
Abstract
Three corbulid species were discriminated from the Cenozoic strata of Kutch, India. Two of them are Caestocorbula gujaratensis n. sp. and Bicorbula kutchensis n. sp. from the Palaeocene to Middle Eocene. The other species, Corbula trigonalis Sowerby, from the Lower Miocene is reassigned to Lenticorbula. The two new species co-occur in the same horizons. Palaeobiogeographical distribution of molluscs in general and corbulids in particular in the Indian subcontinent, especially of India and Pakistan, reveals high endemism. The distribution also evinces palaeobiogeographical closeness among the western Indian basins and Pakistan, thus entailing their inclusion in the same province. This province shows similarity in faunal composition with Africa and Middle East during the Lower Palaeogene whereas with southeast Asian basins during the Miocene. The pattern reflects a shift in biogeographical affinity of the province from Tethyan to Indo-Pacific.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Cenozoic sedimentary rocks are widely distributed in the Indian subcontinent along a vast, almost continuous tract from the Sind-Baluchistan region in Pakistan in the west to Assam-Meghalaya, India in the east through the Himalayan foothills. Contemporaneous deposits are also well exposed in Myanmar, Java and other southeast Asian islands. In peninsular India, the Cenozoic deposits occur extensively along narrow tracts of the east and west coastal belts (Krishnan 1982). Kutch in the west of India houses a thick Cenozoic marine sequence (Biswas 1992), which yielded a rich mollusc fauna. The Sind-Baluchistan region in the west of the extra-peninsular part of the subcontinent is known for its rich mollusc record for a long time (d’Archiac and Haime 1854; Cossmann and Pissarro 1909, 1927; Vredenburg 1925, 1928a, b; Cox 1930, 1931; Eames 1951, 1952; Iqbal 1969a, b, 1972, 1980).
Corbulid bivalves are a small component of this diverse fauna. Most of the corbulid species have been reported from the Sind-Baluchistan and their adjoining areas in the western part of the extra-peninsula (d’Archiac and Haime 1854; Cossmann and Pissarro 1927; Cox 1930, 1931; Vredenburg 1928a; Eames 1951; Iqbal 1969a, b, 1972, 1980).
Sowerby (1840) reported Corbula trigonalis Sowerby 1840 and Corbula rugosa Lamarck 1801 from the Lower Miocene of Kutch for the first time. d’Archiac and Haime (1854) also reported C. trigonalis from Sind and Kutch. Vredenburg (1928a) reported five species of Corbula Bruguière 1797 from Pakistan and Kutch, only two of which come from the Lower Miocene of Kutch. They are C. trigonalis and Corbula tunicosulcata Vredenburg 1928a. The latter is a synonym of C. rugosa Lamarck sensu Sowerby 1840. Tandon (1971, 1976) recorded Corbula subexarata d’Archiac and Haime 1854 and Corbula sp. from the lower Middle Eocene Harudi Formation of Kutch. Kachhara et al. (2011a) recorded four species from the Palaeocene of Kutch—Corbula (Bicorbula) harpa d’Archiac and Haime 1854, Corbula (Bicorbula) samanensis Cox 1930, Corbula (Bicorbula) praexarata Cox 1930 and Corbula (Bicorbula) cf. pseudorakhiensis Eames 1951. Kachhara et al. (2011b) also noted the presence of C. (Bicorbula) subexarata var. lituus de Cotter 1923 from the Upper Eocene of Kutch. These are, so far, the only known fossil corbulid species from the Cenozoic of Kutch. Sowerby (1840) did not provide a detailed description of C. trigonalis. d’Archiac and Haime (1854) described the species very briefly. Neither did Vredenburg (1928a) describe the species. However, he provided a detailed description of the other species from Kutch, i.e., C. tunicosulcata. Tandon (1971, 1976), when dealt with the biostratigraphy of the Middle Eocene rocks of Kutch, erected a zone named after C. subexarata. He did not provide any description or figure of the species. The report by Kachhara et al. (2011a) is based only on a few specimens and the documentation in terms of remarks and figures is inadequate.
Here, we aim to make an account of the corbulids we have found from the Cenozoic succession of Kutch in respect of their systematics and palaeobiogeography. Furthermore, we review the distribution of the Cenozoic corbulid species reported so far from different parts of Pakistan and India to get a picture of the palaeobiogeographical pattern of the Indian subcontinent and its evolution.
Palaeobiogeographical and stratigraphical settings
Palaeobiogeography
It has been known since long that the Cenozoic basins of Pakistan and those of Himachal Pradesh, Rajasthan and Gujarat in western India, share a significant faunal similarity (e.g., Sowerby 1840). This implies continued connectivity of these basins through the Cenozoic. The Pakistan fauna is the richest in diversity and has been representing the western part of the Indian subcontinent in literature. This is also better documented than the western Indian basin faunas. Kutch fauna from the post-Eocene times is relatively better documented and its palaeobiogeography has been studied in some detail in recent literature (Sowerby 1840; Vredenburg 1925, 1928a; Borkar et al. 2004; Harzhauser et al. 2009; Kulkarni et al. 2010; Kachhara et al. 2012). Pre-Oligocene molluscs from Kutch are poorly known and their palaeobiogeographical study is at a nascent stage (Kachhara et al. 2011a, b; Halder 2012).
Palaeobiogeographical affinity of the basins in the western part of the Indian subcontinent with eastern African ones in the Lower Palaeogene has been a commonplace observation. Popov (1993) found strong faunal homogeneity of the Indian fauna with those from Egypt and Somalia and erected the Indo-African subregion consisting of these basins within the Tethyan Region for the Upper Eocene. He observed that the Mediterranean subregion lying to the north of Tethys, constituted of Italy and France, differs significantly from the Indo-African to the south of the ocean. Cox (1930, 1936) found Bahrain and south Iranian faunas of the Eocene similar to faunas from Pakistan, Somalia, and Egypt. Iqbal (1969a, b, 1972) observed general similarity of the Palaeocene and Eocene molluscs from Pakistan with western Tethys faunas from Europe, Africa, and Middle East countries. Haas and Miller (1952) also found similarity of the Eocene nautiloid (Cephalopoda, Mollusca) fauna from Somalia with that from Pakistan. On the other hand, Myanmar, Java, Borneo, and Sumatra were believed to constitute a distinct bio-province, which faunally differed almost entirely from Pakistan and Somalia during the Eocene (de Cotter 1923). Martin (1931) proposed an Indo-Malayian Province based on similarity of the Upper Eocene faunas from Myanmar, Java, and eastern India. This faunal disjunction between Pakistan and southeast Asian countries apparently continued in the Oligocene (Vlerk 1931). Barron and Peterson (1991) demonstrated that Myanmar and Java were separated from north-western India by a strong western boundary current during the Eocene.
Harzhauser et al. (2002, 2007) and Harzhauser (2007) recently formalized the biogeographic framework of the Tethys Realm for the Oligocene and the Lower Miocene. They included Kutch and Sind in the Western Indian Province (sensu Harzhauser 2007) of the Western Tethys Region belonging to the Tethys Realm for the Oligocene. The province is akin to the Eastern African-Arabian Province and also shares similarity to the Mediterranean-Iranian Province. In the Lower Miocene, they observed, the Indian fauna essentially got separated from the Western Tethys Region and started getting influence from southeast Asian faunas such as those from Indonesia, Java, and Borneo (Harzhauser et al. 2002, 2007, 2009). Hence, Harzhauser et al. (2002, 2007) considered this Lower Miocene Indian fauna to belong to the Proto-Indo-West Pacific Region and termed the province Proto-Western Indian Ocean Province.
Stratigraphy
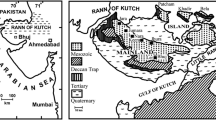
The Palaeogene and Neogene formations of Kutch occur along the present coast line in an arcuate outcrop (Fig. 1). These Cenozoic sedimentary rocks unconformably overlie the Deccan Traps and are tectonically less disturbed. Biswas (1992) classified the sedimentary sequence into eight lithostratigraphical units—the Matanomadh Formation, the Naredi Formation, the Harudi Formation, the Fulra Limestone, the Maniyara Fort Formation, the Khari Nadi Formation, the Chhasra Formation and the Sandhan Formation in ascending order (Fig. 2).

Geographical location of the study area and geological map of the Cenozoic of Kutch with fossil localities (modified after Biswas 1992)

Generalized lithostratigraphical section of part of the Cenozoic of Kutch with the distribution of C. gujaratensis n. sp., B. kutchensis n. sp. and L. trigonalis (Sowerby)
The Upper Palaeocene–Lower Eocene Naredi Formation is composed mostly of gypseous shale interspersed with layers of concretions in the lower part and capped by a ferruginous claystone. Coal seams occur in the lower part of the formation while a persistent limestone studded with the larger foraminifer, Assilina sp., characterizes the middle part (Biswas 1992; Garg et al. 2011). The foraminiferal assemblage indicates the Early Eocene age of the formation. However, suggestions of an earlier (i.e., Palaeocene) inception of deposition are also there (see Biswas 1992; Chattoraj et al. 2009; Kachhara et al. 2011a). The Middle Eocene Harudi Formation is also an argillaceous unit with partings of thin to moderately thick layers of carbonate concretions in its lower part. A few mollusc-rich shell concentrations, which are persistent over a large area, occur in this part of the formation. Sometimes the shell concentrations are associated with carbonate concretions. Tandon (1971) called this fossiliferous horizon the C. subexarata Zone. A persistent key bed studded with Nummulites spp. occurs in the middle of this formation. The early Middle Eocene age has been assigned to this formation (Biswas 1992; Banerjee et al. 2012).
The Lower Miocene Chhasra Formation is very widely distributed and mostly composed of clayey shale in the lower and siltstone in the upper part. Interspersed within this formation are some layers of richly fossiliferous marls and limestones. Most of the fossils from this formation come from these marl/limestone layers. The Burdigalian (Early Miocene) age has been assigned to this formation (Biswas 1992; Kulkarni et al. 2010).
Material and methods
The specimens have been collected from a single locality (Loc. 1, Fig. 1) in the Naredi Formation, four localities (Locs. 2 through 5, Fig. 1) in the Harudi Formation, and one locality (Loc. 6, Fig. 1) in the Chhasra Formation (see Table 1). All the specimens are deposited in the Department of Geology, Presidency University, Kolkata (abbreviated PG). Types and other specimens housed in the Repository of Fossils, Curatorial Division, Geological Survey of India, Kolkata (abbreviated GSI) have also been studied. Most of the specimens were coated with magnesium oxide before photography.
Palaeobiogeographical distribution of the corbulid species from Kutch and other basins of India and Pakistan is studied based on published literatures and plotted in palaeocoastline maps of Smith et al. (1994) for the Middle Eocene and the Lower Miocene. The extent of similarity and endemism of the faunas is used to interpret possible palaeobiogeographical relationships among the basins.
The distribution data of the Cenozoic corbulids are collated primarily from the following literatures—Sowerby (1840); d’Archiac and Haime (1854); Cossmann and Pissarro (1927); Vredenburg (1928a); Eames (1951), and Iqbal (1969a, b, 1972). Mathur (1988) provided an outline of mega-invertebrates reported from India. Later, Lyngdoh et al. (1999) and Kachhara et al. (2011a, b) reported some corbulids, respectively, from the Lower Miocene of Meghalaya and the Palaeogene of Kutch. We have revisited the species reported by Cossmann and Pissarro (1927) and Vredenburg (1928a) but cannot do so for the others because of inaccessibility of most of the collections. The species reported by Kachhara et al. (2011a) from Kutch are based on inadequate documentation. Our knowledge of the fauna from the area supports only two corbulids. The rest of Kachhara et al. (2011a, b) are not considered here.
Systematic palaeontology
Class Bivalvia Linnaeus 1758
Order Myoida Stoliczka 1870
Family Corbulidae Lamarck 1818
Genus Caestocorbula Vincent 1910
Type species Caestocorbula gerardi Vincent 1922 (= Corbula henckeliusiana Nyst sensu Vincent 1890 (as C. henckeliusi, spelling error), by original designation; non C. henckeliusiana Nyst 1836). Eocene of Belgium.
Caestocorbula gujaratensis n. sp.
(Fig. 3)

C. gujaratensis n. sp. from the Eocene Harudi Formation. a-c PG/Cb 134 (holotype)—external view of left valve (a), external view of right valve (b), dorsal view (c). d PG/Cb 192 (paratype), dorsal view. e–g PG/Cb 361 (paratype)—external view of left valve (e), external view of right valve (f), dorsal view (g). h–i PG/Cb 384 (paratype)—external view of left valve (h), external view of right valve (i). j PG/Cb 386 (paratype), dorsal view. k–m PG/Cb 451 (paratype)—external view of left valve (k), external view of right valve (l), dorsal view (m). n–o PG/Cb 452 (paratype)—external view of left valve (n), external view of right valve (o). p–q PG/Cb 511 (paratype)—external view of left valve (p), external view of right valve (q). r PG/Cb 416, right valve, internal view. s–t PG/Cb 95, left valve, internal view (s), dorsal view (t). u–v PG/Cb 398 (paratype)—external view of left valve (u), external view of right valve (v). w–x PG/Cb 512 (paratype)—external view of left valve (w), external view of right valve (x). y PG/Cb 515 (paratype), external view of right valve. z PG/Cb 543, left valve, internal view. aa PG/Cb 595, right valve interior in close-up. ab PG/Cb 596, dorsal margin of left valve in close-up. ac PG/Cb 597, dorsal view of left valve in close-up. ad PG/Cb 598, left valve hinge in close-up. S, pallial sinus, R, chondrophore and L, groove demarcating the position of left valve inside right valve marked by arrow. Specimens except internal and close-up views coated with magnesium oxide. Scale bar for a–z = 10 mm, for the rest = 5 mm
1931 Corbula. (Bicorbula) subexarata d’Archiac and Haime—Cox p. 84, pl. 4, fig. 14a–b.
1936 C. (Bicorbula) subexarata d’Archiac and Haime—Cox p. 33, pl. 4, figs. 4–5.
2011 C. (Bicorbula) praexarata Cox—Kachhara et al., p. 87, pl. 1, fig. 8.
Type specimens
Holotype: PG/Cb 134; paratypes: PG/Cb 192, 361, 384, 386, 398, 451, 452, 511, 512, 515.
Other material examined
More than hundred specimens, many with original shell, a few single valve, others articulated, in the collection of Presidency University.
Type locality, horizon and age
Loc. 4, about 2 km north of village Harudi (23°32′N, 68°40′52″E) (Fig. 1, Table 1), Harudi Formation, lower Middle Eocene.
Etymology
The specific name is after the state of Gujarat, which subsumes the district of Kutch.
Diagnosis
Large, strongly inequivalved; right valve larger, more convex and more rostrate than left valve; hinge of right valve bears a short blunt cardinal tooth, in left valve a nearly horizontal chondrophore posterior to cardinal socket; very shallow pallial sinus; surface of both valves with prominent commarginal ribs—in left valve closer and finer than in right, left valve also bears some blunt faint radial ridges.
Description
Shell large (maximum length observed 30.5 mm), ovate-trigonal, prominently inequivalved with larger right valve; right valve inflated, posteriorly rostrate, umbo of right valve high, projected beyond its hinge, beak incurved, prosogyral; beak of left valve tucked in just below and posterior to beak of right valve; left valve less convex than right valve; anterior margin rounded; posterior part narrow, posterior margin truncated in left valve, in right valve often broken; anterior slope wide, marked by a somewhat prominent border in left valve; posterior slope narrow, traversed by a prominent groove in left valve, slope boundary obscure except a low and sharp ridge in left valve; hinge of right valve with a short blunt cardinal tooth, tooth not projected beyond its beak, a deep resilifer pit posterior to cardinal tooth to receive chondrophore, a small tubercle present posteriorly and between beak and cardinal tooth; left valve hinge with a cardinal socket, socket with a ventral floor, posterior to cardinal socket a chondrophore oblique to hinge with horizontal dorsal surface and upturned ventral surface; dorsal surface of chondrophore bears a deep median longitudinal groove, ridge anterior to groove somewhat longer and higher than posterior ridge, ridges flanked by narrow flat marginal parts, chondrophore transversely striate; two slightly unequal adductor muscle scars, anterior one slightly smaller and reniform, posterior one subelliptical; pallial line with a shallow sinus, sinus with rounded closure; interior of right valve bears a strong venter-parallel groove marking position of left valve margin in closed valves, groove continues into lateral lamellae on either side of hinge; surfaces of both valves characterized by strong commarginal ribs, number of ribs in both valves comparable, ribs stronger and widely spaced in right valve while finer and closer in left valve, left valve also bears faint somewhat distantly spaced radial ridges, faint close-spaced radial lines also visible in delaminated right valve, commarginal growth lines numerous, commarginal lines and ribs mainly of right valve show a small notch in their anterior part; shell thick.
Discussion
The genus Caestocorbula is characterized by a strongly inequivalved shell, rostrate posterior side of the right valve, prominent siphonal plates at the posterior end of the left valve, a higher umbo in the right valve, absence of nepionic cap and discrepant commarginal ornaments, which are stronger in the right valve. The subgenus Parmicorbula Vokes 1944 differs from Caestocorbula s. s. only in the degree of elaboration of the siphonal plates and is difficult to distinguish in the absence of this structure. However, these plates are rarely found and relative rostration of the right valve has generally been used as the primary character for identification of Caestocorbula s. l. Recent descriptions, however, tended to include hinge structure, albeit inadequately, for generic identification (e.g., Kozai 1987; Squires and Saul 2004). We also use the genus sensu lato as siphonal plates are not preserved in any specimen of our collection.
The monogeneric subfamily Caestocorbulinae Vokes 1945, ranging from the Cretaceous to the Eocene, was erected based on the presence of calcified siphonal plates in the left valve (Vokes 1945). Some species of Varicorbula are also known to possess calcified siphonal plates, which is why Mikkelsen and Bieler (2001) placed the genus in this subfamily. Evolutionary significance of the siphonal plates has never been explored. The validity of the subfamily Caestocorbulinae based on a single obscure character is questionable.
The present species bears all the diagnostic characters of the genus. Relative rostration of the right valve in the Kutch species is often less prominent than that in many known representatives of the genus. However, the snout is mostly broken in the specimens of our collection. The hinge of the Kutch species closely resembles that of C. gerardi Vincent 1922, the type species of the genus from the Eocene of Belgium, in having a posteriorly directed narrow projecting chondrophore in the left valve with a prominent longitudinal groove running through its middle (e.g., Fig. 3t, ab, ac). However, the chondrophore in our species is horizontal while that in C. gerardi is inclined. The cardinal tooth in the right valve also seems stronger and more projected in the latter than in the Indian form. Socket in the left valve of our species has a distinct floor in its ventral side (e.g., Fig. 3s, ad), which is absent in C. gerardi. The posterior slope in C. gerardi is steeper and delimited by a more pronounced ridge in the left valve. The right valve of C. gerardi bears fewer, thicker and blunter costae than C. gujaratensis n. sp. (Figs. 1–4 in Vincent 1922).
Caestocorbula henckeliusiana (Nyst 1836), from the Oligocene of Belgium and former USSR is more elongated and has a more prominent rostrum in its right valve than C. gujaratensis n. sp. Its right valve also has a distinct angular carina bordering the distal end of the posterior slope, which is lacking in our species. The other Oligocene Caestocorbula from the USSR is Caestocorbula karaschokensis (Merklin 1974). It has a narrower and longer rostrum than in ours. Its chondrophore is large and inclined and ornamentation is not as strong as in the Kutch species (see Merklin 1974).
Caestocorbula? subtumida (Nagao 1928) from the Middle and Upper Eocene of Japan is much smaller, smooth, and has a nepionic cap. Its posterior rostrum is longer and marked by a keel in the right valve (see Matsubara and Ugai 2006). All other Caestocorbula species known to us come from the Cretaceous and differ significantly from our species mostly in having a more elongated snout-like rostrum (Kozai 1987 and references therein; Squires and Saul 2004 and references therein).
C. (Bicorbula) subexarata d’Archiac and Haime 1854 sensu Cox 1931 is conspecific with our species as it resembles C. gujaratensis n. sp. in all features and is different from d’Archiac and Haime’s (1854) species. The latter is more trigonal in shape, apparently does not have a prominent rostrum in its right valve and has much finer commarginal ribs. Rostrum is broken in the specimen figured by Cox (1931, pl. 4, Fig. 14b). Later, Cox (1936) reported the same species from Bahrain (GSI 17368, 17369). None of the specimens is well preserved; one (GSI 17368) is an external mold of the left valve while the other (GSI 17369) is an internal mold of the right valve. The latter is entirely smooth while the former bears sharp commarginal ribs. The size of the Bahrain form is comparable to the present species.
Some species described from the Indian subcontinent as Corbula Bruguière 1797 or Bicorbula Fischer 1887 are closely comparable to C. gujaratensis n. sp. In our opinion, their generic assignments are equivocal. Corbula (Bicorbula) praexarata Cox 1930 sensu Kachhara et al. 2011a is conspecific with C. gujaratensis n. sp. The posterior rostrum of the right valve of the only specimen they recorded is broken, but resembles our species in all respects including size, shape and ornamentation. B. praexarata (Cox 1930) is characterized by a prominently carinate posterior slope in the right valve, which is absent in C. gujaratensis n. sp. The postero-dorsal margin of the left valve of the former is more steeply inclined than the latter. Commarginal ribs in the right valve of B. praexarata are stouter, closer and flat-topped whereas in C. gujaratensis n. sp. ribs are higher, round-topped, and more distantly spaced. Internal features of B. praexarata are not known. C. (bicorbula) subexarata var. lituus de Cotter 1923 reported by Kachhara et al. (2011b) from a higher stratigraphical horizon of Kutch is equivocal. The figured specimen (Kachhara et al. 2011b, pl. 1, Fig. 7) can be the internal mold of C. gujaratensis n. sp.
Corbula harpa d’Archiac and Haime 1854 from the Palaeocene of Pakistan has a prominent angular posterior carina in the right valve and its left valve is essentially smooth. Corbula daltoni de Cotter 1923 from the Upper Eocene of Myanmar—smaller than C. harpa—is also characterized by the angular carina in the right valve and nearly smooth left valve. C. gujaratensis n. sp., in contrast, does not bear a posterior carina in the right valve and its left valve is ornamented with prominent close-spaced commarginal ribs. The latter is also larger than the two former species.
Bicorbula tjiguhanensis (Martin 1922) from the Miocene of Java is comparable in shape with C. gujaratensis n. sp. However, the Java species differs in having an essentially smooth left valve with only some radial ridges. This valve bears a more prominent posterior carina than that of the latter species. The hinge of the left valve in B. tjiguhanensis is characterized by an inclined chondrophore separated from the cardinal socket by a vertical ridge whereas the chondrophore in C. gujaratensis n. sp. is horizontal and projected. The right valve of the Java species is more strongly rostrate and its cardinal tooth is more projected than the Kutch species (see Leloux and Wesselingh 2009).
Measurements Table 2.
Genus Bicorbula Fischer 1887
Type species Corbula gallica Lamarck 1801. By monotypy, Eocene of France.
Bicorbula kutchensis n. sp.
(Fig. 4)

B. kutchensis n. sp. from the Eocene Harudi Formation. a–c PG/Cb 151 (paratype)—external view of left valve (a), external view of right valve (b), dorsal view (c). d PG/Cb 24 (paratype), dorsal view. e–g PG/Cb 359 (paratype)—external view of left valve (e), external view of right valve (f), dorsal view (g). h–i PG/Cb 368 (paratype)—external view of left valve (h), external view of right valve (i). j–l PG/Cb 500 (holotype)—external view of left valve (j), external view of right valve (k), dorsal view (l). m–n PG/Cb 502 (paratype), dorsal view (m) and its close-up with right valve placed below left valve (n). o–p PG/Cb 369 (paratype)—external view of left valve (o), external view of right valve (p). q PG/Cb 506 (paratype), dorsal view. r–t PG/Cb 501 (paratype)—dorsal view (r), external view of right valve (s), external view of left valve (t). u PG/Cb 509 (paratype), dorsal view. C cardinal tooth, R chondrophore marked by arrow. Specimens except close-up (n) coated with magnesium oxide. Scale bar for n = 5 mm, for the rest = 10 mm
2011 Corbula (Bicorbula) cf. pseudorakhiensis Eames—Kachhara et al. p. 87, pl. 1, fig. 10.
Type specimens
Holotype: PG/Cb 500; paratypes: PG/Cb 24, 151, 359, 368, 369, 501, 502, 506, 509.
Other material examined
More than hundred specimens, some with original shell, in the collection of Presidency University.
Type locality, horizon and age
Loc. 4, about 2 km north of village Harudi (23°32′N, 68°40′52″E) (Fig. 1, Table 1), Harudi Formation, lower Middle Eocene.
Etymology
The species derives its name from the Kutch district, Gujarat, India from where it is being reported.
Diagnosis
Large, ovate-trigonal, strongly inequivalved shell; left valve smaller, less convex and less rostrate; hinge with a large cardinal tooth projected beyond beak in the right valve and a horizontal posteriorly directed grooved chondrophore in the left valve; surface of left valve characterized by prominent commarginal ribs while that of right valve essentially smooth.
Description
Shell large (maximum length observed 44 mm), ovate-trigonal, inequivalved, slightly elongated and rostrate towards posterior; right valve with high umbo projected beyond its hinge and incurved prosogyral beak; umbo relatively low in left valve, beak of left valve prosogyral; anterior slope wide, not clearly demarcated; posterior slope obscure, except a blunt ridge only in left valve; hinge bears a strong tooth in right valve that projects well beyond its beak and a corresponding socket in left; a large horizontal chondrophore extends posteriorly from left valve into a deep resilifer in hinge of right valve, a deep longitudinal groove divides its surface into a broad anterior and a narrow posterior section, anterior section anteriorly inclined and has rounded margin and its surface commarginally striated, posterior section bounded by a strong anterior ridge and a narrow sharp posterior ridge; adductor muscle scars subequal in area; pallial sinus quite prominent, shallow; surface of left valve ornamented by close-spaced sharp fine commarginal ribs and faint radial lines while right valve essentially smooth with distantly spaced fine sharp commarginal lines; faint radial lines also discernible in inner shell layer; right valve bears a small antero-ventral notch; shell thick.
Discussion
The present species has all the characteristics of the genus such as an ovate-trigonal shape, a right valve with strongly incurved prosogyral beak and highly elevated umbo, a hinge with strong and projected cardinal tooth and a large protuberant chondrophore (Anderson and Roopnarine 2003). The chondrophore of the present species is typical of the genus having a rounded outer margin and a deep groove separating a wide anterior section from a narrow posterior section (e.g., Fig. 4m, n, u). The posterior part bears two ridges—a thick one at its anterior border and a fine sharp one at the posterior border. The genus is characterized by almost smooth valves. Our species has a smooth right valve while the left valve is ornamented with fine commarginal ribs. The type species of the genus, B. gallica (Lamarck 1801) resembles B. kutchensis n. sp. but has a more elongated left valve where posterior carina is more prominent. In B. kutchensis n. sp., the left valve somewhat abruptly becomes depressed towards posterior. The shell of B. gallica is also thinner than that of the Kutch species.
We place Corbula (Bicorbula) cf. pseudorakhiensis Eames 1951 sensu Kachhara et al. 2011a in the synonymy of the present species with some hesitation. The only specimen reported by Kachhara et al. (2011a) does not reveal the internal features and is much delaminated and hence, surface features are not detectable. However, it resembles B. kutchensis n. sp. in size and all shape parameters. C. pseudorakhiensis Eames 1951 is much smaller and only slightly inequivalved whereas the present species is very large and strongly inequivalved. C. pseudorakhiensis is characterized by strong posterior carinae in both right and left valves whereas in B. kutchensis n. sp. there is only an obscure carina in the left valve.
The present species is closely comparable to Bicorbula subexarata (d’Archiac and Haime 1854) in many respects including shell shape, nature of beak, and umbo, and in the absence of nepionic cap and posterior carina. However, d’Archiac and Haime’s (1854) species is much smaller and has a narrower and more elongated posterior part. The posterior slope in it is also steeper than that in B. kutchensis n. sp. Ornamentation also differs slightly in being similar in both the valves of B. subexarata whereas in B. kutchensis n. sp. it is discrepant with prominent fine sharp commarginal ribs in the left valve whereas the right valve is essentially smooth. d’Archiac and Haime (1854) reported some differences between their Sind form and those found from Subathu beds, Himachal Pradesh, India. Both valves of the Subathu form is covered by rather strong commarginal ribs and majority of these are smaller than the Sind form. They were in doubt in placing this form in B. subexarata. Mathur (1975) reported six specimens of B. subexarata from the Subathus, all of which are much smaller than the present Kutch species. The only figure he provided resembles our species in shape but the posterior part of the figured specimen is broken and the features of the left valve, as revealed in the figure, are not adequate for specific identification. Tandon (1971, 1976) recorded ‘Corbula subexarata’ from the same stratigraphical level of Kutch (C. subexarata Zone of Tandon 1971) as our species. However, these two cannot be compared as figure or description of the former is not available.
B. subexarata remains an ill-defined species and has been the source of much confusion. B. subexarata and its varieties have been reported from several parts of the Indian subcontinent. Eames (1951) reported it from the Punjab area in Pakistan. Cox (1931, 1936) reported the species from Pakistan and Bahrain, which we have included in C. gujaratensis n. sp. de Cotter (1923) described C. subexarata var. lituus de Cotter 1923 from the Eocene of Myanmar. Among his figured specimens, two are close to our species (de Cotter 1923, pl. 1, Figs. 2, 3) although much smaller and no information about their hinge is available. The other two specimens (de Cotter 1923, pl. 1, Figs. 4, 5) are, however, reminiscent of C. gujaratensis n. sp. Iqbal (1969a, b) reported B. subexarata from different Eocene deposits of Pakistan. The photographs provided by him are inadequate to ascertain their generic and specific identity.
Bicorbula vredenburgi (Cossmann and Pissarro 1927) from the Eocene of Pakistan is distinctly more rostrate than our species and its posterior slope is more clearly demarcated by a carina. However, its surface ornamentation is comparable to ours. B. lunica (Iqbal 1969a) from the Lower Eocene of West Pakistan is much smaller than our species and has a different shell outline. B. ickeae (Martin 1914) from the Middle Eocene of Java resembles the present Kutch species in size, shape and ornamentation. However, the chondrophore of the Java form has a narrower and more elevated anterior section than that in our species. Its posterior margin is more truncated and postero-dorsal margin is somewhat higher than that of the Kutch species.
Measurements Table 3.
Genus Lenticorbula Merklin 1974
Type species Corbula helmerseni Mikhailovsky 1912 by original designation, Lower Oligocene of Romania, Upper Oligocene of Turkmenistan and Kazakhstan, and Lower Miocene of Turkmenistan.
Lenticorbula trigonalis (Sowerby 1840), comb. nov.
(Fig. 5)

L. trigonalis (Sowerby 1840) from the Miocene Chhasra Formation. a–c PG/Cc 12—external view of left valve (a), external view of right valve (b), dorsal view (c). d–f PG/Cc 15—external view of left valve (d), external view of right valve (e), dorsal view (f). g–i PG/Cc 250—external view of left valve (g), dorsal view (h), external view of right valve (i). j–l PG/Cc 199—external view of left valve (j), external view of right valve (k), dorsal view (l). m–n PG/Cc 133—external view of left valve (m), dorsal view (n). o–q PG/Cc 197—external view of left valve (o), external view of right valve (p), dorsal view (q). r PG/Cc 52, external view of left valve. s PG/Cc 38, dorsal view. t–v PG/Cc 259—external view of left valve (t), external view of right valve (u), dorsal view (v). w PG/Cc 262, chondrophore in close–up. x PG/Cc 261, left valve, dorsal view. y–z PG/Cc 260, right valve—internal view (y), dorsal view (z). aa–ac PG/Cc 74—external view of left valve (aa), external view of right valve (ab), dorsal view (ac). C cardinal tooth, R chondrophore marked by arrow. Specimens except in w–z coated with magnesium oxide. Scale bar for w = 2 mm, for the rest = 10 mm
1840 Corbula trigonalis Sowerby pl. 25, fig. 4.
1854 C. trigonalis d’Archiac and Haime p. 235, pl. 16, figs. 6-7.
1879 C. trigonalis Martin p. 93, pl. 15, figs. 12-13.
1928a C. trigonalis Vredenburg p. 461.
Material examined
Two hundred fifteen articulated specimens and several single valves; all from Loc. 6 in Kankawati River near village Vinjan (23°06′N, 69°02′52″E) (Fig. 1, Table 1), Chhasra Formation, Lower Miocene, in the collection of Presidency University. GSI 13535, three articulated specimens.
Description
Shell small (maximum length observed 18.4 mm), trigonal, sub-equivalved with slightly larger right valve, posterior rostrate, posterior side only slightly larger than anterior, shape variable from quite elongated to only slightly elongated and from quite wide to narrow; beaks slightly prosogyral, umbo high but not projected beyond hinge; posterior slope well-demarcated by a strongly angular carina; postero-dorsal margin straight, antero-dorsal margin curved, anterior margin narrowly curved, venter curved; hinge narrow, curved, in right valve contains a strong wedge-shaped cardinal tooth projected beyond hinge margin, behind it a narrow deep resilifer pit; in left valve a socket corresponding to right valve cardinal tooth on a platform at valve interior but without a ventral floor and a horizontally projected chondrophore to its posterior, surface of chondrophore bears two closely spaced posteriorly directed ridges separated by a thin shallow groove, chondrophore margin straight; a narrow groove marking the position of left valve seen on both sides of hinge in right valve; adductor muscle scars, pallial line and sinus not detected; shell surface essentially smooth with incipient to somewhat coarse commarginal ribbing, especially in later ontogeny, often irregularly spaced.
Discussion
C. trigonalis Sowerby 1840 was reported from the Lower Miocene of Kutch, Sind (Sowerby 1840; d’Archiac and Haime 1854; Vredenburg 1928a) and Java (Martin 1879). Kutch is the type area. The specimens referred by Vredenburg (1928a, p. 461) (GSI 13535, three specimens) were examined by us. Our species resembles them in all respects such as size, shape, rostration, and possession of posterior angular carina and fine surface ornamentation, and is considered conspecific. They also come from the same horizon and area of Kutch. However, we have a large collection of specimens, which exhibits a much larger intraspecific variation than was shown by the specimens of Vredenburg (1928a). Vredenburg (1928a) did not describe and illustrate the species. The descriptions provided by Sowerby (1840) and d’Archiac and Haime (1854) were inadequate. That made this revision all the more necessary. Here the species is reassigned to Lenticorbula Merklin 1974. The form shows all the typical features of the genus, such as, small size, triangular shape, strong carination of the posterior, a strong upturned wedge-like cardinal tooth in the right valve (e.g., Fig. 5y, z) and a chondrophore with two prominent ridges through its middle separated by a shallow narrow groove (e.g., Fig. 5w).
The Kutch form resembles the type species, Lenticorbula helmerseni (Mikhailovsky 1912), which comes from the Upper Oligocene and the Lower Miocene of former USSR and the Lower Oligocene of Romania. However, the latter has a narrower umbonal angle, and its chondrophore is larger and inclined while that in L. trigonalis is smaller and nearly horizontal. Anterior section of its chondrophore is more deeply impressed than that in L. trigonalis and is bordered by a sharp anterior ridge.
L. helmerseni major (Merklin 1974) from the Oligocene of former USSR is larger than the Kutch form and has a thicker shell. Among several Oligocene forms from the former USSR the one that is extremely close to L. trigonalis is L. sokolovi slussarevi (Merklin 1974). The latter is known by more than 100 specimens and resembles L. trigonalis in almost all respects except being slightly smaller. Maximum length recorded in it is 13 mm while L. trigonalis reaches up to 18.4 mm. We feel, further studies are required before uniting the two species. L. katherinae (Popov et al. 1993) from the Lower Oligocene of Georgia is more elongated and has a thicker shell than the present species.
The other form described from a coeval Miocene horizon of Kutch by Vredenburg (1928a), C. tunicosulcata Vredenburg 1928a (GSI 13434–13438), is similar in size and shape with the present species. It however, differs in having regularly spaced prominent commarginal costae and a nepionic cap. Ornamentation on the nepionic cap is finer than the rest of the shell surface. In C. tunicosulcata the escutcheon is marked by an angular ridge that occurs between the carina and the postero-dorsal margin. The ridge is more prominent in the right valve. The hinge of the right valve of this species bears a short peg-like cardinal tooth, which is barely projected beyond the beak whereas the tooth is much stronger, wedge-shaped and projected in L. trigonalis. Hinge structure of the left valve of C. tunicosulcata is not known.
Some of the Corbula Bruguière 1797 species described by Eames (1951) from the Eocene of Punjab, Pakistan resemble the present species in external morphology. However, no information on their hinge structure is known. Moreover, they come from an older horizon.
Palaeobiogeography
Distribution patterns of the studied fauna of Kutch
C. gujaratensis n. sp. and B. kutchensis n. sp. appear in the oldest Cenozoic marine deposit exposed in Kutch. This part of the sequence is assigned to the Palaeocene age in recent literature (Chattoraj et al. 2009; Kachhara et al. 2011a). Both the species continue to the lower Middle Eocene beds of Kutch, i.e. C. subexarata Zone (sensu Tandon 1971). C. gujaratensis n. sp. is also known from the lower Middle Eocene of Sind and the Middle Eocene of Bahrain (Cox 1931, 1936). B. kutchensis n. sp. is a close ally of B. subexarata (d’Archiac and Haime 1854). The latter is known to occur widely in the Eocene deposits of the Indian subcontinent. Mathur (1988) found B. subexarata the most widespread bivalve in India during the Eocene. It has been reported from Himachal Pradesh, Haryana, Rajasthan, and Gujarat (Mathur 1988 and references therein). The species was known from the Lower and Middle Eocene from several areas of Pakistan (d’Archiac and Haime 1854; Eames 1951; Iqbal 1969a, b) and the Upper Eocene of Myanmar (de Cotter 1923). The species was also recorded beyond the Indian subcontinent from the Middle Eocene of Egypt and Somalia (see Eames 1951) (see Table 4, Fig. 6).

Palaeobiogeographical distribution of the Middle Eocene corbulids of India and Pakistan shown in palaeocoastline map of Smith et al. (1994)
L. trigonalis (Sowerby 1840) and C. tunicosulcata Vredenburg 1928a come from the Lower Miocene of Kutch. The species were known from the contemporaneous strata of Sind (Vredenburg 1928a; Iqbal 1980). The former was also reported from the Lower Miocene of Java (Martin 1879) whereas the latter also comes from the Miocene of Myanmar (Noetling 1901) (see Table 4, Fig. 7). The distribution of corbulids from Kutch reveals that this Indian basin housed a similar corbulid fauna as that of Pakistan during the Cenozoic. This also roughly suggests a shift in affinity of this basin in terms of corbulid fauna from African-Arabian in the Eocene to western Pacific in the Miocene.

Palaeobiogeographical distribution of the Lower Miocene corbulids of India and Pakistan shown in palaeocoastline map of Smith et al. (1994)
Patterns from the Cenozoic corbulids of the Indian subcontinent
Three corbulid genera are common to the Cenozoic of the Indian subcontinent—Corbula Bruguière 1797, Bicorbula Fischer 1887, and Varicorbula Grant and Gale 1931. These genera are cosmopolitan (e.g., Keen 1969). Only Corbula and Bicorbula were reported from Kutch (Vredenburg 1928a; Tandon 1971, 1976; Kachhara et al. 2011a). Caestocorbula Vincent 1910 and Lenticorbula Merklin 1974 were not known from the Indian subcontinent. Caestocorbula is a cosmopolitan genus whereas Lenticorbula is known mainly from different parts of the former USSR (Keen 1969; Merklin 1974).
Bivalves including corbulids are much more diverse and better known from Pakistan than from any Indian basin. An analysis of the palaeobiogeographical distribution of corbulids from Pakistan reveals that out of 21 species of corbulids known to us from Pakistan, only 7 have also been reported from outside Pakistan. Of the rest, which come exclusively from Pakistan, 12 are from the Palaeocene–Eocene. B. subexarata is the most widespread form there followed by Varicorbula harpa (d’Archiac and Haime 1854). All other species are geographically restricted, often to a small part of the basin. A similar picture emerges from an analysis of all the benthic molluscs known from the Palaeocene–Eocene of Pakistan based on Eames (1951, 1952) and Iqbal (1969a, b, 1972)—out of 231 bivalve species, only 34 are known from outside Pakistan and only 15 from India and out of 233 gastropods only 18 are also known from outside Pakistan including 8 from India. Two features of this distribution scenario are worth mentioning—first, western India and Pakistan display the maximal faunal similarity and second, the Pakistan fauna is characterized by strong endemism. The first feature entails inclusion of western India and Pakistan in a single palaeobiogeographical province. The Western Indian Province (sensu Harzhauser 2007) of the Oligocene may, hence, be considered to be taking shape already since the Lower Palaeogene.
Out of the two Eocene corbulids that also occur beyond this province only B. subexarata was reported from the Indo-Malayian Province (sensu Martin 1931) whereas both were known from western Tethys basins of African and Middle East countries (see Table 4, Fig. 6). This roughly indicates that the Western Indian Province shared greater faunal similarity with the western Tethys basins, especially African and Arabian ones than with the southeast Asian basins during the Palaeogene. V. harpa, known from the Eocene in this province comes from the Upper Cretacous of Lybia, indicating an older relationship between these areas.
In the Neogene, distribution of corbulids reveals a different picture. The two species that have been reported from Kutch and Sind share affinity to the faunas from southeast Asia. Other known corbulids corroborate this pattern of affinity. Corbula acuticosta Martin 1885 from the Miocene of Java was also recorded from the Pliocene of Pakistan (Vredenburg 1928a). Out of the three corbulid species known from the Miocene of Kerala, India two—Varicorbula scaphoides (Hinds 1843) and Anisocorbula socialis (Martin 1879)—are also known from the Miocene of Java. The latter also comes from the contemporaneous horizons of Myanmar (Noetling 1901; Dey 1961; Leloux and Wesselingh 2009) (see Table 4, Fig. 7). Therefore, a shift in the affinity of the western Indian basins from western Tethys in the Palaeogene to western Pacific in the Neogene, as opined by Harzhauser et al. (2002, 2007, 2009), is a strong possibility. The Lower Miocene gastropod fauna from Kutch also displays similar affinity to western Pacific faunas from Java and Sumatra (Kulkarni et al. 2010).
It is pertinent to recall here that in contrast to the general faunal similarity within this province constituted of western Indian and Pakistan basins through the Cenozoic a high specific endemism of the fauna to certain parts of the province especially in Pakistan cannot be overlooked (see also Halder 2012). Out of the 13 Palaeogene corbulids restricted to Pakistan 7 are known from only a single locality, 5 from two localities and the other from three localities. Kulkarni et al. (2010) observed that the Lower Miocene gastropod fauna from Kutch has high endemism—13 % of the gastropod species are endemic to Kutch whereas 47 % are endemic to the western part of the Indian subcontinent. Nautiloids, although relatively rare and nektonic, also display similar restricted occurrences (Halder 2012). Apparently, endemic faunas evolved rapidly and lived simultaneously in these geographically closely linked basins.
Conclusions
-
1.
Corbulid bivalves are known from different parts of the Indian subcontinent through the Cenozoic, most prominently from Pakistan. However, a modern revision of the species is wanting. Systematics, particularly generic status of many of the previously known species is doubtful. A thorough revision of all these forms is not possible because most of the earlier collections are inaccessible. We have made large collection of three species from the Cenozoic succession of Kutch. A detailed systematic revision of these is done here. We have studied C. tunicosulcata from repository specimens. Other species known from Kutch are based on very few specimens and their systematic studies were not rigorous. These species are strongly doubtful and are not considered here for the palaeobiogeographical analysis.
-
2.
Palaeobiogeography of the Indian and Pakistan corbulids is studied here. Similar forms seem to prevail in these basins. Hence, the Western Indian Province constituted of all these Pakistan and western Indian basins, as proposed by Harzhauser (2007) for the Oligocene, may be considered to have evolved earlier during the Lower Palaeogene. This province showed some resemblance in corbulid composition to the African-Arabian Province (sensu Harzhauser 2007) during the Lower Palaeogene.
-
3.
The most striking feature of this distribution pattern, however, is the strong provincialism of the fauna during the Lower Palaeogene. Often, the species are localized and not distributed throughout the province.
-
4.
The Neogene corbulids of this province are not so restricted. They show clear affinity to the southeast Asian corbulids.
References
Anderson LC, Roopnarine PD (2003) Evolution and phylogenetic relationships of Neogene Corbulidae (Bivalvia; Myoidea) of Tropical America. J Paleontol 77:1086–1102
Banerjee S, Chattoraj S, Saraswati PK, Dasgupta S, Sarkar U (2012) Substrate control on formation and maturation of glauconites in the Middle Eocene Harudi Formation, western Kutch, India. Mar Pet Geol 30:144–160
Barron EJ, Peterson WH (1991) The Cenozoic ocean circulation based on ocean General Circulation Model results. Palaeogeogr Palaeoclimatol Palaeoecol 83:1–28
Biswas SK (1992) Tertiary stratigraphy of Kutch. J Palaeontol Soc India 37:1–29
Borkar VD, Kulkarni KG, Bhattacherjee S (2004) Molluscan fauna from the Miocene sediments of Kachchh, Gujarat, India—Part 1. Oysters. Geophytol 34:1–7
Bruguière JG (1797) Plates 190-286. In: Lamarck JB de and Saint-Vincent B (1791–1827) (eds) Tableau Encyclopédique et Méthodique des Trois Règnes de la Nature; vers, Coquilles, Mollusques, et Polypiers. Agasse, Paris
Chattoraj SL, Banerjee S, Saraswati PK (2009) Glauconites from the Upper Palaeocene–Lower Eocene Naredi Formation, western Kutch and their genetic implications. J Geol Soc India 73:567–574
Cossmann M, Pissarro G (1909) The Mollusca of the Ranikot Series. Part 1. Cephalopoda and Gastropoda. Palaeontol Indica, N Ser 3(1):1–83
Cossmann M, Pissarro G (1927) The Mollusca of the Ranikot Series. Part 2. Brachiopoda and Lamellibranchia. Palaeontol Indica, N Ser 10(2):1–31
Cox LR (1930) The fossil fauna of the Samana Range and some neighbouring areas: Part VIII. The Mollusca of the Hangu Shales. Palaeontol Indica, N Ser 15:129–222
Cox LR (1931) A contribution to the molluscan fauna of the Laki and basal Kirthar Groups of the Indian Eocene. Trans R Soc Edinb 57(1):25–92
Cox LR (1936) Fossil Mollusca from southern Persia (Iran) and Bahrein Island. Palaeontol Indica, N Ser 22(2):1–69
d’Archiac AS, Haime J (1854) Description des Animaux Fossiles du Groupe Nummulitique de l’Inde, Seconde livraison: Mollusques. Gide et J. Baudry, Paris, pp 225–373
de Cotter GP (1923) The Lamellibranchiata of the Eocene of Burma. Palaeontol Indica, N Ser 7(2):1–39
Dey AK (1961) The Miocene Mollusca from Quilon, Kerala (India). Palaeontol Indica, N Ser 36:1–119
Eames FE (1951) A contribution to the study of the Eocene in western Pakistan and western India. B. The description of the Lamellibranchia from standard sections in the Rakhi Nala and Zinda Pir areas of the western Punjab and in the Kohat District. Philos Trans R Soc Lond, Ser B, Biol Sci, no. 627, 235:311–482
Eames FE (1952) A contribution to the study of the Eocene in western Pakistan and western India. C. The description of the Scaphopoda and Gastropoda from standard sections in the Rakhi Nala and Zinda Pir areas of the western Punjab and in the Kohat District. Philos Trans R Soc Lon, Ser B, Biol Sci 631(236):1–168
Fischer PH (1887) Manuel de Conchyliologie et de Paleontologie Conchyliologique: ou, Histoire Naturelle des Mollusques vivants et Fossiles, Fascicule 11. F. Savy, Paris, pp 1009–1369
Garg R, Prasad V, Thakur B, Singh IB, Ateequzzaman K (2011) Dinoflagellate cysts from the Naredi Formation, southwestern Kutch, India: implications on age and palaeoenvironment. J Palaeontol Soc India 56:201–218
Grant US IV, Gale HR (1931) Catalogue of the marine Pliocene and Pleistocene Mollusca of California and adjacent regions. Mem San Diego Soc Nat Hist 1:1–1036
Haas O, Miller AK (1952) Eocene nautiloids of British Somaliland. Bull Am Mus Nat Hist 99:313–354
Halder K (2012) Cenozoic fossil nautiloids (Cephalopoda) from Kutch, western India. Palaeoworld 21:116–130
Harzhauser M (2007) Oligocene and Aquitanian gastropod faunas from the Sultanate of Oman and their biogeographic implications for the early western Indo-Pacific. Palaeontogr 280:75–121
Harzhauser M, Piller WE, Steininger FF (2002) Circum-Mediterranean Oligo-Miocene biogeographic evolution—the gastropods’ point of view. Palaeogeogr Palaeoclimatol Palaeoecol 183:103–133
Harzhauser M, Kroh A, Mandic O, Piller WE, Göhlich U, Reuter M, Berning B (2007) Biogeographic responses to geodynamics: A key study all around the Oligo–Miocene Tethyan Seaway. Zool Anz 246:241–256
Harzhauser M, Reuter M, Piller WE, Berning B, Kroh A, Mandic O (2009) Oligocene and Early Miocene gastropods from Kutch (NW India) document an early biogeographic switch from Western Tethys to Indo-Pacific. Paläontol Z 83:333–372
Hinds RB (1843) On new species of Corbula and Potamomya. Proc Zool Soc Lon 11:55–59
Iqbal MWA (1969a) Mega-fauna from the Ghazij Formation (Lower Eocene) Quetta Shahrig area, West Pakistan. Palaeontol Pak 5:1–41
Iqbal MWA (1969b) The Tertiary pelecypod and gastropod fauna from Drug, Zindapir, Vidor (district D. G. Khan), Jhalar and Chharat (district Campbellpore), West Pakistan. Palaeontol Pak 6:1–95
Iqbal MWA (1972) Paleocene bivalve and gastropod fauna from Jherruk-Lakhra-Bara Nai (Sind), Salt Range (Punjab) and Samana Range (N. W. F. P.), Pakistan. Palaeontol Pak 9:1–105
Iqbal MWA (1980) Oligo-Miocene bivalves and gastropods from Kirthar Province, Lower Indus Basin, Pakistan. Rec Geol Surv Pak 51:1–59
Kachhara RP, Devi KB, Jodhawat RL (2011a) Molluscan assemblage from the marine Palaeocene sequence in southwestern Kachchh, Gujarat. J Geol Soc India 78:81–91
Kachhara RP, Jodhawat RL, Devi KB (2011b) Molluscan biostratigraphy of the Palaeogene sediments around Lakhpat, Kachchh, Gujarat, India. J Palaeontol Soc India 56:17–28
Kachhara RP, Jodhawat RL, Devi KB (2012) Lower Oligocene bivalves of Ramanian Stage from Kachchh, Gujarat, India. J Earth Syst Sci 121:405–438
Keen M (1969) Superfamily Myacea Lamarck, 1809. In: Moore RC (ed) Treatise on Invertebrate Paleontology. Part N, Mollusca 6, Bivalvia. Geol Soc Am Inc and Univ Kans, Kansas pp 690-699
Kozai T (1987) Cretaceous Caestocorbula (Bivalvia) from Japan. Trans Proc Palaeontol Soc Jpn, N Ser 148:326–334
Krishnan MS (1982) Geology of India and Burma, 6th edn. CBS, New Delhi
Kulkarni KG, Bhattacharjee Kapoor S, Borkar VD (2010) Molluscan fauna from the Miocene sediments of Kachchh, Gujarat, India—Part 3. Gastropods. J Earth Syst Sci 119:307–341
Lamarck JB de (1801) Systême des Animaux sans Vertèbres. Mus Hist Nat, Paris
Lamarck JB de (1818) Histoire Naturelle des Animaux sans Vertèbres, Présentant les Caractères Généraux et Particuliers de ces Animaux, leur Distribution, leurs Classes, leurs Familles, leurs Genres, et la Citation des Principales Espèces qui s'y Rapportent; Précédée d'une Introduction Offrant la Détermination des Caractères Essentiels de l'Animal, sa Distinction du Végétal et des Autres Corps Naturels, enfin, l'Exposition des Principes Fondamentaux de la Zoologie. Tome 5. Deterville, Paris
Leloux J, Wesselingh FP (2009) Types of Cenozoic Mollusca from Java in the Martin Collection of Naturalis. Natl Natuurhist Mus Tech Bull 11:1–765
Linnaeus C (1758) Systema Naturæ per Regna Tria Naturæ, Secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Tomus 1. Holmiæ, Stockholm
Lyngdoh BC, Tiwari RP, Kachhara RP (1999) Miocene molluscan biostratigraphy of the Garo Hills, Meghalaya India. J Palaeontol Soc India 44:55–67
Martin K (1879) Palaeontologischer Theil: A. Einleitung, B. Systematische Aufzählung und Beschreibung der Petrefacten: I. Cephalopoda, II. Gastropoda, III. Conchifera, IV. Brachiopoda. In: Martin K (1879-1880) Die Tertiärschichten auf Java. Nach den Entdeckungen von Fr. Junghuhn. E. J. Brill, Leiden, pp 1-126
Martin K (1885) Systematischer Theil. I. Glossophora. B. Scaphopoda;II. Lamellibranchiata; Molluscoidea; Vermes; Echinoidea. In: Martin K(1883-1887) Palaeontologische Ergebnisse von Tiefbohrungen auf Java, nebst Allgemeineren Studien ueber das Tertiaer von Java, Timor und Einiger Anderer Inseln. Sammlungen des Geologischen Reichs-Museums in Leiden, Serie 1. Beitr Geol Ost-Asiens Aust 3:185–291
Martin K (1914) Die Fauna des Obereocäns von Nanggulan auf Java. Samml Geol Reichs-Mus Leiden. N F 2:107–200
Martin K (1922) Die Molluscen der Njalindungschichten, Gasteropoda (Fortsetzung), Scaphopoda, Lamellibranchiata. Allgemeiner Theil. Samml Geol Reichs-Mus Leiden. N F 1(2–4):471–496
Martin K (1931) Wann löste sich das Gebiet des Indischen Archipels von der Tethys. Leidsche Geol Meded 4:1–8
Mathur NS (1975) Mollusca from the Subathu Formation (Upper Palaeocene-Middle Eocene), Simla Hills. Bull Indian Geol Assoc 8:1–40
Mathur UB (1988) Marine Tertiary larger invertebrate fauna of India—a review. Geol Surv India Spec Publ 11(2):63–88
Matsubara T, Ugai H (2006) Paleoecology and taxonomy of Middle Eocene molluscs from the Shiratake Formation on Takeshima Island, Amakusa Islands, southwest Japan. Bull Goshoura Cretac Mus 7:1–17
Merklin RL (1974) Opredelitel’ dvustvorchatykh mollyuskov oligotsena yuga SSSR (Handbook of Oligocene Bivalvia of the South of the USSR). Trans Palaeontol Inst, Acad Sci USSR 145:1–189
Mikhailovsky GP (1912) Some new and rare shells from the Tertiary sediments of the northern coast of the Aral Sea. Minutes Soc Nat Tartu Univ 21(1–2):120–138
Mikkelsen PM, Bieler R (2001) Varicorbula (Bivalvia: Corbulidae) of the Western Atlantic: taxonomy, anatomy, life habits, and distribution. Veliger 44:271–293
Nagao T (1928) Palaeogene fossils of the Island of Kyûshû, Japan. Part 1. Sci Rep Tôhoku Imp Univ, Second Ser (Geol) 9:97–128
Noetling F (1901) The fauna of the Miocene beds of Burma. Palaeontol Indica, N Ser 1(3):1–378
Nyst PH (1836) Recherches sur les coquilles fossils de Houselt et de Kleyn-Spauwen (Province du Limbourge). Messag Sci Arts Belg 4:139–181
Popov SV (1993) Zoogeography of the Upper Eocene basins of Western Eurasia based on bivalve molluscs. Stratigr Geol Correl 2:103–118
Popov SV, Voronina АА, Goncharova IA (1993) Stratigraphy and bivalves of the Oligocene—Lower Miocene of the Eastern Paratethys. Trans Paleontol Inst, Russ Acad Sci 256:1–207 (in Russian)
Smith AG, Smith DG, Funnell BM (1994) Atlas of Mesozoic and Cenozoic coastlines. Cambridge University Press, Cambridge
Sowerby JC (1840) Fossils from the Tertiary Formations, Cutch. Trans Geol Soc Lond, Ser 2, 5:pls. 25–26
Squires RL, Saul LR (2004) Cretaceous corbulid bivalves of the Pacific Slope of North America. Veliger 47:102–129
Stoliczka F (1870-1871) Cretaceous fauna of southern India, vol. 3, The Pelecypoda, with a review of all known genera of this class, fossil and Recent. Palaeontol Indica, Ser 6, 3:1-537
Tandon KK (1971) On the discovery of mammalian and reptilian remains from the Middle Eocene rocks of S. W. Kutch, India. Curr Sci 40:436–437
Tandon KK (1976) Biostratigraphical classification of the Middle Eocene rocks of a part of south-western Kutch, India. J Palaeontol Soc India 19:71–88
Vincent E (1890) Sur une plaque appendiculaire observée chez le “Corbula henckeliusi”. Ann Soc R Malacol Belg, Bull Séances 25:7–8
Vincent E (1910) Presence d’un plaque siphonale chez Corbula regulbiensis Morr. Ann Soc R Zool Malacol Belg 44:140–142
Vincent E (1922) Les Corbules de sables de Wemmel. Ann Soc R Zool Malacol Belg 52:94–103
Vlerk M (1931) Caenozoic Amphineura, Gastropoda, Lamellibranchiata, Scaphopoda. Leidsche Geol Meded 5:206–296
Vokes HE (1944) The validity of the molluscan genus Caestocorbula Vincent. Am J Sci 242:614–623
Vokes HE (1945) Supraspecific groups of the pelecypod family Corbulidae. Bull Am Mus Nat Hist 86:1–32
Vredenburg E (1925) Description of Mollusca from the post-Eocene Tertiary formation of North-Western India: Cephalopoda, Opisthobranchiata, Siphonostomata. Mem Geol Surv India 50(1):1–350
Vredenburg E (1928a) Description of Mollusca from the post-Eocene Tertiary formation of North-Western India: Gastropoda (in part) and Lamellibranchiata. Mem Geol Surv India 50(2):351–506
Vredenburg E (1928b) A supplement to the Mollusca of the Ranikot Series. Palaeontol Indica, N Ser 10(4):1–75
Acknowledgments
We are grateful to Laurie C. Anderson (Louisiana State University, USA), Oleg Mandic (Natural History Museum, Vienna, Austraia) and five anonymous reviewers for critical examination of an earlier version of the manuscript and providing valuable suggestions. Irina Goncharova (Russian Academy of Sciences, Moscow, Russia) provided valuable information. The Director and employees of the Curatorial Division, Geological Survey of India (GSI), Kolkata gave access to fossils in the repository of GSI. Ipsita Ghosh (Presidency University) helped in drawing. The first author got financial support from the Department of Science and Technology, India (Project Nos. SR/FTP/ES-49/2003 and SR/S4/ES-653/2012).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Halder, K., Bano, S. Cenozoic Corbulidae (Bivalvia, Mollusca) from the Indian subcontinent—palaeobiogeography and revision of three species from Kutch, India. Arab J Geosci 8, 2019–2034 (2015). https://doi.org/10.1007/s12517-014-1362-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12517-014-1362-6