
Abstract
Stevens et al. (Oxford J Archaeol 29: 407–428, 2010) speculated that the extensive faunal intra-population isotopic variability at Danebury hillfort was due to the animals being husbanded within various ecological isozones (i.e. microenvironments with distinct natural or anthropogenic isotopic baselines) within the Danebury Environs, and subsequently brought to the hillfort. We test this hypothesis through isotopic analysis of 357 animals from five sites in the Danebury Environs (Suddern Farm, Nettlebank Copse, Houghton Down, New Buildings, Bury Hill). Our results demonstrate that the hypothesis is incorrect as the Environs animals also exhibited extensive intra-site isotopic variability. The observed patterns require considerable human management of herds and flocks, either through animal droving over long distances, or movement restriction through personal attendance or penning/corralling within a landscape containing a mosaic of isozones. Large-scale importation of animals to the Danebury sites from distant regions that have different isotopic baseline values is possible. However, the practical requirements of this model (i.e. long distance trade networks maintained in terms of frequency, proportion and source of animals over 500 years) make this improbable. Rather, we propose a model that includes distinct ecological isozones within the Danebury Environs landscape, with some animals from each of the sites consuming foodstuff from different isozones for most of their lives. This likely involves control of the landscape on a communal probably kinship basis, with individuals having access to certain parts of the landscape at certain times. The landscape could be seen as being made up of numerous interlocking activity-territories, defined in both space and time, remembered and inherited. Irrespective of which model proves to be the most accurate, these results paint a picture of complex land management during the Iron Age.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
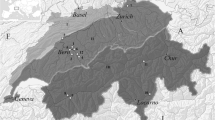
Danebury, one of the largest and most impressive Iron Age hillforts in Southern Britain, is located in Stockbridge, Hampshire (Fig. 1) and was occupied during the latter half of the first millennium BC. Like other hillforts, Danebury is thought to have been both a place of residence and a centre where a range of communal and ritual activities took place, including food storage, processing and exchange (Cunliffe 1984a, 1984b, 1995). Evidence supporting the idea of Danebury being a focus in a redistribution network comes from palaeobotanical, zooarchaeological and stable isotope data. Palaeobotanical remains indicate that grain was being brought to the site for processing and was then stored in pits and granaries prior to redistribution (Jones 1984, 1995; Jones and Nye 1991). Stable isotope analysis of cereal grains indicate that crops from several locations may have been mixed within the hillfort (Lightfoot and Stevens 2012). Stable isotope analysis of animal bone collagen provides further support for the idea of resources being brought to Danebury for redistribution. Extensive heterogeneity is visible in the animal isotope values at Danebury, which indicates the animals consumed a variety of isotopically distinct diets (Stevens et al. 2010). We speculate that the substantial variation in Danebury animal isotope values is due to the animals being husbanded at a number of locations within the Danebury Environs that have different microenvironments with distinct natural or anthropogenic isotopic baselines (hereafter “isozones”). The animals would then be brought to the hillfort. Thus we would expect animals from archaeological sites within the Danebury Environs to have more limited intra-population isotopic variability than at Danebury and considerable inter-population variability. To test this theory we conducted carbon and nitrogen isotope analysis on animal bones from five of the Danebury Environs archaeological sites, Suddern Farm, Nettlebank Copse, Bury Hill, New Buildings and Houghton Down, with the expectation that they would all exhibit a more limited range in faunal isotope values than seen at Danebury hillfort.

Location of Danebury hillfort and Environs sites sampled for this study. Modified from Cunliffe and Poole 2000a
Sites
Danebury (summarized from Cunliffe 1984b, 1993, 1995; Grant 1984, 1991)
Danebury hillfort, Hampshire, is located at relatively low altitude (145 m OD), however, as the surroundings rarely exceed 100 m, Danebury is the dominant feature in the landscape (Fig. 1). The hillfort was occupied during the latter half of the 1st millennium BC. It is found on chalkland, which dips gradually to the south until it becomes overlain by clays and sands in the Hampshire Basin, and dips more steeply to the north to a scarp slope 24 km away, overlooking the valley of the River Enborne, a tributary of the Thames. Soils covering the chalk within the region are thin (200–300 mm thick), but in places patches of clay-with-flints cap the hilltops. The surrounding river valleys contain colluvial and alluvial deposits. Upland pastures were appropriate for grazing sheep, the gentle slopes and light soils lend themselves to cultivation and the wide flood plain of the River Test contained water meadows particularly suitable for grazing cattle. Details of animal mortality profiles and the husbandry focus at Danebury Hillfort are given in Table 1.
Suddern Farm (summarized from Cunliffe and Poole 2000b)
Suddern Farm, Middle Wallop, is a settlement site occupied more or less continuously from the Early Iron Age to the first century AD. It is situated on a low spur of chalk, c. 85 m OD, above the valley of the Wallop Brook, which lies 1 km to the east (Fig. 1). To the west, the land rises steeply to Suddern Hill with the higher ridge of downland, reaching 170 m OD, just beyond. The site is in a prime location, with good arable land, and easy access to well-watered pastures for cattle and open high down where sheep could graze. There is also a small Iron Age cemetery at the site, which has been the subject of palaeodietary isotopic investigations (Stevens et al. 2013). Cattle, sheep, goat, pig, horse, dog, wild mammals and birds were present within the zooarchaeological assemblage. Sheep and cattle predominated, and there were few changes in the assemblage through time. Samples for isotopic analysis were taken from each of the main domesticates (namely, sheep, cattle, pig, horse and dog) throughout the period of occupation, although only one dog bone was sampled. The results of zooarchaeological analysis are presented in Table 1, which details mortality profiles and husbandry emphasis for this and each of the Danebury Environs sites.
Nettlebank Copse (summarized from Cunliffe and Poole 2000d)
Nettlebank Copse, Wherwell, occupies a shallow spur facing eastwards along a dry valley which leads to the River Test some 3 km away (Fig. 1). The chalk bedrock has a discontinuous mantle of clay-with-flints and it is likely, but not proven, that erosion from the valley sides has created a rich colluvial deposit in the dry valleys. There may have been nearby springs but the nearest permanent water supply would probably have been the River Test. Fertile clay-rich soil is available in the immediate region. Nettlebank Copse is a two-phase site, with an Early Iron Age settlement and a Late Iron Age banjo enclosure. The Early Iron Age phase was a small, likely single-family farming unit consisting of an oval shaped enclosure with two circular houses. It was occupied for a short time in the fourth century BC, perhaps 20 to 30 years. The site was likely occupied year-round. The first ditch of the Late Iron Age banjo enclosure was probably dug shortly after the settlement was abandoned, c. 300 BC. The second ditch dates to the mid-first century BC until the mid-first century AD. The lack of young animals suggests that lambing and calving did not take place at the site, and indeed that the site may not have been used at all in spring or summer.
The Early Iron Age animal bone assemblage contains cattle, sheep, horse, pig and dog, with one possible goat. Small mammal and amphibian bones were also noted, but no large or medium wild mammals or fish were recovered. The Late Iron Age animal bone assemblage was very similar, containing sheep, cattle, pig, horse and dog, plus fox, hare and red deer. In both periods sheep were the most numerous species, followed by cattle then pig. Samples were taken for isotopic analysis from the five main domesticated species. Details of the husbandry focus are given in Table 1.
Bury Hill (summarized from Cunliffe and Poole 2000a)
Bury Hill, Upper Clatford, is a two-phase hillfort site dated to the Early Iron Age (not sampled) and the Middle Iron Age in the late second- or early first century BC. It occupies the relatively level summit of a low hill that rises to a maximum of 100 m OD (Fig. 1). The location allows the site to command the confluence of the valleys of the rivers Anna and Anton. The river valleys are floored with alluvium and flanked by narrow gravel terraces. The rivers would have provided a permanent water supply and a continuous band of meadow 200 m wide. This would be suited to cattle rearing, possibly providing hay in the late summer and pasture for much of the early part of the year. The comparatively gentle slopes nearby meant that most of the area was available for arable which, in times of fallow, would have provided good gleaning for sheep and pigs. The hillfort is therefore well situated to exploit both the upland chalk soils and the pasture and hay meadows of the river flood plains.
The zooarchaeological assemblage contained horse, sheep, cattle, pig, dog, fox and two wild bird species, but the assemblage was not well-preserved compared to the other Danebury Environs sites due to the more acidic, clayey soil. The zooarchaeological assemblage contained an unusually high proportion of horse, ten times that at other sites. Samples for isotopic analysis were only taken from horse bones from the later phase of the hillfort due to poor preservation. Details of the husbandry focus are given in Table 1.
New Buildings (summarized from Cunliffe and Poole 2000c)
New Buildings is a mid-first millennium settlement site, dating from c. 470 to 360BC, with a Late Bronze Age occupation which was not sampled and therefore will not be discussed further. It is situated at 100 m OD on the south-facing flank of the chalk ridge that runs eastwards midway between Danebury and the River Test (Fig. 1). Today, the soil is 0.15 m thick and it is unlikely that it was ever significantly thicker. The soil has a comparatively high clay content that adds to its coherence and stability. Immediately to the west of the site there is evidence of extensive arable exploitation on the downland itself. In addition, the site was 1 km away from the water meadows of the Test valley, providing water and food for cattle. The zooarchaeological assemblage was small but indicates the presence of cattle, sheep, horse, pig and dog. Samples for isotopic analysis were taken from all of these species, although only one dog bone was sampled. Details of the husbandry focus are given in Table 1.
Houghton Down (summarized from Cunliffe and Poole 2000e)
Houghton Down is a settlement site situated on Chatgrave Hill on Houghton Down, Stockbridge. It has two phases of occupation, Early (c. 800 to 270BC) and Late (c. 100BC to AD400). As samples were only taken from the former, this discussion will be limited to the Early period. The settlement is situated on the crest of a slight rise in a ridge of undulating downland and stretches from the crest of the hill at 100 m OD down a gentle north-facing slope to 95 m OD (Fig. 1). It is bounded to the east by the valley of the river Test and to the west by a narrow dry valley. From the settlement, the ground slopes gently to the edge of the floodplain c. 1 km to the east. The soil of the ridge in this area is thin and does not have a capping of clay-with-flints. The location of the site near to a permanent water supply would have facilitated cattle husbandry and the floodplain would have provided a wide range of useful resources. The faunal assemblage indicates the presence of sheep, cattle, pig, horse, dog and cat, possibly otter, domestic fowl and other birds at the site, with sheep the most numerous species, followed by cattle and then pig. Samples for isotopic analysis were taken from each of the five main domesticates, although only one dog bone was sampled. Details of the husbandry focus are given in Table 1.
Farming practices in the Danebury environs
The farming system represented at the Danebury Environs sites, and at Danebury itself, was the familiar Iron Age system seen at many southern English sites, based on spelt wheat and hulled six-row barley, with cattle, sheep and pig as the major domesticated animals. In common with many other “Wessex” sites, the proportion of sheep was high (Hambleton 2008), which together with evidence from age distributions and material culture suggests that wool production was important. Bury Hill, with its high proportion of horse and horse-related artefacts, is an exception, and may be a site with a special emphasis on training and management of horses (Hamilton 2000). Animals are vital in mixed farming system as providers of manure (Bogaard 2012). The downland slope soils were vulnerable and had probably degraded considerably by the Iron Age. The increase in sheep during the Iron Age was likely the result of the need to manure large tracts of land to maintain crop yields. The sheep are likely to have been kept for a large part of the year on land which they were manuring which has obvious implications for soil δ15N values which are raised by manuring (see below).
Within this overall uniformity, Campbell (2000) and Hamilton (2000) found evidence of variation in the way the different sites were used, in terms of emphasis on different suites of activities at different times of year. In the Early and Middle Iron Age the picture is of a landscape with occupied sites of various types and sizes, supported by the “normal” Iron Age farming system. Somewhere around 270 BC there was a change in the ceramic style, representing an eastward reorientation of exchange networks away from the Salisbury Plain/Avon domain towards the South Downs zone. Around the same time, Danebury underwent a major refurbishment and intensification of use. Danebury sat virtually alone in the landscape as there is little or no evidence that the Danebury Environs sites were occupied at this time (apart from the previously unoccupied Bury Hill, with its unusual animal bone assemblage, which seems to fall late in the sequence) (Table 2). The evidence implies that Danebury was now the focus of occupation, as well as of exchange systems. It is therefore possible, indeed probable that husbandry practices and economic networks varied significantly over the relevant period. Yet there is no discernible difference in the archaeozoological or archaeobotanical evidence to suggest that the landscape was being used differently—somehow, in spite of the shift in archaeologically visible settlement patterns, the ways in which the different environmental zones were being used appears to have remained the same. In the Late Iron Age, around 100 BC, there was another shift in ceramic tradition, Danebury and Bury Hill were abandoned, and most of the Danebury Environs sites were reoccupied. There is evidence for developing contacts with mainland Europe via the south coast. This time there were changes in the farming system: the proportion of cattle in the animal bone assemblages rose, new crops (oats and peas) appeared, and Campbell (2000) and Hamilton (2000) suggest that the annual regime switched from autumn sowing of cereals to both spring and autumn sowing. These would represent major behavioural changes for people, with considerable reorganisation of activity schedules, and possibly of landscape use.
Background to isotopes
Stable isotope analysis of archaeological fauna is a quantitative method for reconstructing their dietary intake. This method has advantages over traditional archaeozoological methods in that it enables comparisons to be made both between individuals within a population and between populations. However for a holistic investigation of archaeological fauna it is best used alongside traditional zooarchaeological techniques. Stable isotope analysis has been used to investigated the diet and husbandry of archaeological animals from a number of sites in Britain (e.g. Mulville et al. 2009; Millard et al. 2011; Madgwick et al. 2013; Jones et al. 2012). The stable isotope technique is based on the premise that the food consumed by an animal is used to construct the body tissues, thus the body chemistry reflects that of the dietary intake. The isotopic composition of bone collagen, the tissue most frequently analysed in archaeological studies, reflects average diet over the last years of an animal’s life and primarily reflect the protein component of the diet (Hedges and Reynard 2007; Ambrose and Norr 1993; Tieszen and Fagre 1993).
Nitrogen stable isotopes can be used to infer an animal’s trophic position as there is a +3 to +5 ‰ enrichment seen with each increase in trophic level (Bocherens and Drucker 2003). Carbon stable isotope analysis can be used to detect the consumption of either marine resources (e.g. Schoeninger et al. 1983) or C4 plants (e.g. Vogel and Van der Merwe 1977), although neither of these factors are likely to be relevant to this study. A small trophic level enrichment is observed between diet and consumer, typically 0 to +2 ‰ (Bocherens and Drucker 2003).
Although diet is the main control over animal bone collagen isotope values, environmental factors can influence the isotopic signatures of plants and are then passed up the foodchain (Heaton 1999; Stevens and Hedges 2004; Stevens et al. 2008). Small scale variation in C3 plant δ13C values of the magnitude of a few per mille have been shown to be linked to light, water and nutrient availability, temperature, salinity, and atmospheric CO2 δ13C and partial pressure. These factors can cause a plant to increase its stomatal conductance and/or decrease its carboxylation rate. Of these factors, water availability appears to be the most influential parameter (Heaton 1999; Dawson et al. 2002). Soil and plant δ15N have been shown to increase with decreasing mean annual temperature (and to a lesser extent with increasing mean annual precipitation) (Amundson et al. 2003). Within landscape variation of plant and fauna δ 15N may be linked to local variation in nitrogen cycling tied to topography and soil inputs (Ugan and Coltrain 2011).
Methodology
Three hundred fifty-seven animals (cattle, pig, sheep, horse, and dog) were sampled for isotope analysis. Samples were prepared at the RLAHA, Oxford and at the McDonald Institute for Archaeological Research, Cambridge, according to the method used by Privat et al. (2002). Isotopic analysis of samples was performed at RLAHA using an automated Carlo Erba carbon and nitrogen elemental analyser coupled in a continuous flow mode to an isotope ratio-monitoring (PDZ Europa Geo 20/20) mass spectrometer. All samples were analysed at least in duplicate. Carbon and nitrogen results are measured relative to VPDB and AIR standards, respectively and reported in parts per mille (‰) (Hoefs 1997). Repeat analysis of in-house stands (nylon and alanine calibrated against IAEA standards) indicated that measurement errors were less than 0.2 ‰ for both carbon and nitrogen.
Results
Of 357 samples, only 3 failed to produce enough collagen for isotope analysis; a cow, a sheep and a horse all from Nettlebank Copse. Extracted collagen had C/N atomic ratios between 3.1 and 3.4 (Online supplementary dataset 1) which is within the range of 2.9 to 3.6 considered indicative of good collagen preservation (DeNiro 1985; Ubelaker et al. 1995).
Both the carbon and nitrogen isotope results for the different species follow the general pattern observed in archaeological specimens the British Isles (Fig. 2). The δ13C and δ15 N values for all of the fauna are listed in the online supplementary dataset and summary statistics (mean, standard deviation, minimum, maximum and range) for each species at each site are given in Table 3 and outliers can be seen in the boxplots shown in Figs. 3 and 4. The δ13C values are within the typical range observed for animals living in ecosystems dominated by C3 plants. The δ13C of the cattle, sheep and pigs are similar, whereas the horse δ13C are typically lower relative to the ruminants due to them being hind-gut fermenters. The δ15N of the cattle, sheep and horse are similar, whereas the pig δ15N are generally higher due to an omnivorous diet. Therefore isotopic variation in pigs is a reflection of both isotopic variation in the plants they consumed and quantity of animal protein they consumed.

Faunal and human carbon and nitrogen isotopes from Danebury Hillfort and Environs sites

Box plot of faunal carbon isotopes for each species by site; Bury Hill (blue), Danebury (green), Houghton Down (beige), Nettlebank Copse (purple), New Buildings (yellow), Suddern Farm (red)

Box plot of faunal nitrogen isotopes for each species by site; Bury Hill (blue), Danebury (green), Houghton Down (beige), Nettlebank Copse (purple), New Buildings (yellow), Suddern Farm (red)
As different periods of the Iron Age are represented at the different sites (Table 2) it is necessary to check for temporal trends within sites before undertaking an inter-site comparison. Within each multi-period site (Suddern Farm, Danebury, Nettlebank Copse) no differences were observed in the δ13C or δ15 N values for each species between the EIA, MIA, MIA/LIA, LIA, 1st C AD. However, when all sites were considered together some differences in sheep δ15N were observed between time periods. Mean sheep δ15N during the EIA (δ15N = 4.5 ± 0.8 ‰, n = 78) was significantly higher than that of sheep in the MIA/LIA (n = 21, δ15N = 4.0 ± 0.8 ‰) (one-way ANOVA with post-hoc Bonferroni correction, P = 0.023). Furthermore, mean sheep δ15N during the early Iron Age was significantly higher than that of sheep in the LIA (δ15N = 4.0 ± 0.6 ‰, n = 38) (one-way ANOVA with post-hoc Bonferroni correction, P = 0.002). These temporal differences are not driven by one particular site. Given that only a weak temporal trend is observed in the δ15N of only one species, climatic and/or environmental parameters are unlikely to be the driving factor and thus fauna from all time periods at each site are subsequently considered collectively.
In order to assess whether the isotopic variation observed at Danebury could be due to the gathering of animals from the Environs sites, we compare the isotopic signatures of fauna between sites. Our expectation was that each site would have its own distinct isotopic range (i.e. its own “isozone”). Although some statistically significant differences were observed between the mean isotope signatures of particular species at select sites, no consistent pattern was observed between any sites. First, no differences were seen in pig carbon or nitrogen isotope values between sites. In carbon, four statistically significant differences were seen in horse, cattle and sheep. New Buildings mean horse δ13C (n = 13, −22.5 ± 0.6 ‰) is different to both Nettlebank Copse (n = 18, −22.0 ± 0.4 ‰) and Suddern Farm (n = 26, −22.1 ± 0.3 ‰) (Kolmogorov Smirnov Z tests, p = 0.022 and p = 0.013, respectively). New Buildings mean sheep δ13C (n = 19, −21.3 ± 0.3 ‰) is marginally different to Houghton Down (n = 18, −21.0 ± 0.2 ‰) (Kolmogorov Smirnov Z test, p = 0.047). Finally, Danebury mean cattle δ13C (n = 39, −21.6 ± 0.3 ‰) is different to Nettlebank Copse (n = 35, −21.3 ± 0.4 ‰) (Mann Whitney test, p = 0.001). In nitrogen, two statistically significant differences were seen in sheep and cattle. New Buildings mean sheep δ15N (n = 19, 4.8 ± 0.8 ‰) is different to Nettlebank Copse (n = 34, 4.1 ± 0.8 ‰) (Kolmogorov Smirnov Z test, p = 0.012) and New Buildings mean cattle δ15N (n = 17, 4.7 ± 1.2 ‰) is marginally different to Suddern Farm (n = 39, δ15N = 3.7 ± 1.4 ‰) (Kolmogorov Smirnov Z test, p = 0.041). The lack of consistent offsets between sites suggests that the fauna at each site are not feeding within distinct isozones.
Our second expectation was that each Environs site would have a much smaller range of isotopic variation than that seen at Danebury hillfort. As seen in Table 3, the range of variation observed for both carbon and nitrogen in all species at Danebury is similar to the ranges of variation seen at the Environs sites. The one exception to this is the pig range in δ15N values, which is substantially larger at Danebury (6 ‰) than at the other sites (1.8 to 3.7 ‰). However, when the two outliers at Danebury, as identified in Fig. 4 (DBH93 and DBH127), are removed the range of variation is reduced to 2.2 ‰, similar to the other sites. Although the ranges are sometimes slightly less at Houghton Down than the other sites, this appears to be due to the smaller sample size at this site compared to the others.
The amount of intra-species isotopic variation observed at Danebury hillfort and each of the Environs sites is very large, particularly in nitrogen. Up to 5.4, 5.3 and 3.6 ‰ variation in δ15N is seen in cattle, horse and sheep, respectively (Table 3, Fig. 5a–d). When all sites are considered collectively the δ15N ranges increase (Figs. 2 and 5). The variation seen in cattle and horse are 6.1 ‰ and 5.6 ‰ respectively (δ15N typically range from 1.5 to 6.5‰). The δ15N variation in sheep is notably smaller at 4.0 ‰ (δ15N typically range from 3 to 6.5 ‰). The intra-population variability for each species observed at archaeological sites around the world is typically 2 ‰. Furthermore, an increase of 3 to 5 ‰ in δ15N is routinely observed between trophic levels (Bocherens and Drucker 2003). Here, we see an extremely large intra-population variation equivalent to at least one trophic level in sheep and more than one trophic level in cattle and horses.

Faunal δ13C and δ15N values for fauna from Danebury hillfort and the Environs sites (a cattle, b sheep, c horses, d pigs)
Discussion
We speculated that the large amount of intra-population isotopic variability observed at Danebury was due to the animals being husbanded at a number of different ecological isozones within the Danebury Environs and subsequently brought to the hillfort (Model: Fig. 6). Archaeological sites within the Danebury Environs should therefore have more limited intra-population isotopic variability than at Danebury, and greater inter-than intra-population variability. However, the isotope results do not support this straightforward model. Instead, a more nuanced model is required to explain the observed animal isotope signatures.

Economic models 1 (solid lines) and 2 (solid and dashed lines) (see text for description)
In order to produce the large range in isotopic variation seen within Danebury and the Environs sites every species must have had individuals that consumed isotopically different diets. Furthermore, the cattle and horses must have has access to a wider range of isotopically distinct diets than the sheep. The most likely scenario in which this could occur is for subgroups of each species at each site to have spent a significant proportion of their lives feeding from distinct ecological isozones. These isozones could be due to natural isotopic variation between landscapes or within the landscape, for example between watermeadows and upland pasture, or anthropogenic, for example due to collected fodder or intensive manuring of certain crops or areas.
We envisage several economic models in which animals that lived the majority of their lives in different isozones could be found together in the zooarchaeological assemblages at Danebury hillfort and the Environ sites.
A second model (Fig. 6) has each Environs site situated within its own ecological isozone, as in model 1, but this is not detectable in the animal isotopic values because of very large-scale redistribution of animals via Danebury. A very high number of the adult animals from the Environs sites (some of them close to the time of culling) would have had to have been gathered at Danebury. They would then be mixed and randomly redistributed to each of the sites such that each site has the full range of isozones represented. This could also include movement directly between the Environs sites. This model is not parsimonious as it requires an extremely high rate of animal movement through exchange, and the amount of animal movement between sites with distinct isozones to be equal and constant throughout the Iron Age. If each site did have its own unique isozone but also imported some animals from different isozones, we would expect each site to be biased toward the isotopic baseline of its own isozone.
In a third model (Fig. 7), the distinct isozones are not in the Danebury region. Animals are brought into Danebury from at least one area outside of the region and redistributed to the Environs sites. In addition to this, there could also be movement of some animals within the Environs sites, and between the Environs sites and Danebury. Weaknesses of this model relate to the duration of the exchange network. The mechanism by which animals were moved and exchanged over long distances would have had to have been maintained over a 500-year period with little or no variation in the proportion and source of imports. First in support of this model, there is evidence that a range of ecological isozones existed across Iron Age Britain. Low nitrogen isotope values like some of those observed in the Danebury Environs have not been widely found at Iron Age sites outside Hampshire whereas higher nitrogen isotope signatures are seen in other regions of the British Isles (Jay and Richards 2007). Second, various pieces of evidence exist suggesting long-distance animal movement took place during British prehistory. Droveways are thought to have become established at this time. Strontium isotope analysis of a single horse from the nearby Iron Age site of Rooksdown in Hampshire, showed that it was not raised locally and may have been imported from as far away as Wales, Scotland or the continent (Bendrey et al. 2009). Similar analyses of cattle from the Late Neolithic site of Durrington Walls, Wiltshire, demonstrated that a high proportion of cattle were not born locally, and must have travelled substantial distances to the site from a range of areas within the British Isles. However, strontium isotopes of a single horse from Bury Hill indicate that this animal at least was born on chalkland, the dominant geology of the local region (Bendrey et al. 2009). Further strontium isotope analysis of Iron Age fauna from the Danebury Environs would provide evidence in support of or against this model.

Economic model 3 (see text for description)
In our final model (Fig. 8), distinct ecological isozones exist within a relatively small area of the Hampshire landscape, and at least some individual animals of each species had their movement restricted to specific isozones within the landscape. Boundaries of isozones may relate to landscape properties such as gradient, distance from river, underlying geology, soil type, and so forth. Artificial isozones may also exist due to human influence, such as intense manuring of a specific area. In this scenario, people at each of the Environs sites had access to food stuffs from multiple isozones on which the animals were reared, resulting in the wide variation in faunal isotope values. Furthermore, some individuals from each species must have had their movement restricted. Some animals must have been reared on a particular isozone for the majority of their lives, while others from the same site must have spent a similar amount of time feeding from a different isozone. Additionally, some animals may have consumed foodstuffs from two or more isozones (either through animal movement or foddering practices) producing an averaged isotopic signal. This does not necessarily have to happen simultaneously, or even within the same year. However, it must happen over a relatively small number of years, because high isotopic variation is seen within single excavation units. This model could also incorporate the movement of animals within the Environs sites, between the Environs sites and Danebury, and from areas outside of the Danebury region. However, the strength of this model is that it does not require a consistent pattern of large-scale exchange or long-distance movement over a 500-year period.

Economic model 4 (see text for description)
Within this final model, there are two scenarios by which multiple isozones could exist within the region of each of the sites. The first is that anthropogenic isozones could be created within the Danebury Environs landscape by intensive manuring and selective foddering. Long-term intensive manuring has been shown to increase plant δ15N values by up to 9 ‰ (Fraser et al. 2011). In this scenario, the low herbivore δ15N signatures would relate to animals consuming vegetation from unmanured contexts and those with high δ15N values would relate to animals consuming vegetation from intensively manured contexts. Those herbivores with intermediate δ15N values would reflect the consumption of plants that have been subjected to low level manuring (e.g. due to animals being grazed on fields in rotation so that their dung could enrich the soil) or alternatively the consumption of plants from both intensively manured and non-manured contexts. This is possible; however, if such extensive variation in manuring practice was typical in the Iron Age we would expect to see a similar large amount of variation in other locations in the British Isles. This is not to say that manuring does not account for some of the herbivore isotopic variation. Indeed the increase in periodontal disease in sheep through time points to continued degradation of soil, thus manuring must have been important for maintaining soil fertility and crop yields. However, manuring alone cannot fully account for the isotopic variation observed.
The second scenario is that the isozones are primarily natural; however the presence of multiple natural ecological isozones within specific areas of the Hampshire landscape is purely hypothetical. Nevertheless, the areas around Danebury and each of the Environs sites includes a variety of landscape properties that could result in multiple distinct ecological isozones within small areas. On the gentle slopes, the chalk bedrock (highly alkaline) is covered by relatively shallow soils. On the hilltops, the chalk is covered with clays containing flint (generally neutral or slightly acidic). The surrounding river valleys contain colluvial and alluvial deposits that in areas are thought to have been waterlogged for part of the year. To the south and east of the Environs in the Hampshire Basin, more sandy soils (generally acidic) overlie the upper and lower green sand bedrock. Such diversity in landscape types is seen within close proximity in other regions of the British Isles; however, only in regions where chalk is the underlying bedrock have such low nitrogen isotopic values been observed. Thus, it seems the combination of landscape properties in the Hampshire region may increase the isotopic range observed within the isotopic baseline of an area.
Whether the isozones within the Danebury Environs Landscape are natural or anthropogenic, for the observed pattern of isotope signatures to be produced, the movement of some of the horse, sheep and cattle has to have been restricted by humans. This is consistent with evidence from Iron Age Britain, which indicates that the husbandry regime may have included the management of animals through herding or corralling, indeed controlling of flocks or herds is often necessary for protecting arable crops. As noted above, the cattle and horse δ15N typically range from 1.5 to 6.5 ‰ whereas the sheep δ15N typically range from 3 to 6.5 ‰. Thus, no sheep had lifetime access to foodstuffs from isozones with low δ15N values whereas some cattle and horse individuals spent the majority of their lives consuming foodstuffs from such isozones. By contrast, some individuals from all of these species had lifetime access to foodstuffs from isozones with high δ15N values. If the isozones are anthropogenic, some cattle and horses would have to have only fed on non-manured contexts for the majority of their lives whereas other cattle and horses would have to have only fed on intensively manured contexts for the majority of their lives. Sheep, on the other hand, would have to have spent a significant proportion of their lives feeding on vegetation from manured contexts, with no individuals feeding solely on non-manured vegetation. If the isozones are primarily natural, the sheep would not have had access to one of the isozones. This scenario is consistent with the ecological requirements of the species under consideration. Cattle thrive in lush vegetation conditions and will consume as much as 16 gal of water per day (Green 1992), whereas sheep flourish on the relatively poor pasture of hillslopes and are susceptible to foot rot and liver fluke in damp conditions (Albarella 2007). Thus cattle are better able to subsist in the wetland conditions of river valleys that are less suitable for sheep. If cattle were kept in all areas of landscape, but sheep did not have access to the river valley pastures, this could explain the differences in the ranges of variation in cattle and sheep δ15N values.
This final model is consistent with the landscape system described in the Early Irish laws (Kelly 1997). The way in which people and their animals used the landscape is likely to have been articulated via systems of kinship and clientage. Land would be “owned” or held in common by social groups, probably kin-based, rather than individuals. Individuals and their animals would have rights of access to, or use of, certain parts of the landscape at certain times, which might be more or less co-ordinated with the activities of other individuals or families. Individual trajectories of landscape use could be quite variable over time, but might also be consistent from year to year, and animals could be controlled both by personal attendance and more or less temporary enclosure. While both people and their herds/flocks might gather in large groups at certain times of year, at other times they would be dispersed. The landscape could be seen as being made up of numerous interlocking activity-territories, defined in both space and time, and remembered and inherited. This could explain the apparent continuity of landscape use through the settlement changes of the third century BC, such that the Danebury Environs sites could be reoccupied at the same locations two centuries later. Periodic gatherings might occur based on farming activities—e.g. birth of lambs and calves, harvesting and crop-processing—or on a relative lack of agricultural activity giving people time to gather and renew social relationships, e.g. at a midwinter feast. This would allow for exchange at small and large scales, and such gatherings may well have been important components of the hillfort system of the Early and Middle Iron Age.
All the above models assume static agricultural/economic scenarios; however, they may have been dynamic, potentially hinted at by the minor isotopic trends observed in some species. In order to test the proposed models in a more dynamic system would require isotopic data taken from discrete time slices (e.g. ceramic phases). Currently, there is too little isotopic data to test different scenarios in this manner. A more in-depth isotopic investigation may allow links between settlement pattern dynamics and animal husbandry practices over the 500 year occupation to be explored.
Conclusions
We speculated that the large amount of intra-population isotopic variability observed by Stevens et al. (2010) in the faunal isotope signature at Danebury hillfort was due to the animals being husbanded at a number of sites within the Danebury Environs that had distinct ecological isozones, and that the animals were subsequently brought to the hillfort. Our isotopic results from the Danebury Environs sites demonstrate that this is not the case as they also exhibited extensive intra-site isotopic variability. The observed patterns of animal isotopic variation require considerable human management of herds or flocks, either through animal droving over long distances, or movement restriction, possibly through penning or corralling. One possible model that could explain the isotopic variation observed at Danebury and its Environs sites is that large numbers of animals were imported from distant regions that have different isotopic baseline values to Danebury and the Environs sites. The practical requirements of this model (i.e. long distance trade networks maintained in terms of proportion and source of animals over 500 years) make this model improbable. Rather, we propose a model that includes distinct ecological isozones within the Danebury Environs landscape, with some animals from each of the sites consuming foodstuff from different isozones for most of their lives. This interpretation of the isotope results is consistent with previous suggestions that earthwork features of Iron Age sites may have had a pastoral stock management role and have acted as stock enclosures (Fowler 1983; Davis 1987). The control of animal landscape use may be a reflection of the greater need for stock protection due to increasing social tension during the Iron Age or the greater need to fertilize soils (Cunliffe 1986). Further zooarchaeological and isotopic research may help determine which of the possible models is more likely. Irrespective of which model proves to be the most accurate, these results paint a picture of complex land management during the Iron Age.
References
Albarella U (2007) The end of the sheep Age: people and animals in the late iron Age. In: Haselgrove C, Moore T (eds) The late iron Age in Britain and beyond. Oxbow, Oxford, pp 393–406
Ambrose SH, Norr L (1993) Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate. In: Lambert JB, Grupe, G (eds), Prehistoric Human Bone—Archaeology at the Molecular Level. (Berlin) 1–37
Amundson R, Austin AT, Schuur EAG, Yoo K, Matzek V, Kendall C, Uebersax A, Brenner D, Baisden WT (2003) Global patterns of the isotopic composition of soil and plant nitrogen. Global Biogeochem Cycles 17(1):1031
Bendrey R, Hayes TE, Palmer MR (2009) Patterns of Iron Age horse supply: an analysis of strontium isotope ratios in teeth. Archaeometry 51(1):140–150
Bocherens H, Drucker D (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13:46–53
Bogaard A (2012) Middening and manuring in Neolithic Europe: issues of plausibility, intensity and archaeological method. In: Jones RL (ed) Manure: historical, archaeological and ethnographic perspectives. Ashgate, Farnham, pp 25–40
Campbell G (2000) Plant utilization: the evidence from charred plant remains. In: Cunliffe B (ed) The Danebury environs programme volume 1: introduction. English Heritage and Oxford University Committee for Archaeology, Monograph 48, Institute of Archaeology, Oxford, pp 45–59
Cunliffe B (1984a) Danebury: An Iron Age Hillfort in Hampshire. Vol. 2 The Excavations 1969–1978: The Finds. CBA Research Report No. 52b, London
Cunliffe B (1984b) Danebury: An Iron Age Hillfort in Hampshire. Vol. 1 The Excavations 1969–1978: The Site. CBA Research Report No. 52a, London
Cunliffe B (1993) Danebury. English Heritage, London
Cunliffe B (1995) Danebury: An Iron Age Hillfort in Hampshire. Vol. 6. A Hillfort Community in Perspective. CBA Research Report No. 102, York
Cunliffe B (1986) Danebury: anatomy of an Iron Age hillfort. Batsford, London
Cunliffe B (2000) The Danebury environs programme: the prehistory of a Wessex landscape: volume 1—introduction. English Heritage and Oxford University Committee for Archaeology, Oxford
Cunliffe B, Poole C (2000a) The Danebury environs programme: the prehistory of a Wessex landscape: volume 2—part 2: Bury Hill, Upper Clatford, Hants, 1990. English Heritage and Oxford University Committee for Archaeology, Oxford
Cunliffe B, Poole C (2000b) The Danebury environs programme: the prehistory of a Wessex landscape: volume 2—part 3: Suddern Farm, Middle Wallop, Hants, 1991 and 1996. English Heritage and Oxford University Committee for Archaeology, Oxford
Cunliffe B, Poole C (2000c) The Danebury environs programme: the prehistory of a Wessex landscape: volume 2—part 4: New Buildings, Longstock, Hants, 1992 and Fiveways, Longstock, Hants, 1996. English Heritage and Oxford University Committee for Archaeology, Oxford
Cunliffe B, Poole C (2000d) The Danebury environs programme: the prehistory of a Wessex landscape: volume 2—part 5: Nettlebank Copse, Wherwell, Hants, 1993. English Heritage and Oxford University Committee for Archaeology, Oxford
Cunliffe B, Poole C (2000e) The Danebury environs programme: the prehistory of a Wessex landscape: volume 2—part 6: Houghton Down, Stockbridge, Hants, 1994. English Heritage and Oxford University Committee for Archaeology, Oxford
Davis SJM (1987) The archaeology of animals. Batsford, London
Dawson TE, Mambelli S, Plamboeck AH, Templer PH, Tu KP (2002) Stable isotopes in plant ecology. Annu Rev Ecol Syst 33:507–559
DeNiro MJ (1985) Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317:806–809
Fowler P (1983) The farming of prehistoric Britain. Cambridge University Press, Cambridge
Fraser R, Bogaard A, Heaton T, Charles M, Jones G, Christensen BT, Halstead P, Merbach I, Poulton PR, Sparkes D, Styring A (2011) Manuring and stable nitrogen isotope ratios in cereals and pulses: towards a new archaeobotanical approach to the inference of land use and dietary practices. J Archaeol Sci 38:2790–2804
Grant A (1984) Animal husbandry. In Cunliffe B (ed) Danebury: An Iron Age Hillfort in Hampshire. Vol. 2 The Excavations 1969–1978: the Finds. CBA Research Report No. 52b, London
Grant A. (1991) Animal husbandry. In Cunliffe B, Poole C (eds) Danebury: An Iron Age Hillfort in Hampshire. Vol. 5 The Excavations 1979–1988: the Finds. CBA Research Report No. 73b, London
Green M (1992) Animals in Celtic life and myth. Routledge, London, pp 5–21
Hambleton E (2008) Review of Middle Bronze Age—Late Iron Age faunal assemblages from Southern Britain. English Heritage Research Department Report Series 71.
Hamilton J (2000) Animal husbandry: the evidence from the animal bones. In: Cunliffe B (ed) The Danebury Environs programme volume 1: introduction. English Heritage and Oxford University Committee for Archaeology Monograph 48, Oxford, pp 59–76
Heaton THE (1999) Spatial, species, and temporal variations in the 13C/12C ratios of C3 plants: implications for palaeodiet studies. J Archaeol Sci 26:637–649
Hedges REM, Reynard LM (2007) Nitrogen isotopes and the trophic level of humans in archaeology. J Archaeol Sci 34:1240–1251
Hoefs J (1997) Stable isotope geochemistry. Springer, Berlin
Jay M, Richards MP (2007) British iron Age diet: stable isotopes and other evidence. Proc Prehist Soc 73:171–192
Jones J, Mulville J, McGill R, Evershed RJ (2012) Palaeoenvironmental modelling of δ13C and δ15N in the North Atlantic Islands: understanding past marine resource use. Rapid Commun Mass Sp 26(20):2399–2406
Jones MK (1984) The plant remains. In: Cunliffe B (ed) Danebury: An Iron Age Hillfort in Hampshire. The Excavations 1969–1978: The Finds. CBA Research Report No 52b, vol. 2. London, pp. 483–495
Jones MK, Nye S (1991) The plant remains. In: Cunliffe B, Poole C (eds) Danebury: An Iron Age Hillfort in Hampshire. The Excavations 1979–1988: TheFinds. CBA Research Report No 73b, vol. 5. London, pp. 439–447
Jones MK (1995) Patterns in agricultural practice: the archaeobotany of Danebury in its wider context. In: Cunliffe B. (ed) Danebury: An Iron Age Hillfort in Hampshire. A Hillfort Community in Perspective. CBA Research Report No 102, vol.6. London, pp. 43–50
Kelly F (1997) Early Irish farming. School of Celtic studies, Dublin Institute for Advanced Studies
Lightfoot E, Stevens RE (2012) Stable isotope investigations of charred barley (Hordeum vulgare) and wheat (Triticum spelta) grains from Danebury Hillfort: implications for palaeodietary reconstructions. J Archaeol Sci 39(3):656–662
Madgwick R, Mulville J, Stevens RE (2013) Isotopic analysis of fauna from British middens of the Late Bronze Age: Evidence for diversity in foddering strategy and herd management. Environmental Archaeology. In press
Millard AR, Jimenez-Cano NG, Lebrasseur O, Sakai Y (2011) Isotopic investigation of animal husbandry in the welsh and English periods at dryslwyn castle, Carmarthenshire, Wales. Int J Osteoarchaeol. doi:10.1002/oa.1292
Mulville J, Madgwick R, Stevens RE, O’Connell T, Craig O, Powell A, Parker Pearson M, Sharples NM (2009) Isotopic analysis of faunal material from South Uist, Western Isles, Scotland. J North Atl 2:51–59
Privat KL, O’Connell TC, Richards MP (2002) Stable isotope analysis of human and faunal remains from the Anglo-Saxon cemetery at Berinsfield, Oxfordshire: dietary and social implications. J Archaeol Sci 29:779–790
Schoeninger MJ, DeNiro MJ, Tauber H (1983) Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial components of prehistoric human diet. Science 220:1381–1383
Stevens RE, Hedges REM (2004) Carbon and nitrogen stable isotope analysis of northwest European horse bone and tooth collagen, 40,000 BP–present: palaeoclimatic interpretations. Quaternary Sci Rev 23(7–8):977–991
Stevens RE, Jacobi RM, Street M, Germonpré M, Conard NJ, Műnzel SC, Hedges REM (2008) Nitrogen isotope analyses of reindeer (Rangifer tarandus), 45,000 BP to 9,000 BP: Palaeoenvironmental reconstructions. Palaeogeogr Palaeoclimatol Palaeoecol 262(1–2):32–45
Stevens RE, Lightfoot E, Hamilton J, Cunliffe B, Hedges REM (2010) Stable isotope investigations of the Danebury hill fort pit burials. Oxford J Archaeol 29:407–428
Stevens RE, Lightfoot E, Hamilton J, Cunliffe B, Hedges REM (2013) Investigating dietary variation with burial ritual in Iron Age Hampshire: an isotopic comparison of Suddern Farm Cemetery and Danebury hillfort pit burials. Oxford J Archaeol. In press
Tieszen LL, Fagre T (1993) Effect of diet quality and composition on the isotopic composition of respiratory CO2, bone collagen, bioapatite, and soft tissues. In: Lambert JB, Grupe G (eds) Prehistoric human bone—archaeology at the molecular level. Springer Berlin pp.121–155.
Ubelaker DH, Katzenberg MA, Doyon LG (1995) Status and diet in pre-contact highland Ecuador. Am J Phys Anthropol 97:403–412
Ugan A, Coltrain J (2011) Variation in collagen stable nitrogen values in black-tailed jackrabbits (Lepus californicus) in relation to small-scale differences in climate, soil, and topography. J Archaeol Sci 38(7):1417–1429
Vogel JC, van der Merwe NJ (1977) Isotopic evidence for early maize cultivation in New York State. Am Antiq 42:238–242
Acknowledgments
The authors would like to thank Kay Ainsworth at Hampshire Museum stores for providing access to the material, records and unpublished information. Peter Ditchfield is thanked for help with mass spectrometry. Jacqui Mulville is thanked for useful discussions. EL would like to thank Darwin College for financial support; RS would like to thank the Royal Society and NERC for financial support.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Online supplementary dataset 1
Sample provenance, isotope results and atomic C/N ratios. (XLS 91 kb)
Rights and permissions
About this article
Cite this article
Stevens, R.E., Lightfoot, E., Hamilton, J. et al. One for the master and one for the dame: stable isotope investigations of Iron Age animal husbandry in the Danebury Environs. Archaeol Anthropol Sci 5, 95–109 (2013). https://doi.org/10.1007/s12520-012-0114-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-012-0114-3