
Abstract
Scavenger birds can feed on large- to small-sized vertebrates and may contribute in the formation of archaeological sites. To evaluate the modifications and dispersal patterns of bones produced by New World vulture and caracara from Northwestern Patagonia, samples of adult sheep, young sheep, and hare carcasses were offered and subsequently analyzed. New World vultures and caracaras quickly fed on the samples. Taphonomic and bone dispersal patterns suggest two types of accumulations: (1) open-air sites with large and medium-sized vertebrates represented by complete, fractured, scratched, notched, and punctured elements and (2) rock shelter or cave sites dominated by small vertebrates represented by broken, corroded, fractured, and digested elements.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Archaeological sites frequently contain animal bone remains representing past human activity and interaction. For example, remains may reflect pack animals, assistance with prey capture, large predator defense, and most commonly food sources (Lyman 1994). However, faunal accumulations from archaeological sites are not always deposited by humans as at times, sites may not be occupied by humans and may be subject to animal burrowing or nesting (Andrews 1990; Mondini 1995, 2004). Discriminating the accumulating agent is necessary to interpret the relationship between humans and fauna. Studies are vital because they allow for understanding the taphonomic processes and histories of osseous remains from the death of the animal until their recovery.
Researchers examining the disarticulation process of skeletal remains of different species in natural environments report the effects of transportation by different agents, weathering, corrosion by acidic soils, trampling, and modification (e.g., Cáceres et al. 2009; Yravedra et al. 2012). Studies of dispersal and disarticulation of carcasses and bone associations preserved at the site mostly focus on carnivore feeding, gnawing, and scavenging, rather than on the impact of birds (e.g., Binford 1981; Haynes 1982; Borrero et al. 2005; Álvarez et al. 2012).
Scavengers and predators can disperse parts of a carcass over a long distance from the place of death (Hill 1979; Mondini 1995; Martin and Borrero 1997). They reduce bone concentrations and generate secondary concentrations composed by remains from different individuals (Hill 1979). Dispersal patterns can reflect different taphonomic histories. For example, disarticulation can expose the terminal portion of bones to weathering and reduce the skeleton into a large number of parts, which are more easily transported and dispersed (Hill 1979). Also, the factors influencing bone accumulations can affect the relative proportions of the different elements from archaeological sites (Hill 1979). Carnivores like the puma (Puma concolor) can transport the bone segments of its prey from kill sites to rock shelters, showing distinctive digestive corrosion (Martin and Borrero 1997). Numerous Old World vulture examples exist for differential bone accumulation from bird scavenging and carcass transport (Robert and Vigne 2002a, 2002b; Benson et al. 2004; Marín-Arroyo et al. 2009; Robert and Reumer 2009; Marín-Arroyo and Margalida 2012; Lloveras et al. 2014; Sanchis Serra et al. 2014). However, there are few studies examining transport of bones consumed by New World vulture (e.g., Reeves 2009; Spradley et al. 2012; Dadds and Martin 2013).
Scavenger birds mostly feed on large ungulates but can include smaller vertebrates in their diets (del Hoyo et al. 1994; DeVault et al. 2004; Biondi et al. 2005; Kelly et al. 2007; Vargas et al. 2007; Lambertucci et al. 2009; Ballejo and De Santis 2013). These birds can transport and feed on carcass parts at nesting or roosting places on ledges of rocky cliffs and in trees, where bone accumulations can mix with carnivore or human-derived assemblages (Ballejo et al. 2012; Marín-Arroyo and Margalida 2012). Thus, identification of the accumulating agents of faunal assemblages can avoid incorrect paleoeconomic interpretations (Marín-Arroyo and Margalida 2012).
The aim of this study was to investigate how the scavenging of New World vultures (Cathartidae) and caracaras (Falconidae) affects the deposition and recovery of sheep (Ovis aries) and European hare (Lepus europaeus) in Northwestern Patagonia. To develop an actualistic model, we examined taphonomic variables potentially affecting the zooarchaeological record and evaluated the spatial dispersal of adult sheep, young sheep, and hare remains. Additionally, to establish the different anatomical pattern and types of accumulations of these scavenger birds, we compared our results to Ballejo et al. (2012), which examined American black vulture (Coragyps atratus) pellet samples in the same area.
Materials and methods
Study area
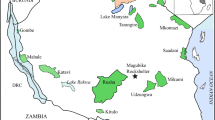
The study area (Fig. 1) includes high elevations of the North Patagonian Andes (2000–2300 masl) and low mountains (1300–1800 masl) of the Patagonian Pre-cordillera (Giacosa and Marquez 1999; Giacosa et al. 2005). The climate is cold with strong westerly winds and a significant west-east rainfall gradient due to the Pacific anticyclone and the topographic barrier of the Andean Cordillera, which generates drier conditions to the east (Paruelo et al. 1998). Phytogeographically, the area belongs to the Occidental District of the Patagonian unit, which is characterized by shrubby and grassy steppe with a vegetation cover about 50 %, constituted mainly by Stipa speciosa (“coirón amargo”), Stipa humilis (“coirón llama”), Senecio filaginoides (“cata mora”), and Mulinum spinosum (“neneo”) (León et al. 1998).

Map of the study area, including the location of carcass samples analyzed in this study and pellet samples reported in Ballejo et al. (2012): 1 Dina Huapi; 2 Estancia El Cóndor; 3 Estancia Chacabuco
This area is also characterized by the presence of several animal species whose importance for the faunal economy is well documented: lesser rhea (Rhea pennata), guanaco (Lama guanicoe), pichi (Zaedyus pichiy), large hairy armadillo ( Chaetophractus villosus ), Patagonian weasel (Lyncodon patagonicus), Molina’s hog-nosed skunk (Conepatus chinga), South American grey fox (Lycalopex griseus), culpeo (L. culpaeus), Geoffroy’s cat (Leopardus geoffroyi), and puma (e.g., Ringuelet 1961). Noteworthy is the existence of a high abundance of domestic livestock mammals: cow (Bos taurus), horse (Equus caballus), sheep, and exotic wild mammals: European hare, red deer (Cervus elaphus), and wild boar (Sus scrofa) hunted for sport (Novillo and Ojeda 2008; Lambertucci et al. 2009).
Carcass samples analyzed in this study were observed in Estancia El Cóndor, Río Negro Province (44° 11′ 55.7″ S, 71° 05′ 43.1″ W, Fig. 1), which produces livestock, mostly sheep (ca. 30,000 individuals), followed by cows, horses, and red deer. Animals that die from sickness or old age are consumed by the substantial populations of scavenger birds nesting in the Estancia El Cóndor and its surroundings. These scavenger birds possess the ability to find carcasses and avoid predators frequenting the area (Lambertucci et al. 2009; Ballejo et al. 2012; Lambertucci et al. 2014).
New World vulture and caracara natural history
Although five species of Cathartidae occur in Argentina, only three (Vultur gryphus, Coragyps atratus, and Cathartes aura) are found in Northwestern Patagonia (del Hoyo et al. 1994). Cathartidae are strictly scavengers that feed on carrion (Wallace and Temple 1987; DeVault et al. 2003). Feeding adaptations include hooked bills, acute vision, the ability to search large areas using energy-efficient soaring flight, and an acute sense of smell (Buckley 1996).
The Andean condor (V. gryphus) is the largest of the South American Cathartidae (100–130 cm; 8.2–10.5 kg) with a 3-m wingspan. It is distributed from the Andes Mountains of Venezuela to southernmost Argentina and Chile. Roosting and nesting occur in mountainous areas (e.g., cliffs, rock shelters), and foraging occurs in open area savannas, grasslands, and beaches along the coast. The condors forage alone or in well-dispersed groups at high altitudes. They scavenge large carcasses of marine mammals and both wild and domestic ungulates (del Hoyo et al. 1994; DeVault et al. 2004; Grzimek’s 2002; Lambertucci et al. 2009).
The American black vulture (C. atratus) is a large-sized bird (56–69 cm and 1.1–1.9 kg). It inhabits open areas and water ways from southern North America to southern South America (South of Chubut, Argentina). The vultures roost, forage, and feed socially in large groups on cliffs, rock shelters, or trees. It feeds on dead animals of any size and occasionally kills injured animals (del Hoyo et al. 1994; Grzimek’s 2002; Ballejo and De Santis 2013).
The Turkey vulture (C. aura) is a large-sized bird (64–81 cm and 0.85–2 kg) that lives in a different kind of habitat from the southern border of Canada to Tierra del Fuego (Argentina). This vulture roosts socially on cliffs, rock shelters, and trees and hunts for carrion in small groups or singly using olfaction. Its diet includes dead animals of any size; however, the species specializes on smaller taxa (del Hoyo et al. 1994; Grzimek’s 2002; Kelly et al. 2007).
In addition to Northwestern Patagonian Cathartidae, two species of Falconidae (Caracara plancus and Milvago chimango) occur as hunters and scavengers (i.e., facultative scavengers) (Wallace and Temple 1987).
The southern caracara (C. plancus) is a large-sized bird (49–59 cm and 0.83 kg) that lives typically in open and semi-open lowlands, grasslands, pastures, and river edges and in Patagonian river valleys, shrub steppes, and grassy foothills. This species is gregarious when scavenging carcasses. Out of the breeding season, they roost communally at the top of isolated trees. They typically feed on carrion, road kills, dead and dying fish, and livestock (Grzimek’s 2002; Vargas et al. 2007).
The chimango caracara (M. chimango) is a medium-sized bird (37–43 cm and 0.25–0.30 kg) that occurs in several kinds of habitats. This includes rural and urban areas from Paraguay to the southern extreme of Brazil to southernmost Chile and Argentina (del Hoyo et al. 1994). This falconid roosts on cliffs, rock shelters, and trees and is a diurnal and generalistic raptor (e.g., Yáñez et al. 1982; Biondi et al. 2005).
Observation and analysis of scavenged remains
We observed and recorded vulture and caracara scavenging and its taphonomic effects on the remains of adult sheep (five in summer, three in autumn, and two in spring of 2013), young sheep (two in spring of 2014), and European hare (one in spring of 2014). Bird scavenging was observed from ca. 150–300 m distance with the naked eye and through binoculars.
Both perimortem and postmortem carcasses not previously fed on by predators or scavengers were observed to determine: (1) vulture and caracara consumption time, (2) number of scavenging birds by species, and (3) skeletal remains dispersed from carcasses. Perimortem carcass consisted of a single adult sheep and hare, which were offered by us to scavenging birds, allowed for observation and quantification of remains immediately after death. For comparison, we also observed postmortem carcasses of adults sheep (n = 9) and young sheep (n = 2) that had naturally died. Carcasses were observed until all scavenger birds ceased feeding, and consumption time was recorded. After each scavenging episode, the compass orientation and dispersal distance of scavenged skeletal parts was measured from point of placement (perimortem) or carcass location (postmortem) to final resting site (Yravedra et al. 2012).
Quantification and statistical analysis
From collected remains, we calculated the number of identified specimens (NISP), minimum number of elements (MNE), minimum number of anatomical units (MAU), standardized MAU (MAU%), and minimum number of individuals (MNI) (Grayson 1984). The relative abundance of skeletal elements was calculated for final accumulations as:
\( MNI={MNE}_{\mathrm{i}}/\left({\mathrm{E}}_{\mathrm{i}}\times MNI\right)\times 100 \)
where MNEi is the minimum number of a given skeletal element in the sample, and Ei is the expected number of that skeletal element in an individual. Skeletal remains were examined using a stereomicroscope for bone integrity (degree of completeness/fracturing), punctures, notching on the edges of bones, and scratches caused by bird scavenging (Brain 1981; Binford 1981; Andrews 1990; Borrero and Martin 1997; Reeves 2009; Lloveras et al. 2014). There are two types of scratches. First, shallow-penetrating scratches that penetrated the surface of the bone and are relatively linear, although irregularly shaped. The second type of marking was non-penetrating scratches; these markings were characterized by a change in color on the surface of the bone (Reeves 2009).
Spearman’s correlation coefficient (r s) was calculated using PAST software (PAleontological STatistics) version 3.02 at α = 0.05 to compare the NISP of sheep between our actualistic findings and from sheep remains previously collected the American black vulture pellet samples at the study area (Fig. 1) by Ballejo et al. (2012).
Results
Carcass consumption and dispersal patterns
Carcasses of adult sheep, young sheep, and hare were consumed by all scavenging bird species except Turkey vulture (Table 1). The absence of scavenging evidence for Turkey vulture is most likely because populations in Northwestern Patagonia are considerably rarer compared to the other scavenger taxa (Bellati 2000).
Table 1 provides scavenging data (consumption time, number of scavenging birds, and number of scattered bones) for Andean condor, American black vulture, and chimango caracara. Overall, all scavenged and dispersed skeletal elements were observed to be clean of tissue except remains of ligaments and skin (Fig. 3a). Herein, we describe the scavenging of Perimortem adult sheep #10 over 4 h 10 min from carcass consumption to abandonment of the resulting skeletal remains:
-
At 3 h 35 min, a chimango caracara arrived and began consuming the sheep’s eyes.
-
At 4 h 1 min, an Andean condor arrived followed by an additional 28 individuals, displacing the chimango caracara. These birds were the major consumer of the carcass.
-
Ten American black vultures joined the other scavengers at consuming the carcass, taking the opportunity to feed when the condors did not eat. A larger group was observed overflying
-
Finally, 12 southern caracara and four chimango caracara fed on the carcass, while catching body parts scattered by larger scavenger birds.
During this scavenging event, the carcass was displaced 7 m in a light depression on the ground from the place of deposition (Fig. 2). Mandibles were disarticulated and displaced 6.35 m from the carcass and 13.35 m from the place of deposition. Scapulae and forelimbs were not scattered because they remained intact and connected to the body by skin. The right hindlimb and pelvis were articulated by ligaments, whereas the left hindlimb was disarticulated from the pelvis, although it was attached to the body similarly to the scapulae and forelimbs.

a Dispersion of the skeletal elements from postmortem (natural death) carcasses (adult sheep #1–9 and young sheep #1–2); b Dispersion of the skeletal elements from perimortem (actualistic) carcasses (adult sheep #10 and hare)
Herein, we describe the scavenging of Perimortem European hare over 3 h 23 min from carcass consumption to abandonment of the resulting skeletal remains:
-
At 50 min, a chimango caracara arrived to the Perimortem European hare carcass.
-
At 1 h 54 min, the first southern caracara arrived, followed by others who carried away parts of the carcass.
-
Al 2 h 50 min, the first Andean condor arrived, followed by an additional five individuals.
A total of seven chimango caracara, six southern caracara, and six Andean condors fed on the carcass. Finally, the left hindlimb was the only part of the skeleton found and recovered and remained articulated by soft tissue. This anatomical part was displaced 13 m from the place of deposition.
Scavenging birds consumed the postmortem adult sheep (sheep #1–9) in an average of 3 h 16 min (Table 1). American black vulture was the most numerous species eating the carcass (74.8 %), following by Andean condor (12.2 %), southern caracara (11.7 %), and chimango caracara (1.4 %). Scavenging resulted in 77 % disarticulation of the mandibles, and limb bones were found separated from the girdles (Fig. 2). The mean dispersal radius of all sheep carcasses was 6.47 m. The maximum radius of bone dispersal was 20 m represented by left hindlimbs.
Young sheep postmortem (#1 and #2) scavenging averaged 2 h 55 min for two specimens. The most representative species of scavenger birds was American black vulture (73.2 %), followed by Andean condor (16.6 %), chimango caracara (6.5 %), and southern caracara (3.6 %). The mandibles of young sheep #1 were absent, whereas those of young sheep #2 were found dispersed 12 m from the carcass. The mean dispersal radius of the two young sheep carcasses was 13.05 m. The maximum dispersal radius recorded was for a left scapula at 31 m.
Table 2 summarizes MNE, MAU, and MAU% data for adult sheep, young sheep, and European hare carcasses recovered after the scavenger activity. MNE values were coincident with NISP values because most of the bone fragments were missing. Scavenging of adult sheep and young sheep carcasses resulted in a mean relative abundance ranging from 51.90–100 %. Only tibiae and autopodia elements were recovered from the scavenged European hare carcass.
Breakage and bone surface modifications
Our examination of scavenged, perimortem European hare limb bones found an absence of bone modification. New World vultures and caracaras produced scarce damage to adult sheep and young sheep carcasses (Table 3). Bone modifications of all scavenged bones from these carcasses included fractures (5.6 %), notches (1.5 %), non-penetrating scratches (1.2 %), shallow-penetrating scratches (0.3 %), and punctures (0.3 %) (see Fig. 3).

Scavenging modifications on adult sheep carcasses. a Perimortem (adult sheep 10) after bird consumption; b Non-penetrating scratch on femur (adult sheep #10); c Notching (2–3 mm length) on the posterior border of the scapula (adult sheep #7); d Fractures on the transverse processes of the lumbar vertebrae (adult sheep #10); e Puncture (3–5 mm length) on the angular process of the mandible (adult sheep 10)
For perimortem adult sheep #10, 80 % of modifications were scratches (non-penetrating- and shallow-penetrating scratches), mainly in scapulae (Table 3). Fractures of the transverse processes and neural spines of vertebrae, notches in vertebrae, and punctures in the angular processes of mandibles were found (Table 3).
For postmortem adult sheep (#1–9), all the fractures were concentrated in the axial skeleton, such as nasal bones, portions of ribs, and transverse apophyses of vertebrae. 37.9 % of modifications were scratches, mostly in girdles, skulls, and femora. Notching in the posterior border of the scapulae and mandibular condyles was also frequent (23.3 %), and a single punctured scapula was found.
In postmortem young sheep (#1 and #2), 75.3 % of all modifications were fractures in skulls and mandibles, mainly in angular and coronoid processes. One skull specimen was badly damaged with nasals, parietals, and maxillae absent. Additionally, we observed fractured young sheep ribs and vertebral apophyses, notched girdles (17.6 %); punctured skulls and mandibles (4.7 %) and non-penetrating scratches on humeri (2.4 %).
In our comparison of scavenged sheep remains in this study to American black vulture pellets from the ledges of rocky cliffs and trees reported by Ballejo et al. (2012), we found our samples to contain a higher NISP for phalanges, metapodia, vertebrae, and ribs. Although sheep carcasses lacked vertebrae and ribs, phalanges were found (Fig. 4). The results of the Spearman correlation coefficient suggest a positive correlation in NISP between sheep carcasses in our study and those skeletal elements in pellets (r s = 0.521, p = 0.0267).

Comparison of adult sheep and young sheep skeletal abundance resulting from black vulture scavenging between this study and pellet samples of Ballejo et al. (2012)
Discussion
The dispersal patterns, the relative abundance of elements, and the bone modifications are attributed to different variables. Abundance and diversity of the scavenging birds and size of the carcasses are the most important. Thus, the small-sized carcasses (European hare) were those which suffered the most losses and dispersal of their skeletal elements, followed by medium-sized carcasses (young sheep). Probably, the smaller body sizes of these carcasses resulted in the easy transportation of their bones over considerable distances, producing a lower relative abundance of elements. We found disarticulated mandibles and articulated limb bones for young sheep. These most likely results from different bird species being more or less adept at disarticulating young sheep limbs. Also, young sheep showed high levels of fracturing on skulls and modifications in the rest of bones, probably due to reduced ossification in young individuals.
The abundance of species and individuals scavenging carcasses was variable. The Andean condor was observed only once across all of our studied carcasses, and the American black vulture was the most abundant species scavenging carcasses. The role played for each species in the dispersal and bone modification was difficult to identify because observations were made from a range of 150–300 m to avoid intimidating the birds. Nevertheless, we consider the interaction between the different bird species scavenging carcasses an actual natural context a good model to interpret past natural contexts. When comparing our results with those previously published (Reeves 2009; Spradley et al. 2012; Dabbs and Martin 2013), we found differences which could be assigned to the presence of the Andean condor and caracaras.
Comparison with other scavenger birds
Both dispersion and taphonomic features of carcasses consumed by scavenger birds have been studied in North America (Reeves 2009; Spradley et al. 2012). Reeves′ (2009) taphonomic study of American black vulture and Turkey vulture, scavenging pig and goat (Capra aegarus hircus) differed from our results for New World vultures and caracaras. He found that scavenging birds waited approximately 24 h after death before scavenging began. Consumption time reported in his study was also greater and averaged 16 h; the greatest consumption time he found took 24 h. Scavenging initiated with the disarticulation of pig and goat mandibles, followed by skulls, scapulae, and forelimbs, whereas autopodia remained articulated. Overall, the pig and goat had little damage, with the only bone modification being surface furrows on skulls and mandibles.
Dabbs and Martin (2013) reported a taphonomic study of American black vultures scavenging pigs in different environments over a period of 1 year. The authors observed the taphonomic effects of scavengers varied with climate and region and the absence of Turkey vulture despite its presence in the area. They pointed out a severe delay in the time of first arrival (up to 28 days), decreased feeding time on remains (14–40 min), a low feeding rate (7 days to 2 months to skeletonization), and few taphonomic marks.
Spradley et al. (2012) examined American black vulture and Turkey vulture scavenging of human remains. Compared to our study, scavenging birds took 37 days to begin consumption and only 5 h for complete skeletonization. Scavenging birds displaced the body across an area less than 83.6 m2 and spread the human remains from higher to lower elevations with the left forelimb dispersed furthest. He also observed scavenger birds returning to the body in subsequent days, which resulted in the further dispersion of remains.
In our study, we found a lower dispersion distance for adult sheep, young sheep, and European hare. Also, the short time of first arrival to carcasses could be attributed to local enhancement (Buckley 1996, Rabenold 1987) and to the great populations of scavenger birds (with the exception of Turkey vulture) in the area (Bellati 2000; Lambertucci et al. 2009; Ballejo et al. 2012; Lambertucci et al. 2014). The rapid arrival of chimango caracara to the carcasses attracts the rest of the scavenging birds.
We observed more modifications to sheep bones than those reported by Reeves (2009) and Dabbs and Martin (2013), including damaged scapulae and to lesser extent damage to vertebrae, mandibles, girdles, skulls, and femora. Our observations suggest that the large body size and beak morphology of the Andean condor is advantageous for opening the abdominal region of the carcass. These conditions expose more food for those smaller birds which specialize in taking meat (Wallace and Temple 1987; Houston 1988). The results of our study show that the bird is in fact a major scavenger that modifies bones and has a rapid consumption time. However, the resulting numerous remains of adult sheep and young sheep found after scavenging indicate low bone attrition. For hare remains, the lack of skeletal elements and their dispersal pattern is most likely explained by the ease with which birds are able to transport small-bodied animals.
We suggest that the bone modifications observed on uneaten adult sheep, young sheep, and hare bones recovered in open landscapes represent a taphonomic signature of New World vultures and caracaras. This is supported by the scavenging results showing a high completeness of skeletal elements remaining for large-sized carcasses and less completeness for small-sized carcasses, with scarce damage on the bones and the absence of digestive corrosion marks. Similarly, this taphonomic signature also occurs with Egyptian vulture (Neophron percnopterus, Accipitridae) scavenging (Sanchis Serra et al. 2014; Lloveras et al. 2014). This bird transports uneaten remains to nests frequently located in caves and rock shelters, and subsequently punctures bones with its beak during consumption. Thus, scavenging birds, such as the Egyptian vulture and southern caracara, produce bone accumulations representing differential consumption of body parts and bone modification (e.g., digestive corrosion) by prey size and diversity (Lloveras et al. 2014; Montalvo and Tallade 2009, 2010; Montalvo et al. 2011). These results, however, are contrary to Ballejo et al. (2012) for American black vulture pellets with phalanges that exhibited strong digestive corrosion.
Implications for understanding the formation of archaeological sites
Different taphonomic studies demonstrate that scavenger birds are active taphonomic agents of bone accumulations in Old World paleontological and archaeological sites (Robert and Vigne 2002a, 2002b; Marín-Arroyo et al. 2009; Robert and Reumer 2009; Marín-Arroyo and Margalida 2012). Our evaluation of New World vulture and caracara scavenging of perimortem and postmortem adult sheep, young sheep, and European hare is the first taphonomic study to integrate actualistic field data of bone remains, bone dispersal patterns, and bone modification. We suggest two taphonomic models of vulture and caracara scavenging that can assist other researchers with the analysis of animal remains found in the paleontological and archaeological record:
-
1.
Open-air sites. In these sites, researchers can expect to find scavenged remains of large and medium-sized vertebrates to be represented by most skeletal elements (e.g., mandibles, articulated limb bones) dispersed in an average radius of c. 7 m. In the dispersal radius, the greatest transported element would most likely be scapulae. Expected bone modification includes non-penetrating scratches and shallow-penetrating scratches and notches and scarce punctures in skulls, mandibles, and scapulae.
-
2.
Rock shelter or cave sites. In these sites, researchers can expect scavenged accumulations to contain a high relative abundance of small vertebrates. Bone modifications expected can include strong digestive corrosion. This will be particularly prevalent for small animal fractured ribs and vertebrae and autopodia elements (e.g., phalanges). By contrast, large-sized mammals are expected to exhibit strong digestive corrosion (Ballejo et al. 2012).
Conclusion
The results support a distinct difference among perimortem and postmortem carcasses and the scavenging of different-sized carcasses (i.e., large [adult sheep], medium [young sheep], and small [European hare]). Moreover, our comparisons of anatomical representation between pellets (located in cliffs, rock shelter, or cave sites) and scavenged carcass (located in open-air sites) suggest significant differences between archaeological and paleontological sites. In addition, taphonomic similarities and differences with scavenger birds from other parts of the world, and with carnivorous mammals that inhabit Northwestern Patagonia, are clear. In fact, the participation of Andean condor plays a distinctive role in reducing the time of carcass consumption and producing bone modifications.
The results of our study demonstrate the potential of New World vulture and caracara scavenging to contribute in the formation of archaeological sites in addition to those produced by other taphonomic agents (e.g., carnivorous mammals and humans). Although this investigation was performed in Northwestern Patagonia, the taphonomic observations we report can assist other researchers in their development of actualistic scavenging models for archaeological and paleontological sites outside of South America where New World vultures and caracaras occur.
References
Álvarez MC, Kaufmann CA, Massigoge A, Gutiérrez MA, Rafuse DJ, Scheiler NA, González ME (2012) Bone modification and destruction patterns of leporid carcasses by Geoffroy’s cat (Leopardus geoffroyi): An experimental study. Quat Int 278:71–80. doi:10.1016/j.quaint.2011.11.029
Andrews P (1990) Owls, caves and fossils. University of Chicago Press, Londres
Ballejo F, De Santis LJM (2013) Dieta estacional del jote cabeza negra (Coragyps atratus) en un área rural y una urbana en el noroeste patagónico. El Hornero 28:7–14
Ballejo F, Fernández JF, De Santis LJM (2012) Tafonomía de restos óseos provenientes de egagrópilas de Coragyps atratus (Jote cabeza negra) en el noroeste de la Patagonia Argentina. Revista del Museo de antropología 5:213–222
Bellati J (2000) Comportamiento y abundancia relativa de rapaces de la Patagonia Extraandina Argentina. Ornitologia Neotropical 11:207–222
Benson PC, Plug I, Dobbs JC (2004) An analysis of bones and other materials collected by Cape Vultures at the Kransberg and Blouberg colonies, Limpopo province, South Africa. Ostrich 75:118–132. doi:10.2989/00306520409485423
Binford LR (1981) Bones: ancient men and modern myths. Academic Press, New York
Biondi LM, Bó MS, Favero M (2005) Dieta del Chimango (Milvago chimango) durante el periodo reproductivo en el sudeste de la provincia de Buenos Aires. Ornitol Neotrop 15:31–42
Borrero LA, Martín FM, Vargas J (2005) Tafonomía de la interacción entre pumas y guanacos en el Parque Nacional Torres del Paine, Chile. Magallania 33:95–114. doi:10.4067/S0718-22442005000100007
Brain CK (1981) The hunters or the hunted? An introduction to African Cave Taphonomy. University of Chicago Press, Chicago
Buckley NJ (1996) Food finding and the influence of information local enhancement, and communal roosting on foraging success of North American vultures. Auk 113:473–488
Cáceres I, Esteban-Nadal M, Bennàsar L, Fernández-Jalvo Y (2009) Disarticulation and Dispersal Processes of Cervid Carcass at the Bosque de Riofrío (Segovia, Spain). Journal of Taphonomy 7:133–145
Dabbs GR, Martin DC (2013) Geographic variation in the taphonomic effect of vulture scavenging: the case for Southern Illinois. J Forensic Sci 58. doi:10.1111/1556-4029.12025
Del Hoyo J, Elliott A, Sargatal J (1994) Handbook of the birds of the world. New World Vultures to Guineafowl. Lynx Editions, Barcelona, Vol II
DeVault TL, Brisbin IL, Rhodes OE (2004) Factors influencing the acquisition of rodent carrion by vertebrate scavengers and decomposers. Can J Zool 82:502–509. doi:10.1139/Z04-022
DeVault TL, Rhodes OE, Shivik JA (2003) Scavenging by vertebrates: behavioral, ecological, and evolutionary perspectives on an important energy transfer pathway in terrestrial Ecosystem. Oikos 102:225–234. doi:10.1034/j.1600-0706.2003.12378.x
Giacosa RE, Alfonso JC, Heredia N, Paredes J (2005) Tertiary tectonics of the sub-Andean region of the North Patagonian Andes, southern central Andes of Argentina. J S A Earth Sci 20:157–170. doi:10.1016/j.jsames.2005.05.013
Giacosa RE, Márquez MJ (1999) Jurásico y Cretácico de la Cordillera Patagónica septentrional y Precordillera Patagónica. In: Caminos R (ed) Geología Argentina 17. SEGEMAR, Buenos Aires, pp. 444–459
Grayson DK (1984) Quantitative zooarchaeology: topics in the analysis of archaeological faunas. Academics Press, Orlando
Grzimek’s B (2002) Animal Life Encyclopedia, 2nd edn. Vol 8, Birds. In: Jackson I, Bock JA, Olendorf WJ (eds) Gale Group, United Estates
Haynes G (1982) Utilization and skeletal disturbances of North American prey carcasses. Artic 35:266–281. doi:10.14430/arctic2325
Hill A (1979) Disarticulation and Scattering of Mammal Skeletons. Paleobio 5:261–274
Houston DC (1988) Competition for food between Neotropical vultures in forest. British Ornithologists Union 130(3):402–417
Kelly NE, Sparks DW, DeVault TL, Rhodes OE (2007) Diet of black and turkey vultures in a forested landscape. Wilson J Ornithol 119:267–270. doi:10.1676/05-095.1
Lambertucci SA, Alarcón PAE, Hiraldo F, Sanchez-Zapata JA, Blanco G, Donázar JA (2014) Apex scavenger movements call for transboundary conservation policies. Biol Conserv 170:145–150. doi:10.1016/j.biocon.2013.12.041
Lambertucci SA, Trejo A, Di Martino S, Sanchez-Zapata JA, Donázar JA, Hiraldo F (2009) Spatial and temporal patterns in the diet of the Andean condor: ecological replacement of native fauna by exotic species. Anim Conserv 12:338–345. doi:10.1111/j.1469-1795.2009.00258.x
León RJ, Bran D, Collantes M, Paruelo JM, Soriano A (1998) Grandes unidades de vegetación de la Patagonia extra andina. Ecología Austral 8:125–144
Lloveras L, Nadal J, Moreno-García M, Thomas R, Anglada J, Baucells J, Martorell J, Vilasís D (2014) The role of the Egyptian Vulture (Neophron percnopterus) as a bone accumulator in cliff rock shelters: an analysis of modern bone nest assemblages from North-eastern Iberia. J Archaeol Sci 44:76–90. doi:10.1016/j.jas.2014.01.018
Lyman RL (1994) Vertebrate Taphonomy. Cambridge University Press, Cambridge
Marín-Arroyo AB, Fosse P, Vigne JD (2009) Probable evidences of bone accumulation by Pleistocene bearded vulture at the archaeological site of El Mirón Cave (Spain). J Archaeol Sci 36:284–296. doi:10.1016/j.jas.2008.09.007
Marín-Arroyo AB, Margalida A (2012) Distinguishing bearded vulture activities within archaeological contexts: identification guidelines. Int J Osteoarchaeol 22:563–576. doi:10.1002/oa.1279
Martin FM, Borrero LA (1997) A Puma Lair in Southern Patagonia: implications for the Archaeological Record. Curr Anthropol 38:453–461
Mondini MN (1995) Artiodactyl prey transport by foxes in puna rock shelters. Curr Anthropol 36(3):520–524. doi:10.1086/204391
Mondini MN (2004) Accumulation of small and large vertebrates by carnivores in Andean South America. In: Brugal JP, Desse J (eds) Petits Animaux et Sociéte′s Humaines Du Complément Alimentaire aux Ressources Utilitaires. Actes des XXIVe’mes Rencontres Internationales d’Archéologie et d’ Histoire d’Antibes. Éditions APDCA, Antibes, pp. 513–517. doi:10.1016/j.quaint.2007.08.013
Montalvo CI, Tallade PO (2009) Taphonomy of the accumulations produced by Caracara plancus (Falconidae) Analysis of Prey Remains and Pellets. Journal of Taphonomy 7:235–248
Montalvo CI, Tallade P (2010) Análisis tafonómico de restos no ingeridos de roedores presa de Caracara plancus (Aves, Falconidae). In: De Nigris M, Fernández PM, Giardina M, Gil AF, Gutiérrez MA, Izeta A, Neme G, Yacobaccio HD (eds) Zooarqueología a Principios del Siglo XXI: Aportes Teóricos, Metodológicos y Casos de Estudio. Ediciones del Espinillo, Buenos Aires, pp. 419–428
Montalvo CI, Tallade PO, Fernández FJ, Moreira GJ, Rafusee DJ, De Santis L (2011) Bone damage patterns found in the avian prey remains of crested caracara Caracara plancus (Aves, Falconiformes). J Archaeol Sci 38:3541–3548. doi:10.1016/j.jas.2011.08.021
Novillo A, Ojeda RA (2008) The exotic mammals of Argentina. Biol Invas 10:1333–1344. doi:10.1007/s10530-007-9208-8
Paruelo JM, Beltran A, Jobbagy E, Sala OE, Golluscio RA (1998) The climate of Patagonia: general patterns and controls on biotic processes. Ecología Austral 8:85–101
Rabenold PP (1987) Recruitment to food in black vultures: evidence for following from communal roosts. Anim Behav 35:1775–1785. doi:10.1016/S0003-3472(87)80070-2
Reeves NM (2009) Taphonomic effects of vulture scavenging. J Forensic Sci 54:523–528. doi:10.1111/j.1556-4029.2009.01020.x
Ringuelet R (1961) Rasgos fundamentales de la zoogeografía Argentina. Physis 22:51–170
Robert I, Reumer JWF (2009) Taphonomic reinterpretation of a bone sample of endemic Pleistocene deer from Crete (Greece): osteoporosis versus regurgitation. Palaeodiversity 2:379–385
Robert I, Vigne JD (2002a) The bearded vulture (Gypaetus barbatus) as an accumulator of archaeological bones. Late glacial assemblages and present-day reference data in Corsica (Western Mediterranean). Acta Zool Cracov 45:319–329. doi:10.1006/jasc.2001.0778
Robert I, Vigne JD (2002b) Bearded vulture Gypaetus barbatus contributions to the constitution of two different bone assemblages: modern reference data and an archaeological example in Corsica. J Archaeol Sci 29:763–777
Sanchis Serra A, Real Margalef C, Morales Pérez JV, Pérez Ripoll M, Tormo Cuñat C, Carrión Marco Y, Pérez Jordá G, Ribera Gómez A, Bolufer Marqués J, Villaverde Bonilla V (2014) Towards the identification of a new taphonomic agent: an analysis of bone accumulations obtained from modern Egyptian vulture (Neophron percnopterus) nests. Quat Int 330:136–149. doi:10.1016/j.quaint.2013.10.047
Spradley MK, Hamilton MD, Giordano A (2012) Spatial patterning of vulture scavenged human remains. Forensic Sci Int 219:57–63. doi:10.1016/j.forsciint.2011.11.030
Vargas RJ, Bo MS, Favero M (2007) Diet of the southern caracara (Caracara plancus) in Mar Chiquita reserve, Southern Argentina. J Raptor Res 41:113–121
Wallace MP, Temple SA (1987) Competitive interactions within and between species in a guild of avian scavengers. Auk 104:290–295
Yáñez JL, Núñez H, Jaksic FM (1982) Food habits and weight of Chimango Caracara in central Chile. Auk 99:170–171
Yravedra J, Lagos L, Bárcena F (2012) The wild wolf (Canis lupus) as a dispersal agent of animal carcasses in Northwestern Spain. Journal of Taphonomy 10:219–238. doi:10.1111/j.1469-7998.2007.00305.x
Acknowledgments
This work has been funded by Project 11/N769 of the Facultad de Ciencias Naturales y Museo, UNLP, and Project 210 of the Facultad de Ciencias Exactas y Naturales, UNLPam. Special thanks are given to Cecilia Deschamps for revising the English, the labor of Mailen De Santis, Carolina Gabriela Sena and Federico Ballejo in the field, and the constructive advice of Luis Del Papa. Guest editors Richard Madgwick and Lee G. Broderick and two anonymous reviewers provided valuable suggestions on an early version of the paper.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Ballejo, F., Fernández, F.J., Montalvo, C.I. et al. Taphonomy and dispersion of bones scavenged by New World vultures and caracaras in Northwestern Patagonia: implications for the formation of archaeological sites. Archaeol Anthropol Sci 8, 305–315 (2016). https://doi.org/10.1007/s12520-015-0277-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-015-0277-9