
Abstract
Cadmium (Cd) is highly toxic metal for plant metabolic processes even in low concentration due to its longer half-life and non-biodegradable nature. The current study was designed to assess the bioremediation potential of a Cd-tolerant phytobeneficial bacterial strain Bacillus sp. SDA-4, isolated, characterized and identified from Chakera wastewater reservoir, Faisalabad, Pakistan, together with spinach (as a test plant) under different Cd regimes. Spinach plants were grown with and without Bacillus sp. SDA-4 inoculation in pots filled with 0, 5 or 10 mg kg−1 CdCl2-spiked soil. Without Bacillus sp. SDA-4 inoculation, spinach plants exhibited reduction in biomass accumulation, antioxidative enzymes and nutrient retention. However, plants inoculated with Bacillus sp. SDA-4 revealed significantly augmented growth, biomass accumulation and efficiency of antioxidative machinery with concomitant reduction in proline and MDA contents under Cd stress. Furthermore, application of Bacillus sp. SDA-4 assisted the Cd-stressed plants to sustain optimal levels of essential nutrients (N, P, K, Ca and Mg). It was inferred that the characterized Cd-tolerant PGPR strain, Bacillus sp. SDA-4 has a potential to reduce Cd uptake and lipid peroxidation which in turn maintained the optimum balance of nutrients and augmented the growth of Cd-stressed spinach. Analysis of bioconcentration factor (BCF) and translocation factor (TF) revealed that Bacillus sp. SDA-4 inoculation with spinach sequestered Cd in rhizospheric zone. Research outcomes are important for understanding morpho-physio-biochemical attributes of spinach-Bacillus sp. SDA-4 synergy which might provide efficient strategies to decrease Cd retention in edible plants and/or bioremediation of Cd polluted soil colloids.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Heavy metal accumulation in agricultural soil have increased to a large extent and causes health and environmental problems worldwide. Cadmium (Cd), which enters the environment from natural and anthropogenic activities such as mining industry and excessive use of fertilizers, is a naturally occurring toxic trace metal found at concentrations of 0.1–0.5 ppm in earth’s crust. Although Cd is non-essential for plants and humans but gets retained easily by plant’s roots and shoots that renders it a metal of high concern regarding its deposition in food chain (Cai et al. 2020). Cadmium-mediated toxicity in plants extends throughout the physiological, morphological and molecular levels and results in stunted growth by disrupting the essential plant mechanisms and by decimating the membrane integrity (Rizwan et al. 2018). Cadmium stress decreases plant height and initiate mechanisms of leaf necrosis and chlorosis (Hussain et al. 2019). Moreover, it obstructs plant photosynthetic apparatus by reduced uptake of mineral elements and denaturation of photosynthetic enzymes thereby causing less plant biomass accumulation (Hussain et al. 2019; Khan et al. 2019). It also trigger the formation of reactive oxygen species (ROS) i.e., hydroxyl radicle and hydrogen peroxide which can denature essential macromolecules like lipids, proteins and nucleic acids (Singh et al. 2019). Being divalent cation, Cd competes for its transport with different nutrients at absorption sites which in turn decreases the uptake of vital nutrient ions from soil i.e., Zn, Mn, Ca, Mg, Fe (Shahid et al. 2019).
Microbial world, especially plant growth-promoting rhizobacteria (PGPR), operates in cooperation with plants to boost soil properties and to establish beneficial relationships with plants even under abiotic stress (Ramakrishna et al. 2020). Various strategies are employed by PGPR to cope with Cd phytotoxicity which include release of metal chelating molecules, biosorption and bioaccumulation (Khanna et al. 2019). Furthermore, PGPR directly influence the growth of plants under stressed conditions by synthesizing phytohormones like indole-3-acetic acid, enzymes like ACC-deaminase and by nutrient mobilization (Shahid et al. 2019).
Application of PGPR is a promising area of research in the field of bacterially-assisted bioremediation technology and efficiency of the process depends on plant’s tolerance to heavy metals and its association with beneficial bacteria (Shahid et al. 2018; Farhangi-Abriz et al. 2020). Bio-augmentation of metal tolerant soil bacteria is significantly important for successful implementation of bioremediation technology, especially in the case where such bacteria are equipped with several phytobeneficial traits (Ojuederie and Babalola 2017). Moreover, ACC-deaminase-producing PGPR play a crucial role in plant growth enhancement, reduction in stress level and bioremediation through different biochemical mechanisms and approaches. Plant inoculation with ACC-deaminase-producing PGPR alleviates stress related drastic effects by metabolizing excessive ACC into α-ketobutyric acid and ammonia which regulates the levels of endogenous stressor ethylene. Furthermore, extensive root system development via IAA production warrants the uptake of essential nutrients and water from soil (Akram et al. 2016; Shahid et al. 2020). A reduction in cellular reactive oxygen species (ROS) levels together with higher levels of antioxidants has also been reported for PGPR inoculated plants (Akram et al. 2016; Khan et al. 2016; Afridi et al. 2018).
Spinach (Spinacia oleracea L.), a member of Amaranthaceae family, is native plant of central and western Asia. Metal accumulation in edible parts of plants species, particularly of leafy vegetables, is dangerous for health of consumers (Chary et al. 2008). Heavy metal entrance and accumulation in edible parts of vegetables generally occurs through roots (Younis et al. 2016). Cadmium intake by vegetables consumption contributes human exposure to Cd from 70% up to more than 90%. Although consumers prefer dark green and undamaged leafy vegetables as a good character but safety of vegetable contamination cannot be guaranteed through mere visual observation of external morphology (Sarwar et al. 2010).
Faisalabad, a city in northeast Punjab, Pakistan, contains several active textile industrial units. Paharang drain is the reservoir for industrial and household wastewater originating from western region of Faisalabad which discharges it to Chakera wastewater collection and subsequently to Chenab River. Since metal concentrations at Chakera’s wastewater generally exceeds the WHO standards (Yasmeen et al. 2014), it was perceived that the site holds heavy metal tolerant-plant growth promoting (HMT-PGP) bacteria. Present study was, therefore, aimed at isolation, identification and characterization of HMT-PGP from Chakera wastewater collection point as well as to evaluate the potential of screened PGPR for spinach (Spinacia oleracea L.) growth and physio-biochemical modulations in Cd polluted soil. Furthermore, dissection of responsible mechanisms for PGPR-mediated adaptations in plant growth, under Cd stress, together with their association with the plant nutrient physiology and antioxidative machinery was also within the scope of present work.
Materials and methods
Wastewater sampling site and isolation of bacteria
Wastewater samples were taken from a collection point near Chakera, (31°32,025″N and 73°28,045″E) Faisalabad, Pakistan in sterilized polythene bags. The bacterial isolation was executed by dilution plate method (Somasegaran and Hoben 1994). An amount of 1 ml from each of the four samples, collected from different wastewater treatment ponds, was poured in saline solution tubes (0.85% NaCl) and serially diluted. The bacterial colonies were isolated by spreading 100 µl from dilution 10–5 and 10–7 on nutrient-agar plates having 25 mM CdCl2. The bacterial colonies were purified by repeated-streaking and confirmation of purity was made under light microscope (Olympus, Japan). The pure cultures were stored at − 80 °C in the form of glycerol stocks.
Characterization for metal tolerance and plant-beneficial traits
Measurement of Cd-tolerance
The Cd-tolerance of isolated bacterial strains was estimated by the minimum inhibitory concentration (MIC) of CdCl2. Each of the purified isolates was cultured in nutrient-broth medium and cell density was equalized by measuring the growth at 600 nm by UV–Vis spectrophotometer (Shimadzu UV/VIS, Kyoto, Japan). An amount of 200 µl from each culture was spread on nutrient-agar plates with different CdCl2 levels (0, 10, 25, 50, 75, 100, 125, and 150 mM). The MIC was calculated by visually observing the growth of bacterial colonies on each plate.
Phosphate solubilization
Phosphate solubilization ability of isolated strains was estimated with 25 mM CdCl2 concentration due to the fact that all the isolates were found tolerant to this level. Purified isolates were spot-inoculated on Pikovskaya’s-agar plates (Pikovskaya 1948) supplemented with different levels (0, 10, 25, 50, 75 and 100 mM) of CdCl2 and incubated for 240 h in an orbital shaker (150 rpm) at 30 ± 2 °C. The cultures were observed for the formation of halo-zones of phosphate solubilization after 240 h and solubilization index was calculated by the formula given below;
Solubilization index = (Diameter of colony + Zone Diameter) / Diameter of colony.
Production of indole-3-acetic acid
To estimate the potential of IAA biosynthesis, each of the isolates was cultured in 100 ml nutrient broth with or without 100 mg L-1 tryptophan as well as supplemented with similar CdCl2 concentrations as mentioned in previous section. The incubation period of the cultures was 48 h at 30 ± 2◦C under constant shaking and non-inoculated control was also run to measure and subtract the abiotic activity. Afterwards, the supernatant was collected by centrifugation of the cultures at 13,000 g and acidified with HCl to pH 2.8. After drying at room temperature, the supernatants were extracted with the help of equal volume of ethyl acetate, mixed with 1 ml methanol and filtered by using nylon filter (0.2 µm) (Tien et al. 1979). Sample analysis was carried out with high-performance liquid chromatography (wavelength = 260 nm) equipped with C-18 column and Turbochrom software (Perkin Elmer, USA). Mobile phase used during analysis was methanol/ acetic acid (30:70, v/v) at a flow rate of 0.5 ml/min.
1-Aminocycloproane-1-carboxylic acid (ACC) deaminase activity
The ability of isolated strains to use ACC (3 μL, 0.5 M) as sole nitrogen source was assessed by quantifying the amount of α-ketobutyrate produced after ACC catabolism on DF salt minimal broth supplemented with the same concentrations of CdCl2 as described in earlier section. The cultures were grown in the medium at 30 ± 2 °C for 48 h and the non-inoculated control was also run to measure and subtract the abiotic enzyme activity. Absorbance of the inoculated tubes was compared with non-inoculated ones in the presence or absence of ACC (Penrose and Glick, 2003). ACC deaminase enzyme activity was assessed by measuring the α-ketobutyrate produced by each isolate through the spectrophotometer (Shimadzu UV/VIS, Kyoto, Japan). A standard curve of α-ketobutyrate in a range from 0.1 to 1.0 μM was drawn for comparative measurements. The protein concentration of cell extracts was calculated by using Bradford methodology (Bradford 1976).
Molecular identification
Isolate SDA-4 was selected based on its Cd tolerance level and plant growth-promoting attributes. The isolate was subjected to DNA isolation by CTAB method (Wilson 2001) followed by the quantification of DNA by the Nano Drop™ 2000/2000c (Thermo Fisher.
Scientific, Waltham, MA, USA). Gene amplification of 16S rRNA was achieved by using the primer set fD1 (5′-AGAGTTTGATCCTGGCTCAG-3′) and rD1 (5′-AAGGAGGTGATCCAGCC-3′) (Weisburg et al. 1991). The amplicons were directly sequenced from Macrogen, South Korea by Sanger method. The sequence retrieved from the commercial company was analyzed, trimmed and subjected to contig formation with the help of online CAP3 Sequence Assembly Program. The final contig sequence was further analyzed by Nucleotide BLAST tool of NCBI and by constructing a phylogenetic tree with the type strain through MEGA 7.0 software package. Furthermore, the sequence was deposited to NCBI database and accession number (MT259045) was obtained.
Rifampicin-resistant derivatives and comparative growth studies
The strain SDA-4 was made resistant to rifampicin antibiotic before inoculating to spinach plants in pots so that it could be recovered from roots on the antibiotic-containing media in the presence of several antibiotic-sensitive indigenous microbes. For this purpose, the strain was cultured in nutrient-broth medium amended with 25 mM CdCl2 and 100 µl volume of this culture was spread on nutrient-agar plates containing 50 µg ml−1 rifampicin. The plate that randomly harbored some resistant colonies was selected and the colonies were sub-cultured on the next antibiotic-containing plates. The resistant strain was compared with the normal one in terms of growth by measuring the absorbance in triplicate at 600 nm using UV–VIS spectrophotometer (Hitachi U-2910, Tokyo, Japan) and by constructing a growth curve (Shahid et al. 2012). In the pot experiment, the strain was recovered again on rifampicin-added agar plates after harvesting by plate count method (Somasegaran and Hoben 1994).
Pot experiment
Experimental soil
Upper soil layer (0–15 cm) was sampled from botanical research station of Government College University, Faisalabad (31°24/N, 73°04/E) situated in flat plains of Punjab, Pakistan. Experimental soil was sandy loam having 1.8% organic matter, EC of 370 dS/m and slightly alkaline pH of 7.64. The Cd concentration of soil was found to 0.26 mg kg−1 with 660 mg kg−1 N, 8.10 mg kg−1 P, 840 mg kg−1 extractable Ca, 71.8 mg kg−1 extractable Mg and 378 mg kg−1 extractable K before any external additions were made. The soil was processed (mixed, air dried, ground and sieved) and autoclaved before using in pot experiment.
Seed sterilization, inoculation and pot experiment
Spinach seeds (cv. All Green SCR-016–202) were provided by vegetable section of Ayub Agricultural Research Institute (AARI), Faisalabad, Pakistan. Surface sterilization was carried out by dipping the seeds in sodium hypochlorite (5%, w/v) solution for 5 min followed by rinsing with sterile water. Seeds were placed on nutrient agar plates for 48 h in order to confirm their sterility. Bacillus sp. SDA-4 inoculum was prepared by growing it up to 109 CFU ml−1. The culture was washed twice with sterile water after centrifugation at 8,000 g and cells were re-suspended in equal volume of ddH2O and diluted to 108 CFU ml−1. The seeds were soaked in the inoculum for 60 min, while non-inoculated seeds dipped in sterile water were taken as control.
Twenty four plastic pots (30 cm width × 60 cm depth) were filled with 12 kg of autoclaved soil to grow Bacillus sp. SDA-4 inoculated and non-inoculated spinach seeds with three CdCl2 treatments which were grouped as [T1: 0 mg CdCl2 without SDA-4 inoculation; T2: 0 mg CdCl2 with SDA-4 inoculation, T3: 5 mg/kg CdCl2 without SDA-4 inoculation, T4: 5 mg/kg CdCl2 with SDA-4 inoculation, T5: 10 mg/kg CdCl2 without SDA-4 inoculation, T6: 10 mg/kg CdCl2 without SDA-4 inoculation].
In each pot, 15 seeds were sown in the beginning which was thinned to 10 plants after emergence. Plants were maintained for 10 weeks and irrigated with half-strength Hoagland’s solution (Arnon and Hoagland 1940) or alternatively with distilled water with three days interval. There were four replicates for each treatment grown in greenhouse in completely randomized design (CRD) with 14/10 h day/night cycle and 25/15 °C ± 2 day/night temperatures. During the course of the experiment, mean relative humidity was maintained to 65% during daytime. Subsequently spinach plants were harvested and roots were thoroughly rinsed with distilled water to remove the debris. Standard methods were used to measure morphological traits (root and leaf length, fresh and dry weight) of inoculated and non-inoculated spinach plants. Leaf area of individual plants was assessed by using leaf area meter (L12000, L1-COR, USA). Half of the plants from each pot were preserved in a freezer for further analysis of morpho-physiological parameters while the other half was oven-dried to be used for analysis of mineral elements.
Lipid peroxidation and protein contents
Spinach leaves were assessed for lipid peroxidation in terms of malondialdehyde (MDA) contents measured by thiobarbituric acid (TBA) reaction according to the methodology of Heath and Packer (1968). Protein estimation in spinach leaf tissues was executed by using Bradford (1976) methodology. Plant material (0.25 g) was extracted in phosphate buffer, maintained at a pH of 7.8 and centrifuged at 10,000 × g for 15 min at 4 °C. An aliquot was subsequently mixed with 2 ml Bradford reagent, heated in waterbath for 20–30 min and the optical density was measured by using UV–VIS spectrophotometer at 595 nm. The bovine serum albumin (BSA) standards were used to make the standard curve and to estimate the protein contents.
Chlorophyll contents
Plant material (0.25 g) was kept overnight after homogenization in 80% acetone for measurement of chlorophyll contents. The solutions were centrifuged for 5 min at 10,000 g and chlorophyll a and b contents were measured with a spectrophotometer according to the methodology of Arnon (1949).
Anthocyanin content
For quantification of anthocyanin contents, 50 mg of fresh plant material was mixed with 250 μl of acidic methanol (1% HCl, w/v). The plant material was incubated for 1 h at 4 °C with thorough shaking after homogenization on ice. The suspensions were centrifuged for 5 min at 14,000 g and absorption of the extracts was measured at 530 and 657 nm photometrically. Following equation was used to calculate plant anthocyanin contents;
Anthocyanin contents = (A 530 – 0.25* A 657) × M−1.
Phenolic content
Plant shoots (0.5 g) were homogenized in 80% acetone and centrifuged at 10,000 g at 4 ºC for 10 min to quantify the phenolic contents. 100 ml supernatant was diluted with water (2 ml) and Folin–Ciocalteau’s phenol reagent (1 ml). Five ml of Na2CO3 (20% w/v) was added, the volume was made up to 10 ml with ddH2O and absorbance was measured spectrophotometrically at 750 nm. The phenolic contents were quantified by comparison with standards as mg g−1 FW of shoots (Julkunen-Tiitto 1985).
Proline content
Proline contents of spinach shoots were calculated by using the methodology of Bates et al. (1973) where the absorbance of the chromophore-containing toluene was measured spectrophotometrically at 520 nm. The free proline content was quantified by comparison with standard curve and expressed as µmol g-1 FW of shoots.
Measurement of POD and SOD activity
Peroxidase (POD) activity of spinach shoots was quantified based upon guaiacol oxidation in reaction mixture comprising of 100 μl enzymatic extract, 50 mM phosphate buffer (with pH 7.8), 20 mM guaiacol and 40 mM H2O2. The absorbance was recorded spectrophotometrically with an interval of 20 s at 470 nm. Unit activity of POD was equated to the change of absorbance 0.01 min−1 mg−1 of protein (Chance and Maehly 1955). The superoxide dismutase (SOD) was measured by photochemical oxidation and nitroblue tetrazolium (NBT) reduction with a spectrophotometer at 560 nm.
Determination of cadmium and mineral elements
Before elemental analysis, plants were rinsed twice in distilled water, dipped in 20 mM EDTA and washed twice with deionized water again in order to remove metals adsorbed on plant surfaces. The washed plant samples were oven-dried at 105 °C for 24 h and 0.1 g dry roots and shoots material from each treatment replicate was wet digested in a digestion mixture of HNO3: HClO4 (7:3 V/V) (EPA3010A 1996). Digested samples thus obtained from each of the four replicates of applied treatments were diluted to 50 ml with distilled water and root and shoot Cd, K, Ca and Mg were determined with atomic absorption spectrophotometer (Hitachi U-2910, Tokyo, Japan). The digested material was analyzed for P and N contents spectrophotometrically by using Barton’s reagent and method of Bremner and Keeney (1965) respectively.
Root bio-concentration and translocation factor
Root bio-concentration factor (BCF) and translocation factor (TF) for Cd were computed as follows;
BCFroot = Root Cd concentration (µg/g) / Soil Cd concentration (µg/g).
TF = Shoot Cd concentration (µg/g) / Root Cd concentration (µg/g).
A given plant species with lower BCF and TF values depicts its inability to extract large amount of metals from soil and limited metal translocation to areal parts respectively.
Statistical analysis
The data, thus, collected was executed for analysis of variance (Steel et al. 1997) and treatment means were compared by least significant difference method (Fisher’s LSD) at p value of ≤ 0.05. Whenever necessary, data was normalized by inverse or logarithmic transformations before statistical analysis. The XLSTAT software version 2016.1 was used for execution of principle component analysis (PCA) and correlation analysis.
Results
Cadmium tolerance and plant-beneficial traits
Out of the eight isolates purified, SDA-4 was found to tolerate highest level of metal (100 mM, of CdCl2) (Table 1). Interestingly, this isolate was characterized to possess all the tested plant-beneficial traits and it dominated the other isolates in terms of phytobeneficial traits. Strain SDA-4 demonstrated highest solubilization index (4.01), IAA synthesis (24.65 µg ml−1), and α-ketobutyrate activity (576.14 nmol mg−1 protein h−1) as compared to other isolates. The other isolates purified in this study were found to have low cadmium tolerance or lacked one or more in vitro plant-beneficial traits, and thus, strain SDA-4 was selected for further experiments.
Taxonomic identification
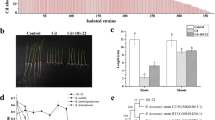
The 1465 bp 16S rRNA gene sequence exhibited 99.93% sequence identity with Bacillus cereus strain SJ37 (MT103054) and D16-1 (MK757979) and same level of identity was also shown by this sequence with the 16S rRNA gene of Bacillus paramycoides strain VITGJ1 (MH107107) and Bacillus thuringiensis strain St-2 (KC201677). In the phylogenetic tree constructed with the maximum likelihood method, the strain SDA-4 clustered with Bacillus cereus NCDO 1771 (X55060). Due to the same level of BLASTn similarity in NCBI with more than one Bacillus species, the strain SDA-4 was not identified up to species level and hence named as Bacillus sp. SDA-4 (Fig. 1).

Phylogenetic relationships of 16S rRNA gene sequences of Bacillus sp. SDA-4 with other type strains of genus Bacillus. The tree was constructed using the Maximum-Likelihood Method and 16S rRNA gene sequence data set. The nucleotide substitutions per site, indicating evolutionary distances, were calculated using Maximum Composite Likelihood model. Bootstrap values (50% or above) are shown at each branch point. To root the tree E. coli ATCC 11775 T was used as out-group
Rifampicin-resistant derivatives of strain SDA-4 and comparative growth studies
The strain SDA-4 was made resistant up to 50 µg ml−1 rifampicin in order to recover it from the spinach rhizosphere on antibiotic-containing nutrient-agar media inhibiting the growth of indigenous microorganisms. In growth studies, the rifampicin-resistant derivative (SDA-4rif) was found to exhibit similar growth behavior to that of wild type strain (SDA-4w), which revealed that SDA-4rif was fit to be inoculated with spinach plants (Fig. 2). The strain SDA-4 was recovered from the rhizosphere of spinach plants up to Log 6 CFU g−1 of rhizosphere soil by the end of pot experiment (Table 4).

Comparative growth curve of SDA-4 wild-type and rifampicin-resistant derivative strains showing the fitness of derivative strain to be inoculated with spinach plants
Biomass accumulation and photosynthetic pigments
Applied Cd stress decreased the biomass accumulation and chlorophyll contents of spinach plants maintained without Bacillus sp. SDA-4rif at 10 mg applied Cd stress (Table 2). Growth parameters of spinach were significantly promoted by the application of Bacillus sp. SDA-4rif strain. Bacillus sp. SDA-4rif inoculated spinach plants depicted 18.46%, 29.22% and 56.44% increase in leaf length, leaf width and leaf area, respectively at highest Cd stress level of 10 mg as compared to non-inoculated control plants. Likewise, Bacillus sp. SDA-4rif significantly improved the fresh weight at higher (10 mg Cd) Cd stress as compared to non-inoculated plants at same Cd stress level. Inoculation of Bacillus sp. SDA-4rif improved the plant fresh and dry weights up to 36.13% and 76.32% respectively when compared with non-inoculated plants at maximum Cd toxicity.
The results revealed that inoculation of Bacillus sp. SDA-4rif significantly improved the Chl a (25.71%), Chl b (58.78%) and total chlorophyll contents (38.20%) at 10 mg Cd stress as compared to non-inoculated ones.
Malonaldehyde (MDA) content
Without Bacillus sp. SDA-4rif application, a significant increase in lipid peroxidation level in spinach plants was observed at all Cd treatments (Table 3). MDA content was significantly reduced up to 10.22% at 10 mg Cd stress in inoculated plants as compared to non-inoculated ones.
Total protein and total phenolics
Significant increase in total protein and phenolic contents was observed in Bacillus sp. SDA-4rif inoculated spinach plants which exhibited 53.53% and 38.98% increase, respectively at 10 mg Cd as compared to non-inoculated ones (Table 3).
Antioxidant activity, proline and anthocyanin contents
Increased proline contents together with reduction in anthocyanin and antioxidants (SOD and POD) were recorded in non-inoculated spinach plants with the increase in applied Cd stress levels (Table 3). At 10 mg applied Cd stress level, spinach plants maintained with Bacillus sp. SDA-4rif exhibited higher biosynthesis for proline (10.39%), anthocyanin (47.23%), SOD (34.46%) and POD (20.36%) as compared to their respective non-inoculated controls.
Analysis of plant nutrient acquisition
The results on effects of Bacillus sp. SDA-4rif inoculation and non-inoculation on N, P, K, Mg and Ca accumulation in spinach roots and shoots at different levels of Cd stress are presented in Table 4. Cd toxicity at all applied levels significantly reduced the root and shoot accumulation of N, P, K, Mg and Ca in non-inoculated spinach plants. However, plants receiving the inoculation of Bacillus sp. SDA-4rif increased the uptake of N, P, K, Mg and Ca in spinach roots (51.9, 12.29, 43.7, 18.6, 16.23% increase, respectively) and shoots (56.62, 15.69, 52.4, 40.7, 28.60% increase), respectively at 10 mg Cd stress as compared to non-inoculated plants.
Cadmium uptake as well as bio-concentration and translocation factors
Cadmium uptake and translocation was increased significantly in non-inoculated spinach plants as the concentrations of applied Cd increased, however, Bacillus sp. SDA-4rif inoculation diminished Cd accumulation (Table 4). The bio-augmentation of Bacillus sp. SDA-4rif decreased Cd uptake by 50.41% and its translocation up to 59.43% at 10 mg Cd level as compared to the same level of Cd stress in non-inoculated plants. Analysis of bio-concentration factor (0.665) and translocation factor (0.692) for Cd depicted lower Cd retention in spinach at 10 mg Cd stress as compared to the corresponding non-inoculated values (1.34 and 0.845 respectively). Soil samples from all treatments were found to have Cd contents of 0.48, 0.50, 3.75, 3.87, 6.773 and 6.96 µg/g of dry soil for T1 to T6 respectively at the end of experiment.
Relationship between Cd uptake and morpho-physiological attributes of spinach
Pearson correlation (correlogram) analysis was conducted to quantify the relationship between Cd uptake in roots and shoots of spinach with plant growth and biomass, chlorophyll contents, lipid peroxidation, antioxidant and nutrient uptake (Fig. 3). Root Cd contents were positively correlated with shoot Cd contents, bioaccumulation factor (BCF) and translocation factor (TF) and proline contents while negatively correlated with plant growth and biomass, photosynthetic pigments, oxidative stress and antioxidative response. Chlorophyll contents were positively correlated with plant biomass accumulation. Positive correlations were also identified among growth attributes and N, P, K, Ca and Mg contents.

Correlations (r values) among different studied parameters of spinach grown under different Cd stress levels and inoculated with and without Bacillus sp. SDA-4rif. L L = leaf length, L W = leaf width, L A = leaf area, P FW = plant fresh weight, P DW = plant dry weight, Chl a = chlorophyll a, Chl b = chlorophyll b, T Chl = total chlorophyll, T Prot = total protein, T Phenol = total phenolics, Pro = proline, Antho = anthocyanin, MDA = malondialdehyde, SOD = superoxide dismutase, POD = peroxidase, N = nitrogen, P = phosphorus, K = potassium, Ca = calcium, Mg = magnesium, Cd = cadmium, BCF = bio-concentration factor, TF = translocation factor
Principal component analysis
In order to evaluate the combinatorial effect of Cd and Bacillus sp. SDA-4rif application on important attributes of spinach plants, score and loading plots of principal component analysis (PCA) were prepared (Fig. 4). All three applied Cd treatments with and without inoculation were successfully dispersed by first two principal components (Fig. 4a). The maximum variance among all the components was based on extracted components i.e., PC1 (Dim1) and PC2 (Dim2) where component Dim1 contributed 77.4% while the contribution of Dim2 was15.2% (Fig. 4b).

Score (a) and loading plot (b) of principal component analysis (PCA) on different study attributes of spinach plants inoculated with or without Bacillus sp. SDA-4rif while grown under Cd stress. Score plot represents separation of treatments as T1: 0 mg CdCl2 without SDA-4 inoculation; T2: 0 mg CdCl2 with SDA-4 inoculation, T3: 5 mg/kg CdCl2 without SDA-4 inoculation, T4: 5 mg/kg CdCl2 with SDA-4 inoculation, T5: 10 mg/kg CdCl2 without SDA-4 inoculation, T6: 10 mg/kg CdCl2 without SDA-4 inoculation. TF = translocation factor, MDA = malondialdehyde, Cd = cadmium, Pro = proline, R L = root length, T Chl = total cholorophyll, N = nitrogen
Such treatment distribution clearly represented the ameliorative effect of Bacillus sp. SDA-4rif application on studied parameters of spinach when compared to non-inoculated control plants. The Cd treatments without Bacillus sp. SDA-4rif i.e., 5 mg/kg (3) and 10 mg/kg (5) were more separated from the treatments such as, non-inoculated control (1); Bacillus sp. SDA-4rif inoculation under 5 mg/kg Cd (4) and Bacillus sp. SDA-4rif inoculation under 10 mg/kg Cd (6) (Fig. 4a), which depicted the Cd-induced hazardous effects on growth and morpho-physiology of spinach plants.
Discussion
Physiological and molecular characterization of Bacillus sp. SDA-4
Wastewater collection point, Chakera, was selected due to its higher probability of prevalence for metal tolerant bacteria as elevated concentrations of heavy metals were received here from various industrial units. These metal tolerant bacteria were presumed to carry PGP traits and hence, eight bacterial strains were purified and evaluated for Cd-tolerance potential along with their in vitro measurement of PGP characteristics. The selection of only one isolate (SDA-4) for further studies was based on the exhibition of superior in vitro PGP traits under Cd stress (Table 1). The molecular identity as Bacillus sp. was based on the phylogenetic analysis of its 16S rRNA gene and we were unable to describe the taxonomic rank up to species level due to same level of sequence similarity with more than one Bacillus species. Species, belonging to genus Bacillus, have previously been reported as PGPR in various studies (Kaymak 2011; Shahid et al. 2015). In vitro phosphate solubilizing ability of Bacillus sp. SDA-4 is attributed to its acid production, an activity responsible to establish halo zone on the culture media. However, the exhibition of maximum phosphate solubilizing ability in Cd-contaminated culture medium not only made this strain a choice but also revealed an added advantage to be inoculated with plants under metal stressed conditions (Shahid et al. 2012). Similarly, there is a proactive relationship between IAA synthesis and bacterial ACC deaminase activity especially when encountering the abiotic stresses (Glick 2014). Hence, the ability of Bacillus sp. SDA-4 to produce both IAA and ACC deaminase under in vitro conditions in the presence of Cd depicted the PGP traits carried by the strain (Akram et al. 2016; Shahid et al. 2018). Before starting the pot experiment, the strain SDA-4 was made rifampicin-resistant to recover the mutant strain from the rhizosphere in the presence of many other indigenous microbes sensitive to the antibiotic. Since, the growth pattern of Bacillus strain SDA-4rif was observed similar to its wild-type counterpart (Fig. 2), therefore, it was anticipated that the rifampicin-resistant strain might have retained PGP traits identical to the wild-type strain.
Impact of Bacillus sp. strain SDA-4rif inoculation on growth of Cd stressed spinach
Plants exhibit impaired growth by various mechanisms i.e., reduced root and leaf length, disrupted photosynthetic rates and oxidation of cellular membranes under metal stress. In the present study, significant reduction of plant fresh and dry biomasses of non-inoculated spinach was recorded under Cd stress as compared to plants without Cd application in a dose-dependent manner (Table 2, Fig. 3). Furthermore, significant decrease in dry biomasses than the fresh ones exhibited that applied Cd stress deciphered more negative impact on photosynthesis of spinach as compared to its water relations. Our results corroborated the findings that soil Cd pollution abridged the biomass accumulation of Linum usitatissimum L. (Shahid et al. 2019).
Inoculation with Bacillus sp. SDA-4rif significantly increased the biomass, growth and chlorophyll contents of spinach plants as compared to non-inoculated ones under applied Cd regimes (Table 2, Fig. 4). Earlier, inoculation of Bacillus sp. PM14 has been reported to augment the growth parameters of metal stressed Sesbania sesban (Din et al. 2020). Likewise, the increase in spinach plant dry and fresh biomass in roots and shoots biomasses might result from PGPR-mediated phosphate solubilization, IAA and siderophore production with concomitant metal stress resistance (Zafar-ul-Hye et al. 2020). The application of Bacillus licheniformis inoculation increased the growth and chlorophyll biosynthesis by modulating the nitrogen fixation, siderophore production, phytohormones production together with secretion of secondary metabolites in spinach plant which acts as insecticides while growing under Cd and As toxicity (Asif et al. 2020). Plants also respond to metal stress by moderating in planta hormonal levels which trigger the expression of different proteins required for plant protection against stresses. Ethylene is one of them, however, its overproduction directly impairs chlorophyll production, plant growth and development (Iqbal et al. 2017). Plant can be relieved from ethylene stress with the help of ACC deaminase-producing bacterial strains co-existing with their roots under harsh environmental conditions which can either store excessive amount of plant’s ACC or by converting it to ammonia and α-ketobutyric acid. Furthermore, IAA production by PGPR enhanced plant growth, adventitious roots production, root elongation and development which improve water and nutrient acquisition from soil (Spaepen et al. 2007; Mohite, 2013). Such a situation is likely to overcome Cd toxicity and stimulate root and shoot growth (Sytar et al. 2018).
The insoluble phosphates in soil matrix are mobilized and made available to plants by bacterial application (Zaidi et al. 2009). In present study, increased spinach biomass was envisioned owing to contribution of ACC-deaminase activity, phosphate-solubilization and IAA production of Bacillus sp. SDA-4 during plant growth.
Reduction in chlorophyll contents of spinach was recorded when grown without Bacillus sp. SDA-4rif under applied Cd stress (Table 2, Fig. 3). The decrease in photosynthetic pigments in Cd treated spinach was possibly linked to Cd-mediated inhibition of protochlorophyllide reductase enzyme, boost of plastoglobuli, interruption of thylakoid membrane, photosystem II, diminished Mg uptake and over-production of ROS (Hussain et al. 2019; Al Mahmud et al. 2018). Improved chlorophyll (a, b) contents of spinach plant were observed under Bacillus sp. SDA-4rif application likely due to the activation of enzymatic pathways of chlorophyll biosynthesis, lower production of ROS or increased bioavailability of minerals including Mg and N (Khan et al. 2016). The microbe-assisted increase in N contents in turn act as an essential component of chlorophyll resulting in higher dry mass production of Medicago lupulina L. under Cd stress (Jian et al. 2019). Improved chlorophyll contents are likely to enhance the photosynthetic activity as depicted from increased dry biomasses of inoculated spinach plants in the present study.
Effect of Bacillus sp. SDA-4rif inoculation on lipid peroxidation and antioxidants of spinach under Cd stress
In non-inoculated spinach plants, malondialdehyde (MDA) contents were found significantly higher under Cd stress which might result from the genesis of ROS due to the breakdown of stable cellular compounds like metal-DNA, proteins-metal DNA-DNA, single or double stranded DNA (Gill et al. 2016). Higher MDA levels depicted the decomposition of proteins and biomolecules which lead to lipid peroxidation of cellular membranes (Islam et al., 2014). ROS, if not properly scavenged, disrupt the plant growth due to higher reaction rate with macro-molecular structures and vital metabolites (Richards et al. 2015) and trigger membrane disintegration and electrolyte leakage. Microbial inoculation to Cd stressed spinach plant is likely to involve in immobilization or transformation of Cd to less toxic forms rendering it unavailable for plant uptake and reduced the oxidative stress by lowering in planta MDA and H2O2 (Malook et al. 2017). Antioxidant phytochemicals such as superoxidase (SOD), peroxidase (POD) proline, phenolics and anthocyanin have a significant role in reducing ROS accumulation, eliminating malondialdehyde (MDA) to resist peroxidation of cellular lipids, maintaining the integrity of cell membrane and protecting the cellular substances form oxidation.
Bacillus sp. SDA-4rif inoculation alleviated Cd toxicity in spinach as it increased activities of SOD, POD as well as phenolics and anthocyanin contents together with decreasing levels of MDA (Table 3, Fig. 4). Our results corroborate the findings of Wan et al. (2012) who reported that Serratia nematodiphila inoculation boosted the growth and yield of Solanum nigrum by increasing the activities of enzymatic and non-enzymatic antioxidants under Cd stress. Elevated activities of antioxidants mitigated the toxic effects of abiotic stresses and application of PGPR up-regulated the antioxidants by enhancing the activities of abiotic stress related genes (El-Esawi et al. 2019). Improvement in antioxidative machinery in inoculated spinach plants might be linked with gene activation or mRNA expression encoding antioxidant enzymes (Gururani et al. 2013). For example, Serratia marcescens BM1 application in soybean increased the antioxidant genes expression (CAT, APX, POD and Cd-SOD) together with stress related genes (CHI, PHD2, CHS, P5CS and VSP) (El-Esawi et al. 2020). The production of CAT and SOD enhanced the production of acidic exopolysaccharides which act as a diffusion barrier against ROS (Wang et al., 2016). Improved antioxidant system of Cd stressed spinach by Bacillus sp. SDA-4rif inoculation can, therefore be envisioned as one of the mitigation strategies to alleviate intracellular Cd by scavenging ROS.
Nutrient acquisition patterns of Cd stressed spinach after Bacillus sp. SDA-4rif inoculation
Increasing Cd stress inhibited the root and shoot accumulation of N, P, K, Ca and Mg in non-inoculated spinach plants (Table 4, Fig. 3). It is likely that Cd-mediated N and P deficiency limits the photosynthetic rate and growth of spinach plant as recorded in the present study. Reduced K uptake in spinach might originate from its leakage into the rhizosphere resulting in decreased biosynthesis of photosynthetic pigments under Cd stress (Kurtyaka et al. 2008). Diminished Ca and Mg uptake probably occurred due to competition among Cd, Ca and Mg at uptake sites (Küpper and Kochian 2010) in non-inoculated spinach plants grown under Cd stress.
Inoculation of Cd stressed spinach plants with Bacillus sp. SDA-4rif significantly increased the accumulation of essential minerals i.e., N, P, K, Ca and Mg which was considered to occur via several mechanisms like production of ACC deaminase enzyme, IAA synthesis and phosphate solubilization (Gamalero et al. 2009). It has been reported earlier that nutrient acquisition was significantly increased with the inoculation of Planomicrobium sp. MSSA-10 in salt stressed pea plants (Shahid et al. 2018). Soil inoculated with PGPR strain initiated soil acidosis by solubilizing colloids bound phosphate through the genesis of organic acids. The release of cationic nutrients (K, Ca and Mg), bound to phosphates is thus increased by rhizospheric acidification which are ultimately bioavailable for plants (Khan et al. 2016). Bacillus sp. improved phosphate retention by siderophores formation, hormonal stimulation and root hair growth which decreased translocation of heavy metals (Gupta et al. 2018). It was also inferred that improved uptake of nutrients (N, P, K, Ca, and Mg) by Cd stressed spinach owing to higher root surface area which offers higher nutrient binding sites upon inoculation with Bacillus sp. SDA-4 as reported earlier for broccoli plants (Yildirim et al. 2011). Root exudation increases the microbial activity of roots, thereby, resulting in improved soil-nutrient solubility and nutrient’s influxes to root cell sap (Ahemad and Kibret 2014). The microbe-assisted increase in plant N contents is acting as an essential component of chlorophyll with concomitant dry mass production of Medicago lupulina L. under Cd stress (Jian et al. 2019).
Effect of Bacillus sp. SDA-4 rif inoculation on Cd uptake by spinach
Root and shoot Cd contents of non-inoculated spinach plants was increased with rise in applied Cd treatment levels (Table 4). Cadmium uptake by plant roots, stems and leaves usually takes place via both apoplastic and symplastic pathways (Lux et al. 2011) and rhizospheric acidosis together with root exudate increased Cd bioavailability for spinach plant. Soil to root Cd transport has been regulated by three main transporters, zinc/iron-regulated transporter-like proteins (ZIP), NRAMP (OsNRAMP5, and AtNRAMP6) and low affinity calcium transporters (TaLCT1) (Corguinha et al. 2015). Root Cd uptake has also been perceived by various cation channels (Lux et al. 2011). Higher Cd treatment levels resulted in higher Cd deposition in apoplast of root cells which in turn resulted in increased plant Cd retention (Sterckeman et al. 2011) as observed in present study.
The Cd contents of Bacillus sp. SDA-4rif inoculated spinach plants were significantly decreased as compared to non-inoculated ones, suggesting that the strain SDA-4 inhibited the Cd toxicity by triggering the formation of biofilm to detoxify the metals. This might be linked with the synthesis of bacterial exopolysaccharides having higher affinity to metallic cations including Cd ions (Gupta and Diwan 2017). Reduction in Cd uptake may also arise from biological dilution as the tissue metal retention capacity diminishes with higher absorption areas originating from increased biomass production in Bacillus sp. SDA-4rif inoculated spinach plants (Javed and Greger 2011).
To keep Cd concentration in balanced physiological ranges and to detoxify the metal, plants initiate mechanisms to control metal retention, transportation and homeostasis. It was also confirmed from the present results that Bacillus sp. SDA-4rif inoculation decreased the Cd uptake (lower values for BCF) by host spinach plants as well as restricted the root to shoot acropetal Cd translocation (lower values for TF) pathways. Generally, PGPR uptake metals from contaminated soil by chemiosmotic gradient of lipomembrane or by consuming ATPs as a source of energy, sometimes, selectively leaving contaminated soil free of heavy metals (Han et al. 2020). Bacillus sp. SDA-4, being a heavy metal immobilizing bacteria, reduced Cd bioavailability to spinach through passivation effect. Bacillus sp. might be immobilizing Cd by Cd adsorption to cell wall, precipitation, or adsorption on other heavy metals with anions produced by microbial metabolism and complexation with phosphate (Han et al. 2020; Wang et al. 2020). Restricted Cd accumulation in inoculated spinach depicted that plants could be protected from metal stress in contaminated soils by synergistic application of Bacillus sp. SDA-4 and will be a potential approach to develop low Cd-accumulating spinach to be used for Cd stabilization in metal contaminated soil colloids.
Conclusions
Bacillus sp. SDA-4 isolated from wastewater reservoir, Chakera, was screened for physio-biochemical and plant growth-promoting abilities under Cd stress. It was concluded that the strain SDA-4 possesses the ability of phosphate solubilization as well as production of IAA and ACC deaminase, which enabled it to survive in stressed environment. Bacillus sp. SDA-4 synergy with spinach augments growth by improving in planta antioxidative potential and nutrient physiology together with reduced lipid peroxidation levels under Cd stress. Use of spinach as a sensitive Cd pollution indicator in our work can be considered a new way of exploring quickly the applicability of PGPR-assisted bioremediation approach. Interestingly, the strain SDA-4rif was successfully recovered from spinach rhizosphere at a considerable cell density after the harvest of experiment, which indicates the rhizosphere competitiveness of the strain. Application of fluorescent dyes like Leadmium Green AM to tag in vivo Cd ions should be executed to localize whether Cd is contained in plant’s roots cell or within the bacterial strain which ultimately restrict Cd accumulation in spinach.
6. References
Afridi MS, Amna MT, Salam A, Bibi M, Khan AA, Ali F, Mehmood S, Chaudhary HJ (2018) Induction of salinity tolerance in wheat genotypes by plant growth promoting endophytes: involvement of ACC deaminase and antioxidant enzymes. Plant Physiol Biochem 139:569–577
Ahemad M, Kibret M (2014) Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J King Saud Uni Sci 26:1–20
Akram MS, Shahid M, Tariq M, Azeem M, Javed MT, Saleem S, Riaz S (2016) Deciphering Staphylococcus sciuri SAT-17 mediated anti-oxidative defense mechanisms and growth modulations in salt stressed maize (Zea mays L.). Front Microbiol 7:867. https://doi.org/10.3389/fmicb.2016.00867
Al Mahmud J, Hasanuzzaman M, Nahar K, Bhuyan MB, Fujita M (2018) Insights into citric acid-induced cadmium tolerance and phytoremediation in Brassica juncea L.: coordinated functions of metal chelation, antioxidant defense and glyoxalase systems. Ecotoxicol Environ Safe 147:990–1001
Arnon DI (1949) Copper enzymes in isolated chloroplasts: poly phenoloxidase in Beta vulgaris. Plant Physiol 24:1–15
Arnon DI, Hoagland DR (1940) Crop production in artificial culture solution, sand and soil with special reference to factors influencing yield and absorption of inorganic nutrient. Soil Sci 50:463–483
Asif M, Pervez A, Irshad U, Mehmood Q, Ahmad R (2020) Melatonin and plant growth-promoting rhizobacteria alleviate the cadmium and arsenic stresses and increase the growth of Spinacia oleracea L. Plant Soil Environ 66(5):234–241
Bates L, Waldren R, Teare I (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39:205–207
Bradford MM (1976) A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Bremner JM, Keeney DR (1965) Steam distillation methods for determination of ammonium, nitrate and nitrite. Anal Chim Acta 32:485–495
Cai Y, Zhang S, Cai K, Huang F, Pan B, Wang W (2020) Cd accumulation, biomass and yield of rice are varied with silicon application at different growth phases under high concentration cadmium-contaminated soil. Chemosphere 242:125–128
Chance B, Maehly AC (1955) Assay of catalase and peroxidase. Method Enzymol 2:764–775
Chary NS, Kamala CT, Raj DSS (2008) Assessing risk of heavy metals from consuming food grown on sewage irrigated soils and food chain transfer. Ecotoxicol Environ Saf 69:513–524
Corguinha APB, de Souza GA, Gonçalves VC, Carvalho CDA, de Lima WEA, Martins FAD, Yamanaka CH, Francisco EAB, Guilherme LRG (2015) Assessing arsenic, cadmium, and lead contents in major crops in Brazil for food safety purposes. J Food Compos Anal 37:143–150
Dabral S, Varma A, Choudhary DK, Bahuguna RN, Nath M (2019) Biopriming with Piriformospora indica ameliorates cadmium stress in rice by lowering oxidative stress and cell death in root cells. Ecotoxicol Environ Safe 186:109741
Din BU, Amna RM, Javed MT, Kamran MA, Mehmood S, Khan M, Sultan T, Munis MFH, Chaudhary HJ (2020) Assisted phytoremediation of chromium spiked soils by Sesbania sesban in association with Bacillus xiamenensis PM14: A biochemical analysis. Plant Physiol Biochem 146:249–258
El-Esawi MA, Al-Ghamdi AA, Ali HM, Alayafi AA (2019) Azospirillum lipoferum FK1 confers improved salt tolerance in chickpea (Cicer arietinum L.) by modulating osmolytes, antioxidant machinery and stress-related genes expression. Environ Exp Bot 159:55–56
El-Esawi MA, Elkelish A, Soliman M, Elansary HO, Zaid A, Wani SH (2020) Serratia marcescens BM1 enhances cadmium stress tolerance and phytoremediation potential of soybean through modulation of osmolytes, leaf gas exchange, antioxidant machinery, and stress-responsive genes expression. Antioxidants 9(1):43
Farhangi-Abriz S, Tavasolee A, Ghassemi-Golezani K, Torabian S, Monirifar H, Rahmani HA (2020) Growth-promoting bacteria and natural regulators mitigate salt toxicity and improve rapeseed plant performance. Protoplasma. https://doi.org/10.1007/s00709-020-01493-1
Gamalero E, Lingua G, Berta G, Glick BR (2009) Beneficial role of plant growth promoting bacteria and arbuscular mycorrhizal fungi on plant responses to heavy metal stress. Can J Microbiol 55:501–514
Gill RA, Zhang NA, Ali B, Farooq MA, Xu J, Gill MB, Mao B, Zhou W (2016) Role of exogenous salicylic acid in regulating physio-morphic and molecular changes under chromium toxicity in black and yellow- seeded Brassica napus L. Environ Sci Pollut Res 23:20483–20496
Glick BR (2014) Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol Res 169:30–39
Gupta P, Diwan B (2017) Bacterial exopolysaccharide mediated heavy metal removal: a review on biosynthesis, mechanism and remediation strategies. Biotechnol Rep 13:58–71
Gupta P, Kumar V, Usmani Z, Rani R, Chandra A (2018) Phosphate solubilization and chromium (VI) remediation potential of Klebsiella sp. strain CPSB4 isolated from the chromium contaminated agricultural soil. Chemosphere 192:318–327
Gururani MA, Upadhyaya CP, Baskar V, Venkatesh J, Nookaraju A, Park SW (2013) Plant growth-promoting rhizobacteria enhance abiotic stress tolerance in Solanum tuberosum through inducing changes in the expression of ROS-scavenging enzymes and improved photosynthetic performance. J Plant Growth Regul 32:245–258
Han H, Cai H, Wang XY, Hu XM, Chen ZJ, Yao LG (2020) Heavy metal-immobilizing bacteria increase the biomass and reduce the Cd and Pb uptake by pakchoi (Brassica chinensis L.) in heavy metalcontaminated soil. Ecotoxicol Environ Safe 195:110375
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125:189–198
Hussain A, Amna KMA, Javed MT, Hayat K, Farooq MA, Ali A, Ali M, Manghwar H, Jan F, Chaudhary HJ (2019) Individual and combinatorial application of Kocuria rhizophila and citric acid on phytoextraction of multi-metal contaminated soils by Glycine max L. Environ Exp Bot 159:23–33
Iqbal N, Khan NA, Ferrante A, Trivellini A, Fracini A, Khan MIR (2017) Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Front Plant Sci 8:475
Islam F, Yasmeen T, Riaz M, Arif MS, Ali S, Raza SH (2014) Proteus mirabilis alleviates zinc toxicity by preventing oxidative stress in maize (Zea mays) plants. Ecotoxicol Environ Safe 110:143–152
Javed MT, Greger M (2011) Cadmium triggers Elodea canadensis to change the surrounding water pH and thereby Cd uptake. Int J Phytoremediat 13:95–106
Jian L, Bai X, Zhang H, Song X, Li Z (2019) Promotion of growth and metal accumulation of alfalfa by coinoculation with Sinorhizobium and Agrobacterium under copper and zinc stress. Peer J 7:e6875
Julkunen-Tiitto R (1985) Phenolic constituents in the leaves of northern willows: methods for the analysis of certain phenolics. J Agric Food Chem 33:213–217
Kaymak HC (2011) Potential of PGPR in agricultural innovations. In: Maheshwari DK (ed) Plant growth and health promoting bacteria. Springer, Berlin, pp 45–79
Khan A, Sirajuddin ZXQ, Javed MT, Khan KS, Bano A, Shen RF, Masood S (2016) Bacillus pumilus enhances tolerance in rice (Oryza sativa L.) to combined stresses of NaCl and high boron due to limited uptake of Na+. Environ Exp Bot 124:120–129
Khan MY, Prakash V, Yadav V, Chauhan DK, Prasad SM, Ramawat N, Sharma S (2019) Regulation of cadmium toxicity in roots of tomato by indole acetic acid with special emphasis on reactive oxygen species production and their scavenging. Plant Physiol Biochem 142:193–201
Khanna K, Jamwal VL, Gandhi SG, Ohri P, Bhardwaj R (2019) Metal resistant PGPR lowered Cd uptake and expression of metal transporter genes with improved growth and photosynthetic pigments in Lycopersicon esculentum under metal toxicity. Sci Rep 9:5855
Küpper H, Kochian LV (2010) Transcriptional regulation of metal transport genes and mineral nutrition during acclimatization to cadmium and zinc in the Cd/Zn hyperaccumulator, Thlaspi caerulescens (Ganges population). New Phytol 185:114–129
Kurtyaka R, Małkowski E, Kita A, Karcz A (2008) Effect of calcium and cadmium on growth and accumulation of cadmium, calcium, potassium and sodium in maize seedlings. Pol J Environ Stud 17:51–56
Lux A, Martinka M, Vaculík M, White PJ (2011) Root responses to cadmium in the rhizosphere: A review. J Exp Bot 62:21–37
Malook I, Rehman SU, Khan MD, El-Hendawy SE, Al-Suhaibani NA, Aslam MM, Jamil M (2017) Heavy metals induced lipid peroxidation in spinach mediated with microbes. Pak J Bot 49(6):2301–2308
Mohite B (2013) Isolation and characterization of indole acetic acid (IAA) producing bacteria from rhizospheric soil and its effect on plant growth. J Sci Plant Nutr 13:638–649
Ojuederie OB, Babalola OO (2017) Microbial and Plant-Assisted Bioremediation of Heavy Metal Polluted Environments: A Review. Int J Environ Res Public Health 14:1504
Penrose DM, Glick BR (2003) Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol Plant 118:10–15
Pikovskaya RI (1948) Metabolism of phosphorous in soil in connection with vital activity of some microbial species. Microbiologia 17:362–370
Ramakrishna W, Rathore P, Kumari R, Yadav R (2020) Brown gold of marginal soil: Plant growth promoting bacteria to overcome plant abiotic stress for agriculture, biofuels and carbon sequestration. Sci Total Environ 711:135062
Richards SL, Wilkins KA, Swarbreck SM, Anderson AA, Habib N, Smith AG, McAinsh M, Davies JM (2015) The hydroxyl radical in plants: from seed to seed. J Exp Bot 66(1):37–46
Rizwan M, Ali S, Abbas T, Adrees M, Zia-ur-Rehman M, Ibrahim M, Nawaz R (2018) Residual effects of biochar on growth, photosynthesis and cadmium uptake in rice (Oryza sativa L.) under Cd stress with different water conditions. J Environ Manage 206:676–683
Sarwar N, Malhi SS, Zia MH, Naeem A, Bibi S, Farid G (2010) Role of mineral nutrition in minimizing cadmium accumulation by plants. J Sci Food Agric 90:925–937
Shahid M, Akram MS, Khan MA, Zubair M, Shah SM, Ismail M, Shabir G, Basheer S, Aslam K, Tariq M (2018) A phytobeneficial strain Planomicrobium sp. MSSA-10 triggered oxidative stress responsive mechanisms and regulated the growth of pea plants under induced saline environment. J Appl Microbiol 124:1566–1579
Shahid M, Hameed S, Imran A, Ali S, Elsas JDV (2012) Root colonization and growth promotion of sunflower (Helianthus annuus L.) by phosphate solubilizing Enterobacter sp. Fs-11. World J Microbiol Biotechnol 28:2749–2758
Shahid M, Hameed S, Tariq M, Zafar M, Ali A, Ahmad N (2015) Characterization of mineral phosphate-solubilizing bacteria for enhanced sunflower growth and yield attributing traits. Ann Microbiol 65:1525–1536
Shahid M, Javed MT, Masood S, Akram MS, Azeem M, Ali Q, Gilani R, Basit F, Abid A, Lindberg S (2019) Serratia sp. CP-13 augments the growth of cadmium (Cd)-stressed Linum usitatissimum L. by limited Cd uptake, enhanced nutrient acquisition and antioxidative potential. J Appl Microbiol 126(6):1708–1721
Shahid M, Shah AA, Basit F, Noman M, Zubair M, Ahmed T, Naqqash T, Manzoor I, Maqsood A (2020) Achromobacter sp FB-14 harboring ACC deaminase activity augmented rice growth by upregulating the expression of stress-responsive CIPK genes under salinity stress. Braz J Microbiol. https://doi.org/10.1007/s42770-019-00199-8
Singh S, Singh VP, Prasad SM, Sharma S, Ramawat N, Dubey NK, Chauhan DK (2019) Interactive effect of silicon (Si) and salicylic acid (SA) in maize seedlings and their mechanisms of cadmium (Cd) toxicity alleviation. J Plant Growth Regul 38(4):1587–1597
Somasegaran P, Hoben HJ (1994) Handbook for rhizobia: Methods in legumerhizobium technology. Springer, Heidelberg, NY
Spaepen S, Vanderleyden J, Remans R (2007) Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol Rev 31:425–448
Steel RGD, Torrie JH, Dickey DA (1997) Principles and procedures of statistics, a biometrical approach, 3rd edn. McGraw-Hill Co., Inc., New York NY
Sterckeman T, Redjala T, Morel JL (2011) Influence of exposure solution composition and of plant cadmium content on root cadmium short-term uptake. Environ Exp Bot 74:131–139
Sytar O, Kumari P, Yadav S, Brestic M, Rastogi A (2018) Phytohormone priming: regulator for heavy metal stress in plants. J Plant Growth Regul 1–14
Tien TM, Gaskins MH, Hubbell DH (1979) Plant growth substances produced by Azospirillum brazilense and their effect on the growth of pearl millet. Appl Environ Microbiol 37:1016–1024
Wan Y, Luo S, Chen J, Xiao X, Chen L, Zeng G, Liu C, He YE (2012) Effect of endophyte-infection on growth parameters and Cd-induced phytotoxicity of Cd-hyperaccumulator Solanum nigrum L. Chemosphere 89:743–750
Wang Q, Chen L, He LY, Sheng XF (2016) Increased biomass and reduced heavy metal accumulation of edible tissues of vegetable crops in the presence of plant growth-promoting Neorhizobium huautlense T1–17 and biochar. Agric Ecosyst Environ 228:9–18
Wang T, Wang X, Tian W, Yao L, Li Y, Chen Z, Han H (2020) Screening of heavy metal-immobilizing bacteria and its effect on reducing Cd2+ and Pb2+ concentrations in water spinach (Ipomoea aquatic Forsk.). Int J Env Res Pub He 17(9):3122
Weisburg WG, Barns SM, Pelletier DA, Lane DJ (1991) 16S ribosomal DNA amplification for phylogenetic study. J Bacteriol 173:697–703
Wilson K (2001) Preparation of genomic DNA from bacteria. Curr Protoc Mol Biol 56:2–4
Yasmeen T, Ali Q, Islam F, Ali N, Akram MS, Javed MT (2014) Biologically treated wastewater fertigation induced growth and yield enhancement effects in Vigna radiata L. Agric Water Manag 146:124–130
Yildirim E, Karlidag H, Turan M, Dursun A, Goktepe F (2011) Growth, nutrient uptake, and yield promotion of broccoli by plant growth promoting rhizobacteria with manure. Hort Sci 46:932–936
Younis U, Malik SA, Rizwan M, Qayyum MF, Ok YS, Shah MHR, Rehman RA, Ahmad N (2016) Biochar enhances the cadmium tolerance in spinach (Spinacia oleracea) through modification of Cd uptake and physiological and biochemical attributes. Environ Sci Pollut Res 23:21385–21394
Zafar-ul-Hye M, Tahzeeb-ul-Hassan M, Abid M, Fahad S, Brtnicky M, Dokulilova T, Danish S (2020) Potential role of compost mixed biochar with rhizobacteria in mitigating lead toxicity in spinach. Sci Rep 10(1):1–12
Zaidi A, Khan M, Ahemad M, Oves M (2009) Plant growth promotion by phosphate solubilizing bacteria. Acta Microbiol Immunol Hung 56:263–284
Acknowledgements
We gratefully acknowledge the provision of funds by Higher Education Commission (HEC) of Pakistan (Grant No: 20-4243/NRPU/R&D/HEC/14/885). The results presented in this paper are the part of MPhil research work of Ms. Sadia Mujtaba.
Author information
Authors and Affiliations
Contributions
MTJ, MS and MSA, planning of study; KT and SM, experimental work; SKT and MHS, statistical analysis and critically review; HJC and SM, data discussion. All the authors approved the final version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Shahid, M., Javed, M.T., Tanwir, K. et al. Plant growth-promoting Bacillus sp. strain SDA-4 confers Cd tolerance by physio-biochemical improvements, better nutrient acquisition and diminished Cd uptake in Spinacia oleracea L.. Physiol Mol Biol Plants 26, 2417–2433 (2020). https://doi.org/10.1007/s12298-020-00900-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12298-020-00900-4