
Abstract
Findings of canid remains in graves at different sites in the northeast of the Iberian Peninsula are evidence of a widespread funerary practice that proliferated between the end of the 3rd and the 2nd millennium BC, in particular, in the Early-Middle Bronze Age contexts. The discovery of four foxes and a large number of dogs at the sites of Can Roqueta (Barcelona) and Minferri (Lleida) respectively, stand out among the many examples of these types of grave goods. In this work, we have made an approximation of the relationship between humans and canids through the study of their diet by analysis of stable isotopes of carbon and nitrogen in bone collagen. These analyses were complemented by archaeozoological, anthropological and archaeobotanical studies. The comparison of human and animal diets comprised a total of 37 canids, 19 domestic ungulates and 64 humans. The results indicate that the diet of the dogs was similar to that of humans, although δ15N values of dogs in Can Roqueta and Minferri are, on the average, 1.4‰ and 1.1‰, respectively, lower than those of humans. The offset between canids and the herbivorous ungulates of each site is not up to the established minimum for a trophic level, which implies an input of C3 plants and human intervention in the feeding of dogs and some of the foxes. Some particular cases in Can Roqueta suggest a specific food preparation, richer in cereals, for larger dogs (probably devoted to carrying loads), and possibly for at least one of the foxes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Dogs in Bronze Age burials in Catalonia
Animal offerings during burials in the Iberian Peninsula are a phenomenon that proliferated from the 5th millennium BC until the end of the Iron Age (Catagnano 2016). But burial dogs are especially important in Chalcolithic and Early-Middle Bronze Age contexts, between the end of the 3rd millennium BC and during the 2nd millennium BC (Albizuri 2011a; Albizuri et al. 2011b; Cámara Serrano et al. 2016; Conlin 2003; Daza 2011; Gómez 2003; Liesau et al. 2013; Nieto et al. 2014; Ruiz et al. 2014; Valera et al. 2010).
The first burials of canids in the Northeast of the Iberian Peninsula (mainly dogs) are recorded in funerary structures from the Middle Neolithic period onwards. Two cases were highlighted in the Northern region of the Northeast area, in Serra del Mas Bonet (Rosillo et al. 2012) and Cova de l’Avellaner (Bosch et al. 1989). Although, the most numerous examples are located within the Catalonian coastline, in the Pit Grave culture of the Middle Neolithic period (ca. 4200–3600 cal BC), as is the case of the necropolis of Bòbila Madurell (Martín Cólliga et al. 2017) or Can Tintorer, Mina 28 (Villalba 1999), the findings of one fox and one dog in Camí de Can Grau (Martí et al. 1997) and the examples of La Serreta and Ca l’Arnella sites (Albizuri et al., currently under study). At the moment, only one case has been found for the Pre-Pyrenees, in Ca l’Oliaire (Martín Cólliga et al. 2005). Yet, the rite was not generalised until the Early-Middle Bronze Age, when dogs played an important role in funerary rituals.
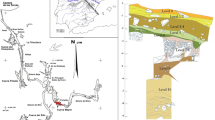
Although, many sites from this timeframe have been the subject of excavations, the most remarkable sites, both for their extension and the quantity of canid deposits, are the settlements of Can Roqueta (Sabadell, Barcelona), situated in the Pre-littoral Plain, and Minferri (Juneda, Lleida) in the Western Catalonian Plain (Fig. 1). In both sites, canids are interpreted as intentional deposits. Using Perri’s typology (Perri 2017), canids are found in isolated structures (buried alone) and in associated structures (co-buried with humans). In most cases, they are fully or partially articulated skeletons (depositions), but we can also find distinct elements only (elemental). In some cases of full skeletons, the absence of certain anatomical parts was observed, although this is unrelated to any detectable taphonomic process. There are also exceptional cases where the right limbs were missing, as well as a unique case of a dog whose paws were removed before deposition. The absence of certain anatomical parts and the presence of some isolated remains of dogs, such as severed heads, have led to speculations as to their occasional consumption in circumstances unrelated to daily contexts. Furthermore, it is noteworthy that these bones never revealed butchery marks—only in some cases did it reveal disarticulation marks in the occipital condyles and, in a single case, disarticulation marks in the distal joints of the two radii. Foxes at both sites are represented by fully articulated skeletons and never revealed butchery marks.

Situation map of Can Roqueta and Minferri and other sites cited in the text: (1) Bòbila Madurell, (2) Can Gambús, (3) Pinetons, (4) Mas d’en Boixos, and (5) Cantorella
Dogs are thus one element of a variety of faunal depositions essentially comprised of domestic animals (cattle, sheep, goat, pig). There are nonetheless instances of wild species, principally, carnivores and birds (Albizuri 2011a, b; Albizuri et al. 2015; Gómez 2003; Nieto et al. 2014), albeit, in a very low proportion.
Other regional Bronze Age sites (see Fig. 1) with similar burial rites include Bòbila Madurell in Sant Quirze del Vallès Occidental (Boquer et al. 1990; Bordas et al. 1994), Can Gambús in Sabadell (Artigues et al. 2007; Roig and Coll 2016) and Pinetons in Ripollet (Balsera et al. 2011). To the south are also the sites of Pacs del Penedès, the vast settlement of Mas d’en Boixos (Bouso et al. 2004, 2005) and Cantorella, near Lleida (currently under study by A. Nieto). Yet, the findings from these sites are not included in the current study, as they remain unpublished.
Dogs and people, tasks and diet
Humans and dogs share a common history from the Palaeolithic period, with putative dog ancestors dating back for more than 30,000 years BP in the Belgian cave of Goyet (Germonpré et al. 2009) or in the Altai Mountains of Siberia (Ovodov et al. 2011), and fully domesticated animals from the Magdalenian period at Late Glacial times (Horard-Herbin et al. 2014). The human–dog relationship is marked by important functional and economic factors, such as hunting, guarding, protection, herd driving, transport and loading, which can be reflected in signs of affection, such as a caring burial (Albizuri et al. 2011b; Coppinger and Schneider 1995; Morey 2010). This is relevant considering the emblematic character of dogs, which also apply to wolves, their wild counterpart. This explains their appearance in burial contexts (Guagnin et al. 2018; Losey et al. 2011; Morey 2010; Perri 2017; Russell 2012).
Among the many tasks entrusted to the dog, some are natural, such as those related to hunting or surveillance, and others require further training, such as grazing livestock or transporting either by pulling travois, carts or sleds, or carrying weights on their backs. The transport of goods by pack dogs is a subject that has not been extensively studied in Europe, although it has been recorded from historical and ethnographic accounts from Roman times to the beginning of the twentieth century AD (Latham, 2016). An exception is the Peterson’s doctoral thesis, which offers an explanation for the diversity of size and robustness of dog bones from the extremities of a large number of examples from Mesolithic sites in Sweden. The variability is interpreted in terms of the diversity of work and transportation tasks (Peterson 2006). A study in Siberia has also recently identified pathologies among Palaeolithic dogs (Pitulko and Kasparov 2017) which indicate that the task of transport was developed from the very outset of the domestication of the wolf. In the case of Can Roqueta, bent spinous processes of lumbar vertebrae found in three dogs were interpreted as sign of a constant bearing of vertical loads on their back (Albizuri et al. 2011b). This confirms the use of the dog as a pack animal in the Iberian Peninsula during the Early-Middle Bronze Age. These pathologies were also observed among dogs of the Palaeo-Indians of Alabama (Dust Cave) and Illinois (Kuehn 2014; Morey 2010; Warren 2004). Although, they were also observed in non-work related and even wild animals (Latham 2016).
The long journey of dogs in the company of humans is evidenced by a diet that for millennia was a proxy to that of humans, usually reflecting the same general trend in the proportion of animal and plant foodstuffs (Cannon et al. 1999; Guiry 2012; Guiry and Grimes 2013; Noe-Nygaard 1988; Pearson et al. 2015; Pechenkina et al. 2005). But this similarity depends to a great extent on other causes, such as cultural habits and environmental conditions, that can positively or negatively influence the dog’s diet (Ewersen et al. 2018; Losey et al. 2013). For example, a scarce diversity of available environmental resources will imply a forced similarity between the feeding of humans and dogs. In the same vein, the consumption of human faeces will lead to new similarities between the diet of dogs and humans (Tankersley and Koster 2009).
In recent societies, dogs that are destined to work are fed expressly and in a similar way to humans so as to ensure their effectiveness (Isik 2009; Koster and Tankersley 2012), but we have no knowledge of whether this happened in the agro-pastoral societies of the Bronze Age. The control of the dog’s diet would be a determining factor in ensuring its effectiveness in the assigned task, freeing up the time spent on obtaining food and encouraging its dependence on the human group.
On the other hand, the exceptional presence in the Iberian Peninsula of four complete and articulated foxes co-buried with humans at Minferri and Can Roqueta led to other questions, especially when considering that domestic species make up 90% of the animal offerings. The presence of these wild animals creates a problem of interpretation as to their potential commensalism with humans—a hypothesis that ought to be taken into account, since they appear in the same burial contexts as dogs. The fact that they are intentionally deposited poses questions as to the symbolism of these wild animals. Although wild carnivores are generally considered to be dangerous, some studies link them as agents of protection (Peters and Schmidt 2004). Wolves and foxes at times form part of the totemic images (Brück 1999). Therefore, it cannot be ruled out that their presence in burials could be symbolic. Foxes, for example, were recorded as deposits in burials sites in Eurasia during the Neolithic period (Horwitz and Goring-Morris 2004; Maher et al. 2011; Brea et al. 2010), as well as in certain Bronze Age and Chalcolithic contexts (Morris 2011; Onar et al. 2005). This species is poorly represented in the Iberian Peninsula’s archaeological sites, with the exceptions of cases in Neolithic and Chalcolithic contexts in the Province of Jaen and in Portugal (Valera and Costa 2013; Camara Serrano et al. 2016). The four foxes considered in the current study, as well as three other single cases, are exceptional to this region. The other cases in the Northeast are one animal buried with a woman at Camí de Can Grau site (Middle Neolithic) (Martí et al. 1997), one fox found in an isolated structure at Cantorella site (Late Neolithic) (under study by A. Nieto) and a third fox buried with two human skeletons in the Early Bronze Age sepulchral cave of Cervereta (Forcadell and Villalbí 1999).
Aim of the study
The dogs associated with the funerary structures as well as the non-funerary annexes of Can Roqueta and Minferri serve as the basis for this study, since they reflect a direct relationship with the deceased, and this allows a better contrast between their diets and the control that humans could have exerted over them.
The central objective of this study is to identify the level of dependence of dogs on humans and the degree of human control over them by studying their diets through the stable isotopes of carbon (C) and nitrogen (N) in bone collagen. The aim is thus to identify the socio-economic utility of dogs and the level of proximity between humans and foxes. We also want to observe if there are any morphological and feeding differences between the dogs studied in order to propose a line of study on the possible specific feeding of some specimens dedicated to hard work.
Archaeological context: Can Roqueta and Minferri
Description of the sites
The settlements of Can Roqueta and Minferri are references for the Early-Middle Bronze Age of the Northeast region of the Iberian Peninsula due to their extension and their similarities, which facilitate comparative study. Both were occupied between the end of the 3rd millennium and the first half of the 2nd millennium BC. Most of the structures of Can Roqueta that have been subject to radiocarbon dating date back to between 1900 and 1600 cal BC (Carlús et al. 2008). Yet, there are concrete examples that are evidence of a phase corresponding to the so-called Bronze inicial from the middle of the 3rd millennium to the last quarter of the 2nd millennium cal BC (Palomo et al. 2016). The dates of Minferri, in turn, place its occupation between 2100 and 1650 cal BC in the Bronze ple (Nieto et al. 2014; Marín et al. 2017). Both settlements are typologically analogous from the viewpoint of their material culture and their models of insertion in the territory. In spite of their similarities, they are located in distinct geographical and climatological areas: Can Roqueta is in Barcelona’s humid Pre-littoral Plain, while Minferri is in the Western Catalonian Plain, which is characterised by a continental climate (see Fig. 1). Both areas are very fertile and conducive for agriculture, with easy access to natural resources (Carlús et al. 2007; Equip Minferri 1997; Alonso and López 2000).
Both sites have also been the subject of continuous archaeological work since the 1990s (Minferri: Agustí et al. 2005; Alonso 1999; Alonso and López 2000; Equip Minferri 1997; Gómez 2000a, 2000b, 2003; López 2000, 2001; Marín 2018; Marín et al. 2017; Moya et al. in press; Nieto et al. 2014; Prats 2013, 2017; Vila 2018; Can Roqueta: Albizuri 2011a, 2011b; Albizuri et al. 2011b, 2015; Boquer et al. 1990, 1992; Carlús et al. 2002, 2007, 2008; Palomo and Rodríguez 2002; Palomo et al. 2016; Rodríguez et al. 2002). Of particular interest to this study is the fact that both offer a very rich assemblage of human and animal remains, unique to the Iberian Peninsula.
The sites are of the open-air, scattered type, with residential sectors represented by dispersed, small, semi-excavated houses, roofed with wood and branches. Both sites had an economy based principally on agriculture. Features linked to production, storage and burials are dug into the geological substrate between the houses (Figs. 2 and 3). The burials containing both the deceased and grave goods take the form of simple semi-circular pits or, more rarely, complex pits with lateral niches of 2 m, and more rarely up to 3 m, in depth, with a maximum diameter spanning 1–2 m. At times, they are very close to other non-funerary pits also containing animal deposits. Some authors interpreted these structures as ritual pits linked to the burials (Albizuri 2011a, 2011b; Nieto et al. 2014).

Plan of the archaeological sectors of Can Roqueta highlighting the structures dating to the Early-Middle Bronze Age. Detail of Can Roqueta II: funerary structures (triangle) non-funerary structures (circle)

Animal deposits at Can Roqueta. a CRII-736: piglet skeleton, b CRII-583: cow skull, c CRTR-162: dog skeleton, d CRII-481: fox skeleton and e CRII-481: sheep skeleton
Bioarchaeological studies of the sites reveal that their populations were dedicated to cereal farming and livestock breeding (cattle, sheep, goats, pigs). There is also evidence of bronze workshops at each settlement.
Funerary structures and animal deposits
The funerary rites are one of the most remarkable parallels between the two sites, as both comprise structures linked to inhumations with intentional animal deposits and other ritual structures containing animal deposits at less than 10 m from burial pits (Table 1). The chronological relationship between funerary and non-funerary structures can only be proven at present in one case, in which fragments of a single ceramic vessel are shared by both a burial of Can Roqueta (CRII-590) and a non-funerary structure (CRII-591) dug a few meters away. This structure also contained an important assemblage of fauna, highlighted by three dog skeletons. Although concentrated in different areas of the settlements, funerary pits are interspersed with domestic features. One of the most significant manifestations of their burial ritual, as noted previously, is the deposition of animals (especially domestic animals), in particular, dogs.
At Can Roqueta, there are more than 150 Early Bronze Age pits distributed at different sectors of the site: Can Roqueta-Torre Romeu (CRTR), Can Roqueta-Diasa, Can Roqueta II (CRII) and Can Roqueta-Can Revella (CRCRV), as seen in Fig. 2. Funerary structures make up a large proportion of the pits of this chronological phase (60 structures, 220 inhumed individuals). Most were found in the CRII sector: 121 structures, of which 29 are funerary structures (40.5%), containing 180 inhumed individuals.
The human skeletal remains of Can Roqueta include all age categories, but the underrepresentation of certain age classes, such as infants less than 2 years as well as mature and elderly individuals, is noteworthy. In general, the animals found in both funerary and non-funerary structures at Can Roqueta are fully or partially skeletons and isolated remains of domestic animals. Birds and wild mammals are occasionally present, as seen in Fig. 3. According to the minimum number of individuals (MNI) values, the most represented are the domestic animals: cattle, sheep and/or goat, pig and dog, arranged in order of importance (Albizuri 2011a, b).
The Minferri funerary record is the largest in the Western Catalan plain for the first half of the Bronze Age. Funerary practices were described based on ten individuals recovered in four structures (Equip Minferri 1997; Guerrero 1990). The number of localised structures increased significantly after the 2001 and 2006 interventions. Therefore, Minferri currently has a set of anthropological remains unearthed in 24 silo-shaped structures, corresponding to a minimum of 56 individuals (Fig. 4). Of the 182 structures, 12.3% were reused for human burials (MNI = 51).

Plan with the position of the Early-Middle Bronze Age Minferri animal deposits. Detail of archaeological sectors: a area 2 and b area 9
Minferri’s largest concentration of burials (63%) is in sector 2 of zone 2, with a total of 32 individuals in the 10 structures. A second important concentration of tombs (zone 9) is located northwest of the main excavation area (zone 2), with six individuals placed in five different structures. The population includes all age categories. There is a significant presence of perinatal individuals and infants, as well as a low number of young people, and only a single elderly individual (Agustí et al. 2005). Grave goods and offerings are not widespread at Minferri. The few exceptions of these funerary structures contain pottery and meat offerings corresponding mainly to full or partial skeletons of cattle, sheep or goat, canids and occasionally foxes and birds, but never pigs, as shown in Fig. 5. There are also other non-funerary structures containing full or partial skeletons of animals (cattle, followed by sheep/goat, pigs and dogs).

Animal deposits at Minferri: a MIN-355: female burial with a complete fox skeleton, b MIN-88: female burial with two complete fox skeletons, c MIN-405: skeletons of two cows and two dogs, d MIN-405: skeletons of one cow and one dog, e MIN-361: dog skeleton, f MIN-354: dog skull, f MIN-354: dog skeleton, h MIN-367: dog skull, i MIN-367: dog skeleton and j MIN-367: dog skull
In summary, the comparison of the findings of animal offerings at Can Roqueta and Minferri suggest the presence of similarities and differences. The first similarity is the predominance of domestic species and the equal representation of dogs with respect to other domestic animals. Another noteworthy aspect is the presence of fully articulated skeletons of foxes at both sites. The dogs are numerous and appear both in tombs and in structures close to the tombs. The most pronounced differences, by contrast, are the higher representation of cattle at Minferri (at times deposited fully) and that of pig at Can Roqueta.
Food resources
The agricultural and plant data at Can Roqueta for the Early-Middle Bronze Age is scarce and comes from a single sediment sample of the Can Revella area (volume: 20 l) and the manual collection of many acorns (1050) during the excavation of various funerary structures. The results of the analyses illustrate the variability of crops consumed by humans and animals. Moreover, acorn offerings in certain tombs point to a symbolic and economic value of plant resources. Can Roqueta’s main crop is barley (Hordeum vulgare s.l.), followed by the token presence of naked wheats (Triticum aestivum/durum) and legumes (probably pea, Pisum sativum). Unlike Western Catalonia, there is no evidence of flax in this territory and in this chronological framework. Yet, it was observed that oleaginous plants are often underrepresented, as their remains are only preserved by carbonisation. However, as noted, acorns abound in various features in the Can Roqueta/Revella sector, suggesting that this fruit could be ground and could have served as a diet supplement. On the other hand, it can be observed that acorns are common to silos and funerary structures elsewhere in the Bronze Age in the Iberian Peninsula (Alonso et al. 2016; Pereira and García 2002).
The plant food resources of Minferri’s Middle Bronze Age sequences are, for the most part, winter cereals gleaned from the systematic sampling of 58 excavated structures, yielding a total of 4559 l of sediment. Although, their number is quite high (5884 grains), their density is very low (only 0.6 finds per litre of sediment). There is a predominance of common/durum wheat (Triticum aestivum/durum), represented by more than 50% of the remains of cultivated plants appearing in 59% of the structures. The importance of this cereal is even more evident when compared to the next most common type of cereal, dressed barley (Hordeum vulgare), represented by only 16.9% of the remains and present in only 17% of the structures. Emmer (Triticum dicoccum), although present, is minor. Furthermore, the total absence of legumes at Minferri is remarkable. There is, nonetheless, evidence of flax cultivation (Linum usitatissimum) (Alonso 1999; Alonso et al. 2006). Harvested fruit comprises acorns (Quercus sp.), mastic (Pistacia lentiscus), blackthorn (Prunus spinosa) and blackberry (Rubus fruticosus) (Alonso et al. 2016).
The study of bones identified as food waste at Minferri and Can Roqueta was carried out on an assemblage comprising 484 remains from 70 silo-type structures (Gómez 2000a, b; Albizuri 2011a). The results indicate a spectrum of domestic mammals: sheep (Ovis aries), goat (Capra hircus), cattle (Bos taurus), pig (Sus domesticus), rarely dog (Canis familiaris) and wild mammals, such as lagomorphs (Oryctolagus cuniculus, Lepus sp.) and red deer (Cervus elaphus). The consumption of domestic species (excluding dog) was clearly predominant. Wild species, in turn, were only occasionally exploited for the human diet. At Can Roqueta, such wild species also include marine mollusc (Palomo et al. 2016), although, this evidence is very limited. Sheep and goat are the most commonly represented domestic species at both sites, with pig being the third most common domestic animal ahead of cattle (Albizuri 2011a).
The evidence indicates that sheep and goats were the most frequently sacrificed as adults or sub-adults, a clear indicator of an attempt to manage the reproduction of the herd so as to profit from their milk, cheese and wool, among other by-products. Cattle, in turn, were mostly slaughtered as sub-adults, after attaining their optimal weight. Only a small number (probably needed for breeding, milk and work) surpassed the age of 36 months. Furthermore, findings of perforated vessels interpreted as strainers at both sites serve as evidence of the processing and filtering of rennet in the preparation of fermented products such as cheese and yogurt. Pigs were sacrificed for the most part around 12 months, an age corresponding to their adult weight. This is evidence of their exploitation for meat. Adults of both sexes were nonetheless retained for reproductive purposes.
Materials and methods
The sample set
In the two sites studied, canids appeared primarily in funerary and ritual structures. For this reason, we have selected 28 funerary structures containing human remains and canids for study, as seen in Table 1. The sampling was completed with samples of deposits containing dogs and other animals in 13 non-funerary structures located in the vicinity of the burials.
For the isotopic study, we selected the best-preserved remains available at the time of the study. Thus, we studied 32 dogs (21 from Can Roqueta and 11 from Minferri), 1 large canid, presumably a wolf, from Can Roqueta and 4 foxes, 3 from Minferri and 1 from Can Roqueta (as seen in Table 2). Only one of the dogs studied was found in a structure far from the funerary zone (MIN-150). This was included in the sample to determine if there are differences between the diet of these dogs and the diets of the dogs included in the funerary structures. The only canid that can be considered to be wild is an animal that we tentatively identify as a wolf (Canis cf. lupus) because of its large size in comparison with the other dogs in the structure (Albizuri et al. 2015).
The assignment of age ranges of canids is based on the period of natural lactation as well as bone and reproductive maturity (see Table 2). They are called puppies in the first 3 months of life, after which they cease to breastfeed definitively. Young dogs are those in the process of replacing their teeth and they have not yet reached bone and reproductive maturity—between 4 and 10 months of age. The adult dog category begins between 10 and 12 months, when their skeleton reaches bone and reproductive maturity. The age range of the canids studied was determined from the degree of epiphyseal fusion of their long bones and dental eruption or wear (Horard-Herbin 2000; Piérard 1967). Sex was estimated by the presence/absence of the penile bone, only when the skeleton was whole or sufficiently represented.
Canid biometric data were obtained according to the work of Von den Driesch (1976) and Onar et al. (2005). Measurements were recorded with a digital calliper on the landmarks described in the literature. The robustness index and body mass were calculated following procedures stated by Onar (2005) and Onar and Belli (2005). The robustness index is based on the mid-shaft diameter (SD) and the great length (GL) of radius and tibia, analogous bones representing the fore and hind limbs, respectively. Body mass was calculated from the femur mid-shaft circumference. The estimate of height at the withers is based on the limb bones great length, according to Harcourt (1974).
The human sample comprises a total of 64 individuals. From Minferri, we have 8 adult males, 14 adult females and 15 sex-indeterminate remains (9 infants, 2 young adults and 2 adults). From Can Roqueta, we have 5 adult males, 8 adult females and 16 sex-indeterminate remains (6 infants, 7 young adults and 3 adults). The low number of males studied is probably the result of a demographic bias in recovered human remains and the difficulty of sex assignment in poorly preserved skeletons. The biological study of the human samples includes the estimation of sex and age at death. Identification of the sex of adult humans was based on the morphological observation of the coxal bone (Bruzek 2002). In absence of the hip bone, sex was also determined according to robustness and morphological characteristics of the skull (Buikstra and Ubelaker 1994; Ferembach et al. 1979). The age estimation of the sub-adult humans was based essentially on dental criteria, evaluating the stages of maturation and eruption of the deciduous and permanent teeth (AlQahtani 2009; Moorrees et al. 1963a, b). The degree of cranial and postcranial bone maturation was also taken into consideration (Scheuer and Black 2000). Infants were identified by the length of their long bones (Fazekas and Kosa 1978; Kosa 1989).
Since the determination of the age of death of adult individuals is very broad and precision is therefore difficult to attain, other criteria had to be considered, including traces of degenerative diseases, the degree of occlusal wear, the stage of pubic symphysis (Todd 1921, in Garvin et al. 2012) and the stages of formation of the auricular surface of the ilium (Lovejoy 1985, from Garvin et al. 2012).
For the isotopic study, the standard age groups were condensed into three groups based on their presumed type of nutrition and, in the case of adults, on their reproductive potential: (1) individuals under 4 years (including perinatal) who could yield isotopic signatures indicative of breastfeeding; (2) young adults from 5 to 14 years of age who no longer consume maternal milk, but have not yet attained physical maturity and possibly had access to a different diet from that of adults; and (3) adults surpassing 15 years of age. In these cases, when possible, the sexes are considered separately.
Other animal species for establishing an isotopic baseline comprise 19 domestic ungulates: 11 cattle (Bos taurus), 6 ovicaprines (Capra hircus/Ovis aries) and 2 pigs (Sus domesticus) (see Table 1).
Methodology of the isotope analysis
Measuring the isotopic composition of collagen from the most compact parts of the bones of adults results in values that offer data about the average diet over a period of time, estimated between 5 and 10 years (Schwarcz and Schoeninger 2012) before the death of an individual. Collagen from non-compact (trabecular) areas of the bones of human adults is representative of the average diet over a period of about four years. The bone collagen turnover rate in dogs, in turn, is much higher than that of humans. Thus, the rate among young adult dogs is reported to be between 6 months and 3 years (Fischer et al. 2007).
The standard treatment of bone collagen extraction begins with at least ten ultrasonic bath wash cycles lasting about 5 min, alternating with acetone and distilled water baths until no turbidity is observed. The samples are then dried for 48 to 72 h at room temperature. About 1 g of each bone is then ground in an agate mortar until reduced to particles measuring < 0.3 mm.
Between 250 to 350 mg of bone powder was used for the extraction of bone collagen following the procedure advanced by Bocherens et al. (1997). The pulverised sample was subject to successive digestions in HCl 1 M (20′) and NaOH 0.125 N (20 h) at room temperature to remove both the bone mineral fraction and potential contaminating organic materials. Each digestion was followed by microfiltration with 5 μm filters to remove the residue. Microfiltration allows separating the dissolved inorganic matter, as well as both soluble organic acids and bone organic components (including degraded collagen) from the well preserved collagen. Finally, the collagen was diluted by incubation in HCl 0.01 M at 90 °C for 17 h, and was subsequently lyophilised and then subjected to combustion analysis in an isotopic ratio mass spectrometer (IRMS), Finnigan Mat Delta Plus coupled to an elemental analyser Carlo-Erba 1108, with an analytical reproducibility greater than 0.2‰ for carbon and greater than 0.2‰ for nitrogen. The IRMS analyses were carried out in the Unit of Instrumental Techniques of Analysis of the Services of Support to Research (SAI) of the University of A Coruña. The internal standard was acetanilide, and quantifications were duplicated. The results are presented under the delta (δ) notation, which reflects the proportion between both isotopes in the sample in relation to the proportion in an international standard, in this case, the Vienna PeeDee Belemnite (VPDB) and atmospheric air (AIR) for carbon and nitrogen respectively, as shown in the equation:
where X is the heavier isotope and R is the 15N/14N or 13C/12C ratio.
All samples had well preserved collagen according to the usual quality criteria: percentage of carbon in collagen was higher than 13%, percentage of nitrogen in collagen was higher than 5% (Ambrose 1990) and C/N atomic ratio in collagen was between 2.9 and 3.6 (DeNiro 1985).
Statistical methods
Statistical tests were carried out using the Paleontological Statistics (PAST) software package (Hammer et al. 2001), version 3.20. The statistics include linear correlation analysis, Cluster Analysis (Ward’s minimum variance method; Ward 1963) as an exploratory method to observe affinity in isotopic values between individuals and a hypothesis contrast to compare distributions of isotopic values between groups. To compare isotopic values between groups, due to the small number of individuals in some of the groups, we chose to perform a nonparametric test: a two-tailed Mann-Whitney U test (Mann and Whitney 1947) with Bonferroni corrected p values and a significance level set at 0.05.
Results
Age and physical traits of dogs in burials
The age at death of the dogs in both sites reveal fairly similar trends. Among the 21 dogs of Can Roqueta, 16 are adults ranging from about 1 year of age up to 3 to 4 years of age. Of the remaining individuals, two fall into the category of old adults, three are puppies between 1 and 3 months of age and two are young dogs around 6 months of age. In Minferri, out of a total of 11 dogs, 4 were adults. Of these, only one is over 4 years old, and the rest are between 2.5 and 3.5 years old. Four individuals are sub-adults (6 to 10 months old), and two dogs were too poorly preserved to determine their age. In summary, dogs in burials seem to have been preferably between 2.5 and 3.5 years of age; that is, they were individuals in transition towards adulthood or in its early stages.
Certain biometric traits yielded an approach to the morphology of the canid populations in both sites, as seen in Fig. 6a, b. All of them are medium-sized animals on the basis of their height at withers (see Tables 2 and 4). The data obtained from the fore and hind limbs of the animals with complete epiphyseal fusion showed average height at withers values of 48.6 cm at Can Roqueta and 44.5 cm at Minferri (Fig. 6a).

Osteometric data of dogs. a Comparison of dog shoulder height by site. b Comparison of radius and tibia dimensions by site
The robustness index was calculated from a small number of animals (Table 3). The results again reflect a greater span in terms of size of the dogs of Can Roqueta (Fig. 6b). The difference in average height between the dogs of the two sites is repeated in the robustness index, with notable differences—especially in the tibia robustness index, in which the dogs of Can Roqueta clearly surpassed those of Minferri.
On the other hand, body weight estimations yielded greater values at Can Roqueta (between 14 and 19.4 kg) than at Minferri (between 8.4 and 14.5 kg), as seen in Table 3.
Dogs at Can Roqueta are therefore generally larger and more robust than those at Minferri. They also revealed pathologies in three cases (CRII-481-1, CRII-591-1 and CRII-505-1) in the lumbar neural apophyses that showed bent spinous processes (Albizuri et al. 2011b). These pathologies have been related to the use of the dog as a work animal, especially in sled dogs (Warren 2004), particularly in specimens from North America and Artic Siberia (Latham 2016). However, these pathological alterations could be related to the age-associated degenerative disease spondylosis deformans, which affects even wild specimens (Latham 2016), although veterinary studies on these pathologies have determined a broader casuistry (Lawler et al. 2016).
Characterising foxes is much more complex, especially considering their small number. The old fox in funerary structure CRII 481 at Can Roqueta, represented by 130 bones, is almost complete (lacking the sacrum, the metatarsals and most of the phalanges). It was unearthed in anatomical connection (Albizuri et al. 2015). The absence of a penile bone indicates that the animal is probably female (Albizuri et al. 2015). This finding is also in line with osteometric data of foxes from current Anatolia (Onar 2005). It is noteworthy that this animal presents several fully healed fractures that occurred before its death. Towards the end of its life, this animal suffered a fracture of the right hind limb that affected the distal middle end of the fourth metatarsal, as evidenced by adhesion of the bone with the metatarsals to each side and the fusion of the distal fibula with the tibia, as seen in Fig. 7. Both fractures occurred at the same moment. The vascularization observed in the callus of the fourth metatarsal indicates that the healing process was still active. The null angulation of both halves of the diaphysis of MT4 is compatible with the immobilisation of this area, presumably by humans.

CRII-481 right hindlimb of a fox with fused fractures on the distal fibula and the fourth metatarsal: a palmar view of the five metatarsals. The arrows highlight the scars on the shaft, on the distal tibiofibular area and on the fourth metatarsal, b dorsal view of the metatarsal and the healing in the fourth metatarsal that affected the other metatarsals, c plantar view of the metatarsals and the healing area in the fourth metatarsal that affects the other metatarsals
The three foxes from Minferri correspond to adults deposited whole and in anatomical connection. A female and a male between 2 and 4 years of age were placed next to a woman in burial MIN-88. A third, in structure MIN-355, is identified as an 18-month-old male placed next to the isolated femurs of a woman.
Results of the isotopic analysis
A total of 120 samples yielded collagen in line with the requested standards of quality, that is, yield values greater than 3.5%, atomic C/N ratios between 2.9 and 3.6 (DeNiro 1985) and percentages of carbon and nitrogen greater than 13 and 5%, respectively. The detailed data, together with the values obtained from δ13C and δ15N, are listed in Table 4 (domestic ungulates), Table 5 (canids) and Table 6 (humans) and represented in Fig. 8.

Bivariate diagrams of the isotopic signatures of C and N at Can Roqueta and Minferri. a, b All individuals. c d Mean values with error bars equivalent to 2 SD except for groups with less than three individuals that are represented individually
The domestic ungulates from both sites (Table 4) serve to establish an isotopic baseline and lead to a better interpretation of canid and human diets. Cattle (Bos taurus) from Minferri (n = 6) show fairly homogeneous values of δ13C = − 19.8 ± 0.5‰, from − 19.0 to − 20.3‰. Yet, the values of nitrogen are more dispersed (δ15N = 6.4 ± 1.4‰, from 4.5 to 7.6‰). The five Bos from Can Roqueta offer δ13C values analogous to those of Minferri (average δ13C = − 19.4 ± 0.6‰, from − 18.9 to − 20.1‰), and δ15N average values and dispersion slightly lower than that of Minferri (average δ15N = 5.8 ± 0.7‰, from 4.8 to 6.2‰). Sheep and/or goat (Ovis/Capra) δ13C values are fairly similar at both sites, in spite of those at Can Roqueta (n = 2; − 19.2 ± 0.4‰) being slightly higher than those at Minferri (n = 4; − 19.7 ± 0.6‰). The average δ15N values of sheep and goat at Minferri reveal higher values (7.5 ± 1.7‰) than those at Can Roqueta (5.1 ± 1.2‰). Both sites offer widely dispersed δ15N values for sheep and goats: a dispersion that is not related, in principle, to the species. This dispersion probably stems from the fact that these animals were fed in different environments. Finally, the data for the pigs of Minferri (n = 2) revealed more negative δ13C values (− 20.2 ± 0.1‰) and higher δ15N values (8.6 ± 0.9‰) than other ungulates.
The isotopic values of the dogs (Canis familiaris), foxes (Vulpes vulpes) and one large canid, a wolf (Canis cf. lupus), are listed in Table 5. The wolf from Can Roqueta shows relatively high values for both isotopes (δ13C = − 18.8‰; δ15N = 9.4‰), greater than the rest of the dogs from the same site and even greater than that of the dogs at Minferri, which generally have higher values than those at Can Roqueta. Adult Minferri dogs (n = 7) yielded δ15N values (average 8.7 ± 0.4‰, from 8.5 to 9.1‰) that were greater than those of Can Roqueta (n = 16; average δ15N = 7.7 ± 0.6‰, from 6.5 to 8.6‰), while δ13C values are also slightly more positive (average − 19.0 ± 0.3‰, from − 19.3 to − 18.5 at Minferri and − 19.3 ± 0.3‰, from − 20.1 to − 18.8‰ at Can Roqueta). The values of the young dogs at Minferri (n = 4) hardly differ from those of adults (δ13C = − 18.7 ± 0.3‰, δ15N = 9.0 ± 0.3‰). At Can Roqueta, the two young dogs revealed very different values for both isotopes—practically at both ends of the adults’ distribution. The δ15N average value of the three puppies is similar to that of adults (7.8 ± 0.9‰), but offers slightly higher δ13C mean values (δ13C = − 18.7 ± 0.3‰). However, one of the three puppies at Can Roqueta shows clear δ15N enrichment, as expected of an animal at the age of lactation.
The four foxes (three from Minferri and one from Can Roqueta) revealed different values. At Minferri, the average δ13C values are slightly more negative than those of adult dogs (− 19.4 ± 0.3‰), and the δ15N values are slightly higher than the adult dogs (9.3 ± 0.6‰). The old fox from Can Roqueta shows an isotopic signature far from the adult dogs at the site, and much farther from the foxes at Minferri. Its combination of high δ13C values (− 18.0‰) and a relatively low δ15N (7.8‰) only finds some similarity with some puppies and one sub-adult from the site, yet it still differs from them in its less negative δ13C.
As for humans, adults at both sites (Table 6) showed quite similar distribution in isotopic values, with averages slightly higher at Minferri (n = 24; average δ13C = − 19.1 ± 0.5‰, average δ15N = 9.8 ± 0.7‰) than those at Can Roqueta (n = 15; average δ13C = − 19.5 ± 0.3‰, average δ15N = 9.1 ± 0.7‰). The apparent similarity disappears when males and females were analysed separately. In Can Roqueta, there is a clear distinction between sexes in δ15N, with the female (n = 8) mean value of 8.7 ± 0.2‰ being lower than that of the site’s males (n = 5; δ15N = 10.0 ± 0.7‰) and lower than the mean of individuals of both sexes at Minferri. Females at Minferri, in turn, showed a greater dispersion of both isotope values (n = 14; δ13C = − 19.3 ± 0.6‰, δ15N = 9.6 ± 0.8‰) than the males (n = 8; δ13C = − 18.9 ± 0.2‰, δ15N = 10.1 ± 0.3‰). Young humans (between 5 and 15 years of age) are isotopically indistinguishable from adults. At Can Roqueta specifically, their average values are equal to those of females. Both sites clearly share high δ15N values in most children (less than 4 years of age). However, there are exceptions: the isotopic signatures of one child from each site are indistinguishable from those of young and adult humans, and at Can Roqueta, the case of a perinatal individual attains much higher δ15N values than those expected when compared to the adults. This is reflected in the great dispersion of the values of the children at Can Roqueta, as opposed to those at Minferri (see Fig. 8).
Discussion
Isotope baseline, environment and inter-site comparison
Today, there are climatic differences between the sites at Can Roqueta and Minferri. Although, both share Mediterranean conditions with a tendency towards a continental climate, and although Can Roqueta is closer to the coast than Minferri, the littoral mountains act as a barrier. The average annual rainfall at Can Roqueta ranges between 550 and 700 mm. Minferri’s continental climate, in turn, is more pronounced, with greater aridity and an average annual rainfall of about 400 mm. The two sites presently also share similar average temperatures, although those of Minferri are slightly higher (Bolós 1979). Carbon isotopic discrimination studies of Pinus halepensis coals from Minferri (Ferrio et al. 2006) indicate a higher average annual rainfall in the past, between 464 ± 50 and 576 mm. Unfortunately, this type of data is not available for Can Roqueta. Yet, its lower aridity during the period of study appears to correspond to a regional trend (Ferrio et al. 2006). If this were the case, the climatic difference could account for the higher general δ15N values at Minferri, since aridity yields an increase in δ15N values in soils and plants (Heaton 1987). The climate also affects the δ13C baseline, as aridity and insolation are positively correlated to δ13C values in plants, while rainfall is negatively correlated (Szpak et al. 2013; Goude and Fontugne 2016). It is therefore possible that the difference in isotopic values between the fauna and humans of both sites is a result of slightly different isotopic baselines.
For δ15N, the average difference between both sites is + 0.6‰ in Bos, but rises to + 2.4‰ between Ovis/Capra. Thus, δ15N values at Minferri among all the herbivore domestic ungulates are + 1.2‰ higher than those of Can Roqueta, while ungulates showed lower values (− 0.3‰). However, the dispersion of the ovicaprine values seems to indicate that not all of them were fed in the same environment. As a whole, strict herbivores from both sites do not differ significantly in their isotope values (Mann-Whitney U test, U = 18.5, p = 0.1102 for δ13C and U = 15.5, p = 0.058 for δ15N).
We only have data from two Minferri pigs, of which there is a clear isotopic difference between them. Clearly, one of the pigs is more omnivorous than the other, and probably received leftover human food in its diet.
Adult dogs also showed more positive mean values at Minferri than at Can Roqueta, with a significant difference of + 1.0‰ for δ15N (U = 2.5, p > 0.0001) and + 0.3‰ for δ13C (U = 29, p = 0.055). Lastly, Minferri adult humans are enriched with + 0.7‰ for δ15N and + 0.4‰ for δ13C over those at Can Roqueta. Both sites have slightly different medians for adult human isotopic values (Mann-Whitney U test, U = 82.5, p = 0.0022 for δ13C, U = 106, p = 0.0172 for δ15N).
Thus, it is not possible to compare the δ15N or δ13C values of the two settlements directly, given the variation of the mean differences in each group (ungulates, canids, humans). Therefore, we are going to consider both sites independently.
Canid diet
When defining the trophic level of an organism by studying stable isotopes, it is common to compare the isotopic signals of the individual in question with the baseline values obtained from individuals with an unequivocally herbivorous diet. In our case, we could use as baseline the δ15N values of cattle and even of sheep or goat of each deposit. Thus, the adult dogs at Can Roqueta showed an offset of + 1.9‰ on the cattle (+ 2.1‰ with respect to all herbivorous ungulates), lower than the minimum of + 3‰ expected for a trophic level (Bocherens and Drucker 2003). In Minferri, the offset is + 2.3‰ in relation to the cattle (but only + 1.9‰ if all herbivorous ungulates are included). We can therefore say that the diet of the dogs from both sites was not particularly rich in animal protein, as would be expected in a wild carnivore. In fact, the wolf at Can Roqueta does reach a characteristic carnivore offset (+ 3.8‰), while the fox at this site resembles dogs in its δ15N value. In the Minferri foxes, the δ15N offset is + 2.9‰ over the cattle.
However, in defining the trophic level of dogs with respect to herbivorous ungulates, we cannot resort to a simple calculation of the offset of the mean values of δ15N. In the case of Can Roqueta, adult dogs have a high dispersion in their values of δ15N for a mean value to be representative of the group. In the case of Minferri, the herbivorous ungulates present extended values of δ15N. In addition, we cannot lose sight of the differences in δ13C that could respond to the introduction of foods with particular signatures, such as marine protein or C4 plants (Chisholm et al. 2006; Schwarcz and Schoeninger 1991, 2012). Therefore, we use a cluster analysis to group individuals together according to their similarities in the values of both isotopes.
The cluster analysis of all the adult animals from each site, as shown in Fig. 9, yielded relevant data. All the dogs at Minferri (Fig. 9a), including the specimen in structure MIN-150, outside of the main assemblage, as well as the foxes, align with the values of one pig and two Ovis/Capra. The pig probably had an omnivorous diet, perhaps including leftover food. Ovicaprines, however, must have come from other environments, given their high δ15N signal compared to other herbivores, which are plainly separated in another cluster. The dogs at Can Roqueta (Fig. 9b), on the other hand, split into two clusters: a set of eight dogs is grouped together with the fox and the wolf, while the remaining eight dogs cluster with some herbivorous ungulates. A statistical analysis (Mann-Whitney’s non-parametric U test) indicates that at both sites, the groups obtained are strongly determined by differences in δ15N rather than δ13C, and that the difference in δ15N median of each cluster is highly significant, and carbon signature is only of relative importance in a secondary sense.

Cluster analysis of adult canids (dogs, foxes and wolf) and domestic ungulates (cattle, sheep/goat, pig) of a Minferri and b Can Roqueta. Results of the Mann-Whitney U test for inequality of medians and distributions are placed between the compared clusters
If the different values observed in δ15N are related to a greater or lesser consumption of animal protein, it is interesting to compare these values with osteometric data to see if the greater intake of animal protein is reflected in the size or robustness of dogs. Figure 10 shows the results of comparing both parameters. Unexpectedly, neither height at withers (Fig. 10a), robustness index (Fig. 10b) nor estimated body mass (Fig. 10c) showed a positive correlation with δ15N values in adult dogs. In fact, the correlation is negative at the height at withers and strongly negative for body mass. The parameters defining the robustness of the correlation (correlation coefficient r, standard error of the correlation coefficient SEr and probability p) indicate an unexpected strong relationship between the lower intake of animal protein and the increase in body size and especially in body mass (see Fig. 9). In summary, dogs with a greater body mass have lower isotopic values, indicating lower consumption of animal protein and, accordingly, a higher input of vegetal food. Interestingly, some of these also show spine pathologies. It could be argued that the scarcity of proteins from animal origin could have caused such pathology in these dogs. However, a veterinary study and diagnosis was made using microcomputed tomography (Philips, Brillance CT 16 Slice®) (Albizuri et al. 2011a, b). The diagnosis was due to lateral deformation from weight borne from an early age. The specimens analysed were complete, with no associated infectious, degenerative or traumatic pathologies in any other part of their skeleton, which would be expected in the case of common ageing processes (Latham 2016), not forgetting that the affected individuals are relatively young (between 1 and 2.5 years). Deformations due to taphonomic causes have also been discarded. The vertebral bodies were normal, but different spinous processes presented a lateral curving in different directions (right, left). This fact rules out a deformation by infectious causes, since in such a case, all the spinous processes would be deflected in the same direction (Lawler et al. 2016). In addition, the radiological study showed that the spinous processes show a thickening of the cortical bone only in the affected section of vertebrae, which is a response to the continuous weight of vertical loads on the lumbar region, possibly being felt from an age when their bones were still in the process of ossification (Albizuri et al. 2011a, b).

Correlation between the isotopic signature of N and the osteometric data of certain Minferri and Can Roqueta dogs: a robustness index, b average height at withers and c body mass. Squares represent young dogs while circles are adult ones. Circles in bold are individuals with vertebral pathologies
Human versus canid diet
Interpreting the diet of the canids at the sites of Can Roqueta and Minferri from the isotopic data requires comparisons with the values of other domestic animals, particularly, with humans. This study has therefore included data from the humans most closely linked to canids, that is, those either sharing burial structures with these animals or those in burials near non-funerary structures containing canids. Hence, when considering the isotopic values of the adult humans of Minferri and Can Roqueta and the values of the respective assemblages of domestic ungulates (see Fig. 8), the 15N enrichment (+ 3.5‰ in Can Roqueta, + 3.0‰ in Minferri) is in the lower limit for one trophic level (O’Connell et al. 2012), suggesting that the overall diet at both settlements was not especially rich in animal protein. The average δ13C offset between humans and herbivores was also reduced (− 0.2‰ in Can Roqueta, + 0.7‰ in Minferri), suggesting that most of the protein came from local terrestrial environment, and no marine or freshwater resources were consumed routinely (Chisholm et al. 2006). In Minferri, the variety of human isotopic values is greater than in Can Roqueta, which points to a greater variety in feeding practices.
Regarding potential differences in animal protein consumption between sexes, females of Can Roqueta showed lower values of δ15N than the males, most of which clearly reflect a higher intake of animal protein than the rest of the individuals, including the canine identified as a wolf. On the contrary, in Minferri, some females reach values of δ15N that are equal to those of the males or even surpass them.
High δ15N values in children younger than 4 years of age are related to breastfeeding, since maternal milk in humans increases δ15N values between 2 and 3‰ with respect to maternal isotopic values (Fogel et al. 1989; Fuller et al. 2006). Children at both Minferri and Can Roqueta show to a greater or lesser degree signs of their stage of lactation, thus, barring the two aforementioned exceptions.
The cluster analysis applied to all dogs and humans revealed different patterns at each site. In Can Roqueta, as shown in Fig. 11, most children form a group that is clearly distinct from all other humans. Most males are grouped together with the wolf. Dogs fall into two groups: (1) dogs with higher input of vegetables (now including the fox); and (2) dogs that cluster with a group of humans, including several young humans, as well as a male and a female. Mann-Whitney U tests show that differences in average δ15N between clusters are highly significant.

Cluster analysis of canids (dogs, fox and wolf) and humans of Can Roqueta, with Mann-Whitney U test results between compared clusters
At Minferri, as shown in Fig. 12, all dogs appear in a group, including young humans and some females, that is marked with lower δ15N values. Two male foxes form part of this group, which is in line with a diet marked by a lower animal protein intake. A third fox, in turn, a female, is associated with another group of female humans with values closer to males, which also indicate a greater consumption of animal protein.

Cluster analysis of canids (dogs and foxes) and humans of Minferri, with Mann-Whitney U test results between compared clusters
Isotopic signatures indicate that dogs at both sites did not consume the same food, nor in the same proportion as humans, but there is an overall similarity, especially with females and young people. The dog isotopic values at both sites indicate an intake of C3 plants, evidencing their mixed diet. By contrast, the only canid classified by its osteometric data as a wolf (Canis cf. lupus) reveals higher values, indicating that it consumed more meat than the other canids, although this is below the level of human males who have more animal protein in their diet. Therefore, the isotopic data does not confirm the classification based on the archaeozoological analysis and raises questions about whether this canid was a larger dog, even a hybrid that fed differently.
The overall isotopic data of humans and dogs shows great correspondence with the palaeoeconomic data gleaned from agriculture and livestock research, which points to the development of cereal crops and legumes, as well as the exploitation of wild plants and products yielded by the domestic animals (meat and milk). Thus, the values of dogs could mirror the human diet—a line of thinking advanced by most isotopic studies that include dogs and humans (Cannon et al. 1999; Guiry 2012; Guiry and Grimes 2013; Noe-Nygaard 1988; Pearson et al. 2015; Pechenkina et al. 2005). Previously reported δ15N differences between archaeological humans and dogs were, on the average, 2 to 3‰ lower in dogs, although, in some cases, both groups showed indistinguishable nitrogen signatures (for a summary, see Guiry 2012). Adult humans from Can Roqueta and Minferri showed δ15N values that are + 1.4 and + 1.1‰ higher, respectively, than adult dogs: a slim difference that indicates their overall similar trophic level. However, except in the case of the robust dogs of Can Roqueta, who have a high proportion of cereals in their diet, the results of the isotopic analysis do not allow us to affirm that the humans fed their dogs specifically. Dogs have an opportunistic eating behaviour, so they could have accessed food waste or human faeces, although, in the case of coprophagy, the values of δ15N would increase, since in humans the faeces are enriched with 15N in the diet (Kuhnle et al. 2013).
The ethnographic parallelisms
In Minferri, dogs cluster clearly within certain human groups (females and youth), at times, from the same structures. Moreover, a similar trend is also discernible at Can Roqueta between a group of dogs and young humans. This correspondence at both sites could indicate that females and young people dedicated greater care to dogs. This type of relationship was observed in the eighteenth century AD among the Hidatsa of North Dakota, whose women were responsible for the training and feeding of these animals (soup of corn and meat scraps), as they were a precious asset, serving as pack dogs, especially for collecting wood (Wilson 1924). The male diet at Can Roqueta and Minferri, in turn, differs from the female diet and that of most of the dogs. Even if the sample of males is smaller than that of females and young people, this distance could potentially indicate a division of labour.
The dogs at both sites share a morphological homogeneity that could point to a certain control over their reproduction. Dogs in most of the funerary structures are young animals (2.5–3.5 years), showing complete epiphyseal fusion with weights corresponding to that of adults. This homogeneity, combined with the data on their diet, leads to speculation as to a selection based on certain traits. Nonetheless, there are differences in height and body volume that indicate a greater corpulence among the Can Roqueta assemblage. Can Roqueta also has a group of dogs with very low δ15N signatures and a greater body mass, that had a singular diet rich in cereals. Certain animals from this group show vertebral pathologies that suggest they served as pack animals. Thus, some dogs of Can Roqueta, that appear to have been assigned to specific tasks requiring a great amount of energy, could had been fed a diet rich in carbohydrates. Cereals are not a foreign food for dogs, because—parallel to the development of agriculture—dogs, like humans, were developing ways of improving digestion of starch through the multiplication of copies of the amylase gene (Ollivier et al. 2016). Ancient written sources confirm this type of food in the canine diet: Columela, Hispano-Roman scholar of the first century AD, recommended for the feeding of dogs “barley flour mixed with whey” or “spelt or wheat bread, mixed with cooked bean broth” (Columela 7.12.10; Álvarez de Sotomayor 1824).
The diet of the dogs at Minferri, by contrast, appears to be more uniform, although they must have also carried out assigned tasks. Work dogs, in order to ensure their performance must not go hungry. In some cases, their food is equivalent to that consumed by humans (Koster and Tankersley 2012). In current cases, this control is visible: the food offered to dogs in the Siberian region of Lake Baikal, for example, is cooked separately, and although it is very similar to human nutrition, it includes less pleasing items, such as tendons, hoofs, periostea, intestines and blood (Bocherens et al. 2015). Moreover, food preparation is at times given special care, such as when dogs must travel long distances in the company of their herds, as is currently the case of the Kangal race in Turkey, which are ancestrally nourished with a mixture of barley flour, oats and wheat with milk whey or milk and yogurt, called yal. Only on very rare occasions are they offered leftovers of lamb bone. Meat is normally available to dogs only in the form of what they can catch, usually rodents in the desert or around the village, or an odd bird or hare (Isik 2009). Ethnographic research on the preparation of food for dogs in Haut-Diois, France (Chevallier 1987) describes the two types of diets. The first, called soupe, is more common and is given to dogs once a day (in the morning). It is in fact a meal of varied content, most often consisting of the leftover soup previously eaten by humans. The second type of food for dogs—especially conceived for working dogs, chien de berger (sheepdogs)—does not contain any animal content. Denis Chevallier further stated that the dog’s daily diet is mainly vegetarian and is only occasionally supplemented by animal castoffs, such as sheep foetuses, varieties of viscera, bones, etc. (Chevallier 1987).
In Can Roqueta, a high proportion of cereals were also observed in some puppies, leading to the hypothesis that in certain cases (absence of the mother, selection of specimens for work require differential feeding patterns), these animals were raised with a higher intake of carbohydrates. Only one puppy at Can Roqueta (CRII-591-2), who was between 2 and 3 months old, showed a high δ15N signature that can be attributed to the trophic effect of ingesting maternal milk (Fogel et al. 1989). The other two puppies (between 1 and 4 months of age) combine isotopic signatures characterised by the absence of maternal milk and a diet rich in cereal. It is noteworthy that ethnoarchaeological studies carried out in Tunisia (Alonso et al. 2014) indicate that women mix part of the cereal grinding by-products with water to feed puppies, as shown in Fig. 13.

The current ethnographic example of feeding puppies cereal by-products by the people of Ouarten tribe (El Kef, Tunisia): a Location of the Ouarten area, b open landscape and isolated houses with their pens and central courtyards in the area, c woman classifying products and by-products by sieving during the milling process of hulled barley with a rotary quern (at her left side), d puppies eat meal consisting of a ground cereal by-product mixed with water and e the grinding by-product placed into a hole in a courtyard. All images from (Alonso et al. 2014)
Diet of foxes
According to their isotopic signatures, the foxes of Minferri consumed various types of food. They cannot, in fact, be linked to any specific diet. One of these foxes, a female, exhibited signs of consumption of a more carnivorous diet. In all three cases, however, they cluster together with human females (with their different types of food) and dogs. On the contrary, the old fox at Can Roqueta revealed a diet with important input of vegetal protein, similar to that of the dogs and closer to two puppies and a young dog (see Fig. 10), albeit with more positive δ13C values. This deviation could be interpreted as resulting in a diet rich in marine molluscs (Chisholm et al. 2006). Yet this is unlikely, due to Can Roqueta’s relative distance from the sea. In addition, the fractures of this animal’s metatarsus and fibula were fused at a late period in its life, probably preventing it from travelling long distances for several weeks. This fracture must have caused a limp that could leave the animal more vulnerable and could mean that it would be hunted more easily. But its particular isotopic signature, which differs significantly from the Minferri foxes, raises the possibility that this fox could have been fed by humans for a period before its death at old age. Although, there is no archaeological evidence to back up this hypothesis, the old fox from Can Roqueta could have received food with cereals, including an appreciable input of the C4 group, such as foxtail millet (Setaria viridis/italica), since there are no wild plants in the archaeological record that can offer these results (Vila 2018). Yet, this interpretation must be taken with caution, as very little is presently known about this type of millet in the Northwest of the Iberian Peninsula and is only recorded at the cave of Punta Farisa in a Middle Bronze Age context (Alonso and Buxó 1993). Although, the presence of foxtail millet at Can Roqueta increases over time in structures of the Late Bronze Age to the Early Iron Age (Antolín, Mensua, Piqué, personal communication, unpublished 2008), evidence on the Early-Middle Bronze Age phases, the subject of the present study, are still lacking.
Conclusions
In this work, we present an isotopic analysis of canids and humans in two Bronze Age sites in the Northeast region of the Iberian Peninsula, Can Roqueta and Minferri, respectively, with the peculiarity that these animals were found deposited during human burials or in nearby ritual structures. This led us to contemplate their possible relationship in life with the humans, which could in fact transcend that of the realms of ceremony and symbolism. Furthermore, the isotopic study of the dogs and foxes also yields information about their diet and their role in the economic and social environment of each settlement.
The dogs at both sites presented a morphological homogeneity that may indicate a certain control over their reproduction. In addition, the inclusion of cereals in their diet, evidenced by their isotopic signatures, leads to the interpretation that animals were subject to human control and possibly some sort of selection based on specific characteristics. The inclusion of cereals in the diet of dogs cannot occur naturally. This indicates that humans may have deliberately prepared the food consumed by the animals at both sites, and that the food probably included the same plants, notably, cereals of group C3, as those consumed by humans. Proteins identified in the animal diet probably came from bones and leftovers of meat, muscles and tendons, or even from dairy products, but these could also be the result of the dogs’ behaviour as opportunistic feeders.
The humans probably fed the dogs to ensure that they fulfilled their assigned tasks, be it the task of guarding or protecting grazing herds. The relationship observed at both sites between the diet of dogs and that of female and young humans appears to suggest a greater dedication by this human group to the nurture and care of dogs.
However, there are differences that point to a diet richer in cereal, detectable in the most robust specimens that also show load pathologies. This coincidence opens up an interesting line of research into the possibility of a differential feeding of dogs intended for work in ancient societies. Isotopic data from some of the foxes indicate a similarity with the feeding patterns of the humans that they accompanied at death and with dogs, raising the possibility of a high level of interaction between these wild canids and societies of the past for reasons that still cannot be identified.
References
Agustí B, López JB, Alonso N (2005) Longevidad en un conjunto funerario prehistórico: Minferri (Juneda, les Garrigues, Catalunya). In: Cañellas A (ed) Nuevas perspectivas del diagnóstico diferencial en paleopatología, Actas del VII Congreso Nacional de Paleopatología (Maó-Menorca, 2-5 octubre 2003), Universitat de les Illes Balears, pp 472–478
Albizuri S (2011a) La ofrenda animal durante el Bronce Inicial en Can Roqueta II (Sabadell, Vallès Occidental). Universitat de Girona, Arqueozoología del ritual funerario. PhD dissertation
Albizuri S (2011b) Animales sacrificados para el cortejo fúnebre durante el Bronce Inicial (2300-1300 cal BC). El asentamiento de Can Roqueta II (Sabadell, Barcelona). Quaderns de Prehistòria i Arqueologia de Castelló 29:7–26
Albizuri S, Alonso N, López Cachero FJ (2011a) Economia i canvi social a Catalunya durant l'edat del bronze i la primer edad del ferro. In: Valenzuela-Lamas S, Padrós N, Belarte MC, Sanmartí J (eds) Economia agropecuària i canvi social a partir de les restes bioarqueològiques. El primer mil·lenni aC a la Mediterrània occidental, V Reunió Internacional d’Arqueologia de Calafell (Abril 2009). Arqueo Mediterrània 12:11–36
Albizuri S, Fernández M, Tomás X (2011b) Evidencias sobre el uso del perro en la carga durante el Bronce Inicial en la Península Ibérica: el caso de Can Roqueta II (Sabadell, Barcelona). Archaeofauna 20:139–155
Albizuri S, Maroto J, Nadal J, Majó T, Sánchez Marco A, Carlús X, Rodríguez A, Palomo A (2015) Wild carnivore and wild bird deposits in an agro-pastoral community during the Bronze Age: Can Roqueta II (Northeast Iberian Peninsula). Munibe Antropologia-Arkeologia 66:163–184
Alonso N, Buxó i Capdevila R (1993) Resultados iniciales del estudio Arqueobotánico de semillas y frutos del yacimiento de Cova Punta Farisa (Fraga, Huesca). In: El complejo arqueológico de Punta Farisa (Fraga, Huesca). Estudios de la Antigüedad 6-7:49–56
Alonso N (1999) De la llavor a la farina. Els processos agrícoles protohistòrics a la Catalunya Occidental. Monographies d’Archéologie Méditerranéenne 4. C.N.R.S. éditions, Lattes
Alonso N, López JB (2000) Minferri (Juneda, Garrigues): un nou tipus d’assentament a l’aire lliure a la plana occidental catalana durant la primera meitat del segon mil·lenni cal. BC Tribuna d’Arqueologia 1997-1998:279–306
Alonso N, Cantero FJ, Jornet R, López D, Montes E, Prats G, Valenzuela S (2014) Milling wheat and barley with rotary querns: the Berber Ouarten women (Dahmani, El kef, Tunisia). In: Selsing L (ed) Seen through a millstone. University of Stavanger, Bergen, Norway, Geology and archaeology of quarries and mills. Museum of Archaeology, pp 193–212
Alonso N, Junyent E, Lafuente A, López JB, Moya A, Tartera E, Vidal A (2006) Agricultura i poblament a la plana occidental catalana durant l’edat del bronze. Condicions de vida al món rural, Institut d’Estudis Ilerdencs, Lleida, pp 711–726
Alonso N, Pérez G, Rovira N, López D (2016) Gathering and consumption of wild fruits in the east of the Iberian Peninsula from the 3rd to the 1st millennium BC. Quat Int 404:69–85
Álvarez De Sotomayor JM (1824) Los Doce Libros de Agricultura que escribió en latín Lucio Junio Moderato Columela. Imprenta de Miguel de Burgos, Madrid
Al Qahtani SJ (2009) Atlas of tooth development and eruption. Barts and the London School of Medicine and Dentistry. Queen Mary University of London, London
Ambrose SH (1990) Preparation and characterization of bone and tooth collagen for isotopic analysis. J Archaeol Sci 17:431–451
Artigues PL, Bravo P, Hinojo E (2007) Excavacions arqueològiques a Can Gambús 2, Sabadell (Vallès Occ.). Tribuna d'Arqueologia 2006:111–140
Balsera R, Matas O, Roig J (2011) Els Pinetons, un assentament prehistòric i medieval a la plana del Vallès. Tribuna d’Arqueologia 2009:237–284
Bocherens H, Billiou D, Patou-Mathis M, Bonjean D, Otte M, Mariotti A (1997) Paleobiological implications of the isotopic signatures (13C, 15N) of fossil mammal collagen in Scladina cave (Sclayn, Belgium). Quat Res 48:370–380
Bocherens H, Drucker D (2003) Trophic level isotopic enrichments for carbon and nitrogen in collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13:46–53
Bocherens H, Drucker D, Germonpré M et al (2015) Reconstruction of the Gravettian food-web at Předmostí I using multi-isotopic tracking (13C, 15N, 34S) of bone collagen. Quat Int 359-360:211–228
Bolós O (1979) Geografia fisica dels Països Catalans. Els sols i la vegetació dels Països Catalans. In: Riba O et al (eds) Geografia fisica dels Països Catalans. Ketres, Barcelona, pp 107–158
Boquer S, González JL, Mercadal O, Rodón T, Saenz L (1990) Les estructures del bronze antic-mitjà al jaciment arqueològic de Can Roqueta (Sabadell, Vallès Occidental). Arrahona 7:9–25
Boquer S, González JL, Mercadal O, Rodón T, Saenz L (1992) Un nou assentament del bronze- ferro al Vallès: Can Roqueta. Tribuna d’Arqueologia 1990-1991:41–51
Bordas A, Pou R, Parpal A, Martín A (1994) Excavacions arqueològiques 1991-1992 a la Bòbila Madurell-Mas Duran (Sant Quirze del Vallès, Vallès Occidental). Tribuna d'Arqueologia 1992-1993:31–47
Bosch A, Mercadal O, Tarrús J (1989) La cova sepulcral del neolític antic de l’Avellaner (Les Planes d’Hostoles, la Garrotxa). Tribuna d’Arqueologia 1988-89:15–28
Bouso M, Esteve X, Farré J, Feliu JM, Mestres J, Palomo A, Rodríguez A, Senabre MR (2004) Anàlisi comparatiu de dos assentaments del bronze inicial a la depressió prelitoral: Can Roqueta II (Sabadell-Valles Occiental) i Mas d'en Boixos- 1 (Pacs del Penedès-Alt Penedès). Cypsela 15:73–101
Bouso M, Esteve X, Farré J, Feliu JM, Mestres J, Palomo A, Rodríguez A, Senabre MR (2005) Can Roqueta II y Mas d’en Boixos-1, dos yacimientos del Bronce inicial situados en la depresión prelitoral catalana. In: Bicho N, Corchón MS (Coord) As idades do Bronce e do Ferro na Península Ibérica. Acta do IV Congreso de Arqueologia. Peninsular. 14-19 de Setembro, Faro, Universidade do Algarve, Promontoria Monográfica, pp 145–158
Brea MB, Mazzieri P, Micheli R (2010) People, dogs and wild game: evidence of human-animal relations from Middle Neolithic burials and personal ornaments in northern Italy. Documenta Praehistorica XXXVII:125–145
Brück J (1999) Ritual and rationality: some problems of interpretation in European archaeology. Eur J Archaeol 2(3):313–314
Bruzek J (2002) A method for visual determination of sex, using the human hip bone. Am J Phys Anthropol 117(2):157–168
Buikstra, JI, Ubelaker DH (1994) Standards for data collection from human skeletal remains: Proceedings of a seminar at the field Museum of Natural History, Organized by Jonathan Haas, Arkansas Archeol Surv Res Series 44
Cámara Serrano JA, Sánchez Susí R, Riquelme JA et al (2016) Culte aux ancêtres dans la période chalcolithique de la péninsule ibérique ? Le sacrifice d’animaux, la circulation des restes humains et la différence de traitement entre hommes et femmes dans les tombes du site archéologique à Marroquíes (Jaén, Espagne) trouvées dans les fouilles de la Tranche 3 du système du tramway. L’Anthropologie 120(2):145–174
Cannon A, Schwarcz HP, Knyf M (1999) Marine based subsistence trends and the stable isotope analysis of dog bones from Namu, British Columbia. J Archaeol Sci 26:399–407
Carlús X, Lara C, López Cachero X, Oliva M, Palomo A, Rodríguez A, Terrats N, Villena N (2002) El paraje arqueológico de Can Roqueta (Sabadell, Vallès Occidental): diacronia y tipología de las ocupaciones. XXVII Congreso Nacional de Arqueología (Huesca, 6 al 8 de Mayo del 2003). Bolskan 19:121–139
Carlús X, López Cachero X, Oliva M, Palomo A, Rodríguez A, Terrats N, Lara C, Villena N (2007) Cabanes, sitges i tombes. El paratge de Can Roqueta (Sabadell, Vallès Occidental) del 1300 al 500 aC. Quaderns d’Arqueologia 4, Museu d’Història de Sabadell. Sabadell
Carlús X, López Cachero FJ, Terrats N, Oliva M, Palomo A, Rodríguez A (2008) Diacronia durant la prehistòria recent a Can Roqueta (Sabadell-Barberà del Vallès, Vallès Occidental) entre el VI i el I mil·lenni cal ANE. Cypsela 17:115–142
Catagnano V (2016) Aproximación morfométrica y paleogenética al estudio de la variabilidad de Canis l. familiaris en la Península Ibérica desde el neolítico hasta época romana y su contextualización en el ámbito del mediterráneo occidental. PhD dissertation, Universitat Autònoma de Barcelona
Chevallier D (1987) L'Homme, le porc, l'abeille et le chien. La relation homme-animal dans le Haut-Diois. Muséum National d'Histoire Naturelle. Mémoires de l'Institut d'Ethnologie XXVI, Paris
Chisholm BS, Nelson DE, Schwarcz HP (2006) Stable carbon isotope ratios as a measure of marine versus terrestrial protein in ancient diets. Science 216:1131–1132
Conlin E (2003) Los inicios del III milenio a.C. en Carmona. Las evidencias arqueológicas. Carel 1:83–143
Coppinger R, Schneider R (1995) Evolution of working dogs. In: Serpell K (ed) The domestic dog: its evolution, behaviour, and interactions with people. Cambridge University Press, Cambridge, pp 21–50
Daza A (2011) Los depósitos de perros. In Blasco C. Liesau C, Ríos P (eds) Yacimientos calcolíticos con campaniforme de la región de Madrid, Patrimonio Arqueológico 6, Madrid, pp 211–222
DeNiro MJ (1985) Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317:806–809
Driesch A von den (1976) a guide to the measurement of animal bones from archaeological sites. Peabody museum bulletin, 1, Harvard
Equip Minferri (1997) Noves dades per a la caracterització dels assentaments a l’aire lliure durant la 1a meitat del II mil·leni cal. BC: primers resultats de les excavacions en el jaciment de Minferri (Juneda, Garrigues). Revista d’Arqueologia de Ponent 7:161–211
Ewersen, J, Ziegler S, Ramminger B, Schmölcke U (2018) Stable isotopic ratios from Mesolithic and Neolithic canids as an indicator of human economic and ritual activity. Journal of Archaeological Science: Reports 17:346–¬357
Fazekas IG, Kosa F (1978) Forensic foetal osteology. Akadémiai Kiado, Budapest
Ferembach D, Schwidetzky I, Stloukal M (1979) Recommandations pour determiner l’âge et le sexe sur le squelette. Bull. et Mém. de la Soc. d’Anthrop. de Paris 6, XIII:7–45
Ferrio JP, Alonso N, López JB, Araus JL, Voltas J (2006) Carbon isotope composition of fossil charcoal reveals aridity changes in the NW Mediterranean Basin. Glob Chang Biol 12:1–14
Fischer A, Olsen J, Richards M, Heinemeier SÁE, Bennike P (2007) Coast-inland mobility and diet in the Danish Mesolithic and Neolithic: evidence from stable isotope values of humans and dogs. J Archaeol Sci 34:2125–2150
Fogel M, Tuross N, Owsley D (1989) Nitrogen isotope tracers of human lactation in modern and archaeological populations. Carnegie Institution of Washington Yearbook, pp:111–117
Forcadell T, Villalbí M (1999) Cova Cervereta (Vinallop, Tortosa). Cavitat sepulcral del calcolític-bronze antic al curs inferior de l'Ebre Quaderns de Prehistòria i Arqueologia de Castelló 20:37–54
Fuller B, Fuller J, Harris D, Hedges R (2006) Detection of breastfeeding and weaning in modern human infants with carbon and nitrogen stable isotope ratios. Am J Phys Anthropol 129:279–293
Garvin HM, Passalacqua NV, Uhl NM, Gipson DR, Overbury RS, Cabo LL (2012) Development in forensic anthropology. Age at death estimation. In: Dirkmaat DC (ed) A companion to forensic anthropology. Blackwell Publishing Ltd, First Ed.
Germonpré M, Sablin MV, Stevens RE, Hedges REM, Hofreiter M, Stiller M, Després VR (2009) Fossil dogs and wolves from Palaeolithic sites in Belgium, the Ukraine and Russia: osteometry, ancient DNA and stable isotopes. J Archaeol Sci 36:473–490
Gómez X (2000a) Noves dades sobre l’explotació de base animal durant la protohistória a la plana occidental catalana. Universitat de Lleida, Tesina de llicenciatura
Gómez X (2000b) Homes i animals al jaciment protohistóric de Minferri (Juneda, Catalunya). II Trobada d’Estudiosos de la Comarca de les Garrigues, Tarrés, pp 11–26
Gómez X (2003) El gos a Minferri (Juneda-Garrigues): Prehistòria del millor amic de l’home. Actes III Trobada d’Estudiosos de la Comarca les Garrigues, Cervià de les Garrigues, pp 219–232
Goude G, Fontugne M (2016) Carbon and nitrogen isotopic variability in bone collagen during the Neolithic period: influence of environmental factors and diet. J Archaeol Sci 70(2016):117–131
Guerrero L (1990) L’antropologia física. In: Llussà A et al. El jaciment del Bronze de Minferri (Juneda, les Garrigues), Quaderns d’Arqueologia del Grup de Recerques Arqueològiques de “La Femosa” 5, Artesa de Lleida, pp 45–54
Guagnin M, Perri A, Petraglia MD (2018) Pre-Neolithic evidence for dog-assisted hunting strategies in Arabia. J Anthropol Archaeol 49:225–236
Guiry EJ (2012) Dog as analogs in human stable isotope based paleodietary reconstructions: a review and consideration for future use. J Archaeol Method Theory 19(3):351–376
Guiry EJ, Grimes V (2013) Domestic dog (Canis familiaris) diets among coastal late archaic groups of northeastern North America: a case study for the canine surrogacy approach. J Anthropol Archaeol 32(4):732–745
Hammer Ø, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeontol Electron 4(1)
Harcourt RA (1974) The dog in prehistoric and early historic Britain. J Archaeol Sci 1:151–175
Heaton THE (1987) The 15N/14N ratios of plants in South Africa and Namibia: relationship to climate and coastal/saline environments. Oecologia 74(2):236–246
Horard-Herbin MP (2000) Dog management and use in the late Iron Age. The evidence from the Gallic site of Levroux (France). In: Crockford SJ (ed) dogs through time: an archaeological perspective, BAR Int. Ser., 889, Archaeopress, Oxford, pp. 115–121
Horard-Herbin MP, Tresset A, Vigne JD (2014) Domestication and uses of the dog in Western Europe from the Paleolithic to the Iron Age. Animal Frontiers 4(3):23–31
Horwitz LK, Goring-Morris L (2004) Animals and ritual during the Levantine PPNB: a case study from the site of Kfar Hahoresh, Israel. Anthropozoologica 39(1):165–178
Isik G (2009) Yal. The traditional dog food in Anatolia. Choban chatter, the official publication of Anatolian shepherd dogs international 19(1)
Kosa F (1989) Age estimation from the foetal skeleton. In: Iscan MY (ed) Age markers in the human skeleton. Charles C. Thomas Publ, Springfield, Illinois, pp 21–54
Koster JM, Tankersley KB (2012) Heterogeneity of hunting ability and nutritional status among domestic dogs in lowland Nicaragua. PNAS 109(8):E463–E470. https://doi.org/10.1073/pnas.1112515109
Kuehn SR (2014) Prehistoric dog pathology in the American bottom: evidence from the Janey B. Goode site (11S1232), St. Clair County, Illinois. Illinois Archaeol 26:97–129
Kuhnle GGC, Joosen AMCP, Catherine J, Kneale CJ, O’Connell TC (2013) Carbon and nitrogen isotopic ratios of urine and faeces as novel nutritional biomarkers of meat and fish intake. Eur J Nutr 52:389–395. https://doi.org/10.1007/s00394-012-0328-2
Latham KJ (2016) Working like dogs: a systematic evaluation of spinal pathologies as indicators of dog transport in the archaeological record. University of Alberta, PhD dissertation
Lawler D, Widga FC, Rubin DA, Reetz JA, Evans RH, Tangredi BP, Thomas RM, Martin TJ, Hildebolt C, Smith K, Leib D, Sackman JE, Avery JG, Smith GK (2016) Differential diagnosis of vertebral spinous process deviations in archaeological and modern domestic dogs. J Archaeol Sci 9:54–63. https://doi.org/10.1016/j.jasrep.2016.06.042
Liesau C, Daza A, Llorente L, Morales A (2013) More questions than answers: the singular animal deposits from Camino de Las Yeseras (chalcolithic, Madrid, Spain). Anthropozoologica 48(2):277–286. https://doi.org/10.5252/az2013n2a6
López JB (2000) L’evolució del poblament protohistòric a la plana occidental catalana: models d’ocupació del territori i urbanisme. Universitat de Lleida, PhD dissertation
López JB (2001) Minferri en el context de l’edat del bronze a la plana occidental catalana, a Grup d’Investigació Prehistòrica. In : Colors de terra. La vida i la mort en una aldea d’ara fa 4.000 anys. Minferri (Juneda), Quaderns de la Sala d’Arqueologia, 1, Institut d’Estudis Ilerdencs, Lleida, pp 13–40
Losey RJ, Bazaliiskii VI, Garvie-Lok S, Germonpré M, Leonard JA, Allen AL, Katzenberg AM, Sablin MV (2011) Canids as persons: early Neolithic dog and wolf burials, Cis-Baikal, Siberia. J Anthropol Archaeol 30(2):174–189
Losey RJ, Garvie-Lok S, Leonard JA et al (2013) Burying dogs in ancient cis-Baikal, Siberia: temporal trends and relationships with human diet and subsistence practices. Plos One 17 8(5):e63740. https://doi.org/10.1371/journal.pone.0063740
Maher L, Stock J, Finney S, Heywood J, Miracle P, Banning E (2011) A unique human-fox burial from a pre-Natufian cemetery in the Levant (Jordan). PLoS One 6(1):e15815. https://doi.org/10.1371/journal.pone.0015815
Mann HB, Whitney DR (1947) On a test of whether one of two random variables is stochastically larger than the other. Ann Math Stat 18(1):50–60
Marín D (2018) Origen, tecnología y funcionalidad del utillaje lítico tallado durante la Edad del Bronce en el Nordeste de la Península Ibérica (2000–1300 cal ANE). Universitat de Lleida, PhD dissertation
Marín D, Gibaja JF, Alonso Martínez A, Ortega i Cobos D, Palomo A, Moya Garra A (2017) Chipped stone tools from the Early Bronze Age settlement of Minferri (2100-1650 cal. BC) (Lleida, Spain). Raw materials, technology and activities inferred. In: Brysbaert A, Gorgues A (eds) artisans versus nobility. Multiple identities of elites and “commoners” viewed through the lens of crafting from the chalcolithic to the Iron ages in Europe and the Mediterranean. Sidestone press, Leiden, pp 139–159
Martí M, Pou R, Carlús X (1997) Excavacions arqueològiques a la Ronda Sud de Granollers, 1994. La necròpolis del Neolític Mitjà i les restes romanes del Camí de Can Grau (La Roca del Vallès, Vallès Oriental) i els jaciments de Cal Jardiner (Granollers, Vallès Oriental). Excavacions Arqueològiques a Catalunya 14. Barcelona
Martín Cólliga A, Martín J, Villalba P, Juan-Tresserras J (2005) Ca l’Oliaire (Berga, Barcelona), un asentamiento neolítico en el umbral del IV milenio con residuos de sal y de productos lácteos. In: III Congreso del Neolítico de la Península Ibérica, pp. 175-185. Universidad de Cantabria. Santander
Martín Colliga A, Blanch RM, Albizuri S, Alaminos A, Mercadal O, Vives E, Lázaro P, Bosch J, Colomer S, Miret J, Enrich R, Aliaga S (2017) El paraje de Bòbila Madurell (Sant Quirze del Vallès, Vallès Occidental, Barcelona). In: Gibaja JF, Subirà ME, Martín A, Mozota M, Roig J (eds) Mirando a la Muerte: Las prácticas funerarias durante el neolítico en el noreste peninsular. E-ditArx - Publicaciones Digitales
Morey DF (2010) Dogs. Cambridge University Press, Domestication and the developement of a social bond
Morris J (2011) Investigating animal burials; ritual. Mundane and Beyond BAR British Series 535
Moorrees CFA, Fanning EA, Hunt EE Jr (1963a) Formation and resorption of three deciduous teeth in children. Am J Phys Anthropol 21:205–213
Moorrees CFA, Fanning EA, Edward E, Hunt EE Jr (1963b) Age variation of formation stages for ten permanent teeth. J Dent Res 42(6):1490–1502
Moya A, López JB, Bergadà MM, Alonso N, Escala Ò, Mateu M, Tartera E, Vidal A (in press) Hornos en fosa para la cocción cerámica en poblados del Neolítico final de la Catalunya occidental: Minferri y Cantorella. VI Congreso del Neolítico en la Península Ibérica (Granada, 22-26 junio 2016)
Nieto A, Moya A, López JB, Agustí B (2014) Ofrenes o deixalles ? El cas dels bovins (Bos taurus) en context funerari del jaciment del bronze ple de Minferri (Lleida, Catalunya). Équidés de la Méditerranée antique. Rites et combats. Jeux et savoirs, Monographies d'Archéologie Méditerranéenne, Hors-série 6:53–112
Noe-Nygaard N (1988) 13C-values of dog bones reveal the nature of changes in man's food resources at the Mesolithic-Neolithic transition, Denmark. Chem Geol: Isot Geosci sect 73(1):87–96
O’Connell TC, Kneale CJ, Tasevska N, Kuhnle GGC (2012) The diet-body offset in human nitrogen isotopic values: a controlled dietary study. Am J Phys Anthropol 149:426–434
Ollivier M, Tresset A, Bastian F, Lagoutte L, Axelsson E, Arendt ML, Bălăşescu A, Marshour M, Sablin MV, Salanova L, Vigne JD, Hitte C, Hänni C (2016) Amy2B copy number variation reveals starch diet adaptations in ancient European dogs. Royal Soc Open Sci 9 3(11):160449. https://doi.org/10.1098/rsos.160449 eCollection 2016 Nov
Onar V (2005) Estimating the body weight of dogs unearthed from the Van-Yoncatepe necropolis in eastern Anatolia. Turk J Vet Anim Sci 29:495–498
Onar V, Belli O (2005) Estimation of shoulder height from long bone measurements on dogs unearthed from the Van-Yoncatepe early Iron age necropolis in eastern Anatolia. Rev Med Vet 156(1):53–60
Onar V, Belli O, Owen PR (2005) Morphometric examination of red fox (Vulpes vulpes) from the Van Yoncatepe necropolis in eastern Anatolia. Int J Morphol 23(3):253–260
Ovodov ND, Crockford SJ, Kuzmin YV, Higham TFG, Hodgins GWL, van der Plicht J (2011) A 33,000-year-old incipient dog from the Altai Mountains of Siberia: evidence of the earliest domestication disrupted by the last glacial maximum. PLoS One 6(7):e22821. https://doi.org/10.1371/journal.pone.0022821
Palomo A, Rodríguez A (2002) Can Roqueta II (Sabadell, Vallès Occidental): un jaciment excepcional de l'edat del bronze. Pirineus i veïns al 3r mil·lenni aC. De la fin del neolític a l'edat del bronze entre l'Ebre i la Garona. XI Col·loqui Internacional d’Arqueologia de Puigcerdà, Puigcerdà
Palomo A, Terrats N, Oliva M, Rodríguez A, Majó T (2016) El complex arqueològic de Can Roqueta: un poblat paradigmàtic del bronze inicial a la Depressió Prelitoral Catalana. Arraona 36:130–141
Pearson JA, Bogaard A, Charles M, Hillson SW, Larsen CS, Russell N, Twiss K (2015) Stable carbon and nitrogen isotope analysis at Neolithic Çatalhöyük: evidence for human and animal diet and their relationship to households. J Archaeol Sci 57:69–79
Pechenkina EA, Ambrose SH, Xiaolin MA, Benfer RA Jr (2005) Reconstructing northern Chinese Neolithic subsistence practices by isotopic analysis. J Archaeol Sci 32(8):1176–1189
Pereira J, García E (2002) Bellotas, el alimento de la edad de oro. Arqueoweb 4(2):1–31
Peters J, Schmidt K (2004) Animals in the symbolic world of PPN Göbekli Tepe (Turkey). Antropozoologica 39:179–218
Peterson N (2006) Hundar kring benen. En analys av belastning och storleksvariation hos hundarna I det Mesolitiska Skane. PhD dissertation, Lunds universitet, Lund, Swedish
Perri A (2017) A typology of dog deposition in archaeological contexts. In Rowley-Conwy P Halstead P Serjeantson D (eds) economic Zooarchaeology: studies in hunting, herding and early agriculture. 1st ed., oxbow books, Ch.11, JSTOR, Oxford; Philadelphia www.jstor.org/stable/j.ctt1s4751q.16
Piérard J (1967) Note d’anatomie appliquée, appréciation de l’âge du chien. Can Vet J 8(9):197–200
Pitulko V, Kasparov AK (2017) Archaeological dogs from the Early Holocene Zhokhov site in the eastern Siberian Arctic. J Archaeol Sci Rep 13:491–515
Prats G (2013) Aproximació tipològica i funcional de les estructures excavades al jaciment del bronze ple de Minferri (Juneda, les Garrigues): emmagatzematge i conservació a la Catalunya occidental. Revista d’Arqueologia de Ponent 23:89–126
Prats G (2017) L’emmagatzematge en sitja entre el Neolític i l’època ibèrica (del 5500 ANE al I ANE). Aproximació socioeconòmica a les comunitats del nord-est de la Península Ibèrica. Universitat de Lleida, PhD Dissertation
Rodríguez A, Palomo T, Majó T (2002) Les estructures funeràries de Can Roqueta II (Sabadell, Vallès Occidental). XII Col·loqui Internacional d’Arqueologia de Puigcerdà, Puigcerdà, pp 659-669
Roig J, Coll JM (2016) El poblament prehistòric del paratge de Can Gambús-1: l’evolució dels assentaments agrícoles des del neolític final fins a l’edat del ferro (3000-500 cal. aC). Arraona 36:90–97
Rosillo R, Palomo A, Tarrús J et al (2012) Darreres troballes de prehistòria a l'Alt Empordà. Dos assentaments a l'aire lliure: La Serra de Mas Bonet (Vilafant) i Els Banys de la Mercè (Capmany). Tribuna d'Arqueologia 2010-2011:41–62
Ruiz C, Vázquez JM, Lomba J, Aviles A, Haber M, Drenes M, Gil Cano F (2014) El yacimiento calcolítico “Camino del Molino” (Caravaca de la Cruz, Murcia): una oportunidad excepcional para estudiarlos cánidos que poblaron el sureste español hace 4000 años. Primeros resultados Orígenes y Raíces 6:5–7
Russell N (2012) Hunting sacrifice at Neolithic Çatalhöyük. In: Porter AM, Schwartz GM (eds) Sacred killing: the archaeology of sacrifice in the ancient near east. Winona Lake, Indiana, Eisenbrauns, pp 79–95
Scheuer L, Black S (2000) Developmental juvenile osteology. Elsevier, London
Schwarcz HP, Schoeninger MJ (1991) Stable isotope analyses in human nutritional ecology. Am J Phys Anthropol 34(S13):283–321
Schwarcz HP, Schoeninger MJ (2012) Stable isotopes of carbon and nitrogen as tracers for palaeo-diet reconstruction. In: Baskaran M (ed) Handbook of environmental isotope geochemistry. Springer-Verlag, Berlin, pp 725–742
Szpak P, White CD, Longstaffe FJ, Millaire J-F, Vásquez Sánchez VF (2013) Carbon and nitrogen isotopic survey of northern peruvian plants: baselines for paleodietary and paleoecological studies. PLoS One 8. https://doi.org/10.1371/journal.pone.0053763
Tankersley KB, Koster JM (2009) Sources of stable isotope variation in archaeological dog remains. North Am Archaeol 30(4):361–375
Valera AC, Costa C (2013) Animal limbs in funerary contexts in southern Portugal and the question of segmentation. Anthropozoologica 48(2):263–275
Valera AC, Nunes T, Costa C (2010) Enterramentos de canídeos no Neolítico: a fossa 5 de Corça 1 (Brinches, Serpa). Apontamentos de Arqueologia e Património 5:7–17
Vila S (2018) L'explotació dels recursos vegetals a la plana occidental catalana durant la protohistòria a partir de l'anàlisi antracològica (III-I mil·lenni). Universitat de Lleida, PhD dissertation
Villalba MJ (1999) Las sepulturas neolíticas del complejo minero de Can Tintorer y el modelo social de la población minera. Revista d’arqueologia de Ponent 9:41–73
Warren DM (2004) Skeletal biology and paleopathology of domestic dogs from prehistoric Alabama, Illinois. Indiana University, Bloomington, Kentucky and Tennessee. PhD dissertation
Ward JH (1963) Hierarchical grouping to optimize an objective function. J Am Stat Assoc 58:236–244
Wilson GL (1924) The horse and the dog in Hidatsa culture. Anthropological Papers of the American Museum of Natural History XV, II, American Museum of Natural History, New York, pp 125–311
Acknowledgements
Our deepest gratitude goes to Mònica Oliva and Noemí Terrats, the directors of the excavation of Can Roqueta, for giving access to unpublished data on the structures of the sectors of Torre Romeu and Can Revella and Javier López Cachero of the University of Barcelona for the recent dating of CRII-591 of Can Roqueta in the framework of projects HAR2013-48010-P and HAR2017-87695-P (MINECO, Spain). We also thank Julià Maroto of the Universitat de Girona for his collaboration in the analysis of animals of Can Roqueta in the framework of project HAR2010-22013 (MINECO, Spain). The research on Minferri was supported by projects HAR2016-78277-R (MINECO, Spain) and SGR 2017-1714 (Generalitat de Catalunya, Catalonia). AGD receives support from the project CGL2014-57209-P (MINECO, Spain). We also acknowledge the veterinary diagnosis of osseous spinal and trauma pathologies to Matias Fernández (608 Farm and Equine Veterinary Surgeons, Quarry Farm, Rowington Green, Rowington, Warwick, Warwickshire, UK) and María Martín Cuervo (Dep. de Medicina Animal, Facultad de Veterinaria, Universidad de Extremadura, Spain). We also acknowledge Timothy Anderson for the English translation.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Grandal-d’Anglade, A., Albizuri, S., Nieto, A. et al. Dogs and foxes in Early-Middle Bronze Age funerary structures in the northeast of the Iberian Peninsula: human control of canid diet at the sites of Can Roqueta (Barcelona) and Minferri (Lleida). Archaeol Anthropol Sci 11, 3949–3978 (2019). https://doi.org/10.1007/s12520-019-00781-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-019-00781-z