
Abstract
Conventional approaches to natural product drug discovery rely mainly on random searches for bioactive compounds using bioassays. These traditional approaches do not incorporate a chemical biology perspective. Searching for bioactive molecules using a chemical and biological rationale constitutes a powerful search paradigm. Here, the authors review recent examples of the discovery of bioactive natural products based on chemical and biological interactions between hosts and symbionts, and propose this method provides a more effective means of exploring natural chemical diversity and eventually of discovering new drugs.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The advent of the antibiotic era, initiated by Alexander Fleming’s discovery of penicillin from the fungal genus Penicillium (Fleming 1929), represented a huge breakthrough in human healthcare against recalcitrant diseases. Based on Fleming’s success, investigations of microorganisms targeting antibiotic natural products resulted in many clinically useful antibiotics, such as, cephalosporins, aminoglycosides, tetracyclines, and erythromycin and its related macrolides, vancomycin, and amphotericin, during the period 1940–1960 (Walsh 2003a). However, over the past three decades, there has been a general decline in the discovery of novel natural products with useful pharmaceutical properties, and at the same time drug resistant diseases became far more widespread (Wright 2007). For example, history confirms that whenever a new antibiotic is utilized widely, clinically significant resistance follows (Walsh 2003b), though the time taken for this to occur varies considerably (e.g., 2 years for penicillin or 30 years for vancomycin) (Plumbi 2001). The same is true of antifungal drug therapies. Amphotericin B provides an excellent example, it was introduced as a clinical treatment in 1950s and Candida resistant isolates were observed almost simultaneously (Littman et al. 1959). Given that life-threatening diseases are becoming more common (Morens et al. 2004), currently available drugs have limitations and the platforms used for the discovery of synthetic therapeutic agents are not as effective as they might be (Ortholand and Ganesan 2004). Accordingly, the discovery of novel classes of drugs from natural sources is a high priority. The drop in the discovery rate of novel natural products that occurred in the late 1980s caused major pharmaceutical companies to change their research paradigms from natural products to combinatorial chemistry. However, it is becoming increasingly clear that combinatorial synthesis, based on previously described platforms, does not provide an effective route to new drugs. On the other hand, the importance of natural products is demonstrated by the facts that more that 60 % of antibacterial and antifungal drugs approved by the FDA between 1981 and 2010 are natural products or natural product-derived molecules. Furthermore, most recently approved antifungal (anidulafungin) and antibacterial drugs (daptomycin) originated from natural products (Newman and Cragg 2012).
Conventional natural product discovery strategies that are dependent on random searches for bioactive compounds may not meet our current needs. Ecologically based search techniques offer a means of improving search efficiencies. However, although we understand the clinical functions of natural compounds in some detail, comparatively little is known about the real natural roles of secondary metabolites in microorganisms (Davies and Ryan 2012). This lack of knowledge has limited the search for natural products to readily accessible environments. Accordingly, there is a need to develop distinctly different strategies for the discovery and characterization of new natural products with clinical potential. Here, we review representative examples of microbial bioactive product discovery from the perspective of chemical biology, in the hope that this study aids the development of new search strategies for bioactive small molecules for drug discovery purposes.
Chemical biology of insect-associated microbes and their bioactive compounds
New antibiotics of symbiotic bacteria mediate multipartite anti-microbe symbiosis
Mutualistic relationships between microorganisms and eukaryotic hosts can be mediated by microbial secondary metabolites, especially by those with antibiotic potential, and these interactions are more widely distributed than previously appreciated. One of the best known examples is the multipartite interactions in the fungus-growing ant (attine ant) systems (Chapela et al. 1994; Currie et al. 1999). Fungus-growing ants (Attini: Formicidae) cultivate a fungal food source by feeding plant material to the fungus. An antagonistic fungus, Escovopsis sp., tries to invade the fungal garden and consume the fungal food source (Currie et al. 2003). It has been demonstrated that secondary metabolites with antifungal properties, produced by a symbiotic actinomycete (Pseudonocardia sp.), control the parasitic fungi and help maintain the beneficial food fungal culture.
The mutualism exhibited by fungus-growing ants and their food source has been well studied from the late 1800s and today the symbiosis shown by these ants provides a good model for studies of symbiosis (Muller et al. 1998; Chapela et al. 1994). Phylogenetic analysis supports that attine ants have cultivated fungi for over 50 million years (Currie et al. 2003; Chapela et al. 1994). How the ants protect their fungal gardens from Escovopsis spp. was longstanding question, but relatively recently it was determined that this successful mutualism is supported by another symbiont, the antibiotic-producing actinomycete, Pseudonocardia. In 1999, Currie and co-workers discovered this previously-unknown association between the attine ants and symbiotic actinomycetes, which secrete putative antifungal chemical substance to inhibit parasitic fungi (Currie et al. 1999). The ants were found to support these mutualistic bacteria with coevolved crypts and exocrine glands (Currie et al. 2006). However, the molecules mediating this interesting symbiotic system have not been identified.
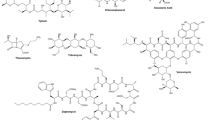
Further chemical and biological investigation of one ant species (Apterostigma dentigerum) based on through comprehensive chromatographic and spectroscopic analyses, led to the isolation of a major secondary metabolite, dentigerumycin (1), from the symbiotic Pseudonocardia strain (Fig. 1). An antifungal bioassay against the specialized parasitic fungus Escovopsis strain confirmed that this molecule clearly inhibits the growth of this parasitic fungus. Spectroscopic analysis of this dentigerumycin revealed that it is structurally novel and bears unusual amino acid units, that is, three piperazic acids, β-hydroxy leucine, N-hydroxy alanine, and a polyketide-derived moiety with a pyran ring (Oh et al. 2009a). The identification of dentigerumycin in symbiotic Pseudonocardia sp. and its inhibition of parasitic fungus demonstrates that the fungus-growing ants coevolved with symbiotic actinomycetes by utilizing their secondary metabolites to protect their food source fungus and maintain the health of the entire symbiotic system. This result also demonstrates that the chemical biology of coevolved chemical interactions between insects and microbes provide valuable insights regarding the discovery of novel natural antibiotics (Hayden 2009). Given that specific groups of fungus-growing ants utilize specific fungal species, these fungi are hosts to specific species of Escovopsis (Currie et al. 2003), and that these associations are modulated by specific Pseudonocardia, it is evident that more new antibiotics could be discovered by studying more symbiotic Pseudonocardia from phylogenetically similar ants.

Microbial bioactive natural products playing key roles in insect–microorganism symbiotic systems in chemical biology perspectives
Chemical biology of the multipartite symbiosis of beetle, bacteria, and fungi in the southern pine beetle ecosystem
The southern pine beetle, Dendroctonus frontalis, is a pest that damages conifers in the southern United States (Pechanova et al. 2008), and engages in one of the most complex symbiotic associations described in nature involving trees, fungi, mites (Klepzig and Wilkens 1997; Klepzig et al. 2001) and bacteria. They typically associate with a beneficial fungus, Entomocorticium sp. by harboring the fungus within specialized body structures (mycangia) of female beetles (Six and Klepzig 2004). Southern pine beetle reproductive adults bore holes through outer bark to create ovipositional galleries within the inner bark and phloem (Six and Klepzig 2004). During the process of gallery construction, female beetles inoculate the phloem and xylem of the tree with their mutualistic fungus, Entomocorticium sp. (Bramble and Holst 1940). Developing larvae obtain nutritional benefits by consuming this fungus. Another fungal symbiont, Ophiostoma minus, has a context dependent relationship with the beetle, and can out-compete the beneficial fungus for uncolonized substrate and thereby disrupt larval development. On the other hand, it can also assist beetles by killing healthy trees (Hofstetter et al. 2006). Furthermore, O. minus has a mutualistic relationship with phoretic tarsonemid mites, which are transported from tree to tree on the exoskeletons of southern pine beetles. The mites feed on O. minus and in exchange the fungus is vectored to new host trees (Klepzig et al. 2001). It was a topic of considerable interest how the beetle controls the antagonistic fungus and protects the beneficial mycangial fungus, because when cultured together O. minus out-competes Entomocorticium sp. (Klepzig and Wilkens 1997). Providing an answer to this chemical biologic question provided crucial clues regarding the discovery of novel antibiotic compounds. The Currie laboratory successfully isolated two previously uncharacterized Streptomyces strains (SPB74 and SPB78) from southern pine beetles in Mississippi and localized one (SPB74) of the two morphotypes to mycangia. Furthermore, Streptomyces strain SPB74 isolated from mycangia inhibited O. minus significantly more than the beneficial Entomocorticium sp. Chemical study of the secondary metabolites of SPB74 revealed that this symbiont produced a novel polyene peroxide, mycangimycin (2) (Scott et al. 2008; Oh et al. 2009b), which is a novel linear 20 carbon carboxylic acid with an endoperoxide and a conjugated heptaene moiety (Fig. 1). The selective toxicity of mycangimycin towards the fungal antagonist, combined with the prevalence and localization of its bacterial source, strongly indicates mycangimycin is the key antifungal that mediates the southern pine beetle symbiotic system (Fig. 2). Mycangimycin has also been reported to be active against a therapeutically relevant amphotericin B resistant Candida albicans strain (Oh et al. 2009b).

The chemical and biological interactions in the southern pine beetle symbiotic system. Southern pine beetle Dendroctonus frontalis utilizes the antifungal natural product, mycangimycin, produced by the symbiotic Streptomyces sp. SPB74 in controlling the antagonistic fungus Ophiostoma minus and thus protecting the larval food fungus Entomocorticium
Continued chemical and biological studies of other Streptomyces strain in the southern pine beetle ecosystem, SPB78, also resulted in the discovery of new antifungal compounds, namely frontalamide A (3) and B, which are polycyclic tetramate macrolactams (Fig. 1) (Blodgett et al. 2010). These antifungals have been proposed to deform hyphae of the antagonistic fungus O. minus. Furthermore, the discovery of the frontalamides and genomic analysis of strain SPB78 also led to the identification their biosynthetic pathways. Once the biosynthetic gene cluster had been identified, several actinomycetes from various environments were screened for frontalamide-like compounds. A degenerate primer-based PCR screen was designed to detect a highly conserved gene region in the biosynthetic pathway, and resultantly, the biosynthetic pathways of frontalamide-like compound were commonly detected among screened bacterial strains. Subsequent chemical analysis of the bacterial cultures by LC/MS detected these frontalamide-type antibiotics (Blodgett et al. 2010).
Given that host associations with actinomycetes resulting in the productions of antibiotics that mediate complex microbial symbiotic interactions are probably common, these results suggest that the identification of chemical biologic associations and the adoption of a chemical biology perspective offer an effective strategy for identifying new natural antibiotics.
Chemical biology of gut bacteria and their secondary metabolites in fungus-growing termites
Fungus-growing termites (Macrotemitinae) are known as decomposers in ecosystems and material cycles. Like the fungus-growing ants, the ecosystems of fungus-growing termites incorporate a lineage of basidiomycete Termitomyces fungi, which aid the degradation of plant material and also serve as food for the termites (Anan et al. 2002). Termites carry plant material to their nests, and this is then partially-digested and accumulated in fungal cultivar combs by colony workers (Rould-Lefevre 2002). Because potentially antagonistic and/or competitive fungi, such as, Pseudoxylaria and Trichoderma, can invade monocultured Termitomyces fungus combs, the success of this mutualistic food-to-shelter symbiosis is likely to be dependent on the effectiveness of the mechanism that protects cultivar fungi (Katoh et al. 2002). It has been documented that fungus-growing termites control inflow materials to their nests by passing the material through the gut, and thus, gut microbiota could play pivotal roles in chemical and biological interactions in this ecosystem. An investigation of the microbiota of the fungus-growing termite Odontotermes formosanus showed that Bacillus sp. present in the gut and in fungus combs secretes a chemical substance that selectively inhibits Trichoderma but not Termitomyces (Katoh et al. 2002). However, unfortunately, the structure of this inhibitory substance has not been deciphered.
In a chemical biologic study conducted on another fungus-growing termite species, Macrotermes natalensis, obtained from South Africa, Poulsen and co-workers demonstrated Bacillus strains inhabiting in the termite gut produce a single major compound that selectively inhibits competitive fungi, such as, Pseudoxylaria and Trichoderma, and the potentially antagonistic fungi Coriolopsis, Umbelopsis, and Fusarium (Um et al. 2013a). Chemical identification of the major metabolite by bioassay-guided purification and spectroscopic analysis revealed that the selective antifungal was bacillaene (4) (Fig. 1). In addition, the symbiotic Bacillus strains were isolated from different M. natalensis colonies and compared with respect to the 80 kb biosynthetic gene cluster of bacillaene and whole genomes. The Bacillus strains were found to have nearly identical genomes, which included biosynthetic gene clusters with almost 100 % identity at the nucleotide and amino acid levels in 15 of 16 genes. This result indicated that fungus-growing termites utilize gut-associated Bacillus symbionts and the chemical compound they produce to maintain the fungal cultivar by suppressing competitive/antagonistic fungi (Um et al. 2013a). In this chemical biology study, the antifungal function of bacillaene, which had been previously shown to be an antibacterial that inhibits prokaryotic protein synthesis, was evaluated for the first time.
European beewolf utilizes a chemical biologic mechanism based on a symbiotic bacterial antibiotic combination to protect offspring
Some wasps construct nest burrows in soil and the larva then feeds on prey that females hunt and provide (Strohm and Linsenmair 1995). Because brood cells are generally humid and warm, immature wasps are prone to fungal and bacterial infections (Strohm and Linsenmair 1995). In the past, it suspected that wasps had probably developed a measure to protect their larva, but its nature was unknown. In 2005, it was reported that one of the solitary hunting wasps, Philanthus triangulum (Hymenoptera), has a unique association with an actinomycete, which presumably protects the cocoon from fungal infestation and significantly enhances larval survival (Kaltenpoth et al. 2005). The endosymbiotic actinomycete strain was detected predominantly in a specialized antennal gland reservoir of female beewolves. Further microbiological studies revealed that this symbiotic bacterium is a new species, Candidatus Streptomyces philanthi (Kaltenpoth et al. 2006). Chemical and biological investigation of this system revealed that the wasp cultivates this specific symbiont and incorporates the bacterium into larval cocoons. Kaltenpoth and co-workers elucidated the underlying chemical biologic mechanism involved. Comprehensive analysis of cocoon extracts by LC/MS/MS and NMR spectroscopy identified piericidin derivatives (5) and streptochlorin (6) as components of the protective antibiotics (Fig. 1) (Kroiss et al. 2010). These antibiotics strongly inhibited various entomopathogenic microbes including Penicillium avellaneum and Paenibacillus larvae, demonstrating that the cocktail of antibiotics produced by a specific actinomycete symbiont constitutes a primary molecular defense mechanism in this system. The distributions of the identified antibiotics on beewolf cocoons were subsequently analyzed by laser desorption/ionization (LDI)–time of flight (TOF) mass spectrometry, and it was concluded that this chemical biologic system utilizes symbiont-produced antibiotics to defend wasp larvae against pathogens, which parallels the ‘combination prophylaxis’ of human medicine (Kroiss et al. 2010).
The continuous search for bioactive natural products in the chemical biologic context of insect–microbe symbiosis
Although complete chemical biology studies of insect–microbe symbiotic systems are not always feasible, such strategies might enable us to tap the huge chemical diversity presented by insect-associated microorganisms for drug discovery (Fig. 3). In 1950s, an Italian research group discovered a toxic compound, pederin, in the hemolymph of the rove beetle Paederus fuscipes (7) (Pavan and Bo 1953). In this study, structural determination was enabled by collecting 250,000 beetles, but later the true producer of pederin was identified by molecular biological analysis of the biosynthetic gene cluster of pederin to be an endosymbiotic Pseudomonas inside female beetles (Piel 2002). To date, this endosymbiotic bacterium has not been cultured, and thus, no further chemical biology studies have been conducted.

Microbial bioactive natural products discovered in the chemical biologic context of insect–microbe symbiosis
Investigations of insect-associated bioactive microbial compounds started in the late 2000s. In 2008, Tan and co-workers isolated the fungus Daldinia eschscholzii strain from the gut of the mantis Tenodera aridifolia; a predator of many insects feeding on endophyte-bearing plants (Zhang et al. 2008). Chemical investigations of culture extracts of this fungus resulted in the discovery of dalesconols A (8) and B, which possessed an unprecedented carbon framework (Zhang et al. 2008). The findings of other relevant investigations include: the production of new cytotoxic furanone analogues, paraconfuranones A (9)-H by a fungal strain belonging to Paraconiothyrium brasiliense isolated from the gut of the grasshopper, Acrida cinerea (Liu et al. 2015); the production of a series of new epipolythiodioxopiperazines, boydines A (10)-D, and structurally novel sesquiterpenes, boydenes A and B (11) by a symbiotic fungus Pseudallescheria boydii, in the gut of the coleopteran larva of the beetle Holotrichia parallela (boydine B displayed significant inhibitory activity against clinically-relevant bacterial strains Bifidobacterium sp., Veillonella parvula, Anaerosterptococcus sp., Bacteroides vulgatus and Peptostreptococcus sp.) (Wu et al. 2014); and the production of a new polyketide glycoside, bionectriol A (12) by a fungal strain belonging to Bionectra sp. associated with the fungus-growing ant (Apterostigma dentigerum) (Freinkman et al. 2009).
Chemical and biological studies of insect-associated bacteria continuously lead to the discovery of bioactive natural products. In 2011, a new 26-membered macrocyclic lactam, sceliphrolactam (13), was reported from a Streptomyces sp. strain associated with the mud dauber wasp, Sceliphron caementarium (Oh et al. 2011). However, this compound is extremely unstable, and thus, its absolute configuration has not been determined. Subsequently, the antifungal activity of sceliphrolactam was confirmed against amphotericin B-resistant Candida albicans (Oh et al. 2011). The above-mentioned studies originated from the chemical analyses of wasp-associated Streptomyces bacteria. Over 200 Streptomyces strains were selectively isolated from solitary mud dauber wasps, S. camentarium and Chalybion californicum (Poulsen et al. 2011). Many bioactive compounds including sceliphrolactam were identified in associated Streptomyces strains, demonstrating that actinobacterial symbionts in Hymenoptera could offer a rich source of bioactive compounds (Poulsen et al. 2011).
New acrylamide (14) and oxazolidin (15) derivatives were isolated from a Streptomyces sp. residing in the gut of the termite Odontotermes formosanus (Bi et al. 2011), and polyketide synthase–nonribosomal peptide synthetase hybrid natural products, macrotermolides A (16) and B, have been discovered from a termite-associated Streptomyces sp. (Carr et al. 2012) Furthermore, continuous research on the secondary metabolites of termite-associated actinomycetes from Macrotermes natalensis colonies led to the isolation of a highly modified geldanamycin compound, natalamycin A (17) (Kim et al. 2014), and the antifungal compounds, tyroscherin and N-methyltyroscherin (18), were identified in a Pseudallescheria boydii fungal strain isolated from the termite Nasutitermes sp. in France (Nirma et al. 2013). In addition, rare actinomycete Amycolatopsis from the grasshopper Oxya chinensis was found to produce new tetrasaccharides, that is, the actinotetraoses A (19)-K (Guo et al. 2012, 2013).
Dung beetle ecosystems are considered interesting in the natural product chemical biology research field. These beetles utilize feces to make brood balls and lay a single egg inside each ball. Because larvae grow inside brood balls and feed on feces, one would imagine that they are potentially vulnerable to pathogens and probably protected by some unknown mechanism. Accordingly, actinomycetes associated with the dung beetle, Copris tripartitus, have been studied in an effort to identify new bioactive natural products. Neuroprotective phenylpyridines, coprismycins A (20) and B, were discovered from in a Streptomyces sp. found in the dung beetle ecosystem (Kim et al. 2011). This class of phenylpyridines constituted a new chemotype and displayed neuroprotective effects against MPP+ (1-methyl-4-phenylpyrimidium)-induced neurotoxicity in neuroblastoma SH-SY5Y cells (Kim et al. 2011). In addition, a brood ball-associated Streptomyces strain yielded the structurally unprecedented tricyclic compound, tripartilactam (21), which incorporates a cyclobutane (Park et al. 2012). Although, its structural instability hampered comprehensive biological evaluation, tripartilactam was found to inhibit Na+/K+ ATPase (Park et al. 2012). Dung beetle larva also harbors diverse bacteria, which could produce bioactive substances. An investigation of larva-associated bacteria in the dung beetle C. tripartitus resulted in the discovery of a dichlorinated indanone, tripartin (22) (Kim et al. 2013), which was found to selectively inhibit histone H3 lysine 9 demethylase KDM4 in HeLa cells, suggesting its use as a chemical probe lead (Kim et al. 2013). In addition, an Amycolatopsis sp. found in a female dung beetle (C. tripartitus) produced the novel 3-furanone-bearing polyketide, actinofuranone C (23) (Um et al. 2013b). As discussed for fungus growing termites above, the gut microfloras of insects exhibit significant chemical biologies, and a recent study of bioactive compounds from bacteria in the gut of the dung beetle C. tripartitus led to discovery of the novel cyclic heptapeptides coprisamides A (24) and B (Um et al. 2015). These unusual compounds incorporate modified amino acid units, such as, β-methylaspartic acid and 2,3-diaminopropanoic acid covalently linked to valine. In addition, the coprisamides contain a previously-unreported 2-hepatrienyl cinnamic acid acyl chain unit. Coprisamides A and B significantly induce quinone reductase activity, and thus are of interest as potential chemopreventives.
Chemical biologies of microbial bioactive secondary metabolites in marine invertebrate ecosystems
Crustacean–episymbiotic bacterial chemical interactions
One of the early examples cited regarding the chemical and biological functions of microbial natural products was a shrimp–microbe chemical interaction, whereby symbiotic marine bacteria defend crustacean embryos from a pathogenic fungus using a bacterial antibiotic (Gil-Turnes et al. 1989). It was previously documented that the pathogenic fungus Lagenidium callinectes infects juveniles and larvae of the shrimp Palaemon macrodactylus. However, interestingly, externally-brooded embryos of this shrimp are resistant to this fungus. Symbiotic bacteria on the surfaces of healthy P. macrodactylus embryos were investigated to decipher the resistance mechanism, and fortunately, a culture-dependent approach resulted in the isolation of several bacterial strains (the culture-independent genomic approach was not available at the time). One strain I-2, Alteromonas sp., was found to be sensitive to penicillin and to inhibit effectively the proliferation of the pathogenic fungus L. callinectes. Fenical and co-workers found that this bacterial strain produced an antifungal in large quantity, and subsequent isolation and structural determination resulted in the identification of 2,3-indolinedione, isatin. In order to investigate the chemical biology of isatin, a series of experiments were performed. Briefly, four groups of embryos were prepared. The first group was used as a control. After eliminating symbiotic bacteria from embryos in second, third and fourth groups using penicillin-G, embryos in the second group received no further treatment; embryos in the third group were re-exposed to the Alteromonas strain; and embryos in the fourth group were dipped periodically in isatin solutions. These four groups were then infected with the pathogenic fungus L. callinectes. The results obtained demonstrated that the embryos treated with only penicillin (the second group) had a dramatically lower survival rate than controls, whereas embryos in the Alteromonas-reinfected group (the third group) and in the isatin supplemented group (the fourth group) did not. In addition, the pathogenic fungus was observed to colonize the surfaces of embryos treated with penicillin-G. These results firmly support the notion that the epi-symbiotic bacterial strain Alteromonas sp. produces the antifungal agent, isatin, and that this protects crustacean embryos from the pathogenic fungus (Gil-Turnes et al. 1989). This study provides a seminal example of the chemical biology of natural products.
Investigation of potential bioactive compounds involved in coral–microbe chemical biology
Corals are marine invertebrates of the phylum Cnidaria and are a rich source of bioactive mevalonate pathway derived secondary metabolites, such as, terpenoids and steroids, some of which are used for chemical defense (Fenical and Pawlik 1991). Ecologically, corals are habitats for photosynthetic dinoflagellates and other diverse microbes that can contribute to the host by providing nutrients and recycling or assimilating host-producing ammonia (Burris 1983; Gates and Ainsworth 2011). In addition to nutrient supply and waste control, the symbiotic microbial communities on corals produce chemicals for host defense against coral pathogens and predators (Ritchie 2006).
Pseudopterosin (25) and its analogs are tricyclic diterpene glycosides that were originally isolated from the Caribbean soft coral Pseudopterogorgia elisabethae and have been reported to have potent anti-inflammatory and analgesic activities (Fig. 4) (Look et al. 1986a, b; Fenical 1987). After differential centrifugation, Mydlarz et al. found pseudopterosins in fractions enriched with the genus Symbiodinium, a symbiotic dinoflagellate of P. elisabethae (Mydlarz et al. 2003). Furthermore, incubation of the Symbiodinium enriched fraction with isotope labeled precursors resulted in the isolation of labeled pseudopterosins (Mydlarz et al. 2003), suggesting symbiotic dinoflagellates of P. elisabethae produced the pseudopterosin. Currently, pseudopterosins are used as anti-inflammatory cosmeuticals in skin care creams (Mayer et al. 1998), which provides an example of the commercial usage of microbial natural products.

Bioactive natural products discovered based on chemical biology of the coral–bacterium association (25), the coral–cyanobacterium association (26–28) and the bryozoan–bacterium association (29–34)
In 2008, a phenomenal microbial bloom was reported by Smith and co-workers on Hawaiian coral reefs (Smith et al. 2008). The bloom-forming microbe was identified as Leptolyngbya crossbyana a filamentous marine cyanobacterium, which had overgrown on the reef coral, Porites compressa, in the Honaunau bay area. Smith and co-workers found dead coral branches and coral bleaching around cyanobacterial colonies, but without direct contact between the two, and speculated the invading cyanobacterium produced toxins. The Gerwick group performed a chemical investigation of this bloom-forming cyanobacterium and two different chemotypes of bioactive natural products were subsequently identified by Choi and co-workers (Choi et al. 2010, 2012). The novel chemical entities of the first chemotype were crossbyanols A–D (heptabrominated pentaphenyl ethers showing sulphate group variations). Honaucins possessing 4-chlorocrotonic acid with a 3S-hydroxy butyrolactone or 3,4-dihydroxy butanoate methyl or ethyl esters composed the second type. In general, the surfaces of marine objects are covered with microbial layers (biofilms) and the corals are known to be well-defended by symbiotic microbe derived chemicals. The two chemotypes isolated from L. crossbyana have been tested using bioassay platforms related to the defense mechanisms of corals, such as, anti-microbial activity, cytotoxicity, quorum sensing inhibition, and anti-inflammatory activities. Of these compounds, crossbyanol B (26) showed potent antimicrobial activity against methicillin resistant Staphylococcus aureus (MRSA) and toxicity on brine shrimp (Fig. 4) (Choi et al. 2010). Interestingly, the honaucins showed potent anti-inflammatory activities on lipopolysaccharide-stimulated murine macrophages and inhibited quorum sensing signaling by Vibrio harveyi BB120 and Escherichia coli JB525 (Choi et al. 2012). In addition, Choi and co-workers prepared a small panel of synthetic derivatives of honaucin A (27) and investigated structure–activity relationships. They found a synergistic effect between a halogenated α,β unsaturated carboxylic acid and a 3S-hydroxy butyrolactone; however, most of the synthetic analogs produced, other than bromohonaucin A (28), showed no improvement in bioactivity or chemical stability (Fig. 4). As a result, they proposed the honaucins are the result of natural evolution.
Chemical biology of the bryozoan Bugula neritina and its associated bacterial metabolites
Bryozoans are sessile filter feeders and a known source of potent cytotoxins. In 1968, a chemical investigation on the extracts of marine organisms collected in Gulf of Mexico was initiated for the purpose of anticancer drug discovery (Halford 2011). Of the extracts of randomly collected marine organisms, the extract prepared from a Bryozoan, Bugula neritina, was discovered to have potent anticancer activities by the National Cancer Institute (NCI). Fourteen years later, Pettit and co-workers elucidated the structure of the compound responsible for toxicity in an extract of B. neritina (Pettit et al. 1982). The compound, which was named “bryostatin 1 (29)”, possesses a 26-membered macrolactone ring incorporating three pyran rings (Fig. 4). To date, 20 different bryostatins have been identified mainly from B. neritina (bryostatin 8 was isolated from Amanthia convolute).
Bryostatin 1 was first found during an anticancer drug discovery program and follow-up study revealed that the bryostatins are potent protein kinase C (PKC) modulators and that this activity is due to binding to the diacylglycerol binding site of the PKC regulatory domain (Nelson and Alkon 2009). In vitro, bryostain 1 stimulates cell differentiation and apoptosis and inhibits cell growth and angiogenesis (Goekjian and Jirousek 2001). Because of its potent activities in in vitro and in animal models, the anticancer drug potential of bryostatin 1 was thoroughly investigated. However, phase II trials were discontinued due to low effectiveness and severe side effects, which included myalgia (Propper et al. 1998). Bryostatins were then investigated in combination therapies with other chemotherapeutics, but unfortunately, the results obtained did not justify further investigation (Kollá et al. 2014). More recently, bryostatin 1 was found to have potential as a treatment for Alzheimer’s disease, based on findings of reduced amyloid β production and protection from neurological injury after acute cerebral ischemia in animal models (Etcheberrigaray et al. 2004; Tan et al. 2013). Currently bryostatin 1 is undergoing phase II clinical trials for the treatment of Alzheimer’s disease (NCT02431468). Bryostatins have also been reported to be potential anti-HIV agents as they induce the activation of latent HIV infected cells and facilitate their elimination (Mehla et al. 2010; DeChristopher et al. 2012).
The chemical biological function of bryostatins in nature was studied by the Haygood and Lindquist groups (Lopanik et al. 2006; Trindade-Silva et al. 2010). Lopanik et al. found bryostatins, particularly bryostatins 10 (30) and 20 (31) (Fig. 4), were used as chemical defense agents to protect B. neritina against fish, corals, and sea anemones, and interestingly, that bryostatin concentrations vary throughout the life-cycle and the levels of these two bryostatins are much lower in the adult than in larvae (Lopanik et al. 2006). It has been speculated that the bryostatins are used by B. neritina larvae to deter predators.
Of the biologically active marine natural products, the bryostatins are chemically and biologically well studied entities. The Pettit group recognized the presence of bryostatin 1 in B. neritina collected in California coast, and isolated 120 mg of bryostatin 1 from 500 kg of the animal (Pettit et al. 1982). In 1991, the NCI undertook a massive collection of B. neritina (~10,000 gallons of wet samples) and successfully isolated 18 g of bryostatin 1 (Schaufelberger et al. 1991), which is sufficient to perform clinical trials.
Scientists have been trying to develop an alternative ways to obtain bryostatin 1 consistently without impacting the environment, and two independent approaches based microbiology and organic chemistry, have been developed.
In the microbiology field, there remains an issue regarding the real producer of the bryostatins. An association between rod-shaped bacteria and B. neritina has been consistently observed through the lifecycle of B. neritina (Woollacott and Zimmer 1975; Woollacott 1981). In particular, the larvae of B. neritina have been found to harbor specific bacteria in the pallial sinus (Woollacott and Zimmer 1975). Later, these associated bacteria were identified as Candidatus Endobugula sertula by molecular biological study (Haygood and Davidson 1997). Based on the chemical structure and the consistent presence of associated bacteria, bryostatins were speculated to be of bacterial origin as shown in Fig. 5. Antibiotic treatment resulted in a decrease in both Ca. E. sertula population and bryostatin concentration in adult B. neritina (Davidson et al. 2001). Furthermore, the larvae of antibiotic-treated B. neritina colonies showed significant reductions in Ca. E. sertula populations and bryostatin concentrations, and larvae free of the symbiont were predated by fish (Lopanik et al. 2004). For some time, microbiologists have tried to isolate this symbiotic bacterium and confirm it produces bryostatin, but as yet, no one has been able to cultivate the bacterium.

The chemical biology in the bryozoan–bacterium association. The bryozoan Bugula neritina uses the bryostatins, produced by associated bacterium Candidatus Endobugula sertula, to protect the larvae from predators. The bryostatins have been found to have therapeutic potential to treat cancers, Alzheimer’s disease and HIV
On the other hand, feeding experiments on B. neritina using isotope labeled precursors established that bryostatins are biosynthesized by a polyketide synthase (PKS) pathway, which is commonly used in the bacteria (Kerr et al. 1996). As a result, a metagenomic approach was applied to B. neritina and PKS genes were rapidly identified in the metagenome samples containing Ca. E. sertula (Davidson et al. 2001), and in addition, a putative biosynthetic gene cluster for bryostatin (bry) was identified (Hildebrand et al. 2004). The bry gene cluster is expressed in Ca. E. sertula cells, but the gene is not expressed after antibiotic treatment (Davidson et al. 2001). The heterologous expression of fragments of bry gene clusters confirmed the use of bry for the biosynthesis of bryostatins (Lopanik et al. 2008; Buchholz et al. 2010). However, the heterologous production of bryostatins has not been achieved as yet.
The structural complexity of the bryostatins prevented their chemical synthesis until 2008. Trost and Dong reported the first synthesis of bryostatin 16 (32) in 2008 (Trost and Dong 2008), and in 2010, the Keck group successfully synthesized bryostatin 1 (Keck et al. 2011). Later in 2011, the Wender and Krische groups successfully synthesized bryostatin 9 (33) and bryostatin 7 (34), respectively (Fig. 4) (Wender and Schrier 2011; Lu et al. 2011).
Advances in the chemical biology of ascidian-associated cyanobacterial secondary metabolites
The ascidians (also known as sea squirts) are sessile filter feeders in the marine ecosystem, and are known to accumulate diverse bioactive compounds. The colonial ascidians (Didemnidae) are known to contain diverse cyanobactins, which are highly modified cyclic peptides. The first two cyanobactins, ulicyclamide (35) and ulithiacyclamide A (36) (Fig. 6), were isolated in 1980 from the extract of Indo-Pacific Lissoclinum patella (Didemnidae) (Ireland and Scheuer 1980), and to date, over 30 cyanobactins, such as, patellamides, ascidiacyclamides, lissoclinamides, ulicyclamides, tawicyclamides, preulithacyclamides, and the patellins, have been reported; most of which were identified in the colonial ascidian, L. patella. Many cyanobactins are potently cytotoxic to various cancer cell lines (IC50 values as low as 0.1 μg/mL), and thus, the NCI investigated the anti-tumor activities of ulithiacyclamide A in two mouse models (Sarcoma M5076 in BCF1 mice and Leukemia P388 in CD2F1 mice) (more detailed information is available on the NCI web site (http://dtp.nci.nih.gov)). However, drug development based on ulithiacyclamide A was terminated due to its high toxicity. A chemical ecological study showed that the other cyanobactin, patellamide C (37) (Fig. 6), deters predators in the marine environment, which suggests patellamide C is a chemical defense principle of L. patella (Paul 1992).

Representative cyanobactins (35–40) discovered from the association of marine ascidian and cyanobacterium
The marine ascidians also harbor a consortium of microorganisms. The symbiotic relationship between the Didemnid ascidians and the cyanobacteria Prochloron sp. (formerly known as photosynthetic algae) was first reported in the mid 1970s (Lewin and Withers 1975; Withers et al. 1978). Prochloron is a photosynthetic unicellular cyanobacterium found embedded in the cavities between the two tunic layers of host ascidians and provides photosynthetic products to the host (Withers et al. 1978; Kühl and Larkum 2002; Hirose et al. 2006). Prochloron sp. are also found in the other marine invertebrates with diverse symbiotic or associated microorganisms (Kühl and Larkum 2002). Interestingly, diverse cyanobactins have been observed in ascidians harboring Prochloron sp. Furthermore, most cyanobactins have been found in L. patella, which harbors Prochloron as the only one symbiont. Therefore, in the past, Prochloron sp. was considered to be the original producer of cyanobactins. Several researchers have isolated Prochloron sp. from L. patella tissues and investigated cyanobactin production, but unfortunately published results are inconsistent (Degnan et al. 1989).
The cyanobactins are all macro-cyclic peptides possessing d-amino acids and two or more heterocyclic rings, such as, thiazole, thiazoline, and methyl oxazoline. Furthermore, these structural features are widely found in microbial and cyanobacterial metabolites produced via a non-ribosomal pathway. Therefore, researchers investigated the non-ribosomal polypeptide synthetase (NRPS) pathways used to produce cyanobactins (Schmidt et al. 2004; Long et al. 2005). However, no NRPS gene clusters related to cyanobactin biosynthesis were found (Schmidt et al. 2004). Schmidt and co-workers performed complete genome sequencing, and searched for all possible linear amino acid combinations of patellamide A (38) in sequence data, and then introduced putative biosynthetic gene clusters into Escherichia coli, heterologously expressed the putative pathway, and observed patellamide A production (Fig. 6) (Schmidt et al. 2005). Jaspars and co-workers adopted a shot-gun technique to clone fragments of the genomic DNA of a cyanobactin producer to E. coli, and discovered the production of patellamide D (39) and ascidiacyclamide (40) (Fig. 6) (Long et al. 2005). These two independent approaches demonstrate Prochloron sp. is the original producer of cyanobactins. Currently, the cyanobactins are considered the products of ribosomally synthesized and post-translationally modified peptide (RiPP) (Arnison et al. 2013).
Schmidt and co-workers further compared the patellamide A producing gene clusters in the genome sequences of 46 other ascidians (Donia et al. 2006), and found highly conserved gene clusters (except for two regions in the gene clusters encoding prepropeptides and the amino acid sequences of variable regions) that exactly corresponded to the amino acid sequence observed in mature peptides before post-translational modification. In addition, they observed conserved patterns in other species.
In this conserved gene clusters, the Schmidt group analyzed the function of each conserved region of three recognition sequences (RSs) for heterocyclase (RSI), N-terminal protease (RSII), and C-terminal protease/macrocyclase (RSIII). Then, they verified that the activities of cyanobactin producing gene clusters are highly tolerant over an extremely high level of mutations on variable sequences (Sardar et al. 2015). The authors concluded that L. patella achieved chemical defense by harboring symbionts and rapidly upgraded the defense by substrate evolution with hypervariable gene cassettes. More recently, the use of a chemical biological approach to biosynthetic gene sequence for patellamide led to combinatorial RiPP library construction by simple mutation of the RiPP sequence (Ruffner et al. 2015).
The above provides an exemplary example of the application of the chemical biology of natural products, as it led to novel research techniques and to the utilization of natural product producing machinery beyond the utilization of bioactive natural products.
Conclusion
Natural products have played key roles in drug discovery and development by providing new chemotypes and new targets. However, conventional approaches to identify and characterize bioactive natural products have been dominated either by efforts that seek activities in specified diseases or targets or simply by the chemical identification of natural products without any knowledge of their biological functions. As reviewed here, microorganisms synthesize and secrete compounds of pharmaceutical potential that affect the physiologies of other neighboring microorganisms or symbiotic hosts, as exemplified by chemical biologic studies of insect–microorganism and marine invertebrate–microorganism symbiotic systems. Continued efforts to identify novel bioactive natural products using natural chemical biologic perspectives that place emphasis on the natural functions of microbial secondary metabolites will undoubtedly lead to the discovery of novel chemotypes and biological activities of potential utility for drug discovery and development.
References
Anan, D.K., P. Egletton, C. Rouland-Lefèvre, T. Guldberg-Frǿslev, S. Rosendahl, and J.J. Boomsma. 2002. The evolution of fungus-growing termites and their mutualistic fungal symbionts. Proceedings of the National Academy of Sciences of the United States of America 99: 14887–14892.
Arnison, P.G., M.J. Bibb, G. Bierbaum, A.A. Bowers, T.S. Bugni, G. Bulaj, J.A. Camarero, D.J. Campopiano, G.L. Challis, J. Clardy, P.D. Cotter, D.J. Craik, M. Dawson, E. Dittmann, S. Donadio, P.C. Dorrestein, K.D. Entian, M.A. Fischbach, J.S. Garavelli, U. Göransson, C.W. Gruber, D.H. Haft, T.K. Hemscheidt, C. Hertweck, C. Hill, A.R. Horswill, M. Jaspars, W.L. Kelly, J.P. Klinman, O.P. Kuipers, A.J. Link, W. Liu, M.A. Marahiel, D.A. Mitchell, G.N. Moll, B.S. Moore, R. Müller, S.K. Nair, I.F. Nes, G.E. Norris, B.M. Olivera, H. Onaka, M.L. Patchett, J. Piel, M.J.T. Reaney, R.P. Ross, H.G. Sahl, E.W. Schmidt, M.E. Selsted, Shen.B. Severinov, K. Sivonen, L. Smith, T.H. Stein, R.D. Süssmuth, J.R. Tagg, G.L. Tang, A.W. Truman, J.C. Vederas, C.T. Walsh, J.D. Walton, S.C. Wenzel, J.M. Willey, and W.A. van der Donk. 2013. Ribosomally synthesized and post-translationally modified peptide natural products: overview and recommendations for a universal nomenclature. Natural Products Reports 30: 108–160.
Bi, S.F., F. Li, Y.C. Song, R.X. Tan, and H.M. Ge. 2011. New acrylamide and oxazolidin derivatives from a termite-associated Streptomyces sp. Natural Products Communications 6: 353–355.
Blodgett, J.A.V., D.C. Oh, S. Cao, C.R. Currie, R. Kolter, and J. Clardy. 2010. Common biosynthetic origins for polycyclic tetramate macrolactams from phylogenetically diverse bacteria. Proceedings of the National Academy of Sciences of the United States of America 107: 11692–11697.
Bramble, W.C., and E.C. Holst. 1940. Fungi associated with Dendroctonus frontalis in killing shortleaf pines and their effect on conduction. Phytopathology 30: 881–899.
Buchholz, T.J., C.M. Rath, N.B. Lopanik, N.P. Gardner, K. Hakansson, and D.H. Sherman. 2010. Polyketide β-branching in bryostatin biosynthesis: identification of surrogate acetyl-ACP donors for BryR, an HMG-ACP synthase. Chemistry and Biology 17: 1092–1100.
Burris, R.H. 1983. Uptake and assimilation of 15NH4 + by a variety of corals. Marine Biology 75: 151–155.
Carr, G., M. Poulsen, J.L. Klassen, Y. Hou, T.P. Wyche, T.S. Bugni, C.R. Currie, and J. Clardy. 2012. Microtermolides A and B from termite-associated Streptomyces sp. and structural revision of vinylamycin. Organic Letters 14: 2822–2825.
Chapela, I.H., S. Rehner, T.R. Schultz, and U.G. Mueller. 1994. Evolutionary history of the symbiosis between fungus-growing ants and their fungi. Science 266: 1691–1694.
Choi, H., N. Engene, J.E. Smith, L.B. Preskitt, and W.H. Gerwick. 2010. Crossbyanols A–D, toxic brominated polyphenyl ethers from the Hawai’ian bloom-forming cyanobacterium Leptolyngbya crossbyana. Journal of Natural Products 73: 517–522.
Choi, H., S.J. Mascuch, F.A. Villa, T. Byrum, M.E. Teasdale, J.E. Smith, L.B. Preskitt, D.C. Rowley, L. Gerwick, and W.H. Gerwick. 2012. Honaucins A–C, potent inhibitors of inflammation and bacterial quorum sensing: synthetic derivatives and structure-activity relationships. Chemistry and Biology 19: 589–598.
Currie, C.R., M. Poulsen, J. Mendenhall, J.J. Boosma, and J. Billen. 2006. Coevolved crypts and exocrine glands support mutualistic bacteria in fungus-growing ants. Science 311: 81–83.
Currie, C.R., J.A. Scott, R.C. Summerbell, and D. Malloch. 1999. Fungus-growing ants use antibiotic-producing bacteria to control garden parasites. Nature 398: 701–704.
Currie, C.R., B. Wong, A.E. Stuart, T.R. Schultz, S.A. Rehner, U.G. Mueller, G.H. Sung, J. Spatafora, and N.A. Straus. 2003. Ancient tripartite coevolution in the attine ant-microbe symbiosis. Science 299: 386–388.
Davidson, S.K., S.W. Allen, G.E. Lim, C.M. Anderson, and M.G. Haygood. 2001. Evidence for the biosynthesis of bryostatins by the bacterial symbiont “Candidatus Endobugula sertula” of the bryozoan Bugula neritina. Applied and Environment Microbiology 67: 4531–4537.
Davies, J., and K.S. Ryan. 2012. Introducing the parvome: bioactive compounds in the microbial world. ACS Chemical Biology 7: 252–259.
DeChristopher, B., B. Loy, M. Marsden, A. Schrier, J. Zack, and P. Wender. 2012. Designed, synthetically accessible bryostatin analogues potently induce activation of latent HIV reservoirs in vitro. Nature Chemistry 4: 705–710.
Degnan, B.M., C.J. Hawkins, M.F. Lavin, E.J. McCaffrey, D.L. Parry, A.L. van den Brenk, and D.J. Watters. 1989. New cyclic peptides with cytotoxic activity from the ascidian Lissoclinum patella. Journal of Medicinal Chemistry 32: 1349–1354.
Donia, M.S., B.J. Hathaway, S. Sudek, M. Haygood, M.J. Rosovitz, J. Ravel, and E.W. Schmidt. 2006. Natural combinatorial peptide libraries in cyanobacterial symbionts of marine ascidians. Nature Chemical Biology 2: 729–735.
Etcheberrigaray, R., M. Tan, I. Dewachter, C. Kuiperi, I. Van der Auwera, S. Wera, L. Qiao, B. Bank, T.J. Nelson, A.P. Kozikowski, F. Van Leuven, and D.L. Alkon. 2004. Therapeutic effects of PKC activators in Alzheimer’s disease transgenic mice. Proceedings of the National Academy of Sciences of the United States of America 101: 11141–11146.
Fenical, W. 1987. Marine soft corals of the genus Pseudopterogorgia: a resource for novel anti-inflammatory diterpenoids. Journal of Natural Products 50: 1001–1008.
Fenical, W., and J.R. Pawlik. 1991. Defensive properties of secondary metabolites from the Caribbean gorgonian coral Erythropodium caribaeorum. Marine Ecology Progress Series 75: 1–8.
Fleming, A. 1929. The antibacterial action of cultures of a Penicillium, with special reference to their use in the isolation of B. influenzae. British Journal of Experimental Pathology 10: 26–236.
Freinkman, E., D.C. Oh, J.J. Scott, C.R. Currie, and J. Clardy. 2009. Bionectriol A, a polyketide glycoside from the fungus Bionectria sp. associated with the fungus-growing ant Apterostigma dentigerum. Tetrahedron Letters 50: 6834–6837.
Gates, R.D., and T.D. Ainsworth. 2011. The nature and taxonomic composition of coral symbiomes as drivers of performance limits in scleractinians corals. Journal of Experimental Marine Biology and Ecology 408: 94–101.
Gil-Turnes, M.S., M.E. Hay, and W. Fenical. 1989. Symbiotic marine bacteria chemically defend crustacean embryos from a pathogenic fungus. Science 246: 116–118.
Goekjian, P.G., and M.R. Jirousek. 2001. Protein kinase C inhibitors as novel anticancer drugs. Expert Opinion on Investigational Drugs 10: 2117–2140.
Guo, Z.K., R.H. Jiao, H.F. Dai, W.L. Mei, R.X. Tan, and H.M. Ge. 2013. Actinotetraoses I-K: tetrasaccharide metabolites produced by an insect-derived actinobacteria, Amycolatopsis sp. HCa1. Chemistry and Biodiversity 10: 296–302.
Guo, Z.K., G.F. Zhang, R.H. Jiao, Y. Shen, Q. Xu, R.X. Tan, and H.M. Ge. 2012. Actinotetraoses A-H: tetrasaccharide derivatives from a grasshopper-associated Amycolatopsis sp. HCa1. Planta Medica 78: 988–994.
Halford, B. 2011. The bryostatins’ tale. Chemical and Engineering News 89: 10–17.
Hayden, E.C. 2009. Fungus farmers show way to new drugs. Nature 458: 558–559.
Haygood, M.G., and S.K. Davidson. 1997. Small-subunit rRNA genes and in situ hybridization with oligonucleotides specific for the bacterial symbionts in the larvae of the bryozoan Bugula neritina and proposal of “Candidatus endobugula sertula”. Applied and Environment Microbiology 63: 4612–4616.
Hildebrand, M., L.E. Waggoner, H. Liu, S. Sudek, S. Allen, C. Anderson, D.H. Sherman, and M. Haygood. 2004. bryA: an unusual modular polyketide synthase gene from the uncultivated bacterial symbiont of the marine bryozoan Bugula neritina. Chemistry and Biology 11: 1543–1552.
Hirose, E., M. Hirose, and B.A. Neilan. 2006. Localization of symbiotic cyanobacteria in the colonial ascidian Trididemnum miniatum (Didemnidae, Ascidiacea). Zoological Science 23: 435–442.
Hofstetter, R.W., J.T. Cronin, K.D. Klepzig, J.C. Moser, and M.P. Ayres. 2006. Antagonisms, mutualisms and commensalisms affect outbreak dynamics of the southern pine beetle. Oecologia 147: 679–691.
Ireland, C.M., and P.J. Scheuer. 1980. Ulicyclamide and ulithiacyclamide, two new small peptides from a marine tunicate. Journal of the American Chemical Society 102: 5688–5691.
Kaltenpoth, M., W. Goettler, C. Dale, J.W. Stubblefield, G. Herzner, K. Roeser-Mueller, and E. Strohm. 2006. ‘Candidatus streptomyces philanthi’, an endosymbiotic streptomycete in the antennae of Philanthus digger wasps. International Journal of Systematic and Evolutionary Microbiology 56: 1403–1411.
Kaltenpoth, M., W. Gottler, G. Herzner, and E. Strohm. 2005. Symbiotic bacteria protect wasp larvae from fungal infestation. Current Biology 15: 475–479.
Katoh, H., T. Miura, K. Maekawa, N. Shinzato, and T. Matsumoto. 2002. Genetic variation of symbiotic fungi cultivated by the macrotermitine termite Odontotermes formosanus (Isoptera: Termitidae) in the Ryukyu Archipelago. Molecular Ecology 11: 1565–1572.
Keck, G.E., Y.M. Poudel, T.J. Cummins, A. Rudra, and J.A. Covel. 2011. Total synthesis of bryostatin 1. Journal of the American Chemical Society 133: 744–747.
Kerr, R.G., J. Lawry, and K.A. Gush. 1996. In vitro biosynthetic studies of the bryostatins, anti-cancer agents from the marine bryozoan Bugula neritina. Tetrahedron Letters 37: 8305–8308.
Kim, S.H., H. Ko, H.S. Bang, S.H. Park, D.G. Kim, H.C. Kwon, S.Y. Kim, J. Shin, and D.C. Oh. 2011. Coprismycins A and B, neuroprotective phenylpyridines from the dung beetle-associated bacterium Streptomyces sp. Bioorganic and Medicinal Chemistry Letters 21: 5715–5718.
Kim, S.H., S.H. Kwon, S.H. Park, J.K. Lee, H.S. Bang, S.J. Nam, H.C. Kwon, J. Shin, and D.C. Oh. 2013. Tripartin, a histone demethylase inhibitor from a bacterium associated with a dung beetle larva. Organic Letters 15: 1834–1837.
Kim, K.H., T.R. Ramadhar, C. Beemelmanns, S. Cao, M. Poulsen, C.R. Currie, and J. Clardy. 2014. Natalamycin A, an ansamycin from a termite-associated Streptomyces sp. Chemical Science 5: 4333–4338.
Klepzig, K.D., J.C. Moser, F.J. Lombardero, R.W. Hofstetter, and M.P. Ayres. 2001. Symbiosis and competition: complex interactions among beetles, fungi and mites. Symbiosis 30: 83–96.
Klepzig, K.D., and R.T. Wilkens. 1997. Competitive interactions among symbiotic fungi of the southern pine beetle. Applied and Environment Microbiology 63: 621–627.
Kollá, P., J. Rajchard, and Pazourek.J. Balounová. 2014. Marine natural products: bryostatins in preclinical and clinical studies. Pharmaceutical Biology 52: 237–242.
Kroiss, J., M. Kaltenpoth, B. Schneider, M.G. Schwinger, C. Hertweck, R.K. Maddula, E. Strohm, and S. Svatoš. 2010. Symbiotic streptomycetes provide antibiotic combination prophylaxis for wasp offspring. Nature Chemical Biology 6: 261–263.
Kühl, M., and A.W.D. Larkum. 2002. The microenvironment and photosynthetic performance of Prochloron sp. in symbiosis with Didemnid ascidians. In Symbiosis—mechanisms and model systems, ed. J. Seckbach, 273–290. Boston: Kluwer.
Lewin, R.A., and N.W. Withers. 1975. Extraordinary pigment composition of a prokaryotic alga. Nature 256: 735.
Littman, M.L., M.A. Pisano, and R.M. Lancaster. 1959. Induced resistance of Candida species to nystatin and amphotericin B. Antibiotics Annual 5: 981–987.
Liu, C.X., L. Wang, J.F. Chen, Z.Y. Guo, X. Tu, Z.S. Deng, and K. Zou. 2015. Paraconfuranones A–H, eight new furanone analogs from the insect-associated fungus Paraconithyrium brasiliense MZ-1. Magnetic Resonance in Chemistry 53: 317–322.
Long, P.F., W.C. Dunlap, C.N. Battershill, and M. Jaspars. 2005. Shotgun cloning and heterologous expression of the patellamide gene cluster as a strategy to achieving sustained metabolite production. ChemBioChem 6: 1760–1765.
Look, S.A., W. Fenical, R.S. Jacobs, and J. Clardy. 1986a. The pseudopterosins: anti-inflammatory and analgesic natural products from the sea whip Pseudopterogorgia elisabethae. Proceedings of the National Academy of Sciences of the United States of America 83: 6238–6240.
Look, S.A., W. Fenical, G. Matsumoto, and J. Clardy. 1986b. The pseudopterosins: a new class of antiinflammatory and analgesic diterpene pentosides from the marine sea whip Pseudopterogorgia elisabethae (Octocorallia). Journal of Organic Chemistry 51: 5140–5145.
Lopanik, N., N. Lindquist, and N. Targett. 2004. Potent cytotoxins produced by a microbial symbiont protect host larvae from predation. Oecologia 139: 131–139.
Lopanik, N.B., J.A. Shields, T.J. Buchholz, C.M. Rath, J. Hothersall, M.G. Haygood, K. Hakansson, C.M. Thomas, and D.H. Sherman. 2008. In vivo and in vitro trans-acylation by BryP, the putative bryostatin pathway acyltransferase derived from an uncultured marine symbiont. Chemistry and Biology 15: 1175–1186.
Lopanik, N.B., N.M. Targett, and N. Lindquist. 2006. Ontogeny of a symbiont-produced chemical defense in Bugula neritina (Bryozoa). Marine Ecology Progress Series 327: 183–191.
Lu, Y., S.K. Woo, and M.J. Krische. 2011. Total synthesis of bryostatin 7 via C–C bond-forming hydrogenation. Journal of the American Chemical Society 133: 13876–13879.
Mayer, A.M.S., P.B. Jacobson, W. Fenical, R.S. Jabocs, and K.B. Glaser. 1998. Pharmacological characterization of the pseudopterosins: novel anti-inflammatory natural products isolated from the Caribbean soft coral, Pseudopterogorgia elisabethae. Life Sciences 62: 401–407.
Mehla, R., S. Bivalkar-Mehla, R. Zhang, I. Handy, H. Albrecht, S. Giri, P. Nagarkatti, M. Nagarkatti, and A. Chauhan. 2010. Bryostatin modulates latent HIV-1 infection via PKC and AMPK signaling but inhibits acute infection in a receptor independent manner. PLoS ONE 5: e11160.
Morens, D.M., G.K. Folkers, and A.S. Fauci. 2004. The challenge of emerging and re-emerging infectious diseases. Nature 430: 242–249.
Muller, U.G., S.A. Rehner, and T.R. Schultz. 1998. The evolution of agriculture in ants. Science 281: 2034–2038.
Mydlarz, L.D., R.S. Jacobs, J. Boehnlein, and R.G. Kerr. 2003. Pseudopterosin biosynthesis in Symbiodinium sp., the dinoflagellate symbiont of Pseudopterogorgia elisabethae. Chemistry and Biology 10: 1051–1056.
Nelson, T.J., and D.L. Alkon. 2009. Neuroprotective versus tumorigenic protein kinase C activators. Trends in Biochemical Sciences 34: 136–145.
Newman, D.J., and G.M. Cragg. 2012. Natural products as sources of new drugs over the 30 years from 1981 to 2010. Journal of Natural Products 75: 311–335.
Nirma, C., V. Eparvier, and D. Stien. 2013. Antifungal agents from Pseudallescheria boydii SNB-CN73 isolated from a Nasutitermes sp. termite. Journal of Natural Products 76: 988–991.
Oh, D.C., M. Poulsen, C.R. Currie, and J. Clardy. 2009a. Dentigerumycin: a bacterial mediator of an ant-fungus symbiosis. Nature Chemical Biology 5: 391–393.
Oh, D.C., M. Poulsen, C.R. Currie, and J. Clardy. 2011. Sceliphrolactam, a polyene macrocyclic lactam from a wasp-associated Streptomyces sp. Organic Letters 13: 752–755.
Oh, D.C., J.J. Scott, C.R. Currie, and J. Clardy. 2009b. Mycangimycin, a polyene peroxide from a mutualist Streptomyces sp. Organic Letters 11: 633–636.
Ortholand, J.Y., and A. Ganesan. 2004. Natural products and combinatorial chemistry: back to the future. Current Opinion in Chemical Biology 8: 271–280.
Park, S.H., K. Moon, H.S. Bang, S.H. Kim, D.G. Kim, K.B. Oh, J. Shin, and D.C. Oh. 2012. Tripartilactam, a cyclobutane-bearing tricyclic lactam from a Streptomyces sp. in a dung beetle’s brood ball. Organic Letters 14: 1258–1261.
Paul, V.J. 1992. Ecology and chemical defense against predation. In Ecological roles of marine natural products, ed. V.J. Paul, 171–175. Ithaca: Cornell University Press.
Pavan, M., and G. Bo. 1953. Pederin, the toxic principle obtained in the crystalline state from blister beetles (Paederus fuscipes). Physiologia Comparata et Oecologia 3: 307–312.
Pechanova, O., W.D. Stone, W. Monroe, T.E. Nebeker, K.D. Klepzig, and C. Yuceer. 2008. Global and comparative protein profiles of the pronotum of the southern pine beetle, Dendroctonus frontalis. Insect Molecular Biology 17: 261–277.
Pettit, G.R., H.L. Cherry, D.L. Doubek, D.L. Herald, E. Arnold, and J. Clardy. 1982. Isolation and structure of bryostatin 1. Journal of the American Chemical Society 104: 6846–6848.
Piel, J. 2002. A polyketide synthase-peptide synthetase gene cluster from an uncultured bacterial symbiont of Paederus beetles. Proceedings of the National Academy of Sciences of the United States of America 99: 14002–14007.
Plumbi, S. 2001. Humans as the world’s greatest evolutionary force. Science 29: 1786–1790.
Poulsen, M., C.D. Oh, J. Clardy, and C.R. Currie. 2011. Chemical analyses of wasp-associated Streptomyces bacteria reveal a prolific potential for natural products discovery. PLoS ONE 6: e16763.
Propper, D.J., V. Macaulay, K.J. O’Byrne, J.P. Braybrooke, S.M. Wilner, T.S. Ganesan, D.C. Talbot, and A.L. Harris. 1998. A phase II study of bryostatin 1 in metastatic malignant melanoma. British Journal of Cancer 78: 1337–1341.
Ritchie, K.B. 2006. Regulation of microbial populations by coral surface mucus and mucus-associated bacteria. Marine Ecology Progress Series 322: 1–14.
Rould-Lefevre, C. 2002. Termites: evolution, sociality, symbioses, ecology. Dordrecht: Kluwer.
Ruffner, D.E., E.W. Schmidt, and J.R. Heemstra. 2015. Assessing the combinatorial potential of the RiPP cyanobactin tru pathway. ACS Synthetic Biology 4: 482–492.
Sardar, D., E. Pierce, J.A. McIntosh, and E.W. Schmidt. 2015. Recognition sequences and substrate evolution in cyanobactin biosynthesis. ACS Synthetic Biology 4: 167–176.
Schaufelberger, D.E., M.P. Koleck, J.A. Beutler, A.M. Vatakis, A.B. Alvarado, P. Andrews, L.V. Marzo, G.M. Muschik, J. Roach, J.T. Ross, W.B. Lebherz, M.P. Reeves, R.M. Eberwein, L.L. Rodgers, R.P. Testerman, K.M. Snader, and S. Forenza. 1991. The large-scale isolation of bryostatin 1 from Bugula neritina following current good manufacturing practices. Journal of Natural Products 54: 1265–1270.
Schmidt, E.W., J.T. Nelson, D.A. Rasko, S. Sudek, J.A. Eisen, M.G. Haygood, and J. Ravel. 2005. Patellamide A and C biosynthesis by a microcin-like pathway in Prochloron didemni, the cyanobacterial symbiont of Lissoclinum patella. Proceedings of the National Academy of Sciences of the United States of America 102: 7315–7320.
Schmidt, E.W., S. Sudek, and M.G. Haygood. 2004. Genetic evidence supports secondary metabolic diversity in Prochloron spp., the cyanobacterial symbiont of a tropical ascidian. Journal of Natural Products 67: 1341–1345.
Scott, J.J., D.C. Oh, M.C. Yuceer, K.D. Klepzig, J. Clardy, and C.R. Currie. 2008. Bacterial protection of beetle-fungus mutualism. Science 322: 63.
Six, D.L., and K.D. Klepzig. 2004. Dendroctonus bark beetles as model systems for studies on symbiosis. Symbiosis 37: 207–232.
Smith, J.E., J. Kuwabara, K. Flanaga, S. duPlessis, J. Coney, J. Beets, M. Takabayashi, S. Barnes, J. Turner, D. Brown, B.K. Griesemeret, and F. Stanton. 2008. An unusual cyanobacterial bloom in Hawai’i. Coral Reefs 27: 851.
Strohm, E., and K.E. Linsenmair. 1995. Leaving the cradle: how beewolves (Philanthus triangulum F.) obtain the necessary spatial information for emergence. Zoology 98: 137–146.
Tan, Z., R.C. Turner, R.L. Leon, X. Li, J. Hongpaisan, W. Zheng, A.F. Logsdon, Z.J. Naser, D.L. Alkon, C.L. Rosen, and J.D. Huber. 2013. Bryostatin improves survival and reduces ischemic brain injury in aged rats after acute ischemic stroke. Stroke 44: 3490–3497.
Trindade-Silva, A.E., G.E. Lim-Fong, K.H. Sharp, and M.G. Haygood. 2010. Bryostatins: biological context and biotechnological prospects. Current Opinion in Biotechnology 21: 834–842.
Trost, B.M., and G. Dong. 2008. Total synthesis of bryostatin 16 using atom-economical and chemoselective approaches. Nature 456: 485–488.
Um, S., H.S. Bang, J. Shin, and D.C. Oh. 2013a. Actinofuranone C, a new 3-furanone-bearing polyketide from a dung beetle-associated bacterium. Natural Product Sciences 19: 71–75.
Um, S., A. Fraimout, P. Sapountzis, D.C. Oh, and M. Poulsen. 2013b. The fungus-growing termite Macrotermes natalensis harbors bacillaene-producing Bacillus sp. that inhibit potentially antagonistic fungi. Scientific Reports 3: 3250.
Um, S., S.H. Park, J. Kim, H.K. Park, K. Ko, H.S. Bang, S.K. Lee, J. Shin, and D.C. Oh. 2015. Coprisamides A and B, new cyclic peptides from a gut bacterium of the dung beetle Copris tripartitus. Organic Letters 17: 1272–1275.
Walsh, C. 2003a. Where will new antibiotics come from? Nature Reviews Microbiology 1: 65–70.
Walsh, C. 2003b. Antibiotics: actions, origins, resistance. Washington D. C.: ASM Press.
Wender, P.A., and A.J. Schrier. 2011. Total synthesis of bryostatin 9. Journal of the American Chemical Society 133: 9228–9231.
Withers, N., W. Vidaver, and R.A. Lewin. 1978. Pigment composition, photosynthesis and fine structure of a non-blue-green prokaryotic algal symbiont (Prochloron sp.) in a didemnid ascidian from Hawaiian waters. Phycologia 17: 167–171.
Woollacott, R.M. 1981. Association of bacteria with bryozoan larvae. Marine Biology 65: 155–158.
Woollacott, R.M., and R.L. Zimmer. 1975. A simplified placenta-like system for the transport of extraembryonic nutrients during embryogenesis of Bugula neritina (bryozoa). Journal of Morphology 147: 355–377.
Wright, G.D. 2007. The antibiotic resistome: the nexus of chemical and genetic diversity. Nature Reviews Microbiology 5: 175–186.
Wu, Q., N. Jiang, B.H. Wen, N.M. Ya, M.G. Hui, K.G. Zhi, S.W. Ng, and R.X. Tan. 2014. Antibacterial epipolythiodioxopiperazine and unprecedented sesquiterpene from Pseudallescheria boydii, a beetle (Coleptera)-associated fungus. Organic and Biomolecular Chemistry 12: 9412–9505.
Zhang, Y.L., H.M. Ge, W. Zhao, H. Dong, Q. Xu, S.H. Li, J. Li, J. Zhang, Y.C. Song, and R.X. Tan. 2008. Unprecedented immunosuppressive polyketides from Daldinia eschscholzii, a mantis-associated fungus. Angewandte Chemie International Edition 47: 5823–5826.
Acknowledgments
This work was supported by a Korean National Research Foundation (NRF) grants funded by the Korean government (No. 2009-0083533 and 2014R1A1A2057302).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare no conflict of interest with any person or any organization.
Rights and permissions
About this article

Cite this article
Choi, H., Oh, DC. Considerations of the chemical biology of microbial natural products provide an effective drug discovery strategy. Arch. Pharm. Res. 38, 1591–1605 (2015). https://doi.org/10.1007/s12272-015-0639-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12272-015-0639-y