sand matrix (7.5YR 6/3) up to 1 m thick. Below this lies large dolomite blocks (some more than 1 m in height) that have fallen from the rock shelter ceiling. The falling of the blocks produced the hydroplastic injection of clays (Level D) containing some lithic and faunal remains from Level F. Level F lies below Level D, forming a bed up to 0.85 m thick composed of clay/sand (10YR 4/3) and carbonate clasts with a long axis of up to 0.35 m. Burnt sediments from this level have been dated by thermoluminiscence to be between 71.685 ± 5.082 and 77.230 ± 6.016 ka, i.e. the final part of MIS5a or the first part of the MIS4. According to pollen analyses, Level F reflects an open ecosystem (Ruiz Zapata et al. 2015). Under Level F are at least 2 m of allochthonous fluvial facies of siliceous gravel and sands deposited by the Valmaíllo stream, which drains Variscan gneisses before flowing into the Lozoya River.

a Stratigraphic sequence of the site (for a complete view of the sequence and its legend, see Fig. 2 in Arriaza et al. 2017a). Note that the archaeological materials of Level D are found in a secondary position due to the effects of hydroplasticity; b orthophotograph of the stratigraphic sequence of the Navalmaíllo Rock Shelter in the archaeological pit (see Fig. 1). Orthophotograph by Alfonso Dávila Lucio/MAR
The remains of lithic industry make up some 60% of the archaeological record of Level F (Márquez et al. 2016b). Most of these remains (~ 78%) are made of locally collected quartz (Abrunhosa et al. 2014, 2019); the most common technological category recorded is that of simple flakes (Márquez et al. 2013). There is a clear trend towards the production of microlithics, an apparently intentional choice, at least for those tools made of quartz. The presence of anvils near cores, along with bipolar products, indicates bipolar knapping to have been the best way of handling this type of raw material, especially for small format products (Márquez et al. 2013, 2016b, 2017).
The faunal remains of Level F belong to animals of different size (Table 1) (Huguet et al. 2010; Arsuaga et al. 2011; Moclán et al. 2017, 2018a, 2018b; Arriaza et al. 2017a). Previous studies have highlighted the major presence of large animals such as Bos primigenius/Bison priscus in the assemblage, as well as Equus ferus and Stephanorhinus hemitoechus (Huguet et al. 2010). Medium-sized animals (such as cervids) are also well represented, but small-sized animals are clearly underrepresented (Moclán et al. 2017, 2018a). The remains of the very large-, large- and medium-sized mammals show clear evidence of anthropic activity, such as cut marks, percussion marks and evidence of burning to varying degrees of intensity. Taphonomic alterations produced by carnivores are not common in the faunal assemblage, and it would appear that Neanderthals were the main drivers of bone accumulation at the site (Huguet et al. 2010; Moclán et al. 2017, 2018a). The remains of small-sized animals (such as rabbits and tortoises) and carnivores are not the result of anthropic activity. The rabbits, for example, were almost certainly brought into the shelter by lynxes when the shelter was not occupied by human groups (Arriaza et al. 2017a).
Materials
The materials examined in this work were bones from the archaeological record of NV Levels D and F, plus the non-archaeological materials presented by Moclán et al. (2019). The experimental (non-archaeological) materials were divided into three sets: set 1—40 anthropically broken long bones (10 humeri, 10 radii-ulnae, 10 femurs, 10 tibiae) from Cervus elaphus (a medium-sized species), set 2—medium-size bone remains from a hyena (Crocuta crocuta) den near Lake Eyasi in Tanzania, and set 3—the bones of medium-sized animals (Cervus elaphus and Sus scrofa) broken by wolves (Canis lupus) living semi-free in the Hosquillo Natural Park (Serranía de Cuenca, Spain). All three sets have been previously described in detail (Prendergast and Domínguez-Rodrigo 2008; Moclán and Domínguez-Rodrigo 2018; i.e. Moclán et al. 2019). All the bone fragments represented in the latter two sets were identifiable as belonging to long bones; no metapodials were included since previous work has shown since, given the very thick cortical cross-section, they provide little evidence about the bone-breaking agent (Capaldo and Blumenschine 1994). The anthropically generated sample (n = 332) provided 881 fracture planes for analysis (oblique = 549; longitudinal = 297; transverse = 35), the hyaena-generated samples (n = 66) provided 202 fracture planes (oblique = 87; longitudinal = 91; transverse = 24), and the wolf-generated sample (n = 61) provided 237 (oblique = 273; longitudinal = 287; transverse = 50).
The NV archaeological remains examined were 12,966 bone fragments (unearthed between 2002 and 2018), with 11,919 from Level F and 1047 from Level D. The materials from Level D were originally from Level F, but were moved by the hydroplastic processes that affected the ceiling of Level F, where they eventually settled. Metapodials were excluded from the analysis. One thousand one hundred four specimens of medium-sized animals were detected (long bones = 63.77%) with a total of 377 fracture planes.
Methods
Zooarchaeological and taphonomic methods
For taxonomic identifications and the assignment of a size (via body weight), the reference collections of the Institut Català de Paleoecologia Humana i Evolució Social (IPHES) and the Museo Arqueológico Regional (MAR), and manuals of comparative anatomy were used (Pales and Lambert 1971; Schmid 1972; Barone 1976; France 2009).
The sizes of the animals represented by the bone remains were defined via their weight as very large (> 800 kg), large (200–800 kg), medium (50–200 kg), small (10–50 kg) and very small (< 10 kg). The assignment of a size was independent of any taxonomic identification.
BSM were identified following different methods: cut marks were distinguished using the criteria of Domínguez-Rodrigo et al. (2009), tooth marks were identified following the method of Blumenschine (1988, 1995) and percussion marks were distinguished following the methods of Blumenschine and Selvaggio (1988) and Blumenschine (1995).
Breakage patterns were identified following the method of Villa and Mahieu (1991), differentiating between those in dry and fresh bones. The fracture planes were divided into three types—transversal, longitudinal and oblique—according to the trajectory followed with respect to the long axis of the bone (Alcántara García et al. 2006). Gifford-Gonzalez (1989) and Haynes (1983) defined transverse planes as fractures occurring at right angles to the long axis of the bone, while longitudinal planes are fractures parallel to the long axis of the bone. Pickering et al. (2005) described oblique fracture planes on either straight or curved in a helical pattern with a subparallel angle in relation to the long axis of the bone.
Only remains over 4 cm in length were examined, following the indications of Alcántara García et al. (2006)—a requirement for determining some of the variables measured (see below). The angles of the fracture planes were measured using a goniometer at the point of greatest inflexion.
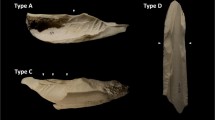
Notches were defined as “semi-circular to arcuate indentations on the fracture edge of a long bone that are produced by dynamic or static loading on cortical surfaces […] leaving a negative flake scar onto the medullary surface” as reported by Capaldo and Blumenschine (1994). Here we identified notches according to the typological classification proposed by Pickering and Egeland 2006; modified from Capaldo and Blumenschine 1994):
• Complete or type A notches: those with two inflection points on the cortical surfaces and a non-overlapping negative flake scar.
• Incomplete or type B notches: those missing one of the inflection points.
• Double opposing or type C notches: those with negative flake scars that overlap an adjacent notch.
• Double opposing complete or type D notches: the two notches that appear on opposite sides of a fragment and that result from two opposing loading points.
• Micronotches or type E notches: always < 1 cm in length.
Following the method of Moclán et al. (2019), the fracture planes were examined using ML algorithms. This required collecting data for 12 variables that allowed the planes to be individualised with no loss of a fragment’s general information, such as follows:
- 1.
The presence or absence of an epiphyseal section in the bone fragment.
- 2.
Length (mm) of the bone fragment.
- 3.
Length category (the fragments were included in categories related to their maximum length; see Table 2).
- 4.
The number of total fracture planes measurable in the fragment, including green transversal planes.
- 5.
Type of fracture plane, i.e. longitudinal or oblique (a variable used only when comparing all the samples at the same time).
- 6.
Angle of the fracture plane.
- 7.
Type of angle: if the angle is right or close (i.e. 85–95°) or not (obtuse or acute).
- 8.
Fracture plane longer than 4 cm or not.
- 9.
Presence or absence of notches in the fragment.
- 10.
Presence or absence of type A notches.
- 11.
Presence or absence of type C notches.
- 12.
Presence or absence of type D notches.
Note that some variables refer to the structure of the fragment studied (1, 2, 3, 4, 9, 10, 11, 12) and others to the fracture plane (5, 6, 7, 8), allowing for multivariate analysis (Domínguez-Rodrigo and Yravedra 2009; Domínguez-Rodrigo and Pickering 2010) rather than the univariate analysis covered by the original method of Alcántara García et al. (2006).
Statistical methods
The present work follows the methodology of Moclán et al. (2019), and thus involves classic univariate and bivariate analyses followed by the use of ML algorithms. All analyses were performed using R v.3.2.3 software (R Core Team 2015).
Firstly, the ratio of longitudinal to oblique fracture planes was established. According to Moclán et al. (2019), this ratio provides an initial means of testing whether a sample of fragment was subject to anthropic breakage (ratio < 1) or breakage via the action of carnivores (ratio > 1). This was followed by comparing of the angles of the fracture planes using the non-parametric Wilcoxon signed-rank test due to the Shapiro-Wilk test confirmed the normal distribution of the data.
Following the indications of Alcántara García et al. (2006), and making use of the “plotrix” library in R (Lemon 2006), the mean and 95% confidence intervals of the fracture plane angles were compared to examine the variation between the different experimental non-archaeological sets of bones (sets 1–3) and the NV sample.
Following the method of Moclán and Domínguez-Rodrigo (2018), and using the “cabootcrs” library in R (Ringrose 2013), the notches on alternative sample materials made by known agents—lions (Arriaza et al. 2016), hyaenas (Domínguez-Rodrigo et al. 2007), anthropic percussion action on small-, medium- and large-sized animal bones (Domínguez-Rodrigo et al. 2007; Blasco et al. 2014; Moclán and Domínguez-Rodrigo 2018), and anthropic percussion via battering (Blasco et al. 2014)—were compared via correspondence analysis of notch types A, B and C, thus providing a further comparator for the present archaeological materials. To facilitate understanding of the correspondence diagram, the examples of anthropic action on all large-sized animals in the alternative sample (Domínguez-Rodrigo et al. 2007; Blasco et al. 2014) were taken together.
Finally, ML statistical analysis was performed using the “caret” library in R (Kuhn 2017). The powerful statistical methods involved, which were first used in a taphonomic context by Arriaza and Domínguez-Rodrigo (2016), allow the classification of data, and predict into which categories new data will fall (Kuhn and Johnson 2013). A standard procedure in these ML analyses is the use of bootstrapping (Efron 1979) to render the results more robust. Following the method of Moclán et al. (2019), raw data for the experimental materials were bootstrapped 1000 times to generate a predictive model for comparison with the NV archaeological sample.
The functioning of the different ML algorithms differs mathematically, but the results are interpreted in the same way. In previous taphonomic studies (Arriaza and Domínguez-Rodrigo 2016; Domínguez-Rodrigo and Baquedano 2018; Domínguez-Rodrigo 2018; Moclán et al. 2019; Courtenay et al. 2019), kappa agreement indices were calculated. The kappa index, which accounts for the possibility of a correct prediction occurring by chance alone, takes a value of − 1 to 1, with values of 0.80 to 1 reflecting results in “very good agreement” (Lantz 2013). However, other variables also need to be taken into account, such as accuracy, sensitivity, specificity and balanced accuracy. Accuracy refers to the percentage success of the classification generated by an algorithm (represented on an ascending 0–1 scale). Sensitivity and specificity provide a view of how good the methods of delivering the kappa and accuracy values are, sensitivity describes the proportion of correctly classified positive results, and specificity describes the proportion of correctly classified negative results. The balanced accuracy corrects the final result taking into account both true positives and true negatives.
To generate the predictive model, the material in sets 1–3 was divided into “training” and “testing” groups (70 and 30%, respectively) (Moclán et al. 2019). This allows one to check whether the algorithms used make correct classifications (via the generation of kappa, accuracy, sensitivity, specificity and balanced accuracy values). For further details on the learning process, see Tables 5–9 of Moclán et al. (2019).
Since the present work involves archaeological material, a second step was required to examine that material with the predictive model generated by the algorithms. In this step, the algorithms process the data for each of the archaeological materials, and determine the probability that they were broken by one agent or another. This provides a final view of whether, as a whole, these materials were broken by wolves, hyaenas or anthropic percussion (the agents used in the training stage).
As in Moclán et al. (2019), the algorithms used in the present work were the neural network (NN), support vector machines (SVM), k-nearest neighbour (KNN), random forest (RF), mixture discriminant analysis (MDA) and naive Bayes (NB) algorithms. The partial least squares (PLS) algorithm and “decision trees using the C5.0 algorithm” (DTC5.0) were also used since these have been reported useful in taphonomic contexts (Arriaza and Domínguez-Rodrigo 2016; Domínguez-Rodrigo and Baquedano 2018; Domínguez-Rodrigo 2018; Moclán et al. 2019). Accuracy, kappa, sensitivity, specificity and balanced accuracy are reported for these algorithms since they were not included in Moclán et al. (2019).
Results
Univariate and bivariate analyses
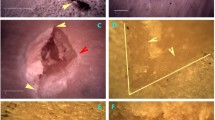
A total of 1104 NV remains were eligible for analysis out of the original 12,966 available. The sample represented two types of medium-sized herbivore—Cervus elaphus and Dama dama. Some samples were identified at the family level of Cervidae when the species could not be confirmed. These remains belonged to all anatomical areas, although remains of the long bones were the most common (n = 704), followed by those of cranial remains (n = 198) and flat bones (n = 122). BSM of different types were detected, including cut marks (n = 72; 7.17% of identified specimens [%NISP]), percussion marks (n = 32; 3.19% NISP) (Fig. 3), tooth marks (n = 13; 1.29% NISP) and thermal alterations (n = 136; 13.55% NISP). Notches were also seen at the fracture planes of the long bones (n = 30; 2.99% NISP).

Left: NV specimens of medium-sized animals with green fracture planes. Right: an example of a percussion mark on a shaft fragment of a medium-sized animal (Mag: ×35). Photos: Abel Moclán
Three hundred and thirteen remains of these medium-sized animals showed evidence of being broken when fresh; 168 fragments were over 4 cm long with a total 377 green fracture planes, of which 327 were in long bones of the upper and intermediate anatomical sections (it should be remembered that no metapodials were included in the sample). Twenty-two transverse fracture planes were seen, along with 136 longitudinal planes and 169 oblique planes (see Table 3); the longitudinal/oblique ratio was 0.80.
Comparison of the mean values for the angles for all sample sets (Wilcoxon signed-rank test) indicated the NV set to differ significantly (p < 0.05) from sets 1–3 (whether taking all specimens of each set together or analysing by type of fracture).
Following the method of Alcántara García et al. (2006), large differences were revealed among the NV materials and those of sets 1–3 in terms of the types of fracture planes present (Table 3; Fig. 4).

Comparison of mean and 95% confidence intervals for fracture angles in the NV and sets 1–3 material
As indicated by other authors (Alcántara García et al. 2006; Coil et al. 2017; Moclán et al. 2019), the transverse fracture planes showed too much variation in their angles to allow any conclusions to be drawn regarding the bone-breaking agent. The longitudinal and oblique planes, however, showed much less variation; < 90° longitudinal fracture planes were seen in both anthropically and carnivore-generated samples, while those of > 90° were seen only in the latter. In contrast, > 90° oblique fracture planes were seen in all samples generated by wolves, while those of < 90° were seen only in the specimens generated by percussion.
Thirty of the NV remains had notches; 15 were type A, 8 were type B, 1 was type C, 1 was type D and 2 were type E. Correspondence analysis showed the medium-sized NV remains to be clearly associated with an anthropic origin; certainly, the NV sample contained many simple notches (Fig. 5).

Bootstrapped correspondence analysis distinguishing the alternative experimental samples produced by carnivores, battering and percussion, and the NV sample of medium-sized animals. Ellipses cover the 95% confidence intervals. Left, distribution of the different types of notches. Right, distribution of the different samples
Machine learning analysis
The two ML new algorithms proposed (PLS and DTC5.0) returned results very similar to those reported by Moclán et al. (2019). PLS returned 81–89% correct classifications for the experimental pieces, while DTC5.0 returned 91–99% correct classifications (Table 4). This result shows DTC5.0, along with NN, RF and SVM, to be among the best algorithms available for determining the identity of bone-breakers. Like KNN, MDA and NB (Moclán et al. 2019), PLS is not as reliable.
Analysing the longitudinal and oblique fracture planes together revealed anthropic action to be the most likely agent that broke the NV bones. Depending on the algorithm used, the probabilities returned varied between 80.33% (DTC5.0) and 100% (PLS). The probability that hyaenas were the bone-breakers was just 18.36% according to the DTC5.0 algorithm, and 0% according to PLS. Wolves were the least probable bone-breakers; the NB algorithm returned a value of 4.59%, while PLS returned a value of 0% (Table 5).
When the same analysis was performed using only the longitudinal fracture planes, the same conclusion was reached. The probability that the NV material was broken anthropically was determined as 55.43% by the KNN algorithm and 98.91% by the NB algorithm when contemplating fracture planes of > 90°, and as 86.27% by NNET and 100% by PLS when contemplating fracture planes of < 90°.
The values returned for hyaenas ranged from 26.09 to 36.96% according to the NNET, SVM, KNN, RF and DTC5.0 algorithms, while the highest probability that wolves were responsible was returned by the KNN algorithm (7.61%) (Table 6).
A similar pattern was seen when the oblique fracture planes were examined alone (Table 7); once again, the most likely bone-breaker was determined to be hominins (60.80–100% depending on the algorithm). When considering fracture planes of < 90°, the maximum probability that hyaenas were responsible was 33.60%. When considering fracture planes of > 90°, the highest values suggesting hominins were responsible were recorded, ranging between 86.67% for the MDA algorithm, and 100% for NB and PLS.
Discussion
The statistical analysis of the green fracture planes of the NV material returned different results depending on the complexity of the test employed. The ratio of the types of fracture plane provided a clue that the bones were broken by hominins, although this kind of result needs to be understood with caution since large sets of material can have different proportions of different bones (i.e. humerus, radius-ulna, femur and tibia), which could affect over/underrepresented the results obtained (Moclán et al. 2019). Analysing the mean values of the fracture plane angles provided no clear result either; the Wilcoxon signed-rank test showed differences between the different sets of samples, while the 95% confidence test (Alcántara García et al. 2006) indicated one type of fracture plane had been generated by carnivores, and the other by anthropic action.
The results obtained in the analysis of the notches are also a little problematic. The correspondence analysis revealed a clear relationship between the NV notches and all the anthropic-origin samples. However, very wide variation was detected, and indeed, more overlap was seen with other small- and large-sized remains than other medium-sized remains.
The NV site has been described as a camp used by Homo neanderthalensis with evidence of intense anthropic action, including the fracturing of bones, on carcasses weighing over 50 kg (Huguet et al. 2010; Moclán et al. 2017, 2018a, 2018b). Here the anthropic activity is clearly confirmed by the use of ML algorithms. The examination of the longitudinal and fracture planes, and their combination, by these algorithms indicated anthropic action to be that main cause of bone breakage at the site.
That said, the different algorithms returned different probability values regarding this anthropic breakage. PLS, for example, returned a probability of 100% when oblique or longitudinal fracture planes of > 90° were contemplated, and when the entire sample was studied as a whole. These results should be understood with some caution, however, since the presence and the distribution of tooth marks on the NV materials and the absence of epiphyseal fragments indicate some degree of ravaging—although rather small (Huguet et al. 2010; Moclán et al. 2018a, 2018b, 2017). It should also be remembered that the accuracy of PLS was well below 100% when the experimental materials in sets 1–3 were examined; errors might also occur, therefore, when archaeological material is examined. In fact, the NNET, RF, SVM (Moclán et al. 2019) and DTC5.0 (current study) algorithms were shown to be the safest to use, and indeed these identified anthropic activity to be behind the breakages too (the RF algorithm returned a probability of 97.78% when contemplating oblique fractures of > 90°) (Fig. 6).

Differences in the classification of the bone-breaker of the NV material according to the NNET, SVM, RF and DTC5.0 algorithms (contemplating all fracture planes together, and longitudinal and oblique fracture planes separately)
It should not be overlooked that all these algorithms also returned a high probability that hyaenas—rather than wolves—were responsible for some modifications to the remains. Both taxa have been identified among the remains at the rock shelter, and previous work has shown that hyaenas probably ravaged some bones (Moclán et al. 2017, 2018a).
The present work shows the usefulness of ML algorithms for analysing bone breakages in archaeological materials. The study of BSM has been a priority of neotaphonomic studies, as the literature reveals, for example, with respect to cut marks (Shipman and Rose 1983; Bromage and Boyde 1984; Andrews and Cook 1985; Behrensmeyer et al. 1986; Olsen and Shipman 1988; Domínguez-Rodrigo 1997; Greenfield 1999, 2006; e.g. Abe et al. 2002; Domínguez-Rodrigo et al. 2005, 2009; Dewbury and Russell 2007; Bello and Soligo 2008; Bello et al. 2009; de Juana et al. 2010; Maté-González et al. 2015, 2018, 2019; Braun et al. 2016; Yravedra et al. 2017; Palomeque-González et al. 2017; Courtenay et al. 2017; Wallduck and Bello 2018). However, the present and earlier studies show that bone breakage is an important variable to bear in mind. It can used to help reveal the origin of faunal assemblages. Future work should focus on expanding the reference base for comparisons, and include studies on large- and small-sized bones, and different taxa.
Conclusions
This work, based on the methodology of Moclán et al. (2019), shows that ML algorithms can be used to identify bone-breakers, even those of archaeological material, without the need to examine BSM. The present results show that hominins were the bone-breakers at the Navalmaíllo Rock Shelter, but also indicate the remains suffered slight intervention by carnivores, probably hyaenas.
References
Abe Y, Marean CW, Nilssen PJ et al (2002) The analysis of cutmarks on archaeofauna: a review and critique of quantification procedures, and a new image-analysis GIS approach. Am Antiq 67:643–663. https://doi.org/10.2307/1593796
Abrunhosa A, Márquez B, Baquedano E et al (2014) Raw material study of the Mousterian lithic assemblage of Navalmaíllo rockshelter (Pinilla del Valle, Spain): preliminary results. Estudos do Quaternário Revista da Associação Portuguesa para o Estudo do Quaternário 11:19–25
Abrunhosa A, Pereira T, Márquez B et al (2019) Understanding Neanderthal technological adaptation at Navalmaíllo Rock Shelter (Spain) by measuring lithic raw materials performance variability. Archaeol Anthropol Sci 11:5949–5962. https://doi.org/10.1007/s12520-019-00826-3
Alcántara García V, Barba Egido R, Barral del Pino JM et al (2006) Determinación de procesos de fractura sobre huesos frescos: un sistema de análisis de los ángulos de los planos de fracturación como discriminador de agentes bióticos. Trab Prehist 63:37–45
Alférez F, Brea P, Buitrago AM et al (1982) Descubrimiento del primer yacimiento cuaternario (Riss-Würm) de vertebrados con restos humanos en la provincia de Madrid (Pinilla del Valle). Coloquios de Paleontología 37:15–32. https://doi.org/10.5209/rev_COPA.1982.v37.35543
Álvarez-Lao D, Arsuaga JL, Baquedano E, Pérez-González A (2013) Last Interglacial (MIS 5) ungulate assemblage from the Central Iberian Peninsula: the Camino Cave (Pinilla del Valle, Madrid, Spain). Paleogeogr Paleoclimatol Paleoecol 374:327–337
Análisis y Gestión del Subsuelo S.L. [AGS] (2006) Prospección geofísica para la caracterización del subsuelo en las excavaciones de Pinilla del Valle (Madrid). c/ Luxemburgo no 4, portal 1, oficina 3. 28224-Pozuelo de Alarcón, Madrid, Spain
Andrés M, Gidna A, Yravedra J, Domínguez-Rodrigo M (2013) A study of dimensional differences of tooth marks (pits and scores) on bones modified by small and large carnivores. Archaeol Anthropol Sci 4:209–219
Andrews P, Cook J (1985) Natural modifications to bones in a temperate setting. Man 20(4):675–691. https://doi.org/10.2307/2802756
Aramendi J, Maté-González MA, Yravedra J et al (2017) Discerning carnivore agency through the three-dimensional study of tooth pits: revisiting crocodile feeding behaviour at FLK- Zinj and FLK NN3 (Olduvai Gorge, Tanzania). Palaeogeogr Palaeoclimatol Palaeoecol 488:93–102. https://doi.org/10.1016/j.palaeo.2017.05.021
Arilla M, Rosell J, Blasco R et al (2014) The “bear” essentials: actualistic research on Ursus arctos arctos in the Spanish Pyrenees and its implications for paleontology and archaeology. PLoS One 9:e102457. https://doi.org/10.1371/journal.pone.0102457
Arriaza MC, Domínguez-Rodrigo M (2016) When felids and hominins ruled at Olduvai Gorge: a machine learning analysis of the skeletal profiles of the non-anthropogenic Bed I sites. Quat Sci Rev 139:43–52. https://doi.org/10.1016/j.quascirev.2016.03.005
Arriaza MC, Domínguez-Rodrigo M, Yravedra J, Baquedano E (2016) Lions as bone accumulators? Paleontological and ecological implications of a modern bone assemblage from Olduvai Gorge. PLoS One 11. https://doi.org/10.1371/journal.pone.0153797
Arriaza MC, Huguet R, Laplana C et al (2017a) Lagomorph predation represented in a middle Palaeolithic level of the Navalmaíllo Rock Shelter site (Pinilla del Valle, Spain), as inferred via a new use of classical taphonomic criteria. Quat Int 436:294–306. https://doi.org/10.1016/j.quaint.2015.03.040
Arriaza MC, Yravedra J, Domínguez-Rodrigo M et al (2017b) On applications of micro-photogrammetry and geometric morphometrics to studies of tooth mark morphology: the modern Olduvai Carnivore Site (Tanzania). Palaeogeogr Palaeoclimatol Palaeoecol 488:103–112. https://doi.org/10.1016/j.palaeo.2017.01.036
Arsuaga JL, Baquedano E, Pérez-González A (2009) Neanderthal and carnivore occupations in Pinilla del Valle sites (Community of Madrid, Spain). In: Oosterbeek L (ed) Proceedings of the XV World Congress of the International Union for Prehistoric and Protohistoric Sciences. Lisbonne, pp 111–119
Arsuaga JL, Baquedano E, Pérez-González A et al (2010) El yacimiento arqueopaleontológico del Pleistoceno Superior de la Cueva del Camino en el Calvero de la Higuera (Pinilla del Valle, Madrid). Zona Arqueológica 13:421–442
Arsuaga JL, Baquedano E, Pérez-González A (2011) Neanderthal and carnivore occupations in Pinilla del Valle sites (Community of Madrid, Spain). In: Oosterbeek L, Fidalgo C (eds) Proceedings of the XV World Congress of the International Union for Prehistoric and Protohistoric Sciences. Archaeopress, Inglaterra, pp 111–119
Arsuaga JL, Baquedano E, Pérez-González A et al (2012) Understanding the ancient habitats of the last-interglacial (late MIS 5) Neanderthals of central Iberia: Paleoenvironmental and taphonomic evidence from the Cueva del Camino (Spain) site. Quat Int 275:55–75. https://doi.org/10.1016/j.quaint.2012.04.019
Baquedano E, Arsuaga JL, Pérez-González A (2010) Homínidos y carnívoros: competencia en un mismo nicho ecológico pleistoceno: los yacimientos del Calvero de la Higuera en Pinilla del Valle. Actas de las Quintas Jornadas de Patrimonio Arqueológico en la Comunidad de Madrid 61–72
Baquedano E, Domínguez-Rodrigo M, Musiba C (2012a) An experimental study of large mammal bone modification by crocodiles and its bearing on the interpretation of crocodile predation at FLK Zinj and FLK NN3. J Archaeol Sci 39:1728–1737. https://doi.org/10.1016/j.jas.2012.01.010
Baquedano E, Márquez B, Pérez-González A et al (2012b) Neandertales en el valle del Lozoya: los yacimientos paleolíticos del Calvero de la Higuera (Pinilla del Valle, Madrid). Mainake:83–100
Baquedano E, Márquez B, Laplana C et al (2014) The archaeological sites at Pinilla del Valle: (Madrid, Spain). In: Sala R (ed) Pleistocene and Holocene hunter-gatherers in Iberia and the Gibraltar strait: the current archaeological record. Fundación Atapuerca, Burgos, pp 577–584
Baquedano E, Arsuaga JL, Pérez-González A et al (2016) The Des-Cubierta Cave (Pinilla del Valle, Comunidad de Madrid, Spain): a Neanderthal site with a likely funerary/realistic connection. Alcalá de Henares
Barone R (1976) Anatomie comparée des mamiferes domestiques: Ostéologie. Vigot Frères, Paris
Behrensmeyer AK, Kathleen DG, Yanagi GT (1986) Trampling as a cause of bone surface damage and pseudocutmarks. Nature 319:768–771. https://doi.org/10.1038/319768a0
Bello SM, Soligo C (2008) A new method for the quantitative analysis of cutmark micromorphology. J Archaeol Sci 35:1542–1552. https://doi.org/10.1016/j.jas.2007.10.018
Bello SM, Parfitt SA, Stringer C (2009) Quantitative micromorphological analyses of cut marks produced by ancient and modern handaxes. J Archaeol Sci 36:1869–1880. https://doi.org/10.1016/j.jas.2009.04.014
Binford LR (1978) Nunamiut Ethnoarchaeology. Academic Press, New York
Binford LR (1981) Bones: ancient men and modern myths. Academic Press, New York
Blain H-A, Laplana C, Sevilla P et al (2014) MIS 5/4 transition in a mountain environment: herpetofaunal assemblages from Cueva del Camino, central Spain. Boreas 43:107–120. https://doi.org/10.1111/bor.12024
Blasco R, Rosell J, Fernández Peris J et al (2008) A new element of trampling: an experimental application on the Level XII faunal record of Bolomor Cave (Valencia, Spain). J Archaeol Sci 35:1605–1618. https://doi.org/10.1016/j.jas.2007.11.007
Blasco R, Domínguez-Rodrigo M, Arilla M et al (2014) Breaking bones to obtain marrow: a comparative study between percussion by batting bone on an anvil and hammerstone percussion. Archaeometry 56:1085–1104. https://doi.org/10.1111/arcm.12084
Blumenschine RJ (1988) An experimental model of the timing of hominid and carnivore influence on archaeological bone assemblages. J Archaeol Sci 15:483–502. https://doi.org/10.1016/0305-4403(88)90078-7
Blumenschine RJ (1995) Percussion marks, tooth marks, and experimental determinations of the timing of hominid and carnivore access to long bones at FLK Zinjanthropus, Olduvai Gorge, Tanzania. J Hum Evol 29:21–51. https://doi.org/10.1006/jhev.1995.1046
Blumenschine RJ, Selvaggio MM (1988) Percussion marks on bone surfaces as a new diagnostic of hominid behaviour. Nature 333:763–765. https://doi.org/10.1038/333763a0
Blumenschine RJ, Marean CW, Capaldo SD (1996) Blind tests of inter-analyst correspondence and accuracy in the identification of cut marks, percussión marks, and carnivore toothmarks on bone surfaces. J Archaeol Sci 23:493–507
Braun DR, Pante M, Archer W (2016) Cut marks on bone surfaces: influences on variation in the form of traces of ancient behaviour. Interface Focus 6:20160006. https://doi.org/10.1098/rsfs.2016.0006
Bromage TG, Boyde A (1984) Microscopic criteria for the determination of directionality of cutmarks on bone. Am J Phys Anthropol 65:359–366. https://doi.org/10.1002/ajpa.1330650404
Bunn HT (1981) Archaeological evidence for meat-eating by Plio-Pleistocene hominids from Koobi Fora and Olduvai Gorge. Nature 291:574–577. https://doi.org/10.1038/291574a0
Bunn HT (1982) Meat eating and human evolution: studies on the diet and subsistence patterns of Plio-Pleiostecene hominids in East Africa. University of California, California
Bunn HT, Ezzo JA (1993) Hunting and scavenging by Plio-Pleistocene hominids: nutritional constraints, archaeological patterns, and behavioural implications. J Archaeol Sci 20:365–398. https://doi.org/10.1006/jasc.1993.1023
Bunn HT, Kroll EM, Ambrose SH et al (1986) Systematic butchery by Plio/Pleistocene hominids at Olduvai Gorge, Tanzania [and comments and reply]. Curr Anthropol 27:431–452. https://doi.org/10.1086/203467
Capaldo SD, Blumenschine RJ (1994) A quantitative diagnosis of notches made by hammerstone percussion and carnivore gnawing in bovid long bones. Am Antiq 59:724–748
Coil R, Tappen M, Yezzi-Woodley K (2017) New analytical methods for comparing bone fracture angles: a controlled study of hammerstone and hyena (Crocuta crocuta) long bone breakage. Archaeometry 59:900–917. https://doi.org/10.1111/arcm.12285
Costamagno S, Rigaud J-P (2014) L’exploitation de la graisse au Paléolithique. In: Costamagno S (ed) Histoire de l’alimentation humaine : entre choix et contraintes (édition électronique). CTHS, París, pp 134–152
Courtenay LA, Yravedra J, Mate-González MÁ et al (2017) 3D analysis of cut marks using a new geometric morphometric methodological approach. Archaeol Anthropol Sci 11:1–15. https://doi.org/10.1007/s12520-017-0554-x
Courtenay LA, Yravedra J, Huguet R et al (2018) New taphonomic advances in 3D digital microscopy: a morphological characterisation of trampling marks. Quat Int 517:55–66. https://doi.org/10.1016/j.quaint.2018.12.019
Courtenay LA, Yravedra J, Huguet R et al (2019) Combining machine learning algorithms and geometric morphometrics: a study of carnivore tooth marks. Palaeogeogr Palaeoclimatol Palaeoecol 522:28–39. https://doi.org/10.1016/j.palaeo.2019.03.007
de Juana S, Galán AB, Domínguez-Rodrigo M (2010) Taphonomic identification of cut marks made with lithic handaxes: an experimental study. J Archaeol Sci 37:1841–1850. https://doi.org/10.1016/j.jas.2010.02.002
Dewbury AG, Russell N (2007) Relative frequency of butchering cutmarks produced by obsidian and flint: an experimental approach. J Archaeol Sci 34:354–357. https://doi.org/10.1016/j.jas.2006.05.009
Domıínguez-Rodrigo M, Piqueras A (2003) The use of tooth pits to identify carnivore taxa in tooth-marked archaeofaunas and their relevance to reconstruct hominid carcass processing behaviours. J Archaeol Sci 30:1385–1391. https://doi.org/10.1016/S0305-4403(03)00027-X
Domínguez-Rodrigo M (1997) A reassessment of the study of cut mark patterns to infer hominid manipulation of fleshed carcasses at the FLK Zinj 22 site, Olduvai Gorge, Tanzania. Trab Prehist 54(2):29–42. https://doi.org/10.3989/tp.1997.v54.i2.364
Domínguez-Rodrigo M (2002) Hunting and scavenging by early humans: the state of the debate. J World Prehist 16:1–54. https://doi.org/10.1023/A:1014507129795
Domínguez-Rodrigo M (2015) Taphonomy in early African archaeological sites: questioning some bone surface modification models for inferring fossil hominin and carnivore feeding interactions. J Afr Earth Sci 108:42–46. https://doi.org/10.1016/j.jafrearsci.2015.04.011
Domínguez-Rodrigo M (2018) Successful classification of experimental bone surface modifications (BSM) through machine learning algorithms: a solution to the controversial use of BSM in paleoanthropology? Archaeol Anthropol Sci 11:2711–2725. https://doi.org/10.1007/s12520-018-0684-9
Domínguez-Rodrigo M, Baquedano E (2018) Distinguishing butchery cut marks from crocodile bite marks through machine learning methods. Sci Rep 8:5786. https://doi.org/10.1038/s41598-018-24071-1
Domínguez-Rodrigo M, Barba R (2006) New estimates of tooth mark and percussion mark frequencies at the FLK Zinj site: the carnivore-hominid-carnivore hypothesis falsified. J Hum Evol 50:170–194. https://doi.org/10.1016/j.jhevol.2005.09.005
Domínguez-Rodrigo M, Martínez-Navarro B (2012) Taphonomic analysis of the early Pleistocene (2.4 Ma) faunal assemblage from A.L. 894 (Hadar, Ethiopia). J Hum Evol 62:315–327
Domínguez-Rodrigo M, Pickering TR (2010) A multivariate approach for discriminating bone accumulations created by spotted hyenas and leopards: harnessing actualistic data from east and Southern Africa. J Taphonomy 8:155–179
Domínguez-Rodrigo M, Yravedra J (2009) Why are cut mark frequencies in archaeofaunal assemblages so variable? A multivariate analysis. J Archaeol Sci 36:884–894. https://doi.org/10.1016/j.jas.2008.11.007
Domínguez-Rodrigo M, Rayne Pickering T, Semaw S, Rogers MJ (2005) Cutmarked bones from Pliocene archaeological sites at Gona, Afar, Ethiopia: implications for the function of the world’s oldest stone tools. J Hum Evol 48:109–121. https://doi.org/10.1016/j.jhevol.2004.09.004
Domínguez-Rodrigo M, Egeland CP, Barba R (2007) The “physical attribute” taphonomic approach. In: Deconstructing Olduvai. Springer, Dordrecht, pp 23–32
Domínguez-Rodrigo M, de Juana S, Galán AB, Rodríguez M (2009) A new protocol to differentiate trampling marks from butchery cut marks. J Archaeol Sci 36:2643–2654. https://doi.org/10.1016/j.jas.2009.07.017
Domínguez-Rodrigo M, Saladié P, Cáceres I et al (2017) Use and abuse of cut mark analyses: the Rorschach effect. J Archaeol Sci 86:14–23. https://doi.org/10.1016/j.jas.2017.08.001
Efron B (1979) Bootstrap methods: another look at the jackknife. Ann Stat 7:1–26. https://doi.org/10.1214/aos/1176344552
Egeland CP, Domínguez-Rodrigo M (2008) Taphonomic perspectives on hominid site use and foraging strategies during Bed II times at Olduvai Gorge, Tanzania. J Hum Evol 55:1031–1052. https://doi.org/10.1016/j.jhevol.2008.05.021
Egeland CP, Domínguez-Rodrigo M, Barba R (2007) The hunting-versus-scavenging debate. In: Domínguez-Rodrigo M, Egido RB, Egeland CP (eds) Deconstructing Olduvai: a taphonomic study of the bed I sites. Springer, Dordrecht, pp 11–22
Egeland CP, Domínguez-Rodrigo M, Pickering TR et al (2018) Hominin skeletal part abundances and claims of deliberate disposal of corpses in the Middle Pleistocene. PNAS 201718678. https://doi.org/10.1073/pnas.1718678115
France DL (2009) Human and non-human bone identification:A color atlas. CRC Press, Boca Raton
Galán AB, Rodríguez M, de Juana S, Domínguez-Rodrigo M (2009) A new experimental study on percussion marks and notches and their bearing on the interpretation of hammerstone-broken faunal assemblages. J Archaeol Sci 36:776–784
Gifford-Gonzalez DP (1989) Ethnographic analogues for interpreting modified bones: some cases from East Africa. In: Bonnichsen R, Sorg MH (eds) Bone modifications. Center for the Study of the First Americans, Institute for Quaternary Studies. University of Maine, Orono, pp 179–246
Greenfield HJ (1999) The origins of metallurgy: distinguishing stone from metal cut-marks on bones from archaeological sites. J Archaeol Sci 26:797–808. https://doi.org/10.1006/jasc.1998.0348
Greenfield HJ (2006) Slicing cut marks on animal bones: diagnostics for identifying stone tool type and raw material. J Field Archaeol 31:147–163
Harris JA, Marean CW, Ogle K, Thompson J (2017) The trajectory of bone surface modification studies in paleoanthropology and a new Bayesian solution to the identification controversy. J Hum Evol 110:69–81. https://doi.org/10.1016/j.jhevol.2017.06.011
Haynes G (1983) Frequencies of spiral and green-bone fractures on ungulate limb bones in modern surface assemblages. Am Antiq 48:102–114. https://doi.org/10.2307/279822
Huguet R, Arsuaga JL, Pérez-González A et al (2010) Homínidos y hienas en el Calvero de la Higuera (Pinilla del Valle, Madrid) durante el Pleistoceno Superior. Resultados preliminares. Zona Arqueológica 13:443–458
James EC, Thompson JC (2015) On bad terms: problems and solutions within zooarchaeological bone surface modification studies. Environ Archaeol 20:89–103. https://doi.org/10.1179/1749631414Y.0000000023
Johnson E (1985) Current developments in bone technology. In: Schiffer MB (ed) Advances in archaeological method and theory. Academic Press, San Diego, pp 157–235
Jones KT, Metcalfe D (1988) Bare bones archaeology: bone marrow indices and efficiency. J Archaeol Sci 15:415–423. https://doi.org/10.1016/0305-4403(88)90039-8
Karampaglidis T (2015) La evolución geomorfológica de la cuenca de drenaje del río Lozoya (Comunidad de Madrid, España). Universidad Complutense de Madrid, Madrid
Kuhn M (2017) Caret: classification and regression training
Kuhn M, Johnson K (2013) Applied predictive modeling. Springer-Verlag, New York
Lantz B (2013) Machine learning with R. Packt Publishing, Birmingham
Laplana C, Blain H-A, Sevilla P et al (2013) Un assemblage de petits vertébrés hautement diversifié de la fin du MSI5 dans un environment montagnard au centre de l’Espagne (Cueva del Camino, Pinilla del Valle, Communauté Autonome de Madrid). Quaternaire 24(2):207–216
Laplana C, Sevilla P, Arsuaga JL et al (2015) How far into Europe did pikas (Lagomorpha: Ochotonidae) go during the Pleistocene? New evidence from Central Iberia. PLoS One 10:e0140513. https://doi.org/10.1371/journal.pone.0140513
Laplana C, Sevilla P, Blain H-A et al (2016) Cold-climate rodent indicators for the Late Pleistocene of Central Iberia: new data from the Buena Pinta Cave (Pinilla del Valle, Madrid Region, Spain). Comptes Rendus Palevol 15:696–706. https://doi.org/10.1016/j.crpv.2015.05.010
Lemon J (2006) Plotrix: a package in the red light district of R. R-News 6(4):8–12
Malet C (2007) L’alimentation lipidique en milieu froid. In: Actes des XVIIIe Rencontres Internationales d’Archéologie et d’Histoire d’Antibes. APDCA - CNRS, Antibes, pp 295–308
Márquez B, Mosquera M, Baquedano E et al (2013) Evidence of a neanderthal-made quartz-based technology at Navalmaíllo rockshelter (Pinilla del Valle, Madrid Region, Spain). J Anthropol Res 69:373–395
Márquez B, Baquedano E, Pérez-González A, et al (2016a) El Abrigo de Navalmaíllo (Pinilla del Valle, Madrid, España). Un campamento de Neandertales en el centro de la Península Ibérica. Alcalá de Henares
Márquez B, Baquedano E, Pérez-González A, Arsuaga JL (2016b) Microwear analysis of Mousterian quartz tools from the Navalmaíllo Rock Shelter (Pinilla del Valle, Madrid, Spain). Quat Int 424:84–97. https://doi.org/10.1016/j.quaint.2015.08.052
Márquez B, Baquedano E, Pérez-González A, Arsuaga JL (2017) Denticulados y muescas: ¿para qué sirven? Estudio funcional de una muestra musteriense en cuarzo del Abrigo de Navalmaíllo (Pinilla del Valle, Madrid, España). Trab Prehist 74(1):26–46
Maté-González MÁ, Yravedra J, González-Aguilera D et al (2015) Micro-photogrammetric characterization of cut marks on bones. J Archaeol Sci 62:128–142. https://doi.org/10.1016/j.jas.2015.08.006
Maté-González MÁ, Yravedra J, Martín-Perea DM et al (2018) Flint and quartzite: distinguishing raw material through bone cut marks. Archaeometry 60:437–452. https://doi.org/10.1111/arcm.12327
Maté-González MÁ, Courtenay LA, Aramendi J et al (2019) Application of geometric morphometrics to the analysis of cut mark morphology on different bones of differently sized animals. Does size really matter? Quat Int 517:33–44. https://doi.org/10.1016/j.quaint.2019.01.021
Metcalfe D, Jones KT (1988) A reconsideration of animal body-part utility indices. Am Antiq 53:486–504. https://doi.org/10.2307/281213
Moclán A, Domínguez-Rodrigo M (2018) An experimental study of the patterned nature of anthropogenic bone breakage and its impact on bone surface modification frequencies. J Archaeol Sci 96:1–13. https://doi.org/10.1016/j.jas.2018.05.007
Moclán A, Huguet R, Márquez B, et al (2017) El Abrigo de Navalmaíllo: nuevos datos preliminares para entender el poblamiento neandertal en la Meseta Ibérica. Aproximación desde la Zooarqueología y la Tafonomía. Burgos, España
Moclán A, Huguet R, Márquez B, et al (2018a) Pinilla del Valle sites: new preliminary data to understand Neanderthal-carnivore interaction in the Iberian Plateau. In: 60th Annual Meeting of the Hugo Obermaier-Society. Hugo Obermaier Society for Quaternary Research and Archaeology of the Stone Age, Tarragona, Spain p 106
Moclán A, Huguet R, Márquez B et al (2018b) Cut marks made with quartz tools: an experimental framework for understanding cut mark morphology, and its use at the Middle Palaeolithic site of the Navalmaíllo Rock Shelter (Pinilla del Valle, Madrid, Spain). Quat Int 493:1–18. https://doi.org/10.1016/j.quaint.2018.09.033
Moclán A, Domínguez-Rodrigo M, Yravedra J (2019) Classifying agency in bone breakage: an experimental analysis of fracture planes to differentiate between hominin and carnivore dynamic and static loading using machine learning (ML) algorithms. Archaeol Anthropol Sci 11(9):4663–4680. https://doi.org/10.1007/s12520-019-00815-6
Njau JK, Blumenschine RJ (2006) A diagnosis of crocodile feeding traces on larger mammal bone, with fossil examples from the Plio-Pleistocene Olduvai Basin, Tanzania. J Hum Evol 50:142–162. https://doi.org/10.1016/j.jhevol.2005.08.008
Olsen SL, Shipman P (1988) Surface modification on bone: trampling versus butchery. J Archaeol Sci 15:535–553. https://doi.org/10.1016/0305-4403(88)90081-7
Pales L, Lambert C (1971) Atlas ostéologique pour servir à l’identification des mammifères du Quaternaire. Editions du centre national de la recherche scientifique, París
Palomeque-González JF, Maté-González MÁ, Yravedra J et al (2017) Pandora: a new morphometric and statistical software for analysing and distinguishing cut marks on bones. J Archaeol Sci Rep 13:60–66. https://doi.org/10.1016/j.jasrep.2017.03.033
Pante MC, Blumenschine RJ, Capaldo SD, Scott RS (2012) Validation of bone surface modification models for inferring fossil hominin and carnivore feeding interactions, with reapplication to FLK 22, Olduvai Gorge, Tanzania. J Hum Evol 63:395–407. https://doi.org/10.1016/j.jhevol.2011.09.002
Parkinson JA (2018) Revisiting the hunting-versus-scavenging debate at FLK Zinj: a GIS spatial analysis of bone surface modifications produced by hominins and carnivores in the FLK 22 assemblage, Olduvai Gorge, Tanzania. Palaeogeogr Palaeoclimatol Palaeoecol 511:29–51. https://doi.org/10.1016/j.palaeo.2018.06.044
Pérez-González A, Karampaglidis T, Arsuaga JL et al (2010) Aproximación geomorfológica a los yacimientos del Pleistoceno Superior del Calvero de la Higuera en el Valle Alto del Lozoya (Sistema Central Español, Madrid). Zona Arqueológica 13:403–420
Pickering TR, Egeland CP (2006) Experimental patterns of hammerstone percussion damage on bones: implications for inferences of carcass processing by humans. J Archaeol Sci 33:459–469. https://doi.org/10.1016/j.jas.2005.09.001
Pickering TR, Domínguez-Rodrigo M, Egeland CP, Brain CK (2005) The contribution of limb bone fracture patterns to reconstructing early hominid behaviour at Swartkrans cave (South Africa): archaeological application of a new analytical method. Int J Osteoarchaeol 15:247–260. https://doi.org/10.1002/oa.780
Pineda A, Saladié P (2018) The Middle Pleistocene site of Torralba (Soria, Spain): a taphonomic view of the Marquis of Cerralbo and Howell faunal collections. Archaeol Anthropol Sci 11:2539–2556. https://doi.org/10.1007/s12520-018-0686-7
Pineda A, Saladié P, Vergès JM et al (2014) Trampling versus cut marks on chemically altered surfaces: an experimental approach and archaeological application at the Barranc de la Boella site (la Canonja, Tarragona, Spain). J Archaeol Sci 50:84–93. https://doi.org/10.1016/j.jas.2014.06.018
Pineda A, Saladié P, Huguet R et al (2017) Changing competition dynamics among predators at the late Early Pleistocene site Barranc de la Boella (Tarragona, Spain). Palaeogeogr Palaeoclimatol Palaeoecol 477:10–26. https://doi.org/10.1016/j.palaeo.2017.03.030
Pineda A, Cáceres I, Saladié P et al (2019) Tumbling effects on bone surface modifications (BSM): an experimental application on archaeological deposits from the Barranc de la Boella site (Tarragona, Spain). J Archaeol Sci 102:35–47. https://doi.org/10.1016/j.jas.2018.12.011
Prendergast ME, Domínguez-Rodrigo M (2008) Taphonomic analyses of a hyena den and a natural-death assemblage near Lake Eyasi (Tanzania). J Taphonomy 6:301–335
R Core Team (2015) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Ringrose TJ (2013) Cabootcrs: bootstrap confidence regions for correspondence analysis
Ruiz Zapata B, García G, José M et al (2015) Vegetación, clima y recursos naturales durante el Pleistoceno Superior en los alrededores del Abrigo de Navalmaíllo (Calvero de la Higuera, Pinilla del Valle, Madrid). Geogaceta 58:115–118
Saint-Germain C (2005) Animal fat in the cultural world of the Native Peoples of Northeastern America. In: Actes du Colloque de l’ICAZ, 2002. Oxbow books, Durham, pp 107–113
Saladié P, Huguet R, Díez C et al (2013) Taphonomic modifications produced by modern brown bears (Ursus arctos). Int J Osteoarchaeol 23:13–33. https://doi.org/10.1002/oa.1237
Schmid E (1972) Atlas of animal bones for prehistorians, archaeologists and quaternary geologists. Elsevier Publishing Company, Amsterdam
Selvaggio MM (1994) Carnivore tooth marks and stone tool butchery marks on scavenged bones: archaeological implications. J Hum Evol 27:215–228. https://doi.org/10.1006/jhev.1994.1043
Shipman P, Rose J (1983) Early hominid hunting, butchering and carcass processing behaviors: approaches to the fossil record. J Anthropol Archaeol 2(1):57–98. https://doi.org/10.1016/0278-4165(83)90008-9
Villa P, Mahieu E (1991) Breakage patterns of human long bones. J Hum Evol 21:27–48. https://doi.org/10.1016/0047-2484(91)90034-S
Wallduck R, Bello SM (2018) Cut mark micro-morphometrics associated with the stage of carcass decay: a pilot study using three-dimensional microscopy. J Archaeol Sci Rep 18:174–185. https://doi.org/10.1016/j.jasrep.2018.01.005
Yravedra J, Domínguez-Rodrigo M, Santonja M et al (2016) The larger mammal palimpsest from TK (Thiongo Korongo), Bed II, Olduvai Gorge, Tanzania. Quat Int 417:3–15. https://doi.org/10.1016/j.quaint.2015.04.013
Yravedra J, Maté-González MÁ, Palomeque-González JF, et al (2017) A new approach to raw material use in the exploitation of animal carcasses at BK (Upper Bed II, Olduvai Gorge, Tanzania): a micro-photogrammetric and geometric morphometric analysis of fossil cut marks. Boreas https://doi.org/10.1111/bor.12224
Yravedra J, Aramendi J, Maté-González MÁ et al (2018) Differentiating percussion pits and carnivore tooth pits using 3D reconstructions and geometric morphometrics. PLoS One 13:e0194324. https://doi.org/10.1371/journal.pone.0194324
Acknowledgements
The authors are grateful to the Pinilla del Valle research team for archaeopalaeontological discussions, especially A. Abrunhosa, A. Álvarez, D. Álvarez-Lao, M.A. Galindo-Pellicena, N. García-García and M.C. Ortega. The authors are very grateful to those who assisted during the experimental phase, particularly Alicia Caboblanco for her assistance, and Cárnicas DIBE S.L., which provided all the carcasses used in the preparation of the set 1 material. Special thanks are owed to M. Domínguez-Rodrigo who assisted with the statistical analysis. The authors also thank Adrian Burton for language and editing assistance.
Funding
AM is funded by a grant from the Junta de Castilla y León financed in turn by the European Social Funds through the Consejería de Educación (BDNS 376062). This research was conducted as part of competitive projects PGC 2018-094125-B-100 (MCIU/AEI/FEDER, UE), PGC 2018-093925-B-C32 (MICINN-FEDER), AGAUR (2017SGR1040), URV (2014, 2015 and 2016 PFR-URV-B2-17) and funded by the I+D activities program for research groups run by the Education Secretariat of the Madrid Regional Government. The study was also partly funded by the Museo Arqueológico Regional de la Comunidad de Madrid (MAR), Grupo Mahou and Canal de Isabel II-Gestión. This work is a contribution to the Valle de los Neandertales project (H2019/HUM-5840) funded by the Comunidad de Madrid and the Fondo Social Europeo.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Moclán, A., Huguet, R., Márquez, B. et al. Identifying the bone-breaker at the Navalmaíllo Rock Shelter (Pinilla del Valle, Madrid) using machine learning algorithms. Archaeol Anthropol Sci 12, 46 (2020). https://doi.org/10.1007/s12520-020-01017-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-020-01017-1