
Abstract
The analysis of benthic foraminiferal assemblages was performed in the Oxfordian deposits of the “Argile de Saïda” in northwestern Algeria. This formation consists mainly of clay and sandstone alternations of the Upper Jurassic. It is located in the western part of the Tlemcenian domain. The first results of the micropaleontological study of Djebel Brame indicated that the microfauna of Foraminifera was quite original and widely abundant during that period. In addition, their biostratigraphic distribution exhibited three microfaunal associations, largely dominated by Nodosariidae. These results also revealed the renewal of the assemblage composition and allowed identifying several species of biostratigraphic divisions which were found consistent with those established for the ammonites. In addition, the palaeoenvironmental interpretations were based on the spatio-temporal distribution of benthic foraminiferal associations with hyaline and agglutinated tests. Besides, the geochemical analyses (calcimetry) reflected the basin infill during the Late Oxfordian.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
During the Callovo-Oxfordian, tectono-eustatic events affected the entire Tlemcenian domain (Elmi 1978a; Elmi and Benest 1978; Benest 1985; Benest et al. 1995, 1997). Their consequences on the study region, i.e., Djebel Brame, led to the installation of a platform where clay-sand formations, known as the “Argiles de Saïda” formation, were deposited; these formations were very favorable to the development of benthic organisms. The formation of Saïda clays in the eastern part of the Tlemcenian domain was first described by time by Atger and Verdier (1965) in the region of Sidi Kadda (ex Kechrou), south of the city of Mascara; it has been the subject of numerous studies (Lucas 1952; Augier 1967; Delfaud 1973; Elmi 1978b; Elmi and Benest 1978; Touahria 1979; Benest et al. 1995; 1997; Bendella et al. 2011; Cherif et al. 2015; Halamski and Cherif 2017). The evolution of this formation, during the Oxfordian, showed a tendency to filling which started, during the Middle Oxfordian, with the detritic sedimentation to pass then to the carbonate sedimentation at the beginning of the Upper Oxfordian. It was later completed with the formation of the Bou-Medine sandstones at the end of the Oxfordian-Kimmeridgian. The sediments in the region of Djebel Brame (north-west of Algeria) are represented by interbedded clayey sand beds from the Upper Jurassic, rich in foraminiferous microfauna. This section was subdivided into three lithostratigraphic units, namely a lower clayey sandstone unit, a medium clayey carbonate unit, and a higher clayey sandstone unit. The study of the Callovo-Oxfordian Foraminifera in the Saïda Mountains has been undertaken for a few years. Results have already been obtained in the Saïda region (Touahria and Sebane 2019). The study of Foraminifera in the Djebel Brame section made it possible to identify several species in the biostratigraphic division. The findings obtained in this biostratigraphic division turned out to be consistent with those established for the ammonites (Cherif et al. 2015). Observations based on the Foraminifera test made it possible to distinguish two different forms, namely the hyaline form and the agglutinated form. The tectono-eustatic events of these two forms affected the entire area; they seemed to have a certain impact on the vertical sequence of Foraminifera. In addition, a calcimetric analysis was also carried out on the marls at the foraminiferous levels in order to formulate a paleoenvironmental interpretation.
Geological setting
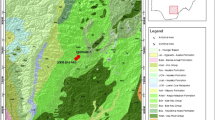
The study section is located in the Saïda Mountains; this area is located approximately in the eastern part of the Tlemcenian domain (Fig. 1a). This area is bordered to the north by the Tellian Atlas and to the south by the pre-Atlas (Hauts Plateaux s.s.), as clearly shown in Fig. 1a. The outcrop understudied, a section of Djebel Brame, with a height of 906 m, is located 21 km to the north of the town of Takhemaret (Tiaret, northern Algeria) and 8 km southeast of the village of Hadj El Arbi (Fig. 1b). The section already studied on the biostratigraphic and sedimentological level (Bendella et al. 2011; Cherif et al. 2015; Cherif 2017) is represented by alternating sandy and clayey levels, rich in Foraminifera from the Upper Jurassic. This section was subdivided into three lithostratigraphic units which probably form the upper member of the “Argile de Saïda” formation. These three units are the lower clay-sandstone unit, the medium clay-carbonate unit, and the higher clay-sandstone unit. The other two lower and middle members, which date back to the Callovian stage, were defined by Touahria (1979) in the region of Saïda.

Geographical location of the study region. a Geographic location of the studied section in northern Algeria. b Geographic location of the studied outcrop
Lith of Djebel Brame section
It is useful to recall that the “Argile de Saïda” formation, in the study area, was subdivided into three distinct lithostratigraphic units, with uneven development (Bendella et al. 2011; Cherif et al. 2015; Cherif 2017), namely the lower clayey sandstone unit, the medium clayey carbonate unit, and the higher clayey sandstone unit, as shown in Fig. 2:

Lithostratigraphic and biostratigraphic subdivision of Brame mountain (Cherif 2017)
Lower clayey sandstone unit (210 m)
This formation starts with a comb of clay, 20 m thick, interspersed with a few micaceous sandstone layers, of centimetric thickness, and numerous hydrodynamic structures. The base of this formation is marked by the intercalation of two conglomeratic channels, with a sandy-clay matrix including some polygenic centimetric pebbles (limestone, sandstone), associated with clay balls, the debris of silicified wood, and solitary polypiers. The base of the sandstone beds is often irregular, erosive, and rich in erosion figures (sole marks). These are essentially turbulent scour figures (gutter casts and flute casts) and figures of dragged objects (groove marks, prod marks, skip marks, and bounce marks). Most often, the bed interface exhibits multi-directional hydrodynamic structures; these are mainly horizontal laminations, sometimes accompanied by hummocky cross-stratification (HCS). The upper and lower surfaces of the sandstone beds include numerous ichnofossils (Bendella et al. 2011; Cherif et al. 2015; Cherif 2017), such as the Chondrites, Cochlichnus, Didymaulichnus, Diplichtines, Megagrapton, Ophiomorpha, Palaeophycus, Planolites, Taenidium, and Thalassinoides. The ammonites collected by Cherif (2017) and Cherif et al. (2015) in this unit are represented by the following species: Subdiscophinctes sp., and Discosphinctes sp. This association indicates the Transversarium zone (Shilli subzone)
Median clay-carbonate unit
These are greenish clays with strato-increasing beds of oolithic or oncolithic limestones, of grayish color, hard, and rich in calcified gastropods and crinoid stems. This lithostratigraphic succession continues with an alternation of greenish clays and greso-carbonate lenses, with oolitic particles, arranged in cross laminations, as well as micritic limestones in centimetric and decimetric beds. Towards the summit, this formation ends with a bed of oolithic limestones. The rare ammonites reported in this unit by Cherif (2017) are represented by an association of Dichotomoceras crassus (Enay 1966) and Larcheria gr. Schilli (Oppel 1863). This association indicates the Rotoides subzone (Transversarium zone).
Upper clayey sandstone unit
This is basically an alternation of greenish clays with fine-grained sandstone streaks that are most often affected by slump-like deformations. The base of the sandstone beds exhibits erosion figures (scours and dragged objects) with small-size loads; however, their roofs show symmetrical or asymmetrical current ripples. The bed interface presents hummocky cross-stratification, as shown in Fig. 2, horizontal laminations, and oblique or cross laminations. The middle part, which is marked by the appearance of a fossiliferous limestone slab with a plurikilometric lateral extension equivalent to the sub-reef level defined by Auclair and Biehler (1967) and Benest et al. (1998), about 1 m thick, reveals a bioclastic fraction composed of solitary polypiers, sea urchins, brachiopods, gastropods, crinoid stems, ammonites, and belemnites. The top part of the formation is characterized by the appearance of a metric bar that is composed of sandstone levels belonging to the “Sidi Amar sandstone formation” (Upper Oxfordian-Lower Kimmeridgian) (Auclair and Biehler 1967; Ghali 1984; Benest 1985). Rare ammonites are indicated in this unit by Cherif (2017). The Bifurcatus Zone is identified by the record of Passendorferia gr. ziegleri or Teresiformis (Brochwicz-Lewinski 1973).
Materials and methods
The Foraminifera assemblages described in this study were recovered from the Jebel Brame section. The samples, weighing approximately 2 kg, were collected from the clay levels of the section. A mass of 0.2 kg of clay was used in each level. These clay samples were soaked in tap water for 24 h. After disintegration, the rinsing procedure involved washing the sediments under a gentle jet of water over a set of standard stainless steel sieves (250, 125, and 65 μm). The collected residues were oven-dried at 40 °C. The Foraminifera were hand-picked with a hair paint-brush on a standard black picking grid-tray under a binocular microscope and the microfossils extracted. The Foraminifera were analyzed, mainly in the fraction > 125 μm, although they were occasionally abundant in the fraction 125–65 μm. In the present work, the classification of Foraminifera is adopted from Tappan and Loeblich Jr. (1988). This made it possible to distinguish 28 species, divided into 10 genera (Fig. 2). The assemblages obtained are well suited for a paleoecological analysis that is based on foraminiferous tests (agglutinative test and hyaline test). Some of these marls are also intended for the calcimetric study in order to quantify the richness of Caco3. The CaCO3 content was determined using the Dietrich-Frùling calcimeter. The test carried out consisted of measuring, using a gas burette, the volume of CO2 released by the reaction of HCl with the calcium carbonate contained in the sample.
Results and discussion
Biostratigraphy: foraminiferal assemblages
A detailed study of the Foraminifera and its stratigraphic distribution in the Djebel Brame section is shown Fig. 3. It allows identifying three successive assemblages during the Oxfordian, each one defined by the characteristics species. The Foraminifera taxa observed are exclusively benthic and are dominated by Nodosariidae belong to the sub-orders Textulariina, Lagenina, and Rotalina. These Foraminifera zones are defined and described herein from oldest to youngest as follows:
-
Assemblage 1.
It is characterized by a predominance of hyaline Foraminifera mainly composed of Ammodiscus sp., Citharina clathrata, Garentilla ampasindavensis, Lenticulina aff batrakiens mg. Marginulopsis, Triplasia bertensteini, Lenticulina munsteri mg. L, Spirillina infima, and Leniculina sp. All these forms indicate the Oxfordian.
-
Assemblage 2.
It includes agglutinated forms, in association with the first planktonic forms. It is comprised of the following forms: Lenticulina fraasi mg. Astacolus, Textularia jurassica, Dentalina vetusta, Globuligerina axfordiana, Lenticulina Polymorpha mg. L, Spirillina elongala, Lenticulina gottimgensis. mg. L, Lenticulina musnsteri mg. L, Spirilina infima, and Lenticulina sp. The ammonite fauna harvested by Cherif (2017) in the same clay levels of Subdiscosphinctes sp., Dichotomoceras crassus (Enay 1966), and Larcheria gr. Schilli which characterizes the Middle Oxfordian (Transversarium zone).
-
Assemblage 3.
It is mainly characterized by Nodosariidae such as Lenticulina musnsteri mg. L, Spirillina infima, Lenticulina sp., Lagena sp., Lenticulina varians mg. L, Marginulina jurassica, and Ammodiscus sp. The associated agglutinated forms are reduced quantitatively and they are represented by Textularia jurassica, Trochammina inflata, Ammobaculites fontinensis, and Ammobaculites agglutinans. This assemblage belongs to Upper Oxfordian (Bifurcatus zone). It was noted the presence of the ammonite macrofauna (Cherif 2017), Dichotomoceras bifurcatoides (Enay 1966), more often with Dichotomoceras aff. Stenocycloides (Siemiradzki 1898) and Subdiscosphinctes sp. This assemblage characterizes the Upper Oxfordian (Bifurcatus zone).

Stratigraphic range of main species of Foraminifera from Oxfordian in the Jebel Brame
Paleoecology
The analysis of benthic Foraminifera from the region of Djebel Brame revealed several assemblages, largely dominated by benthic Foraminifera (Plate 1). These assemblages are made up of two groups. The first one gathers the agglutinated test forms; the suborder Textulariina is exclusively represented by Ammobaculites. The second one is that of the hyaline test forms which is dominated by the Nodosariidae (Fig. 3), whose temporal distribution is not random but seems to depend on sedimentary and tectono-eustatic variations. It was found that the evolution of the composition of foraminiferal populations is consistent with the sequential division previously established by Cherif (2017). In addition, variations in the proportions of agglutinants, Spirillins and Lenticulins, can be used to characterize the main paleobathymetric variations. Furthermore, different types of sedimentary discontinuities were pointed out in the Oxfordian of Djebel Brame.

Agglutinated Foraminifera from Djebel Brame section: (1) Ammobaculites fontinensis and (2) Textularia jurassica. Hyaline benthic Foraminifera from Jebel Brame section: (3) Spirillina infima. (4) Lenticulina musnsteri mg. L. (5) Dentalina varians. (6) Marginulina jurassica
The bottom part of the lower clay-sandstone formation has a CaCO3 contents close to 12%. It is characterized by the predominance of Foraminifera with hyaline tests (80%), where the Spirillins proportion is higher than those of the other groups. In addition, the proliferation of Spirillins reflects a decrease in depth where the abundance of this group could be explained by the wide availability of trophic resources linked to photosynthetic production in a shallow proximal platform environment (Bouhamdi et al. 2001). Then, as the CaCO3 proportions diminish, an increase in lenticulins is noted, with the appearance of dentalines. Note that the ornamentation is well preserved, and the normal size of the Foraminifera rather suggests an increase in depth in an external, well-oxygenated platform environment that is favorable to benthic life.
The transgressive interval from the end of the Middle Oxfordian period is characterized by a detrital sedimentation materialized by a reduction in the proportion of CaCO3 and a high clay level (Fig. 4). It is worth indicating that the microfauna collected in the clay levels becomes more abundant; it is largely dominated by agglutinative forms (68%), especially small agglutinated Foraminifera with siliceous test (Trochammina) associated with pelagic forms (Globuligerina) (Plate 2) and dentalinids. A decrease in spirilllins and lenticulins was also noted, which testifies to the depression of the platform, in the form of a basin (Elmi and Benest 1978) and an increase in salinity during the progressing transgression. This corresponds to a period which was followed by a progradation phase that was materialized by a high-sea-level prism. It should be noted that the increase in lenticulins implies the start of a filling sequence.

Relative frequency of foraminifera (hyaline and agglutinated), variation in the level of CaCo3, the palaeoenvironment and the sea-level fluctuations

Planktonic Foraminifera. Globigerina oxfordiana: (a) ventral side and (b) dorsal side
During the Upper Oxfordian, the facies, which is represented by a clay-carbonate formation, begins first with oolithic limestone levels with fossil accumulations and then continues with an alternation of clays with sandstone lenses. The proportion of calcium carbonate reaches higher values than those recorded in the underlying deposits; they are around 15%. Foraminifera are characterized by the abundance of hyaline forms (80%), with high proportions of lenticulins and Spirillins. The microfaunal renewal is marked by the predominance of Nodosariidae with coiled forms. It should be noted that the agglutinating group is scarce and represents an average of 20%, which indicates a shallower environment with a more or less carbonated platform.
Conclusions
The study of foraminiferal populations in the region of Djebel Brame made it possible to collect quite important micropaleontological, biostratigraphic, and paleoecological information. From the micropaleontological point of view, 28 species of Foraminifera belonging to 10 genera were found. Foraminifera biostratigraphy allowed identifying three associations of benthic Foraminifera during the Oxfordian. Their distribution depends on the variation of the relative sea level and also on the tectonics that affects the whole area and which shows an abundance of calcareous hyaline forms in shallow environments, with a wide availability of Spirillins in the flat-proximal form environment that is rich in trophic resources due to elevated photosynthetic production. Moreover, high proportions of lenticulins were observed as depth increased. In addition, the transgressive interval was characterized by the development of agglutinated forms associated with pelagic forms (Globuligerina) and dentalines in a deeper environment, rich in terrigenous material.
References
Atger M, Verdier J (1965) Etude géologique du plateau jurassique de Cacherou (Sidi Kada), bordure Nord des Monts de Saïda. Géoscience, Société Nationale de Recherche et d’Exploitation de Pétrole en Algérie
Auclair D, Biehler J (1967) Etude géologique des Hautes Plaines oranaises entre Tlemcen et Saïda. Publ Serv Géol Algérie (Nouvelle Série) 34:3–45
Augier C (1967) Quelques éléments essentiels de la couverture sédimentaire des Hauts Plateaux. Publ Serv Géol Algérie 34:47–80
Bendella M, Benyoucef M, Cherif A, Benhamou M (2011) Ichnology and sedimentology of the “Argiles de Saïda” formation (Callovo-Oxfordian) of the Djebel Brame (Tiaret, Algeria). Bull Soc Géol France 182:417–425
Benest M (1985) Evolution de la plateforme de l’Ouest algérien et du Nord-Est Marocain au cours du Jurassique supérieur et au Crétacé: Stratigraphie, milieux de dépôts et dynamique sédimentaire. Doc. Lab. Géol., Univ, Lyon1. Fasc. I et II
Benest ES, Ouardas T, Perriaux J, Ghali M, Benhamou M (1995) Dynamique de mise en place d’un cône détritique d’âge callovo-oxfordien dans le sillon intra- cratonique du Nador de Tiaret sur la marge téthysienne de l’Ouest algérien. Compt Rendus Acad Sci Paris 321:103–110
Benest M, Elmi S, Ouardas T, Perriaux, Behamou M (1997) Les facteurs de contrôle de la sédimentation de l’éventail détritique callovo-oxfordien du Djebel Nodor de Tiaret sur marge continentale sud-téthysienne en Algérie occidentale. Bull Serv Géol Algérie 8(1):3–17
Benest M, Ouardas T, Boullier A (1998) Conditions d’installation d’un seuil corallien dans l’intervalle transgressif Oxfordien moyen et supérieur p.p. entre le bassin tellien et son avant-pays (Ouest algérien) ; implications paléographiques. Comptes Rendus l’Académie des Sciences, Paris:326: 399–405
Bouhamdi A, Gaillard C, Ruget C (2001) Spirillines versus agglutinants: impact du flux organique et intérêt paléoenvironnemental (Oxfordien moyen du Sud-Est de la France). Geobios 34:266–277
Brochwicz-Lewinski W (1973) Some remarks on the origin of the subfamily Idoceratinae Spath, 1924 (Perisphinctidae, Ammonoidea). Acta Palaeontol Pol 18
Cherif A (2017) Sédimentologie et dynamique sédimentaire de la Formation des Argiles de Saïda (Oxfordien moy-sup) de la partie orientale du domaine tlemcénien (Algérie ouest). Université d’Oran2 Mohamed ben Ahmed
Cherif A, Bert D, Benhamou M, Benyoucef M (2015) La Formation des Argiles de Saïda (Jurassique supérieur) dans le domaine tlemcenien oriental (Takhemaret, Algérie): données biostratigraphiques, ichnologiques et sédimentologiques
Delfaud J (1973) Sur l’appartenance de certains “pseudo-flyschs” aux faciès prodeltaïques de plateforme. Compt Rendes Acad Sci Paris 277:1125–1128
Elmi S (1978a) Polarité tectono-sédimentaire pendant l’effritement des marges septentrionales du bâti africain au cours du Mésozoïque (Maghreb). Ann Soc Géol Nord 97:315–323
Elmi S (1978b) Polarité tectono-sédimentaire pendant l’effritement des marges septentrionales du bati africain au cours du Mésozoique (Magreb). Annales de la Société Géologic du Nord,XCVII:315–323
Elmi S, Benest M (1978) Les “argiles de Saïda”: faciès flyschoïde du Callovo-Oxfordien du sillon tlemcenien (Ouest algérien); stratonomie, environnement, interprétation et évolution paléogéographique. Livre jubilaire Jacques FLANDRIN.-Documents du Laboratoire de Géologie de la Faculté des Sciences de Lyon, Villeurbanne. Hors Série 4:203–261
Enay R (1966) L’Oxfordien dans la moitié Sud du Jura français. Etude Stratigraphique Publ Musée Confluences 8:1–323
Ghali M (1984) Le Jurassique supérieur et le Crétacé basal des Monts de Saïda et de Tiaret-Frenda. Université Claude-Bernard Lyon 1
Halamski AT, Cherif A (2017) Oxfordian brachiopods from the Saïda and Frenda mountains (Tlemcenian Domain, north-western Algeria).in Annales Societatis Geologorum Poloniae
Lucas G (1952) Bordure Nord des Hautes Plaines dans l’Algérie occidentale: primaire, jurassique, analyse structurale. Verlag nicht ermittelbar
Oppel A (1863) III- Über jurassische Cephalopoden:. Paleontologische Mittheilungen aus dem Museum des Koeniglischen Bayerischen Staates, Stuttgart:127–266
Siemiradzki J (1898) Monographic description of the ammonite group Perisphinctidae. Palaeontographia 45:69–296
Tappan H, Loeblich AR Jr (1988) Foraminiferal evolution, diversification, and extinction. J Paleontol:695–714
Touahria A (1979) Biostratigraphie du Callovien des environs de Saïda (Algérie occidentale). Les Reineckeiidae (Ammonitina, Perisphinctaceae). Thèse 3ème Cycle, Univ Lyon:152 p
Touahria A, Sebane A (2019) Paleobathymetric influence on the distribution of ammonite and foraminifer settlements in the Callovian of the Saïda Region (Western Algeria). Pages 75-78 paleobiodiversity and tectono-sedimentary records in the Mediterranean Tethys and related eastern areas. Springer
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Attila Ciner
This paper was selected from the 2nd Conference of the Arabian Journal of Geosciences (CAJG), Tunisia 2019
Rights and permissions
About this article

Cite this article
Ziouit, K., Sebane, A., Sebane, A.T. et al. Benthic Foraminifera from the Saïda clay formation (Djebel Brame, northwestern Algeria) biostratigraphy and paleoecology. Arab J Geosci 14, 5 (2021). https://doi.org/10.1007/s12517-020-06204-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12517-020-06204-x