
Abstract
We aimed to study the genetic diversity and population structure of eight Iranian terrestrial orchid species, including Anacamptis coriophora (L.) R. M. Bateman, Pridgeon and M. W. Chase, Dactylorhiza umbrosa (Kar. & Kir.) Nevski, Himantoglossum affine (Boiss.) Schltr., Orchis collina Banks and Solander, Orchis mascula (L.) L., Orchis simia Lam., Ophrys schulzei Bornm. and Fleischm., and Ophrys straussii H. Fleischm. and Bornm. using start target codon markers (SCoT) and finding markers associated with seed morphometric traits. A total of 254 reproducible SCoT fragments were generated, of which 248 fragments were polymorphic (average polymorphism of 96.18%). The SCoT markers showed a narrow range of polymorphism information content (PIC) varied from 0.397 for S9 primer to 0.499 for S11 and S20 primers. Based on the population analysis results, the Orchis simia accessions collected from Paveh region (Os.P) represented the lowest observed number of alleles (Na) (1.13) and effective number of alleles (Ne) (1.09). At the same time, the highest Na (1.29) and Ne (1.18) values were obtained in O. schulzei collected from Javanrood (Oyst.JA). Shannon’s information index (I) was ranged from 0.03 for D. umbrosa accessions collected from Marivan (Du.M population) to 0.263 for Ha.Ja population (H. affine accessions collected from Javanrood). The UPGMA dendrogram obtained with the Jaccard similarity coefficient (r = 0.97295) divided 97 studied terrestrial orchid accessions into eight groups mainly based on species type and geographical origin. Based on the Bayesian statistical index, the highest probability of the data was achieved when accessions were divided into eight groups (K = 8). Multiple association analysis (MRA) revealed significant associations between some of SCoT bands with seed morphometric traits. Our findings can be useful for germplasm characterization, conservation, and improvement of Iranian terrestrial orchid species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
With more than 28,000 species comprising over 763 genera, Orchidaceae is the second largest family within the plant kingdom. Orchids have spread worldwide from tropical to high alpine areas (Phillips et al. 2020). Remarkable flower beauty and diversity, unusual seeds, exceptional and complicated pollination, and reproduction system, as well as particular symbiotic interactions with mycorrhiza fungi, have led to focusing comprehensive scientific researches on various biological aspects of orchid species (Cozzolino and Widmer 2005; Sherif et al. 2020). Among different life forms of orchids, terrestrial orchids are usually grown in soil and generate a round or palmate underground fleshy tuber (de Boer et al. 2017). These fleshy tubers are currently used to make traditional ice-cream and beverages with unique rheological properties in the minor Asia and the Middle East (Farhoosh and Riazi 2007).
Compared with neighboring regions, orchids are rather rare and usually not numerous in Iran. This is due to its situation, where the western, north-western and eastern marginal zones are the only hotspots of much richer orchid centers. The Iranian orchids have spread over various habitats not only over the Hyrcanian and dry oak forests but also throughout the alpine wet meadows, primarily the Alborz and the Zagros Mountains. They mainly belong to European-Mediterranean species within Dactylorhiza, Orchis, Cephalanthera, Epipactis and Ophrys genera (Renz 1978). No connection is present between the two hotspots of orchids distribution in the western and eastern periphery of the Iran plateau. This is because the deserts are located at the center of the country which has separated the European-Mediterranean species from the Himalayan species that can be seen in the far east of the country (Kreutz and Spencer 2011). This wide gap has limited the possible gene flow of orchids in either direction. Based on the Flora Iranica, 46 tuberous orchid species are grown in the Iran, mainly in western regions, including Kurdistan, Kermanshah, and West Azerbaijan provinces (Kreutz and Spencer 2011; Renz 1978). Among Iranian terrestrial orchids, 36 species have been exploited for their fleshy tubers, and only in 2013 around 7–11 million orchid tubers have been dug and collected causing depletion of natural populations (Ghorbani et al. 2014a, b). All Iranian orchids are listed in the Convention on International Trade of Endangered Species of Fauna and Flora (CITES) appendices (Appendix I or II) (CITES 2014). Besides, over-harvesting, the Iranian terrestrial orchids have also experienced an extensive range of fragmentation and decreased the population sizes due to climate change (Nosrati et al. 2011). Despite the extensive commercial usage, Iranian tuberous orchids have not already been domesticated and are exclusively collected from nature, leading to even the local distinction of some species (Moradi et al. 2015; Ghorbani et al. 2014b).
Comprehensive and detailed information on genetic variability and population structure is fundamental to highlight the long-term population viability and adaptation under dynamic environmental conditions (Ebadi et al. 2019; Vafaee et al. 2008). The knowledge on genetic variability of terrestrial orchids can provide baseline genetic information for the development of sustainable conservation programs and for breeding and improvement strategies (Vafaee et al. 2017).
Little information is available on the genetic diversity of Iranian terrestrial orchid species, and there are only few studies available on these threatened species. Vafaee et al. (2021) evaluated genetic diversity of a collection of 107 individuals of Iranian terrestrial orchids using floral- and tuber-related traits and seed micromorphology characteristics. Ghorbani et al. (2017) determined the phylogeny of the Iranian orchids based on DNA-metabarcoding analyses at the genus and species levels. Kaki et al. (2020) investigated intra- and inter-population genetic variation and genetic distance among five medicinal orchid species using the IRAP marker. ISSR marker was also successfully applied to elucidate the genetic variation among individuals and inter- and intra-population diversity in a collection of Iranian terrestrial orchids (Gholami et al. 2021). Hence, DNA-based molecular markers like SCoT have not yet been used to study genetic variation in medicinal Iranian orchid species. On other side, seed and testa micro-morphological traits have been used as taxonomical and phylogenetic markers on a vast range of orchid species (Barthlott et al. 2014). In this regard, scanning electron microscopy (SEM) has come to aid the analyses of orchid seeds and their microstructures (Aybeke 2007).
Among DNA-based molecular markers, start codon targeted (SCoT) polymorphism has been introduced as a gene-targeted molecular marker system principally different from random DNA markers and it generally relies on the short conserved nucleotide region surrounding the ATG initiation codon of plant genes (Collard and Mackill 2009). SCoT markers are advantageous over other PCR-based molecular markers because of low cost, easy operation, high loci amplification and polymorphism, high reproducibility and reliability and suitability for marker-assisted selection programs (Bhattacharyya et al. 2013; Collard and Mackill 2009). The SCoT markers have been effectively employed to study the genetic variation and population structure of different species within the Orchidaceae family, including Dendrobium nobile (Bhattacharyya et al. 2013), Cypripedium japonicum (Tian et al. 2018), Eulophia sp. (Bhattacharyya and van Staden 2018), Huperzia serrata (Minh et al. 2019), and Paris polyphylla (Zhao et al. 2020). We assume that there is different genetic variation pattern among individuals of geographically distant populations and SCoT marker could be a good choice to elucidate this diversity pattern. On the other hand, as the seed morphometry of orchids are species-specific, we hypothesize that some seed micro-morphology traits can possibly be associated with some SCoT loci. This is the first study on the usability of SCoT markers on genetic diversity of endangered Iranian orchid species as well as the first report on the possible association between seed microstructure traits with molecular markers in species within Orchidaceae family which could also be exploited for breeding and conservation purposes. Therefore, the present research study was aimed to (i) evaluate important seed micro-structure characteristics and their variation, (ii) study genetic diversity and population structure using 15 SCoT primers, and (iii) estimate associations between SCoT loci and evaluated seed micro-structure characteristics in a collection of 97 accessions belonging to eight terrestrial orchid species.
Material and methods
Plant material
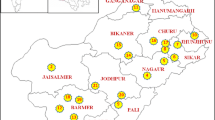
A collection of 97 accessions belonging to sixteen geographically isolated populations of eight Iranian terrestrial orchids species including Anacamptis coriophora (L.) R.M.Bateman, Pridgeon and M.W.Chase, Dactylorhiza umbrosa (Kar. & Kir.) Nevski, Himantoglossum affine (Boiss.) Schltr., Orchis collina Banks and Solander, Orchis mascula (L.) L., Orchis simia Lam., Ophrys schulzei Bornm. and Fleischm., and Ophrys straussii H. Fleischm. and Bornm. were evaluated (Fig. 1). Table 1 represents the population origin, code, accession number, and geographical coordinates of studied orchid populations. The distribution map of the collected populations has been shown in Fig. 1. The systematic identification of plant material was performed in the Herbarium of University of Kurdistan, where the voucher specimens of collected species have been deposited. A minimum of five accessions per population was considered for genetic diversity analysis. For SCoT analysis, young leaf samples were collected, labeled, and frozen in liquid nitrogen and finally kept at − 80 °C until DNA extraction.

Geographic locations and collection sites of the sample populations and inflorescence of studied terrestrial orchid species. a Dactylorhiza umbrosa (Kar. & Kir.) Nevski, b Himantoglossum affine (Boiss.) Schltr., c Orchis simia Lam., d Orchis mascula (L.) L., e Anacamptis coriophora (L.) R.M.Bateman, Pridgeon & M.W.Chase, f Ophrys schulzei Bornm. & Fleischm., g Ophrys straussii H. Fleischm. & Bornm., h Orchis collina Banks and Solander
DNA extraction and quantification
The total DNA was extracted from ~ 100 mg leaf tissue according to the cetyltrimethylammonium (CTAB) based on the Doyle and Doyle (1987) procedure with minor modifications. The quantity and quality of the extracted DNAs were determined through both spectrophotometry (using a ND 1000 spectrophotometer, NanoDrop Technologies, Wilmington, DC, USA) and agarose gel electrophoresis methods. The DNA samples with optimal quality and purity were used for PCR amplification.
PCR reactions and SCoT analysis
To optimize PCR amplification, two DNA samples for each population were chosen for preliminary evaluation of primers by determining samples with satisfactory band profile and exclusion of primers with low quality and weak bands. From 19 assessed SCoT primers, 15 primers produced clear and consistent bands, and these primers were used for the final SCoT amplification. PCR reactions carried out in a final volume of 10 µl, comprising 20 ng pure DNA, 200 µM dNTPs, 0.4 U Taq DNA polymerase (SinaClone, Tehran, Iran), 1 µl 10X PCR buffer, 2 mM MgCl2 and 10 pmol/μl SCoT primer. PCR reactions performed in a Biorad thermocycler (Bio-Rad, Hercules, CA, USA) based on the following program: 5 min initial denaturation at 94 °C followed by 30 cycles of 45 s denaturation at 94 °C, 60 s primer annealing at 50.7–58 °C and 90 s extension at 72 °C, with a final extension of 7 min at 72 °C. In addition to the main PCR amplifications, negative (water) and positive control samples were also tested to identify possible self-dimers or DNA contaminations and to check the consistency of amplified bands, respectively. The obtained DNA bands were separated on 1.5% agarose gel (SinaGene, Iran) at 90 V in 1 × TAE buffer for 120–150 min. The amplified DNA fragments were visualized by staining the agarose gels in 0.0015% w/v ethidium bromide solution and photographing under UV light by a Bio-Rad Gel Doc 2000 model (Bio-Rad, Hercules, CA, USA). Weak and ambiguous fragments were eliminated from scoring, and only clear and sharp fragments were considered for the next steps where the bands scored as 1 and 0 for the presence and absence, respectively. The PCR product size was determined using a 1 kb SinaGene DNA ladder (250–3500 bp).
Seed morphometric analysis
To analyze seed morphometric traits, fully ripen capsules were obtained for each individual and transported to the research lab at the Department of Horticultural Sciences and Engineering, University of Kurdistan. The obtained seed were further dehydrated for a minimum of 1 month and were maintained in closed paper bags. The light microscopic images taken by Olympus BX51 stereomicroscope (fitted with an Olympus DP72 camera) were used to determine seed testa color. To perform scanning electron microscope (SEM) experiment, the seed of each terrestrial orchid species were fixed on an aluminum SEM stub and were then coated with 10-nm gold particles. The SEM experiment was carried out using a FESEM Mira 3 scanning electron microscope (Tescan, Breno, Czech Republic) at 30 kV voltage. Rating and scaling were used to analyze seed qualitative micromorphology characteristics including testa color (TeCo), testa cell pattern (TeCePa), overall seed shape (OvSeSh), ridges on periclinal cell walls (RiPeWa), height of anticlinal walls (HeAnWa), and anticlinal wall curvature (AnWaCu). The obtained SEM images were used to evaluate quantitative attributes including seed width (SeWi), seed length (SeLe), seed length/width ratio (SeLe/Wi), seed cell length (SeCeLe), seed/cell length ratio (Se/CeLe), cell number per seed (CeNu/Se), and cell number in the longitudinal axis (CeNuLoAx). In this connection, the terminologies and procedures introduced by Barthlott et al. (2014), Akçin et al. (2009); Arditti and Ghani (2000), Szendrak (1997), Molvray and Kores (1995), and Chase and Pippen (1988) were used. The SEM experiment was performed in triplicate (3 capsules) where five seed were randomly selected for analyses.
Data analyses
The obtained mean values for seed micro-structure quantitative traits were used for statistical analysis. The descriptive statistical measures such as mean, minimum and maximum values, standard deviation (SD) and coefficient of variation (CV%) were estimated for the evaluated seed traits using PAST software (Hammer et al. 2001). The CV percentage as a variation index was also calculated using PAST software.
The genetic similarity among terrestrial orchid accessions based on Jaccard’s coefficient was estimaed using the SIMQUAL implemented in the numerical taxonomy multivariate analysis system NTSYS-pc version 2.10 (Rohlf 2000) and the dendrogram were drawn using SAHN clustering program with the unweighted pair group method with arithmetic means (UPGMA) as recommended in most diversity studies. To assess the efficiency of SCoT primers, the total number of polymorphic bands, polymorphism information content (PIC), effective multiplex ratio (EMR), marker index (MI), and resolving power (RP) parameters were estimated according to Powell et al. (1996). GenAlEx software version 6.5 (Peakall and Smouse 2006) was used to calculate genetic diversity indices including the number of different alleles (Na), the number of effective alleles (Ne), Shannon index (I), Nei's gene diversity (H) and the percentage of polymorphic loci (Pp%) at both population and species levels. Moreover, the principal coordinate analysis (PCoA) was carried out to show the relationships among the terrestrial orchid accessions based on the variance–covariance matrix obtained from SCoT data, and a biplot was then drawn using the first two main coordinates (PCo1 and PCo2).
To analyze population structure, a clustering method based on the Bayesian model implemented in the STRUCTURE software package 2.3.4 was used (Falush et al. 2007; Pritchard et al. 2000). This model identifies subpopulations, assigns their individuals, and estimates the allele frequency. The structure analysis was conducted by 10 independent runs per each K with a cluster number ranging from K = 1 to K = 10. For each individual run, a 50,000 burn-in length was set with 5000 Markov Chain Monte Carlo (MCMC) replication using the admixture model and independent allele frequency. To calculate the best K value, both log-probability of the data (LnP(D)) and delta K values determined with the online program Structure Harvester (Earl and vonHoldt 2011).
Multiple regression analysis (MRA) was used to identify SCoT loci (as independent variables) associated with the studied seed micro-structure characteristics (as dependent variables). For this purpose, stepping criteria using SPSS software were employed. The model \({\text{Y }} = a + b_{1} m_{1} + b_{2} m_{2} + \ldots + b_{j} m_{j} + \ldots + b_{n} m_{n} + d + e\) was used to perform MRA. This model attributes the variation in the dependent variable Y to a linear function of the set of SCoT markers (defined as independent variables, mj). The coefficient of partial regression (bj) indicates the empirical connection between Y and mj, d shows residuals among individuals (remaining after regression) representing the random error of Y, and e represents the environmental variation (Virk et al. 1996). The SCoT loci with significant regression values were assumed to be associated with the related seed micro-structure trait. The independent variables were selected based on F-values with the probability of 0.045 and 0.099 for entry and removal, respectively. Besides, the evaluated SCoT loci were individually tested with a linear curve fitting by linear models to verify the significance of standardized beta coefficients (β-statistics) for each locus calculated by MRA.
Results
Seed morphometric analysis
Seed micro-structure characteristics were studied by SEM in all individuals within collected populations, and there was a high variation among these traits (Table 2). The CV values for qualitative characteristics, including anticlinal wall curvature (138.23%), ridges on periclinal cell walls (82.06%), testa color (54.19%), and overall seed shape (53.47%) were higher compared to quantitative traits. For the height of anticlinal wall, H. affine accessions and all accessions within the genus Orchis excluding O. mascula was prominent. On the other hand, the height of anticlinal wall was low in D. umbrosa accessions (Fig. 2a), while it was medium–high in O. mascula and Ophrys species. Overall, in most of evaluated terrestrial orchid accessions, the cells in apical and basal ends were short and smaller, while the cells in the middle part of seed were rectangular and elongated. The seed testa color of studied terrestrial orchid accessions was mainly brown and dark brown. Regarding seed dimensions, the highest and the lowest seed width was observed in Os.Pa1 (127.6 µm) and Du.M3 (276.6 µm), respectively (Electronic Supplementary 1). On the contrary, Ocol.K6 and Os.J1 had the shortest (304.4 µm) and longest (788.4 µm) seed, respectively. Fusiform was the most prevalent seed shape among the studied terrestrial orchid accessions (Fig. 2). This type of seed shape was observed in accessions within O. simia (Fig. 2d), O. straussii (Fig. 2f), and O. schulzei (Fig. 2g) species. The studied terrestrial orchid accessions represented a significant difference in seed cell number. In this connection, Du.S1 (D. umbrosa, Salian population) and Ac.S4 (A. coriophora, Salian population) represented the highest (139.3) and the lowest (36.6) seed cell number, respectively.

Seed morphology, view in light microscopy and SEM. a Dactylorhiza umbrosa, b Himantoglossum affine, c Orchis mascula, d Orchis simia, e Anacamptis coriophora, f Ophrys schulzei, g Ophrys straussii, h Orchis collina
Molecular analysis
SCoT polymorphism
Among 19 preliminary studied SCoT primers, 15 primers produced clear and sharp banding patterns, which were considered for final reactions and analyses. As represented in Table 3, a total of 254 fragments were generated, of which 248 fragments were polymorphic (average polymorphism of 96.18%). The total band per primer was varied from 9 (S9 primer) to 22 (S20, S21, and S23 primers) with a size range 100–2800 bp. Most of the studied SCoT primers represented 100% of polymorphism, and only S5, S9 and S11 had lower polymorphism percentages. The allele frequency of each primer was used to calculate polymorphism information content (PIC). The highest PIC value (0.499) was obtained with S11 and S20 primers, confirming their higher efficiency to determine genetic diversity and distance (Table 3). On the contrary, the lowest PIC value (0.397) was calculated for S9 primer. The marker index (MI) was ranged from 2.19 by S5 primer to 7.50 by S20 primer with a mean of 5.26 for all evaluated SCoT primers. The average RP value of the SCoT primers used in the present study was observed as 15.66. The highest (29.89) and the lowest (9.19) RP values were obtained for S5 and S3 primers, respectively.
Genetic diversity and differentiation
The genetic diversity indices of the studied populations are represented in Table 4. The O. simia accessions collected from the Paveh region (Os.P) represented the lowest observed number of alleles (Na) (1.13) and effective number of alleles (Ne) (1.09), while the highest Na (1.29) and Ne (1.18) values were obtained in O. schulzei collected from Javanrood region (Oyst.JA) (Table 4). With an average of 0.114, heterozygosity (H) was found to be highest (0.195) in population D of A. collina (O.colK) collected from Divandareh region, while the lowest H (0.51) was obtained in Os.P population of O. simia collected from Paveh region. Furthermore, Shannon’s information index (I) was ranged from 0.03 for D. umbrosa accessions collected from Marivan region (Du.M population) to 0.263 for Ha.Ja population (H. affine accessions collected from Javanrood region). Based on AMOVA results, a significant (P < 0.001) genetic differences were obtained within and among populations (Table 5). Of the total genetic variation, inter-population variations were 68%, and the rest 32% was intra-population variation.
Cluster analysis
The most appropriate cut-off line to classify orchid individuals and assigning clusters and sub-clusters was set based on the inter- and intra-group variance. Based on obtained dendrogram, the 97 studied terrestrial orchid accessions were clustered into eight main groups (Fig. 3). The first group included 17 accessions of D. umbrosa belonged to population Du.M (collected from Marivan region), Du.S (collected from Salian region), and Du.D (collected from Dehgolan region). All samples of A. coriophora belonging to Oc.J (collected from Javanrood) and Oc.S (collected from Salian) fell within the IInd group. In the IIIrd group, 15 accessions within Ocol.K and Ocol.D populations belonging to O. collina species were grouped together. The individuals within Om.P from Pave region belonging to O. mascula were classified under the IVth group. The Orchis simia accessions of Os.Pa and Os.J populations (from Paveh and Javanrood regions, respectively) were clustered under the Vth group. All accessions within VIth group were H. affine accessions belonging to A (Ha.JA1–Ha.JA5) and B (Ha.JB1–Ha.JB7) populations collected from Javanrood region. Interestingly, the accessions within Oysc.JA, Oysc.JB, Oyst.JA, and Oyst.JB belonging to O. sculzei and O. straussii species were closed under VIIth group.

UPGMA dendrogram of genetic similarity based on the SCoT data generated from Jaccard coefficients
The PCoA biplot was constructed based on the the first two principal coordinates (PCo1 and PCo2), which somewhat confirmed the results of UPGMA clustering. As illustrated in Fig. 4, most of the studied terrestrial orchid accessions were separated mainly based on species taxonomy except for O. schulzei, O. straussi, O. collina, and A. coriophora while their accessions were placed under common groups. Phylogenetically, O. collina and A. coriophora are the species with a close genetic relationship, which can also generate interspecific hybrids (Dafni and Ivri 1979). Both the UPGMA clustering and PCoA plot represented the close relationship between A. coriophora and O. collina accessions. On the other hand, few individuals from different populations of each species were clustered together. This genetic similarity between individuals of different regions can probably be due to anemochoric dispersal of their seeds over long distances and during long times (Pinheiro et al. 2012).

The PCoA biplot of terrestrial orchid populations based on the two first principal coordinates (PCo1 = 39.40%, PCo2 = 30.76%) using SCoT data
Population structure
In our study, the highest probability of the data was obtained when individuals were divided into eight populations (K = 8). Therefore, K = 8 was optimum to elucidate population structure and to estimate the membership matrix of individual clusters. Figure 5 shows the bar plot obtained for K values 7, 8, and 9. The obtained Bayesian matrix clustering was matched with the UPGMA dendrogram grouping and particularly with the PCoA plot, which also divided accessions into eight major clusters. In the bar plot obtained for K = 8, the studied accessions of D. umbrosa, H. affine, O. mascula, O. simia, A. coriophora, O. schulzei, O. straussii and O. collina were placed in corresponding clusters, respectively.

Population structure of 97 Iranian terrestrial orchids based on SCoT data analyzed by STRUCTURE software and Bayesian model. a K = 7; b K = 8; c K = 9. Number of each horizontal lane is the individual code and each color shows a subpopulation
Association between seed micro-structure traits and SCoT markers
The associations between SCoT amplified bands and seed micro-structure traits were determined using multiple regression analysis (MRA) (Table 6). Seed morphometric characteristics and SCoT markers were considered as dependent and independent variables where they were associated and showed highly significant correlation values. From 254 amplified SCoT bands, 63 bands were associated with seed microstructure traits. In this regard, S1-15, S13-13, S13-11, and S13-7 loci represented the highest associations with seed width (SeWe). Seed Length (SeLe) was significantly correlated with S11-2, S22-16, S20-22, and S21-10 markers. Significant associations were also observed between seed length/width ratio (SeLe/Wi) with S7-4, S20-22, S11-5, and S21-2 SCoT loci. Four SCoT bands, including S17-11, S7-5, S23-22, and S22-16 represented association with seed cell length (SeCeLe). Seed/Cell Length Ratio (Se/CeLe) was associated with S5-5, S22-12, S1-8, and S21-11, where S5-5 provided the highest positive β-value (0.646) among all analyzed SCoT bands. Cell number per seed (CeNu/Se) was significantly associated with S1-15, S23-19, S23-7, and S14-9 markers. Finally, there were significant association values between cells number in longitudinal axis (CeNuLoAx) with S3-2, S3-10, S7-11, and S17-10.
Discussion
Seed microstructure traits of testa and embryos are important in genetic diversity and phylogeny studies of different taxa in the Orchidaceae family. Fusiform seed was the most prevalent seed shape in the present research which is known for their narrow apical and basal ends and broad median part due to embryo placentation. Fusiform has been suggested as the most common shape of seed in Orchidoideae subfamily (Chase and Pippen 1988). In contrast, the accessions of D. umbrosa and H. affine (2A and 2B) had flask- and oval-shaped seed, respectively. These results are in agreement with the findings of Akçin et al. (2009) and Arditti and Ghani (2000). The overall seed shape can be used as a complementary tool for molecular markers to study genetic diversity and classification of terrestrial orchid species (Phillips et al. 2020; Şeker and Şenel 2017; Gallo et al. 2016; Gamarra et al. 2015b). Seed cell number and cell surface pattern have also been proposed as important micro-morphology characteristics to study genetic diversity and phylogenetic classification of orchid species as they are consistent and typical for any given orchid species (Gamarra et al. 2012, 2015a; Arditti and Ghani 2000). In line with previous reports, a species-specific pattern of seed microstructure traits was also revealed for Iranian terrestrial orchid species. From a conservational point of view, these findings can be particularly useful when the identity of ripen seed employed for in vitro conservation programs is unknown if the collection has been performed after flowering as the flower is the main tool to identify species (Verma et al. 2014). Therefore, the identification of orchid species using microstructure traits helps scientists and growers to establish the best conditions specific for given species or genera from seed germination to tuber formation and flowering. More importantly, seed morphometry analysis of orchids could be important in managing seed samples maintained in gene banks to determine taxa identity for further usage in in vitro and in situ conservation programs, particularly in case of threatened and endangered species.
Different range of PIC, MI, and Rp value were obtained for evaluated SCoT primers. PIC can be efficiently employed in genetic diversity studies of plant populations and it shows the discriminating power of markers in revealing polymorphism within the populations and differentiate genotypes based on the known allele frequency and distribution (Kaki et al. 2020; Botstein et al. 1980). PIC equals genetic variation, and higher PIC values imply a higher polymorphism and higher number of rare alleles in a single gene locus. In this regard, PIC determines the marker efficiency by providing a benchmark of primers employed in genetic diversity studies (Botstein et al. 1980). Variables including allele number generated by loci and genotype and primer number can affect the PIC range in genetic diversity and population structure analyses, which could be from 0 to 1 (Powell et al. 1996). MI is another important genetic index employed to determine the overall utility of marker systems and estimate primer efficiency to discriminate plant germplasm resources based on the polymorphic band frequency. The higher MI values thus suggest more coverage and information of the corresponding genome (Spooner et al. 2005). Similar to PIC, the higher MI implies the more efficiency and informativeness of the primer. This indicator is more informative when it exploits for inter-specific diversity studies (Medhi et al. 2014). RP refers to the primer ability to determine diversity level among sampled accessions based on the allele distribution and therefore, can be used as a powerful indicator to choose the suitable primers as it is impacted by the proportion of genotypes with bands and the number of bands produced (Azizi et al. 2009).
The fitness of loci to evaluate the genetic variation of plant accessions and populations depend on the average number of each marker (Rao and Hodgkin 2002). In this connection, primers that generate higher number of bands could be better for genetic variation studies as they are more effective in the coverage of genomic regions (Powell et al. 1996). The observed number of alleles is a pivotal index of genetic differentiation attributed to populations, types, and geographical locations. Na is 0 when accessions have the same allele frequency, and it equals 1 when the accessions are completely different for allele frequency. The effective number of alleles (Ne) is a reciprocal of expected homozygosity, so it is entirely correlated with the genetic diversity measured as the expected heterozygosity (or a sum of squares of allele frequencies at a locus) (Kimura and Crow 1963).
Both I and H are important genetic indices to study the polymorphism of populations gathered from geographically different locations. The higher I value represents higher diversity within populations, therefore, Ha.Ja had the highest diversity compared to other evaluated terrestrial orchid populations. This was further confirmed by its percentage of polymorphic loci (45.67%), which was also highest among all studied 16 populations. The level of genetic diversity is affected by different factors, including the current dynamical conditions of local populations, their adaptation to the climatic and micro-climatic conditions, and historical events. Size reduction and population fragmentation would decrease the level of genetic diversity within the population of terrestrial orchids due to genetic drift arising from restricted intra-population gene flow. Here, the observed low level of diversity among Iranian orchid populations, is probably due to exploitation and depletion of dried orchid tubers by local people of rural districts in West of Iran (de Boer et al. 2017; Kaki et al. 2020). The indices for intra-population variations estimated in our study were comparable with those obtained by the IRAP marker (Kaki et al. 2020) and ISSR markers (Gholami et al. 2021) in Iranian terrestrial orchids. This was also reported by Giri et al. (2017) (Habenaria edgeworthii), Liu et al. (2014) in Paphiopedilum micranthum, and Cai et al. (2011) in Dendrobium loddigesii.
Information on genetic variation is a key factor in defining conservation programs of rare orchid species (Bhattacharyya et al. 2013). The AMOVA results obtained in the present research were in line with the results obtained in Dendrobium nobile (Bhattacharyya et al. 2013, 2017), Cypripedium kentuckiense (Pandey et al. 2015) and Cypripedium japonicum (Tian et al. 2018) which shows a higher distribution of genetic variation amongst populations as compared to the distribution within populations. These findings further show the fragmentation of Iranian terrestrial orchid populations and their decreased connectivity and relationship, which have been previously indicated by the IRAP marker (Kaki et al. 2020).
Clustering is among the most important multivariate analyses to study the genetic variation of plant species. In the present study, Jaccard similarity provided the highest correlation coefficient (r = 0.97295), and therefore, it was used to draw dendrogram based on the UPGMA clustering algorithm. Excluding Ophrys species, the studied species has been well-grouped in separate clusters and its populations within each species fall in individual sub-clusters. Only in case of Ophrys genera, SCoT marker data set couldn’t differentiate species and population levels. In this regard, O. schulzei populations were not separated from O. straussii counterparts even as sub-clusters. This is probably because Ophrys has been known as the most diverse genus within terrestrial orchids (with over 250 identified species) where their interspecific relationship is a complicated classification of the Orchidaceae family due to the diverse intra-species hybridization (Breitkopf et al. 2015). The classification of remaining groups was mainly based on the species; however, the studied accessions were also separated relatively based on their populations and geographical collection site. A similar pattern of clustering was observed in Iranian terrestrial orchid species using IRAP molecular markers by Kaki et al. (2020). PCoA has been suggested as a suitable and complementary tool to interpret UPGMA or NJ clustering methods (Peakall and Smouse 2006). Therefore, PCoA was further performed to provide more details on the relationship among the evaluated accessions.
The Bayesian clustering method assigns accessions to hypothetical populations. The studied accesions of Iranian terrestrial orchis species were grouped mainly based on species where each species has been classified as a separate group. Population structure results were in agreement with the AMOVA results which revealed a higher level of the inter-population variation compared to intra-population diversity. Due to high sensitive to low degrees of genetic diversity, the SCoT technique is a powerful tool to study population genetics and structure in various plant species and also to identify species or population of the same species (Bhattacharyya et al. 2013).
In this paper, some SCoT markers were associated with more than one seed microstructure traits. This scheme of association could be due to correlation among traits and/or pleiotropy effect imposed by linked quantitative trait loci (QTL) on different characters (Culp et al. 1979). On the other hand, a single SCoT locus can include the tightly linked QTLs exhibiting different seed micromorphology traits. In this connection, S1-15 marker had association with both SeWi (β = − 0.799) and CeNu/Se (β = 1.268). Furthermore, S22-16 marker showed a strong association with SeLe and SeCeLe traits. As the SCoT primers target short conserved nucleotide regions surrounding the ATG initiation codon of plant genes, the loci associated with seed morphometric trait can contain the gene responsible for seed and testa development. This hypothesis can be tested by excision, purification, and sequencing of SCoT markers associated with seed morphometry traits, which can provide more genetic information on distinct seed coat features. This information could be useful in understanding seed germination process as seed testa is one of the main germination barriers in terrestrial orchids (Yeung 2017).
Conclusion
The preservation of genetic diversity is crucial in maintaining the evolutionary capacity of natural resources. The limited genetic variation generally results in reduced fitness of populations. Therefore, preliminary information on the genetic variability is pivotal to plan conservation and domestication programs for rare, endangered and threatened (RET) terrestrial orchid species. The SCoT primers employed in the current research considered to be appropriate to elucidate genetic variability and structure of 97 studied orchid accessions belonging to 16 populations and 8 Iranian tuberous orchid species. A high inter- population and low intra-population variability was observed in the present study, showing population fragmentation of terrestrial orchid species from the western part of Iran. This probably is the result of habitat deterioration mediated by anthropogenic activities and underground tuber over-harvesting, particularly in case of D. umbrosa and O. simia. All the studied Iranian orchids are classified as temperate hardy orchids with undeveloped embryos having internal seed dormancy. The features that result in low rate of germination in nature as these species need complicated symbiosis relationships with mycorrhiza fungi. Moreover, terrestrial orchid species need specific ecological niches and particular pollinators. Therefore, Iranian terrestrial orchid species deserve particular conservation attention on a national and international scale and our observations can be useful for germplasm characterization, conservation, and improvement of these valuable species.
Abbreviations
- CITES :
-
Convention on International Trade in Endangered Species of Wild Fauna and Flora
- MRA :
-
Multiple Association Analysis
- PCoA :
-
Principal Coordinate Analysis
- PIC :
-
Polymorphism Information Content
- SEM :
-
Scanning Electron Microscope
- UPGMA :
-
Un-weighted Paired Group Methods with Arithmetic Mean
References
Akçin TA, Ozdener Y, Akçin A (2009) Taxonomic value of seed characters in orchids from Turkey. Belg J Bot 142(2):124–139
Arditti J, Ghani AKA (2000) Numerical and physical properties of orchid seeds and their biological implications. New Phytol 145:367–421
Aybeke M (2007) Pollen and seed morphology of some Ophrys L. (Orchidaceae) taxa. J Plant Biol 50(4):387–395. https://doi.org/10.1007/bf03030673
Azizi A, Wagner C, Honermeier B, Friedt W (2009) Intraspecific diversity and relationship between subspecies of Origanum vulgare revealed by comparative AFLP and SAMPL marker analysis. Plant Syst Evol 281(1–4):151–160
Barthlott W, Große-Veldmann B, Korotkova N (2014) Orchid seed diversity: a scanning electron microscopy survey. Englera 32:1–245
Bhattacharyya P, van Staden J (2018) Molecular insights into genetic diversity and population dynamics of five medicinal Eulophia species: a threatened orchid taxa of Africa. Physiol Mol Biol Plants 24(4):631–641
Bhattacharyya P, Kumaria S, Kumar S, Tandon P (2013) Start Codon Targeted (SCoT) marker reveals genetic diversity of Dendrobium nobile Lindl., an endangered medicinal orchid species. Gene 529(1):21–26. https://doi.org/10.1016/j.gene.2013.07.096
Bhattacharyya P, Ghosh S, Mandi SS, Kumaria S, Tandon P (2017) Genetic variability and association of AFLP markers with some important biochemical traits in Dendrobium thyrsiflorum, a threatened medicinal orchid. S Afr J Bot 109:214–222. https://doi.org/10.1016/j.sajb.2016.12.012
Botstein D, White RL, Skolnick M, Davis RW (1980) Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am J Hum Genet 32(3):314
Breitkopf H, Onstein RE, Cafasso D, Schlüter PM, Cozzolino S (2015) Multiple shifts to different pollinators fuelled rapid diversification in sexually deceptiveOphrysorchids. New Phytol 207(2):377–389. https://doi.org/10.1111/nph.13219
Cai X, Feng Z, Zhang X, Xu W, Hou B, Ding X (2011) Genetic diversity and population structure of an endangered Orchid (Dendrobium loddigesii Rolfe) from China revealed by SRAP markers. Sci Hortic 129(4):877–881
Chase MW, Pippen JS (1988) Seed morphology in the Oncidiinae and related subtribes (Orchidaceae). Syst Bot 13:313–323
CITES (2014) The convention on international trade in endangered species of Wild Fauna and Flora: appendices. See https://cites.org/eng/app/appendices.php
Collard BC, Mackill DJ (2009) Start codon targeted (SCoT) polymorphism: a simple, novel DNA marker technique for generating gene-targeted markers in plants. Plant Mol Biol Report 27(1):86–93
Cozzolino S, Widmer A (2005) Orchid diversity: an evolutionary consequence of deception? Trends Ecol Evol 20(9):487–494
Culp TW, Harrell DC, Kerr T (1979) Some genetic implications in the transfer of high fiber strength genes to upland cotton. Crop Sci 19(4):481–484
Dafni A, Ivri Y (1979) Pollination Ecology of, and Hybridization between, Orchis coriophora L. and O. Collina Sol. Ex Russ. (Orchidaceae) in Israel. New Phytologist 83(1):181–187
de Boer HJ, Ghorbani A, Manzanilla V, Raclariu AC, Kreziou A, Ounjai S, Osathanunkul M, Gravendeel B (2017) DNA metabarcoding of orchid-derived products reveals widespread illegal orchid trade. Proc Biol Sci 284 (1863)
Doyle JJ, Doyle JL (1987) A rapid isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull 19:11–15
Earl DA, vonHoldt BM (2011) Structure Harvester: a website and program for visualizing Structure output and implementing the Evanno method. Conserv Genet Resour 4(2):359–361
Ebadi A, Ghaderi N, Vafaee Y (2019) Genetic diversity of Iranian and some European grapes as revealed by nuclear and chloroplast microsatellite and SNP molecular markers. J Hortic Sci Biotechnol. https://doi.org/10.1080/14620316.2019.1585210
Falush D, Stephens M, Pritchard JK (2007) Inference of population structure using multilocus genotype data: dominant markers and null alleles. Mol Ecol Notes 7:74–578
Farhoosh R, Riazi A (2007) A compositional study on two current types of salep in Iran and their rheological properties as a function of concentration and temperature. Food Hydrocoll 21(4):660–666. https://doi.org/10.1016/j.foodhyd.2006.07.021
Gallo FR, Souza LA, Milaneze-Gutierre MA, Almeida OJ (2016) Seed structure and in vitro seedling development of certain Laeliinae species (Orchidaceae). Revista Mexicana de Biodiversidad 87(1):68–73. https://doi.org/10.1016/j.rmb.2016.01.005
Gamarra R, Ortúñez E, Cela PG, Guadaño V (2012) Anacamptis versus Orchis (Orchidaceae): seed micromorphology and its taxonomic significance. Plant Syst Evol 298(3):597–607
Gamarra R, Galán Cela P, Seligrat I, Ortúñez E, Vivar A, Scrugli A (2015) A study of seed micromorphology in the genus Ophrys (Orchidaceae). An Jard Bot Madr. https://doi.org/10.3989/ajbm.2370
Gamarra R, GalÁN P, Pedersen HÆ, OrtÚÑEz E, Sanz E (2015) Seed micromorphology in Dactylorhiza Necker ex Nevski (Orchidaceae)and allied genera. Turk J Bot 39:298–309. https://doi.org/10.3906/bot-1401-66
Gholami S, Vafaee Y, Nazari F, Ghorbani A (2021) Molecular characterization of endangered Iranian terrestrial orchids using ISSR markers and association with floral and tuber-related phenotypic traits. Physiol Mol Biol Plants. https://doi.org/10.1007/s12298-020-00920-0
Ghorbani A, Gravendeel B, Zare S, De Boer HJ (2014) Illegal wild collection and international trade of CITES-listed terrestrial orchid tubers in Iran. Traffic Bull 26(2):52–58
Ghorbani A, Gravendeel B, Naghibi F, de Boer H (2014) Wild orchid tuber collection in Iran: a wake-up call for conservation. Biodivers Conserv 23(11):2749–2760
Ghorbani A, Gravendeel B, Selliah S, Zarre S, de Boer H (2017) DNA barcoding of tuberous Orchidoideae: a resource for identification of orchids used in Salep. Mol Ecol Resour 17(2):342–352
Giri L, Jugran AK, Bahukhandi A, Dhyani P, Bhatt ID, Rawal RS, Nandi SK, Dhar U (2017) Population genetic structure and marker trait associations using morphological, phytochemical and molecular parameters in Habenaria edgeworthii-a threatened medicinal orchid of West Himalaya. India Appl Biochem Biotechnol 181(1):267–282
Hammer Ø, Harper DAT, Ryan PD (2001) Paleontological statistics software package for education and data analysis. Palaeontol Electron 4(1):1–9
Kaki A, Vafaee Y, Khadivi A (2020) Genetic variation of Anacamptis coriophora, Dactylorhiza umbrosa, Himantoglossum affine, Orchis mascula, and Ophrys schulzei in the western parts of Iran. Ind Crops Prod. https://doi.org/10.1016/j.indcrop.2020.112854
Kimura M, Crow JF (1963) The measurement of effective population number. Evol Dev 17(3):279–288
Kreutz K, Spencer J (2011) In the Footsteps of Renz: Orchids in Iran. JHOS 8(1):12–24
Liu X, Huang Y, Li F, Xu C, Chen K (2014) Genetic diversity of 129 spring orchid (Cymbidium goeringii) cultivars and its relationship to horticultural types as assessed by EST-SSR markers. Sci Horticult 174:178–184. https://doi.org/10.1016/j.scienta.2014.05.015
Medhi K, Sarmah DK, Deka M, Bhau BS (2014) High gene flow and genetic diversity in three economically important Zanthoxylum Spp. of upper Brahmaputra valley zone of NE India using molecular markers. Meta Gene 2:706–721. https://doi.org/10.1016/j.mgene.2014.09.009
Minh NTA, Van TT, Hau HV, Trieu LN, Tien CV, Vinh TT, Van DN (2019) Genetic diversity and variation of Huperzia serrata (Thunb. ex Murray) Trevis. population in Vietnam revealed by ISSR and SCoT markers. Biotechnol Biotechnol Equip 33(1):1525–1534. https://doi.org/10.1080/13102818.2019.1671896
Molvray M, Kores PJ (1995) Character analysis of the seed coat in Spiranthoideae and Orchidoideae, with special reference to the Diurideae (Orchidaceae). Am J Bot 82:1443–1454
Moradi S, Dianati Daylami S, Vahdati K, Arab M (2015) Direct somatic embryogenesis of two Iranian native orchid species using protocorm explants. Iran J Horticult Sci Technol 16(1):137–148
Nosrati H, Hajiboland R, Razban-Haghighi A, Nikniazi M (2011) A comparative assessment of fruit formation in some orchid species from the southern Caucasus region. Turk J Bot 35(5):553–560
Pandey M, Richards M, Sharma J (2015) Microsatellite-based genetic diversity patterns in disjunct populations of a rare orchid. Genetica 143(6):693–704. https://doi.org/10.1007/s10709-015-9867-9
Peakall ROD, Smouse PE (2006) genalex 6: genetic analysis in excel. Population genetic software for teaching and research. Mol Ecol Notes 6(1):288–295
Phillips RD, Reiter N, Peakall R (2020) Orchid conservation: from theory to practice. Ann Bot. https://doi.org/10.1093/aob/mcaa093
Pinheiro LR, Rabbani ARC, da Silva AVC, da Silva LA, Pereira KLG, Diniz LEC (2012) Genetic diversity and population structure in the Brazilian Cattleya labiata (Orchidaceae) using RAPD and ISSR markers. Plant Syst Evol 298(10):1815–1825
Powell W, Morgante M, Andre C, Hanafey M, Vogel J, Tingey S, Rafalski A (1996) The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol Breed 2:225–238
Pritchard JK, Stephens M, Donnelly P (2000) Infernce of population structure using multilocus genotype data. Genet Biol Drosoph 155:945–959
Rao VR, Hodgkin T (2002) Genetic diversity and conservation and utilization of plant genetic resources. Plant Cell Tissue Organ Cult 68(1):1–19
Renz J (1978) Flora Iranica. Part: 126 orchidaceae. Naturhistorisches Museum, Vienna
Rohlf FJ (2000) NTSYS-pc numerical taxonomy and multivariate analysis system. Version 21 Exeter Software, Setauket
Şeker ŞS, Şenel G (2017) Comparative seed micromorphology and morphometry of some orchid species (Orchidaceae) belong to the related Anacamptis Orchis Neotinea Genera. Biologia. https://doi.org/10.1515/biolog-2017-0006
Sherif NA, Senthil Kumar T, Rao MV (2020) DNA barcoding and genetic fidelity assessment of micropropagated Aenhenryarotundifolia (Blatt.) C.S. Kumar and F.N. Rasm: a critically endangered jewel orchid. Physiol Mol Biol Plants 26(12):2391–2405. https://doi.org/10.1007/s12298-020-00917-9
Spooner DM, McLean K, Ramsay G, Waugh R, Bryan GJ (2005) A single domestication for potato based on multilocus amplified fragment length polymorphism genotyping. Proc Natl Acad Sci 102(41):14694–14699
Szendrak E (1997) Asymbiotic in vitro seed germination, micropropagation and scanning electron microscopy of several temperate terrestrial orchids (Orchidaceae). University of Nebraska, Lincol, Nebraska
Tian HZ, Han LX, Zhang JL, Li XL, Kawahara T, Yukawa T, López-Pujol J, Kumar P, Chung MG, Chung MY (2018) Genetic diversity in the endangered terrestrial orchid Cypripedium japonicum in East Asia: insights into population history and implications for conservation. Sci Rep 8(1):6467
Vafaee Y, Mozafari AA, Baghalian K (2008) Diversity evaluation of Iranian and several exotic garlic (Allium sativum L.) clones using morphological traits Iranian. J Horticult Sci Technol 8(4):259–270
Vafaee Y, Ghaderi N, Khadivi A (2017) Morphological variation and marker-fruit trait associations in a collection of grape (Vitis vinifera L.). Sci Hortic 225:771–782
Vafaee Y, Mohammadi G, Nazari F, Fatahi M, Kaki A, Gholami S, Ghorbani A, Khadivi A (2021) Phenotypic characterization and seed-micromorphology diversity of the threatened terrestrial orchids: implications for conservation. S Afr J Bot 137:386–398. https://doi.org/10.1016/j.sajb.2020.11.010
Verma J, Sharma K, Thakur K, Sembi JK, Vij SP (2014) Study on seed morphometry of some threatened Western Himalayan orchids. Turk J Bot 38:234–251. https://doi.org/10.3906/bot-1307-14
Virk PS, Ford-Lloyd BV, Jackson MT, Pooni HS, Clemeno TP, Newbury HJ (1996) Predicting quantitative variation within rice germplasm using molecularvmarkers. Heredity 76:296–304
Yeung EC (2017) A perspective on orchid seed and protocorm development. Bot Stud. https://doi.org/10.1186/s40529-017-0188-4
Zhao X, Zou G, Zhao J, Hu L, Lan Y, He J (2020) Genetic relationships and diversity among populations of Paris polyphylla assessed using SCoT and SRAP markers. Physiol Mol Biol Plants 26(6):1281–1293. https://doi.org/10.1007/s12298-020-00808-z
Acknowledgements
This work was financially supported by the Iran National Science Foundation [Grant No. 95826350] and the University of Kurdistan. The authors are thankful to Dr. Sirwan Babaie, Department of plant breeding and genetic, University of Kurdistan for photography of the orchid species.
Funding
The current work was financially supported by the Iran National Science Foundation [grant No. 95826350] and the University of Kurdistan, Iran.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no confict of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Gholami, S., Vafaee, Y., Nazari, F. et al. Exploring genetic variations in threatened medicinal orchids using start codon targeted (SCoT) polymorphism and marker-association with seed morphometric traits. Physiol Mol Biol Plants 27, 769–785 (2021). https://doi.org/10.1007/s12298-021-00978-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12298-021-00978-4