
Abstract
Japanese rice cultivation in paddy fields has 2,400∼3,000 years of history. Most of modern Japanese rice varieties are classified as Temperate-japonica (Tm-J). Few landraces are recognized as Tropical-japonica (Tr-J) only in southwestern Japan. However, ancient DNA studies and phytolith analysis suggest that Tr-J strains were more popular in the past than now. Maekawa is a complex archaeological site composed of paddies dated from the Yayoi (2,100 years BP) to the Heian (1,100 years BP) periods. Phytolith analysis indicates that intensive rice cultivation was practiced in both periods, but there was no cultivation in the intervening period. Morphological features of bulliform phytoliths suggest that Tr-J was cultivated during both periods. Locally, rice cultivation during the Heian period was brought to a close by a flood event, in which immature rice plants were pulled down and buried in silt to be preserved in a quasi-carbonized/ waterlogged state. Ancient DNA (aDNA) analysis of the carbonized plant culm from Heian Maekawa recovered chloroplast DNA sequences of the 6C7A plastid subtype, which is common to both Tr-J and Tm-J, whereas two plastid subtypes, such as 6C7A and also 7C6A, were found in aDNA of carbonized grains from the Tareyanagi site of the Yayoi period. The latter plastid subtype was specific only to Tr-J. In order to better characterize the past rice populations, modern landraces collected in the local area were classified with morphophysiological traits. Some of the landraces were found to carry several traits of Tr-J, including bulliform phytolith types, but mixed with Tm-J traits. Based on the discontinuous distribution of rice phytoliths between the Yayoi and the Heian period, the early introduction of rice cultivation may have been discontinuous and locally reintroduced after a ∼1,000-year hiatus, but with a genetically different rice population. Such populations were composed from Tr-J like strains as shown by landraces but with reduced diversity in plastid types. Through such changes, since the Yayoi era, Tr-J was largely replaced by Tm-J, although ancient Tr-J continued to participate in the genetic makeup of later rice populations and may have aided the local adaptation of introduced Tm-J.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Rice is one of the important cereals in East Asia. Archaeological rice remains and sometimes accompanying fields have been recorded in back to 4,000–5,000 BC in the Middle and Lower Yangtze region of China (see Nakamura 2010; Zhao 2010; Fuller and Qin 2009). While there is a long record of rice use, including wild rice, in China, indicated by Late Pleistocene sites such as Xianrendong, by contrast, there were no wild rice populations in Japan. People living in Japan during the Jōmon periods (from about 12,800 BC up to 300 BC) based their subsistence foods mainly on various wild sources including chestnuts, fishes, berries, and so on, together with some cultivation of vegetables, beans, and millets in some areas in the Later Jōmon, but without wild rice use (Habu 2004). Rice cultivated in irrigated paddy systems is probably introduced around 900 BC into south/west Japan (Takahashi 2009). Recent data, especially early AMS dates on rice, have created an area of active debate about chronological boundaries between the Jōmon and the Yayoi periods, whether in the earlier or later First Millennium BC (e.g., Keally 2004).
Rice, with evidence for paddy field agriculture, was widespread in Japan by approximately 400 BC, having reached the most northerly areas of Honshu Island. The Tareyanagi site, the Takahi (III) site (belonging to the same culture as Tareyanagi), and the Maekawa site all produced evidence for paddies dated to the Yayoi period (Agency of Education of Inakadate Village 2002; Agency of Education of Aomori prefecture 1984; Agency of Education of Aomori Prefecture 2009). Maekawa also preserved paddies from the Heian period. However, such rice cultivation can be inferred to have not succeeded in persisting continuously in this area from the Yayoi period. Sakaguchi (1983) suggested fluctuating paleoclimate intervals of warm and cool periods based on pollen analysis. It was implied that such intervals would influence agricultural activity severely in the northern areas. Indeed, there was found to be a lack of rice cultivation in cool periods and more activity in warm periods in this region. For example, there are few paddies dated in the Kofun (1760 to 1268 BP) in this area. Paddies reappeared again at the Heian period (794 BP to 1192 BP). By contrast, rice cultivation was continuously performed in the southern areas of Japan. The reasons for an apparent suspension of rice cultivation on northerly Honshu Island between the Yayoi and the Heian periods is still an open question.
Another question concerns which kinds of rice varieties were cultivated, and whether these were the same or different over time? Given a history of rice paddy agriculture of nearly 3,000 years, which might ask whether there has been selection for particular rice varieties, Tropical-japonica (Tr-J) is one candidate varietal group, based on previous ancient DNA research, in which excavated ancient rice grains were used for amplification of chloroplast (plastid) DNA (cpDNA) by polymerase chain reaction (PCR) technology to differentiate japonica and indica rice subspecies (Sato 1996). However, most modern varieties in Japan are classified as Temperate japonica (Tm-J). In this paper, we report the amplification of ancient DNA from carbonized plants dated to around 1000 BP. Then, morphological and physiological characteristics of modern local genetic resources were examined in order to assess which lineages contributed some genetic components to rice populations cultivated in the northern parts of Japan.
Study area
The Maekawa site belongs to the Tsugaru area, where rice cultivation is still popular at present. Water is supplied from the Hakkoda mountain areas. The water flow often causes sudden floods of the alluvial plain. Thus, paddies at the Maekawa site were also damaged by floods. Within 50-km distance, there is the Sannai-Maruyama, a famous Jōmon settlement that prospered from 5900 BP to 3500 BP (Habu 2008; Kawahata et al. 2009). In general, subsistence of the period is regarded as hunting and gathering. Several evidences, however, suggest that a food resource management system had been developed to control chestnut trees, wild grape, and millets, especially Echinochloa crus-galli (Habu 2004; Sato and Ishikawa 2004; Crawford 2008). The ancient city collapsed due to unknown reasons, but a climatic transition from warm to cold conditions may have contributed (Sakaguchi 1983; Kawahata et al. 2009). Rice cultivation had not reached this site/region by that time. Subsequently, there was a gap from approximately 3500 BP when the Sannai villages were abandoned, but before rice agriculture was in evidence, to approximately 2400 BP when rice cultivation became popular in the northern area. Elsewhere in the region, the earliest possible evidence for introduced millet (Setaria italica) and rice cultivation is dated at the Kazahari site from approximately 900 BC (D'Andrea et al. 1995), although debates persist about determining the timing and process of rice introduction. One of our goals in this study was to investigate how rice cultivation with paddy systems was introduced over time.
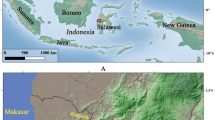
The Maekawa site is composed of the Yayoi paddies, the Heian paddies, and houses for the middle ages (Fig. 1). Several sections were divided into smaller areas as blocks to excavate. Dates of these paddy fields were determined with radiocarbon dates on carbonized fragments reported on the Maekawa site by Agency of Education of Aomori prefecture (2009). Among four sections (sections A to D), layer IV in section B was dated to approximately 650–710 cal. AD (50.0%) or 740–770 cal. AD (18.2%) (IAAA-71833, 1320 ± 30 BP). There were numerous remains of whole rice plants oriented towards the downstream direction, probably pulled down by a sudden flood in Heian times (Fig. 1c, d). Numerous human footprints were also covered over with silts by the flood (Fig. 1b). These plants were used to amplify DNA in this study. The dating is confirmed by a volcanic tephra caused by the eruption known as “Towada A tephra,” dating to 915 AD (Machida and Arai 2003). In section C1, there were other paddy fields at layer VII. A radiocarbon date on carbonized material from in the layer was dated 370–340 cal. BC (8.0%) or 320–200 cal. BC (60.2%) (IAAA-71827, 2210 ± 30 years BP) (Agency of Education of Aomori prefecture 2009). Other materials were also dated around the Yayoi period. Based on the dated materials, the paddies were determined to originate in the Yayoi period. Close to the Maekawa site, the Takahi (III) and Tareyanagi sites are also well known for archaeologically preserved paddies dating to the Yayoi period (Agency of Education of Inakadate village 2002; Agency of Education of Aomori prefecture 1984).

The Maekawa site which preserves Heian-aged paddies. a Locations of the Maekawa site, Tareyanagi site, Takahi (III) site, Sannai-Maruyama site, and Kazahari site. b Footprints preserved in the paddy surface, from the middle-aged paddies, which were covered with flood silts. c Plant remains pulled down by a flood silt, about 1,000 years ago. d Paddy fields dated to 1330 BP
Materials and methods
Soil samples
From Yayoi period paddy paleosols, multiple samples were taken from localities A to D (blocks A to D; Table 1). Twenty samples from A block and ten of each other block were used for phytolith analysis. Multiple samples were also prepared for 14 Heian period paddies (no. 3, 4, 5, 6, 7, 8, 10, 30, 31, 32, 33, 34, 46, and 47; following the numbering system in the Maekawa site report; Agency of Education of Aomori prefecture 2009). Replicate numbers were shown in Table 2. Ten samples from quasi-carbonized/ waterlogged plants sealed in layer VI, section B, dated from the Heian period, were subjected to aDNA investigation in order to confirm which plant species they originated from.
Phytoliths
Quantitative analysis of phytoliths was carried out following the method of (Fujiwara et al. 1987, 1993) This procedure included six steps: (1) soil samples were collected from site; (2) sample was dried at 105°C for 24 h; (3) a fixed amount of microscopic glass beads were added to the sample; (4) samples were diffused with ultrasonic waves (300 W, 42 kHz, 10 min); (5) samples were sieved to remove particles under 20 μm; (6) samples were pipetted onto slides and examined with transmitted light at magnifications of ×400.
Bulliform phytoliths, derived from silicification of the motor cells in the leaf blades of grasses (family Poaceae), were counted by microscopic examination (×400 magnification) alongside the counting of glass beads. Phytoliths and beads were counted until the number of glass beads reached up to around 500. As the number of glass beads added in the sample was known, this allowed for an accurate estimate of the density of bulliform phytoliths in the sample.
Ancient DNA extraction
DNA extraction was carried out to target DNA in organics trapped inside silicified cells (phytoliths). This procedure involved removing DNA that might contaminate the sample, followed by breaking open the phytoliths and extracting the included DNA. Ancient DNA extraction was carried out in a purpose-built lab, dedicated to ancient DNA work at the Research Institute for Humanity and Nature (RIHN), Kyoto, Japan. Samples were first treated to prevent the contamination of results by microbial DNA. This involved taking 20-mg samples of the ancient rice, which were softly ground and then allowed to stand at 37°C for 2 h after being mixed with the following reagents: 500 μl G buffer (0.3 M sucrose, 50 mM Tris–HCl (pH 7.5), 0.1 mM ethylenediaminetetraacetic acid (EDTA), 0.05% BSA, and 10 mM MgCl2) and 50 μg DNase I (Sigma Co.). Then, in order to remove the digested DNA and DNase, a Shelf buffer (0.6 M sucrose, 10 mM Tris–HCl, 20 mM EDTA) was added (on shelf buffer, see Umbeck and Gengenback 1983). After centrifugation (10,000×g, 20 min), the pellet was resuspended with an equal amount of the Shelf buffer again. After the solution was mixed, it was centrifuged and supernatant was removed again. All nonchloroplast DNA should now have been removed.
The following steps were used to amplify the archaeological rice DNA. Each sample was added with 20 μl of extraction buffer A (1 M NaOH, 50 mM Tris–HCl). As the samples were presumed to consist from nuclei included within phytoliths, the solution of each sample was treated with Multi-bead shocker (Yasui Kikai Co. Ltd.) to break and homogenize the phytoliths. Then, to each solution was added 380 μl of extraction buffer B (0.1 M Tris–HCl, pH 8.0). Mixed samples were treated at 65°C for 10 min and centrifuged at 2,000 rpm for 10 min. Three hundred microliters of the supernatant was transferred into a new tube, to which was added 900 μl of 100% ethanol. The mixture was stirred and centrifuged at 15,000 rpm for 10 min. The supernatant was transferred to a new tube to which was added 30 μl 3 M sodium acetate and 3 μl Ethachinmate (Nippongene Co., Japan). The mixture was again centrifuged, at 15,000 rpm, for 10 minutes, and supernatant was drained. Debris was added to 300 μl of 70% ethanol and centrifuged at 15,000 rpm for 10 min, and then the supernatant was drained again. The debris was added with 20 μl TE and treated at 65°C for 10 min. The mixture was centrifuged at 15,000 rpm for 10 min and purified with ProbeQuant G-50 Micro Columns (GE Co., Japan). A solution was used for enzyme repair, PreCR™ repair mix (NEB, Japan) based on the company's recommended protocol.
The targeted DNA site was parts of rpl16–rpl14 cluster in the chloroplast DNA. Two succeeding, nested PCRs were performed to amplify enough DNA. The first primer pair was a PS-ID A primer sequence (bases = aaagaaccagatttcgtaaacaacatagaggaagaa) and a PS-ID B primer (bases = atctgctacatttaaaagggtctgaggttgaatcat). As the second primer pair, PS-ID 2F (bases = caatttcttcggttagaaata) and PS-ID 2R (bases = gaaagaaatattgtctttccag) were designed from the rpl16–rpl14 gene cluster in chloroplast DNA (Fig. 2). PCR conditions were 95°C for 1 min as preheating, 40 rounds of 95°C for 10 s, 58°C for 30 s, and 72°C for 10 s, followed by 72°C for 1 min with exTaq polymerase (Takara Co., Japan). For the second PCR reaction, total volume of the first PCR solution was added to a new solution, and the same timing and temperature conditions were used.

Amplified DNA that is part of the rpl16–rpl14 cluster in the chloroplast genome. a Primer distribution along the targeted sequence. b Amplified sequences in this study, compared with those from the Takahi (III) site close to Maekawa. These sequences obtained at the Takahi (III) originated from carbonized grains dated as the Yayoi period
Discriminating Tropical-japonica and Temperate-japonica with morphophysiological characters
Local rice landraces were collected in the Tsugaru area. Morphophysiological characteristics were characterized within these 15 landraces (Akaiga, Iwaka, Shiroshine, Akamoro, Akamoro-K, Akamoro-Kuroishi, Akamoro-M, Shiromoro, Akamochi, Akamochi2, Shijyuunichi-wase, Hosogawa, Kuromochi, Somewake, Derokowase), with Nipponbare as a control example (prototype) of Tm-J and two strains from the Philippines (ACC220 and ACC221) as prototypes of Tr-J. They were discriminated into Tr-J or Tm-J with morphophysiological characters by the following formula (Sato 1987):
in which measurements include mesocotyl length (M; mm), ratio of hull length/width (R; L/W), and endosperm alkaline digestion (E; percentage for remain of weight per total weight after 1.5% KOH treatment). Strains with Z2 scores under 0 correspond to Tm-J and those above 0 to Tr-J.
Results
Phytolith analysis
Phytoliths were tabulated as morphotypes. Some morphotypes were assigned taxonomic identification to several taxa, or probable taxa, including Oryza, Echinochloa, Paniceae, Phragmites, Miscanthus, Pleioblastus, and Sasa. Other phytolith morphotypes were grouped into other shape categories. The number of these phytoliths was quantified for each layer to provide some proxy for the relative frequency of the various plant types in each layer. In addition, the density of rice phytoliths was calculated for soil samples from ancient paddy paleosols, which were dated to the Yayoi period based on radiocarbon dates. Ten samples were counted at blocks A, B, C, and D (Table 1). The densities ranged from 1,200 to 6,000 pieces per gram at block A (average 4,000 pieces per gram), from 3,000 to 10,200 pieces per gram at block B (average 5,900 pieces per gram), from 1,200 to 7,300 pieces per gram at block C (average 4,100 pieces per gram), and from 600 to 2,400 pieces per gram at block D (average 1,100 pieces per gram). As a general standard, rice phytolith density from paddy fields is expected to be 5,000 pieces per gram. (Fujiwara et al. 1987; Fujiwara 1993); this is defined as the standard density for a rice paddy field. Block B represented a higher density of rice phytoliths than the standard density, while blocks A and C were close to the standard density.
All paleosols in layer IV, section B included rice phytoliths. Averaged concentrations of rice phytoliths ranged from 4,900 pieces per gram at “Paddy 32” to 15,500 pieces per gram at “Paddy 10” (Table 2). Other plant phytolith categories, originating from Echinochloa, Paniceae, Phragmites, Miscanthus, and Bambusoideae were relatively rare. Relatively higher density was observed for Sasa sp., which recorded densities from 2,700 to 6,400 pieces per gram.
In “Paddy 20,” plant macroremains were also recognized (Fig. 1). These appeared black as though carbonized. These were remains of intact (articulated) plant leaf and culm that appeared to be horizontally aligned in alluvial silts, suggesting that they had been pulled in the downstream direction by a flood event but were still in situ in the field. These plants were fairly evenly spaced, suggesting that they had been planted at regular intervals. Samples from the remains showed high density of rice phytoliths ranging from 15,700 to 24,000 pieces per gram (Table 3). The average density, 13,300 pieces per gram, was apparently higher than those of other taxa from the same samples, which included Paniceae, Phragmites, and Miscanthus, which ranged from 0 to 1,200 pieces per gram in single samples. We interpret this as confirming that these were primarily the remains of rice plants, with a minor contamination from the sediment of other contemporary, nearby plants and weeds, represented by the other types of phytoliths.
DNA analysis
The preserved plants of rice, just described above, came from “Paddy 20” of the Heian period (approximately 900 AD). An ancient DNA extraction was carried out on ten soil samples which included the preserved plant bodies, following the method outlined above, which was designed to extract the nuclear DNA that is presumed to be trapped within the phytoliths within the plant body. PCR amplification was confirmed with six out of the ten samples, i.e., in samples 2, 4, 7, 8, 9, and 10 (Fig. 3). Additional bands other than the expected one were seen on gels, but similar bands occurred in the positive control represented but not in the negative control, and therefore these DNA bands may be interpreted as artifacts derived from the combination of various smaller amplified products, i.e., “chimera” DNA. Further analyses were not carried out on such bands. Amplified bands in expected bands can be presumed to originate from plants rather than microorganisms. These bands, of targeted fragment length, were sliced off from the gels. DNA was purified and sequenced (as described in “Materials and methods”). All amplified samples showed the 6C7A chloroplast plastid type, which is the type common to most of Tm-J and parts of Tr-J.

Amplified DNA from plant remains. M1 and M2 were size markers, 100-bp ladders and 20-bp ladders (Biolabs, Co. USA). Ct is the positive control amplified product from genomic DNA of rice plant. Ne is the negative control with water. No. 1 to 10 were samples. a First electrophoresis. b Second electrophoresis, in which amplified fragments are observed clearly
Paleo-Tropical-japonica in landraces around the Maekawa site
Local landraces which had been cultivated in this area were characterized with methods based on morphophysiological characters. All 15 landraces represented the 6C7A plastid subtype rather than 7C6A which predominates in Tr-J. This genetic characterization indicated a japonica identification but does not determine whether the landraces are in the tropical or temperate subgroups (Table 4).
In several other morphophysiological characters, a Tm-J identification was suggested. The percentage of undigested endosperm after alkaline treatment is one of the characteristics to determine Tr-J: Tr-J strains tend to show hardness for the digestion. For example, landrace numbers 220 and 221, as control Tr-J strains, presented 2.8% and 1.5%, respectively. Nipponbare, as the control Tm-J strain, presented 31.0%. Digested percentage of local landraces ranged from 72.4% of “Akamochi” to 18.9% of “Akamoro.” Mean of the percentage was 42.6 ± 13.4%. Also, the seed shape suggested Tm-J. Length-to-width ratio ranged from 1.84 to 2.06 among local landraces. These ratios were not different from general Japanese varieties. “Nipponbare” has 1.72 but is much lower than standard Tr-J strains (represented by ACC220 and ACC221, which showed 2.97 and 2.34, respectively).
However, a few other traits suggested some presence of Tr-J genetic material in a few landraces. One difference between Tm-J and Tr-J is the elongation ability of the mesocotyl under dark conditions. In general, Tm-J shows very little elongation. For example, our representative “Nipponbare” elongated 3.2 mm. In contrast, Tr-J representatives ACC220 and ACC221 showed 13.4 and 21.2 mm, respectively. In two local landraces, relatively longer mesocotyl elongation was noted: in “Shiromoro” it was 39 mm, while “Akamoro” also showed 25.2 mm. The mean was 10.23 ± 11.0 mm among all local strains used. Landraces were, then, examined for morphophysiological traits to calculate Z2 score to discriminate them into Tr-J or Tm-J. “Nipponbare” and Tr-J showed negative score and positive scores, respectively. Those of the landraces ranged from −2.31 to 2.49. The maximum scores, suggesting classification with Tr-J, were found in “Shiromoro” followed by “Akamoro.”
Discussion
Tropical (Tr-J) and temperate (Tm-J) subgroups of japonica subspecies of rice have been separated morphologically and physiologically for a long time (Oka 1958). Temperate-japonica predominates in current landraces in Japan (Ishikawa et al. 1992), although few landraces were recognized morphologically as Tr-J (Sato 1992). Based on aDNA of archaeological rice grains, a few represent Tr-J at molecular level (Sato 2002). For example, Takahi (III) site, which is close to the Maekawa site and Tareyanagi site, had both Tr-J plastid type and Tm-J plastid type. Generally, however, Tr-J is rare in Japanese landraces, especially as one moves north. As far north as central Japan, Tr-J could be recognized in a few landraces but it is more common in southwestern Japan, but generally in lowlands rather than in hills. Sato (1992) suggested that ancient outcrossing between Tr-J and Tm-J generated highly adapted descendants. However, subsequent rice populations (including modern landraces) have included more Tm-J. More pure Tr-J are therefore rare and have been missed in modern rice breeding programs in Japan. In addition, recent selection has focused on substituting two major Tm-J cultivars, “Aikoku” and “Shinriki,” for many diverse landraces. These two cultivars have been used to create most modern Japanese breeding lineages.
Based on amplified DNA fragments from the archaeological materials from the Takahi (III) site dated to the Yayoi period, two plastid types, 6C7A and 7C6A, were identified (Sato 2002). The latter is attributed to Tr-J origin because paleoclimatic proxy data suggest that the Yayoi climate was warmer than modern times and allowed people to adopt rice cultivation even in the most northerly areas of Honshu island. After that period, ancient paddies disappeared in the northern region until the Heian period, equivalent to another warmer period. The Maekawa site suggests such an interruption based on the presence and absence of rice phytoliths (Tanaka et al. 2009; this paper). More systematically cultivated paddy fields were created at the same site once again in the Heian period. During a sudden flash flood, water and silt covered over the fields, and plants in the paddies were pushed down. They were blackened, perhaps partly carbonized, and contained articulated rice phytoliths. Phytolith analysis indicated that the plants were rice, and the fields were paddies based on the density of rice bulliforms. Thus, DNA amplification was tried, targeting the occluded nuclei in the phytoliths contained in the waterlogged, quasi-carbonized plants. The 7C6A plastid type was not recognized although sequences were obtained from multiple samples.
Rice phytoliths gave us evidence from which to infer which kinds of rice was cultivated, whether tropical or temperate japonica. Morphological characters indicated that relatively larger-size phytolith was found both in the paddies dated as the Yayoi period and also the Heian period, and these suggest Tr-J (Tanaka et al. 2009). We therefore infer that Tr-J, carrying the 6C7A, plastid type was cultivated at 2400 BP and 1,000 years later. After that, although we do not know exactly when, Tm-J was substituted in this region. Although hybridization with the older Tr-J forms is likely.
We attempted to confirm the presence of older genes from the Tr-J landraces through morphological analyses of modern local landraces. All modern landraces represented typical Tm-J in many traits, like grain size and shape. However, hardness of endosperm digested with alkaline solution and/or ability of elongation at the first internode (mesocotyl) under completely dark conditions suggested traits like those typical of upland rice and Tr-J. Hardness of the endosperm is controlled by a SSIIa gene related to soluble starch synthase (Umemoto et al. 2002). The hardness is regarded as a nonpreferred character in starch quality for modern Japanese consumers. Morphology of rice phytolith was another characteristic which linked typical Tr-J with some of the landraces in Tsugaru area. These observations support the conclusion that some of the Tr-J genetic compositions have remained in local landraces even in northeastern Japan. This suggests that there was an original Tropical-japonica substratum of rice that has been mostly replaced by temperate-japonica types introduced in the past millennium. Changes in the local rice variety may have been promoted by climatic change and or preferences for cooking traits and taste. This highlights that an appreciation of the history of rice in northern Japan, near the northern limits of this crop, requires more than just dating the earliest occurrence of rice but also a long-term perspective on local crop adaptation and the introduction of later landraces.
References
Agency of Education of Aomori prefecture (1984) Tareyanagi site. Agency of Education of Aomori prefecture, Aomori, in Japanese
Agency of Education of Aomori prefecture (2009) Maekawa site, vol 2. Agency of Education of Aomori prefecture, Aomori, in Japanese
Agency of Education of Inakadate village (2002) Takahi (3) site excavation report. Agency of Education of Inakadate village, Aomori, in Japanese
Crawford GW (2008) The Jōmon in early agriculture discourse: issues arising from Matsui, Kenehara and Pearson. World Archaeol 40(4):445–465
D'Andrea AC, Crawford GW, Yoshizaki M, Kudo T (1995) Late Jōmon cultigens in northeastern Japan. Antiquity 69:146–152
Fujiwara H (1993) Research into the history of rice cultivation using plant opal analysis. In: Pearsall DM, Piperno DR (eds) Current research in phytolith analysis: applications in archaeology and paleoecology. MASCA research papers in archaeology, vol 10. The University of Pennsylvania Museum, Philadelphia, pp 147–159
Fujiwara H, Matsuda R, Sugiyama S (1987) The experimental study on the estimation of the boundaries of the paddy fields and the changes in the Tareyanagi site in Aomori Pref. Archaeol Nat Sci 22:29–41, in Japanese
Fuller DQ, Qin L (2009) Water management and labour in the origins and dispersal of Asian rice. World Archaeol 41:88–111
Habu J (2004) Ancient Jōmon of Japan. Cambridge University Press, Cambridge
Habu J (2008) Growth and decline in complex hunter–gatherer societies: a case study from the Jōmon period Sannai Maruyama site, Japan. Antiquity 82:571–584
Ishikawa R, Maeda K, Harada T, Niizeki M, Saito K (1992) Genotypic variation for 17 isozyme genes among Japanese upland varieties in rice. Japan J Breed 42:737–746
Kawahata H, Yamamoto H, Ohkushi K-I, Yokoyama Y, Kimoto K, Ohshima H, Matsuzakai H (2009) Changes of environments and human activity at the Sannai-Maruyama ruins in Japan during the mid-Holocene Hypsithermal climatic interval. Quatern Sci Rev 28:964–974
Keally CT (2004) Bad science and the distortion of history: radiocarbon dating in Japanese archaeology. Sophia Int Rev 26:1–16, http://www.t-net.ne.jp/∼keally/Reports/sir2004.html. Accessed 30 March 2010
Machida H, Arai F (2003) Atlas of tephra in and around Japan. Tokyo University Press, Tokyo, in Japanese
Nakamura S (2010) The origin of rice cultivation in the lower Yangtze Region, China. Archaeological and Anthropological Sciences 2(2) (this issue)
Oka H-I (1958) Intervarietal variation and classification of cultivated rice. Indian J Genet Plant Breed 18:79–89
Sakaguchi Y (1983) Warm and cold stages in the past 7600 years in Japan and their global correlation—especially on climatic impacts to the global sea level changes and the ancient Japanese history. Bull Depart Geogr Univ Tokyo 15:1–31, In Japanese
Sato Y-I (1987) Variation in spikelet shape of the indica and japonica rice cultivars in Asian origin. Jpn J Breed 41:121–134, in Japanese
Sato Y-I (1992) The road by which rice was introduced. Shoukabou, Tokyo, In Japanese
Sato Y-I (1996) Civilization based on rice domestication inferred from DNA. NHK, Tokyo, in Japanese
Sato Y-I (2002) History of Japanese rice. Kadokawa, Tokyo, in Japanese
Sato Y-I, Ishikawa R (2004) Plant world in Sannai-Maruyama site. SHOKABO, Tokyo, in Japanese
Takahashi R (2009) Symbiotic relations between paddy-field cultivators and hunter–gatherer–fishers in Japanese prehistory: archaeological considerations of the transition from the Jōmon Age to Yayoi Age. In: Ikeya K, Ogawa H, Mitchell P (eds) Interactions between hunter–gatherers and farmers: from prehistory to present. Senri ethnological studies, 73. National Museum of Ethnology, Osaka, pp 71–98
Tanaka K, Udatsu T, Ishikawa R (2009) DNA analysis from plant remains and plant opal analysis excavated from the Maekawa site. Report of cultural deposits research in Aomori prefecture, no. 475, vol. 2, pp 97–100 [in Japanese]
Umbeck PF, Gengenback BG (1983) Reversion of male-sterile T-cytoplasm maize to male fertility in tissue culture. Crop Sci 23:584–588
Umemoto T, Yano M, Satoh H, Shomura A, Nakamura Y (2002) Mapping of a gene responsible for the difference in amylopectin structure between japonica-type and indica-type rice varieties. Theor Appl Genet 104:1–8
Zhao Z (2010) New data and new issues for the study of origin of rice agriculture in China. Archaeol Anthropol Sci. doi:10.1007/s12520-010-0028-x
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Tanaka, K., Honda, T. & Ishikawa, R. Rice archaeological remains and the possibility of DNA archaeology: examples from Yayoi and Heian periods of Northern Japan. Archaeol Anthropol Sci 2, 69–78 (2010). https://doi.org/10.1007/s12520-010-0036-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-010-0036-x