
Abstract
Sugarcane secrets a reddish pigment presumed to contain phytoalexin compounds in the stalk tissues in response to infection of Colletotrichum falcatum, causing red rot. To identify the phytoalexin compounds involved in red rot resistance, detailed HPLC analyses were conducted using the phytoalexin standards in a set of sugarcane cultivars varying in red rot resistance, after pathogen inoculation. We found nine different 3-deoxyanthocyanidin phytoalexin compounds, which are differentially induced upon C. falcatum infection in sugarcane varieties with varying degrees of disease resistance. Among the nine compounds, four compounds were identified as luteolinidin, 5-methoxy luteolinidin, apigeninidin and arabinosyl-5-O-apigeninidin. Multifold induction of luteolinidin, apigeninidin and arabinosyl-5-O-apigeninidinin were recorded in the stalk tissues in disease resistant cvs Co 93009, BO 91 and Baragua, respectively after pathogen inoculation as compared to the susceptible cv CoC 671. In addition to the known compounds one to three unknown compounds were detected in the resistant cultivars. The susceptible cultivars exhibited a placid induction of phytoalexin compounds after pathogen infections. The results very clearly demonstrated a specific induction of 3-deoxyanthocyanidins in resistant cultivars at a higher level as compared to susceptible cultivars after C. falcatum infection in sugarcane. Total extracts of the induced phytoalexins in the calorimetric assays, also revealed similar results. Probably induction of either luteolinidin or apigeninidin compounds alone or in combination at higher concentrations in the resistant cultivars may enable the effective arrest of pathogen invasion and further development inside the stalk tissues.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Phytoalexins are antimicrobial compounds of low-molecular weight synthesized in planta as they are accumulated in tissues upon pathogen invasion, which represent a module, among the multicomponent antimicrobial defense response induced upon pathogen attack. In plants antimicrobial compounds fall into two categories viz., phytoanticipins and phytoalexins depending upon the accumulating time in the cells (Mansfield 1999). Phytoanticipins are preexisting, low molecular weight antimicrobial compounds, but phytoalexins are synthesized de novo by plants and accumulate rapidly in areas of incompatible pathogen infection (Van Etten et al. 1994). Phytoalexins are chemically diversified among plant kingdom as terpenoids, glycosteroids and alkaloids, the example includes isoflavonoids and pterocarpans in legumes, sulfur-containing indole derivatives in cruciferous plants (Tsuji et al. 1992), sesquiterpenoids in solanaceous plants, and coumarins in umbelliferous plants (Knogge et al. 1987). Phytoalexins produced in planta may act upon invading organisms by affecting the hyphal formation, retarding maturation, disrupting metabolism or preclude reproduction. On recognition of pathogenic invasion, it launches a two-pronged resistance: a general short-term response and a delayed long-term specific response. The common short-term response is the hypersensitive response (HR), where infected cells and its neighbor are signaled to undergo apoptosis in order to prevent pathogen progression further. Upon HR, the plant deploys reactive oxygen species such as superoxide and hydrogen peroxide to kill invading cells followed by the accumulation of phytoalexin at the site of pathogen invasion (Hammerschmidt 1999).The pigmented phytoalexins produced by sorghum are clear exception among the phytoalexins (Snyder and Nicholson 1990). The visibility of these compounds in tissue establishing a role for these compounds in the resistance of sorghum to fungal pathogen Colletotrichum sublineolum (Snyder et al. 1991) and these studies elucidated the biosynthetic pathway for 3-deoxyanthocyanidins. Thus the importance of phytoalexins in plant defense is well proven, but still need to be understood in detail in order to effectively utilize their potential in applied disease resistance programs.
Sugarcane, a major crop cultivated in tropical and subtropical regions across the globe meets the need for sugar and ethanol. It is prone to various diseases, amongst them red rot, a fungal disease caused by Colletotrichum falcatum, causes colossal loss to cane growth and yield (Viswanathan 2010). During the pathogen invasion, a reddish compound is produced at the site of invasion and surrounding tissues (Fig. 1) and this gives the name ‘red rot’ to the disease. In our earlier studies, we found specific accumulation of 3-deoxyanthocyanidins, luteolinidin and apigeninidin in sugarcane after red rot pathogen C. falcatum challenge (Viswanathan et al. 1996a, b; Malathi et al. 2008). As the induction of these compounds in different cultivars will provide more insight into the role played during compatible and incompatible interactions with the host, we further studied 3-deoxyanthocyanidin accumulation in different cultivars varying in disease resistance to red rot through HPLC analysis and calorimetric assays. The outcome shows differential accumulation of the 3-deoxyanthocyanidins in sugarcane cultivars varying in resistance to C. falcatum.

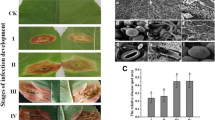
Variation in symptom expression in sugarcane varieties resistant and susceptible to red rot pathogen Colletotrichum falcatum: 30 days after pathogen inoculation by plug method. Arrows indicate point of inoculation
Materials and Methods
Sample Collection
Six month old sugarcane cultivars like CoC 671, CoC 90063, CoC 92061, Co 419, Co 997, Co 1148 and Co 86032 which are susceptible (S), moderately susceptible (MS) and cultivars like Baragua, BO 91, Co 94008 and Co 93009 which are resistant (R) to C. falcatum respectively, were inoculated with 7 days old sporulating cultures of C. falcatum (1 × 106 conidia ml−1) in the field. Inoculation was done by making a bore-hole (12 mm depth and 8 mm diameter) using a red rot inoculator in the middle of the third exposed internode (Viswanathan, 2010). Two drops of the above prepared conidial suspension was dropped into the bore-hole using a Pasteur pipette, the tissue block was replaced into the bore-hole incision, and covered with china clay. While the sterile distilled water was used in place of conidial suspension as mock inoculated samples. The stalk samples were collected at different time point’s after pathogen challenge and immediately stored at −80 °C until processing.
Extraction and Analysis of Phytoalexin Compounds
Tissue samples surrounding the site of inoculation (~150 mg) were removed 24 and 48 h after pathogen inoculation from each variety, and cut into small pieces and immersed in 1 ml of HPLC grade methanol. The pigments were allowed to leach from the tissues for 18 h at 4 °C; later the extracts were filtered through a 0.4 µm membrane and analyzed by HPLC and spectroscopy. The phytoalexins were separated with reverse phase C-18 column using perchloric acid: methanol solvent system (Shimadzu LC-8A). Solvent A was 0.6 % perchloric acid and solvent B was 100 % HPLC-grade methanol. Samples (20 µl) were injected and eluted isocratically with 40 % solvent B at a flow rate of 0.8 ml min−1 and the phytoalexin compounds were detected at 480 nm. Twenty micro litres of crude extract from sorghum mesocotyls extract (generous gift from Late Dr. R. L. Nicholson, Purdue University, West Lafayette, Indiana, USA) was used as the phytoalexin standard (Nicholson et al. 1987). Phytoalexin standards luteolinidin, apigeninidin, and cyanidin (Extrasynthese, France) were dissolved in HPLC-grade methanol, chromatographed and their retention times and peak areas were determined. Retention times of standards were used to identify the 3-deoxyanthocyanidin compounds in the test samples, and the amount of each compound in a sample was determined by plotting its peak area in the standard curve of the respective standard.
In addition, total anthocyanidin content in sugarcane tissues after pathogen inoculation was assayed through spectrophotometric analysis. Here tissue samples were taken 24 h after pathogen inoculation and were processed by the same method as stated above, and the pigment induction was quantified at 480 nm in a spectrophotometer (T80+ UV spectrophotometer, PG Instruments, Germany) and the concentration was expressed as micromoles of 3-deoxyanthocyanidins using the molar extinction coefficient of luteolinidin (13,800 M−1 cm−1).
Results and Discussion
The HPLC analysis revealed induction of nine 3-deoxyanthocyanidin compounds induced differentially across the sugarcane cultivars tested. Among them, 4 were identified as luteolinidin (Retention time (tR) = 16.31 min), 5-methoxy luteolinidin (tR = 19.45 min), apigeninidin (tR = 21.88 min) and Arabinosyl-5-O-Apigeninidin (tR = 24.11 min) using sorghum mesocotyl extract as standard (Nicholson et al. 1987). The remaining 5 unknown compounds were named as X1 (tR = 15.07 min), X2 (tR = 18.33 min), X3 (tR = 20.43 min), X4 (tR = 22.97 min) and X5 (tR = 26.32 min) (Fig. 2). Among the identified compounds (Fig. 3), luteolinidin was found to be expressed at higher levels in resistant cultivars when compared to the susceptible cultivars. The highest induction was noticed in the pathogen inoculated canes of the resistant cv Co 93009, where pathogen inoculation triggered multi-fold induction of luteolinidin, whereas its induction probably suppressed in pathogen inoculated cane tissues of susceptible cv CoC 671. Furthermore its induction was found to be absent in other susceptible cvs Co 419 and Co 997. 5-methoxy luteolinidin was found to be induced only in the susceptible cultivars with a highest induction in the cv CoC 92061 followed by Co 1148 and CoC 671. In Co 86032, which is moderately susceptible to red rot, it was found to be specifically induced after pathogen inoculation.

HPLC chromatograms represent different fractions of phytoalexin compounds in Sorghum mesocotyl extract (reference) and from sugarcane cultivars after Colletotrichum falcatum inoculation a Sorghum mesocotyl extract; b CoC 671; c Co 93009; d Baragua; e BO 91

Induction of different phytoalexins in stalk tissue after pathogen inoculation in varying resistance to Colletotrichum falcatum C—mock inoculated control (48 h), I—pathogen inoculated (48 h), tR—retention time
The resistant cv Co 93009 accumulated highest concentration of luteolinidin (0.0331 µg mg−1 tissue) (~122 µM) followed by another resistant cv BO 91 (0.0133 µg mg−1 tissue) (~49 µM). Whereas the susceptible cultivars accumulated very low levels, nearly 100 folds lower than the resistant cultivars. The susceptible cv CoC 671 recorded 0.0004 µg mg−1 tissue (~1.47 µM) and CoC 92061 recorded 0.0020 µg mg−1 tissue (~7.37 µM). In an in vitro bioassay luteolinidin and 5-methoxyluteolinidin were found to completely inhibit the growth of sorghum anthracnose pathogen C. sublineolum, at concentration of as low as 5–7.5 µM (Lo et al. 1996; Tenkouano et al. 1993). Thus the levels of luteolinidin accumulated in the resistant sugarcane cvs Co 93009 and BO 91 were well above the necessary inhibitory concentrations and may be sufficient to arrest the fungal growth. The suboptimal accumulations of luteolinidin induction in susceptible cultivars might be a major reason for the compatible reaction.
Luteolinidin and 5-methyoxy luteolinidin were found to be significantly more fungitoxic than the other 3-deoxyanthocyanidin phytoalexins (Lo et al. 1996; Nicholson et al. 1987). In sorghum, resistance to C. sublineolum (causal agent of sorghum anthracnose) was correlated to the more complex phytoalexin compounds, in particular luteolinidin and luteolinidin 5-methylether, which were found to be synthesized only in incompatible interactions (Boddu et al. 2004). When infected with C. sublineolum, a resistant sorghum cultivar had a greater and faster accumulation of phytoalexins than a susceptible cultivar (Lo et al. 1999). Snyder et al. (1991) revealed the accumulation of luteolinidin in cells responding to pathogen infection at concentrations in the range of 0.48–1.20 ng cell−1 in incompatible interaction of sorghum-C. sublineolum. The phytoalexin inclusions in the resistant sorghum cultivar were found to be more darkly pigmented than those of susceptible cultivar. In sugarcane also similar observations were reported (Viswanathan et al. 1996a, b). Since luteolinidin has a higher absorption maximum than apigeninidin and its compounds (Nicholson et al. 1987), the resulted high level accumulation in resistant sugarcane cultivars could therefore account for darker or intense pigmentation around the inoculation site.
In sorghum, 5-methoxy luteolinidin was found to accumulate along with luteolinidin in the resistant cultivar in response to C. sublineolum inoculation. However, the concentration of 5-methoxy luteolinidin is very low than luteolinidin. The level accumulation may, in part, due to the more potent activity of 5-methoxy luteolinidin against C. sublineolum, as it was found to completely inhibit C. sublineolum germination even at concentrations of 3 µM (Lo et al. 1996). However in sugarcane the methylated derivatives were found more in compatible interaction than incompatible interactions. Although the methylated derivatives of flavonoids were generally more potent antibiotics than their un-methylated counterparts (Van Etten et al. 1980; Kodama et al. 1992; Aida et al. 1996) but a few phytopathogenic fungi were able to degrade these methylated phytoalexins and mount successful infection. Since 5-methoxy luteolinidin is completely lacking in all the resistant cultivars studied, it is possible that demethylation mechanism may operate in sugarcane - C. falcatum interaction, which needs further investigation. If that is the case, accumulation of 5-methoxy luteolinidin will be of no use and may also trigger the pathogen virulence genes. Hence the absence of 5-methoxy luteolinidin in resistant varieties will help them to avoid the activation of pathogen virulence factors and alternative accumulation of new phytoalexin compounds will complement its absence. This postulation was further supported by the fact that, two of the susceptible cvs Co 1148 and CoC 92061 were able to accumulate 5-methoxy luteolinidin at concentrations of 0.0127 and 0.0079 µg mg−1 tissue (corresponds to 46 and 29 µM), respectively and still showing compatible interaction. Another possible reason for this failure in pathogen inhibition may be due to the lack of targeted accumulation of this compound around the infection site.
Apigeninidin induction was observed only in three resistant cvs Baragua, BO 91 and Co 94008. Among them the cv BO 91showed higher induction when compared to Co 94008. Arabinosyl-5-O-apigeninidin was found to be induced in resistant cvs Baragua and Co 94008 and also in susceptible cv Co 997. Its induction was highest in Baragua and lowest in Co 997. It was specifically induced upon pathogen inoculation in the cv Baragua. In sorghum, apigeninidin accumulation was found to be the first response upon inoculation of non pathogenic fungus Bipolaris maydis in resistant cultivars, however during course of time (by 24 h) caffeic acid ester of arabinosyl 5-O-apigeninidin and luteolinidin were found to accumulate at higher levels (Wharton and Nicholson, 2000). Luteolinidin and its derivative were found to be more fungitoxic than apigenenidin compounds (Lo et al. 1996). Further to check the invading pathogen effectively, highier quantities of apigeninidin need to induced, as in the case of the resistant cv BO 91. The absence of induction in other varieties tested may be complemented by the induction of alternative, more fungitoxic compounds. Similarly the induction of Arabinosyl-5-O-Apigeninidin was found to be limited in three of the 11 cultivars studied viz., Baragua, Co 997 and Co 94008. Its induction was highest in Baragua upon pathogen inoculation. Since, other resistant cultivars accumulated luteolinidin and other unknown compounds at higher levels upon pathogen inoculation, it may be concluded that relatively minor distribution of apigenenidin compounds may be due to the synthesis of other related compounds, probably using apigenenidin as the precursor. Accumulation of phytoalexin varies in sugarcane tissues depending on the site of pathogen intrusion, its progression and adjoining tissue. Induction of different phytoalexin compounds is dynamic one and we found accumulation of one or two phytoalexins separately or together during the host-pathogen interaction. In this study we found higher quantities of luteolinidin induction with no accumulation of apigeninidin in resistant cv Co 93009. This suggests that in incompatible interaction, the pathogen progress is restricted either by increased accumulation of a single phytoalexin or different compounds. However further studies are required on the level of phytoalexins required to inhibit the fungal growth inside the host tissue. It was proposed that the flavonoid 3′ hydroxylase (F3′H) activity will lead to the conversion apigeninidin to luteolinidin (Liu et al. 2010). In support for this, in our transcript profiling study, a unique F3′H transcript was found to accumulate in the resistant sugarcane cv Co 93009 (Viswanathan, Unpublished). The remaining five compounds (X1–X5) yet to be characterized due to the lack of their similarity with the standards used in the present study. Amongst the unknown compounds, X1 was found to be present only in the susceptible cultivars. The compounds X2, X4 and X5 were found to be present only in the resistant cultivars at high concentrations (Fig. 4). Hence the unknown compounds identified in this study may play important role in the incompatible reaction of sugarcane against C. falcatum. In sorghum also, induction of several unknown compounds with spectral characteristics similar to 3-deoxyanthocyanidins were reported to be induced upon inoculation of pathogens (Nicholson et al. 1987).

Induction of different uncharacterized phytoalexin compounds during the interaction with C. falcatum C—Mock inoculated control (48 h), I—pathogen inoculated (48 h), tR—retention time, X1–X5—uncharacterized peaks
Accumulation of phytoalexins varies with varieties during stress. It was not only induced during the fungal pathogen intrusion, it is also been induced upon insect attack as well as during mechanical injury which was shown by the induction of these compounds in the mock control due to mechanical injury. Previous studies revealed that partially purified pathogen toxin also induced higher quantities of phytoalexins in both the resistant and susceptible varieties. Also there was a considerable degradation of phytoalexin compounds in the susceptible variety (Viswanathan et al. 1996b). We found relatively higher induction of apigeninidin and luteolinidin in the cvs BO 91 and Baragua in the injured cane tissues than the inoculated tissues. This phenomenon may be due to partial loss of the induced chemicals or the smaller quantity induced may be sufficient to restrict the pathogen progress.
Analysis of total anthocyanidin content in 11 sugarcane cultivars varying on red rot resistance by spectrophotometric quantification revealed differential accumulation patterns in the stalk tissues upon pathogen challenge. The accumulation was found to be higher in the resistant cultivars at 24 h after pathogen inoculation than the susceptible cultivars. This indicates that susceptible cultivars were unable to synthesize phytoalexin compounds in relation to resistant cultivars (Fig. 5).

Quantification of 3-deoxyanthocyanidin phytoalexin compounds in different sugarcane varieties varying in red rot resistance by calorimetry (24 h after pathogen inoculation)
In conclusion, the present study revealed the differential accumulation of phytoalexin compounds among red rot resistant and susceptible sugarcane cultivars, indicating their probable role in resistance mechanism. Presence of four 3-deoxyanthocyanidin compounds viz., apigeninidin, arabinosyl 5-O apigeninidin, luteolinidin and 5-methoxy luteolinidin in sugarcane was confirmed and five unknown compounds with their possible roles in red rot resistance mechanism were identified. The differential expression pattern of different 3-deoxyanthocyanidin fractions even among the resistant cultivars, indicate the multiple modes of phytoalexin mediated resistance mechanism operating in sugarcane. The consistent induction of luteolinidin at high levels in the resistant cultivars and the relatively low or absence of induction in the susceptible cultivars clearly indicates the major contribution of luteolinidin sugarcane resistance reaction to C. falcatum (Fig. 3). Our studies very clearly established that luteolinidin may be the major phytoalexin which determines host resistance to C. falcatum which needs further investigation. Few of the resistance cultivars showed involvement of apigeninidin. Also specific induction of unidentified compounds found in both compatible and incompatible interactions. Further studies are in progress to identify the compounds and to relate their role in red rot resistance and degradation of phytoalexin compounds in compatible interactions. Our studies also indicate contribution of two or more phytoalexin compounds in red rot resistance.
References
Aida, Y., S. Tamogami, O. Kodama, and T. Tsukiboshi. 1996. Synthesis of apigeninidin 7-methylether and its fungicidal activity against. Gloeocercospora sorghi. Bioscience Biotechnology and Biochemistry 60: 1495–1496.
Boddu, J., C. Svabek, R. Sekhon, A. Gevens, R.L. Nicholson, D.A. Jones, J.F. Pedersen, D.L. Gustine, and S. Chopra. 2004. Expression of a putative flavonoid 3′-hydroxylase in sorghum mesocotyls synthesizing 3-deoxyanthocyanidin phytoalexins. Physiological and Molecular Plant Pathology 65: 101–113.
Hammerschmidt, R. 1999. Phytoalexins: What have we learned after 60 years? Annual Review Phytopathology 37: 285–306.
Knogge, W., E. Kombrink, E. Schmelzer, and K. Hahlbrock. 1987. Occurrence of phytoalexins and other putative defense-related substances in uninfected parsley plants. Planta 171: 279–287.
Kodama, O., J. Miyakawa, T. Akatsuka, and S. Kiyosawa. 1992. Sakuranetin, a flavanone phytoalexin from ultraviolet-irradiated rice leaves. Phytochemistry 31: 3807–3813.
Liu, H., Y. Du, H. Chu, C.H. Shih, Y.W. Wong, M. Wang, I.K. Chu, Y. Tao, and C. Lo. 2010. Molecular dissection of the pathogen-inducible 3-deoxyanthocyanidin biosynthesis pathway in sorghum. Plant Cell Physiology 51: 1173–1185.
Lo, S.-C., I. Weiergang, C. Bonham, J. Hipskind, K. Wood, and R.L. Nicholson. 1996. Phytoalexin accumulation in sorghum: Identification of a methyl ether of luteolinidin. Physiological and Molecular Plant Pathology 49: 21–31.
Lo, S.-C.C., K. de Verdier, and R.L. Nicholson. 1999. Accumulation of 3-deoxyanthocyanidin phytoalexins and resistance to Colletotrichum sublineolum in sorghum. Physiological and Molecular Plant Pathology 55: 263–273.
Malathi, P., R. Viswanathan, P. Padmanaban, D. Mohanraj, V. Ganesh Kumar, and K.P. Salin. 2008. Differential accumulation of 3-deoxy anthocyanidin phytoalexins in sugarcane varieties varying in red rot resistance in response to Colletotrichum falcatum infection. Sugar Tech 10: 154–157.
Mansfield, J.W. 1999. Antimicrobial compounds and resistance: the role of phytoalexins and anti anticipins. In Mechanisms of resistance to plant diseases, ed. A.J. Slusarenko, R.S.S. Fraser, and L.C. VanLoon, 325–370. Amsterdam: Kluwer.
Nicholson, R.L., S.S. Kollipara, J.R. Vincent, P.C. Lyons, and G. Cadena-Gomez. 1987. Phytoalexin synthesis by the sorghum mesocotyl in response to infection by pathogenic and nonpathogenic fungi. Proceedings of the National Academy of Sciences USA 84: 5520–5524.
Snyder, B.A., and R.L. Nicholson. 1990. Synthesis of phytoalexins in sorghum as a site specific response to fungal ingress. Science 248: 1637–1639.
Snyder, B.A., B. Leite, J. Hipskind, L.G. Butler, and R.L. Nicholson. 1991. Accumulation of sorghum phytoalexins induced by Colletotrichum graminicola at the infection site. Physiological and Molecular Plant Pathology 39: 463–470.
Tenkouano, A., F.R. Miller, G.E. Hart, R.A. Frederiksen, and R.L. Nicholson. 1993. Phytoalexin assay in juvenile sorghum: An aid to breeding for anthracnose resistance. Crop Science 33: 243–248.
Tsuji, J., E.P. Jackson, D.A. Gage, R. Hammerschmidt, and S.C. Somerville. 1992. Phytoalexin accumulation in Arabidopsis thaliana during the hypersensitive reaction to Pseudomonas syringae pv. syringae. Plant Physiology 98: 1304–1309.
Van Etten, H.D., P.S. Matthews, K.J. Tegtmeier, M.F. Dietert, and J.I. Stein. 1980. The association of pisatin tolerance and demethylation with virulence on pea in Nectria haematococca. Physiological Plant Pathology 16: 257–268.
Van Etten, H.D., J.W. Mansfield, J. Bailey, and E.E. Farmer. 1994. Letter to the editor: Two classes of plant antibiotics: Phytoalexins versus phytoanticipins. Plant Cell 6: 1191–1192.
Viswanathan, R. 2010. Plant disease: red rot of sugarcane. New Delhi: Anmol Publications Pvt. Ltd.
Viswanathan, R., D. Mohanraj, and P. Padmanaban. 1996a. Role of red rot pigments in relation to red rot resistance in sugarcane. Indian Journal of Sugarcane Technology 11: 151–154.
Viswanathan, R., D. Mohanraj, P. Padmanaban, and K.C. Alexander. 1996b. Synthesis of phytoalexins in sugarcane in response to infection by Colletotrichum falcatum Went. Acta Phytopathologica Entomologica Hungarica 31: 229–237.
Wharton, P.S., and R.L. Nicholson. 2000. Temporal synthesis and radiolabelling of the sorghum 3-deoxyanthocyanidin phytoalexins and the anthocyanin, cyanidin3-dimalonyl glucoside. New Phytologist 145: 457–469.
Acknowledgments
Authors are thankful to Dr. N. Vijayan Nair, Director of the Institute for the support and encouragement. The research work was supported by Sugar Development Fund, Ministry of Consumer Affairs, Food and Public Distribution, New Delhi.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Ganesh Kumar, V., Viswanathan, R., Malathi, P. et al. Differential Induction of 3-deoxyanthocyanidin Phytoalexins in Relation to Colletotrichum falcatum Resistance in Sugarcane. Sugar Tech 17, 314–321 (2015). https://doi.org/10.1007/s12355-014-0334-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12355-014-0334-1