
Abstract
Veronica peregrina has a wide range of types of constituents with various pharmacological properties. Here in this study, we isolated protocatechuic acid (PCA) from V. peregrina and examined PCAs effects on the lifespan and stress tolerance using Caenorhabditis elegans model system. We found that lifespan of wild-type worms was significantly lengthened in the presence of PCA in a dose dependent manner. PCA also elevated tolerance of worms against osmotic, heat shock, and oxidative stress. We also demonstrated antioxidant capacity of PCA by checking intracellular reactive oxygen species level and antioxidant enzyme activities such as catalase and superoxide dismutase. We further investigated several factors including pharyngeal pumping rate and progeny production that might influence prolonged lifespan and enhanced stress tolerance by PCA. Interestingly, both factors were significantly reduced after PCA exposure, indicating PCA exerts longevity activity by shifting food intake and reproduction at least in part. In addition, PCA-treated aged worms showed increased body movement compared to untreated controls suggesting PCA could enhance healthspan as well as lifespan.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Aging is the accumulation of changes over time which is associated with the increasing susceptibility to different diseases and death. Although there are several hypotheses to explain how aging occurs, exact aging process remains to be defined. Nonetheless, many evidences led to the general acceptance of the oxidative stress theory that the accumulation of molecular damage caused by reactive oxygen species (ROS) is a major factor in aging (Bokov et al. 2004; Finkel and Holbrook 2000). All organisms living in an aerobic condition are exposed to ROS consistently, due to the mitochondrial redox imbalance. Oxidative stress induced by ROS can lead to oxidation of biomolecules such as protein, lipid, and DNA which is suggested to be the central cause factor promoting aging process (Oliveira et al. 2010). Therefore, antioxidants which can prevent oxidative stress by scavenging radicals or interfering radical generation may delay aging and prolong the lifespan. Indeed, many previous studies on the correlation between antioxidants and aging support this notion (Wu et al. 2002; Ishii et al. 2004; Adachi and Ishii 2000; Harrington and Harley 1988).
Veronica peregrina (Scrophulariaceae) which is widely distributed through Korea and Japan has been used as a traditional medicine for the treatment of various diseases such as hemorrhage, leucorrhoea and pain. Recently, it has been reported that this plant has anti-inflammatory (Jeon 2012) and antioxidant properties (Ahn et al. 2011; Kwak et al. 2009). In addition, previous phytochemical studies revealed that V. peregrina has been found to possess flavonoids, iridoid glycosides, and phenolic compounds (Ahn et al. 2011; Kwak et al. 2009). Since phenolic compounds have been shown to increase lifespan in a wide range of species (Aires et al. 2012; Peng et al. 2011; Canuelo et al. 2012), we hypothesized that this plant may affect lifespan and aging. Indeed, our preliminary data revealed that the methanolic extract of V. peregrina has lifespan extension activity with increased stress resistance. Moreover, longevity activity of caffeic acid which is one of constituent of this plant has already been reported (Pietsch et al. 2011). In this regard, we assumed that another phenolic compound of V. peregrina such as protocatechuic acid (PCA) may also have similar properties.
To determine whether PCA can alter the lifespan and aging, we used the nematode, Caenorhabditis elegans as an in vivo model system. Recently, this model has become a popular for evaluating pharmacological properties of natural products on the aging, aging-related diseases, and stress-response mechanism (Tazearslan et al. 2009; Golden and Melov 2007; Antebi 2007; Brown et al. 2006). C. elegans has the ability of short lifespan, rapid generation, and ease of handling (Guarente and Kenyon 2000), and thus, can provide excellent environment for aging or lifespan research. Moreover, nematodes and mammals share many specific features of aging including loss of muscle mass (Herndon et al. 2002), behavioral declines (Glenn et al. 2004) as well as aging-related factors such as oxidative stress (Finkel and Holbrook 2000) and caloric restriction (Lakowski and Hekimi 1998).
The aim of this work was to study the effect of PCA on the lifespan and stress tolerance in C. elegans. In addition, antioxidant capacity of this compound was analyzed by measuring intracellular ROS level and antioxidant enzyme activities of nematodes. We also checked whether this compound affects aging parameters such as pharyngeal pumping, movement, and reproduction.
Materials and methods
Chemicals
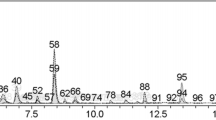
The isolation of PCA from V. peregrina has been detailed in a previous report (Ahn et al. 2011). Selected peptone and yeast extracts were obtained from BD bioscience (USA). Agar, 2′,7′-dichlorodihydrofluorescein diacetate, methyl viologen dichloride hydrate (paraquat), catalase, xanthine, xanthine oxidase, and nitroblue tetrazolium were purchased from Sigma (St. Louis, MO, USA).
Nematodes maintenance and PCA treatment
Bristol N2 (wild-type) was kindly provided by Dr. Myon-Hee Lee (East Carolina University, NC, USA). The worms were grown at 20 °C on nematode growth medium (NGM) agar plate with Escherichia coli OP50 as described by Brenner (Brenner 1974). To prepare plates supplemented with PCA, the stock solution in DMSO was inserted into autoclaved NGM plates (at 50 °C). A final DMSO concentration of 0.2 % (v/v) was maintained under all conditions.
Lifespan assay
The lifespan assays were performed five times independently at 20 °C. To obtain age-synchronized nematodes, eggs were transferred to NGM plate in the absence or presence of various concentration of PCA after embryo isolation. Test worms were considered dead when they failed to respond to prodding with the tip of a platinum wire. The worms were transferred to fresh NGM plate every 2 days. In order to test whether PCA may alter the lifespan of aged worms, on the 8th day of adulthood, worms were transferred to PCA or solvent containing plate, and then counted dead bodies as described above.
Determination of stress resistance
The age-synchronized N2 worms were bred on NGM agar plates with or without various concentrations of PCA. For the heat tolerance assay the adult day 4 worms were transferred to fresh plates and then incubated at 36 °C. The viability was scored over 10 h as previously described (Lee et al. 2005). Oxidative stress tolerance was assessed as described previously with minor modification (Mekheimer et al. 2012). Briefly, the adult day 7 worms were subjected to plate containing 60 mM paraquat and then survivals were recorded over 24 h. Resistance to osmotic stress was measured by placing the adult day 3 worms to NGM agar plate containing 500 mM NaCl (Horikawa and Sakamoto 2009). Survival rate of the worms was calculated after 12 h incubation. The survival of worms was determined touch-provoked movement (Lithgow et al. 1995). Worms which failed to respond to gentle touch with a platinum wire were considered to be dead. Each test was performed at least three times.
Measurement of antioxidant enzyme activities
To assess enzymatic activity, the worm homogenates were prepared. Briefly, the worms were harvested from plate with M9 buffer on the adult day 5 and washed three times. Then, the collected worms were resuspended in homogenization buffer (10 mM Tris–HCl, 150 mM NaCl, 0.1 mM EDTA, pH 7.5) and homogenized on ice. SOD activity was measured spectrophotometrically analysing the decolorization of formazan using enzymatic reaction between xanthine and xanthine oxidase. The reaction mixture contained 20 μL of worm homogenates and 480 μL of 1.6 mM xanthine, 0.48 mM nitroblue tetrazolium (NBT) in 10 mM phosphate buffer (pH 8.0). After pre-incubation at room temperature for 5 min, the reaction was initiated by adding 1 ml of xanthine oxidase (0.05 U/ml) and incubation at 37 °C for 20 min. The reaction was stopped by adding 500 μL of 69 mM SDS, and the absorbance at 570 nm was measured. SOD activity was expressed as a percentage of the scavenged amount per control. Catalase activity was calculated by spectrophotometry as previously described (Aebi 1984). Briefly, the prepared homogenates were mixed with the 25 mM H2O2 and after 5 min incubation, absorbance was determined at 240 nm. Catalase activity was expressed in U/mg protein (1 unit will decompose 1.0 μM of H2O2 per min at pH 7.0 at 25 °C).
Analysis of intracellular ROS
Intracellular ROS in the nematodes was measured using molecular probe 2′,7′-dichlorodihydrofluorescein diacetate (H2DCF-DA). Equal number of worms was incubated in the absence or presence of PCA. On the 4th day of adulthood, animals were exposed to NGM agar plate containing 30 mM paraquat for 3 h. Subsequently, five worms were transferred into the wells of a 96-well plate containing 50 μL of M9 buffer. Immediately after addition of 50 μL of 25 μM H2-DCF-DA solution resulting in a final concentration 12.5 μM, basal fluorescence was quantified in a microplate fluorescence reader at excitation 485 nm and emission 535 nm. Plates were read every 30 min for 2 h.
Measurement of pharyngeal pumping and body movement
The age-synchronized N2 worms were bred on NGM agar plates with or without various concentrations of PCA. On the 4th and 8th days of adulthood, single worms were transferred to fresh plate followed by pharynx contractions and body movements of animals were counted under an inverted microscope for 1 min. All the tests were repeated at least three times.
Reproduction assay
N2 worms were raised from embryo as in the lifespan assay. L4 larvae were individually transferred to the fresh plate every day to distinguish the parent from the progeny. The progeny was counted at the L2 or L3 stage. The test was completed in triplicate.
Data analysis
The values are expressed as the mean ± standard error of the mean. Animal survival rate was plotted using Kaplan–Meier analysis and analyzed by log-rank test. The statistical significance of differences between the control and treated groups were analyzed by one-way analysis of variance (ANOVA). p-values less than 0.01 were considered significant.
Results and discussion
Protocatechuic acid (PCA, 3,4-dihydroxybenzoic acid) is a catechol-type o-diphenol phenolic acid and has been shown to exert many pharmacological properties including anti-inflammatory (Tsai and Yin 2012; Wei et al. 2013), antiplatelet (Kim et al. 2012), antibacterial (Stojkovic et al. 2013), anti-metastatic (Lin et al. 2011), and antidiabetic activities (Harini and Pugalendi 2010). Above all, PCA has recently received a lot of attention because of the strong antioxidant potential (Sroka and Cisowski 2003; Masella et al. 2012). It is well known that excess of ROS causes chronic and degenerative diseases by oxidative modification of biomolecules including DNA mutation, protein denaturation, and lipid peroxidation. In particular, ROS-mediated oxidative stress plays a pivotal role in the process of aging, and thus, antioxidants can be an excellent candidate for effective anti-aging agent (Harman 1956; Queen and Tollefsbol 2010). In this regard, we dissected the effect of PCA on the lifespan and stress tolerance using C. elegans model system.
To determine the effect of PCA on the lifespan of wild-type worms, the survival curve for untreated (control) worms and PCA-treated worms was compared. Here in this study, PCA was shown to prolong the lifespan significantly compared to untreated controls under standard laboratory conditions at 20 °C (Fig. 1a). As shown in Fig. 1b, 200 μM PCA treatment caused a significant increased average lifespan (16.34 ± 0.78 days, p < 0.001), while the untreated worms was 11.76 ± 0.28 days. In addition, the maximum lifespan was also lengthened by PCA in a dose dependent manner from 20 to 25 days (100 μM) and 28 days (200 μM), respectively.

Effects of PCA on the lifespan of wild-type N2 nematodes. Worms were grown in the NGM agar plate at 20 °C in the absence or presence of PCA. The number of worms used per each lifespan assay experiment was 49–72 and three independent experiments were repeated (N = 3). a The mortality of each group was determined by daily counting of surviving and dead animals. b The mean lifespan of the N2 worms was calculated from the survival curves in (a). Statistical difference between the curves was analyzed by log-rank test. Error bars represent the standard error of mean (SEM). Differences compared to the control were considered significant at **p < 0.001 by one-way ANOVA
Many evidences revealed that increased viability under stress condition is closely linked with longevity (Munoz and Riddle 2003; Larsen 1993). Thus, in the current study, we evaluated whether PCAs lifespan extension activity is associated with increased stress tolerance. To answer this question, we compared the viability of PCA-treated worms with untreated controls under various stress conditions including thermal, osmotic, and oxidative stress. Our results showed that the PCA-treated worms exhibited enhanced resistance to osmotic stress as well as increased thermotolerance suggesting PCA could protect the worms under osmotic stress and heat shock conditions and might result in lifespan extension (Fig. 2a and b). Further, we checked the survival rate under oxidative stress induced by paraquat and, as expected, PCA showed significant enhanced tolerance against oxidative stress (Fig. 2c).

Effects of PCA on the stress tolerance of wild-type N2 nematodes. a Resistance to osmotic stress was measured by placing worms to NGM agar plate containing 500 mM NaCl and survival rate was calculated after 12 h incubation. b To assess thermal tolerance, worms were incubated at 36 °C and then their viability was scored. c For the oxidative stress assays, worms were transferred to NGM agar plate containing 60 mM of paraquat, and then their viability was scored. Statistical difference between the curves was analyzed by log-rank test. Error bars represent the standard error of mean (SEM). Differences compared to the control were considered significant at *p < 0.01 by one-way ANOVA
Then, we evaluate the possibility whether PCAs regulatory effect on the antioxidant enzyme activity might be involved in increased survival rate under oxidative stress condition. In the current study, we found that PCA was able to elevate catalase and superoxide dismutase activities of worms (Fig. 3a and b). These results are consistent with earlier findings that PCA enhances endogenous antioxidant enzyme activities in vitro (An et al. 2006) and in vivo (Zhang et al. 2011). In order to confirm whether enhanced resistance by PCA under oxidative stress condition is due to attenuation of excessive ROS accumulation in the cells, we quantified intracellular ROS levels of PCA-treated worms compared to untreated controls. As can be seen in Fig. 4, treatment of PCA effectively reduced intracellular ROS level in a dose dependent manner. Taken together, we could explain that PCAs radical scavenging and up-regulation of antioxidant enzyme activities resulting in attenuation of intracellular ROS level might give rise to enhanced resistance under stress condition and prolonged lifespan, at least to some extent.

Effects of PCA on the antioxidant enzyme activity of wild-type N2 nematodes. a The enzymatic reaction of xanthine with xanthine oxidase was used to generate •O2 − and the SOD activity was estimated spectrophotometrically through formazan formation by NBT reduction. SOD activity was expressed as a percentage of the scavenged amount per control. b Catalase activity was calculated from the concentration of residual H2O2, as determined by a spectrophotometric method. Catalase activity was expressed in U/mg protein. Data are expressed as the mean ± SEM of three independent experiments (N = 3). Differences compared to the control were considered significant at *p < 0.01 by one-way ANOVA

Effects of PCA on the intracellular ROS accumulation of wild-type N2 nematodes. The worms were incubated with 30 mM paraquat for 3 h, and subsequently treated with the fluorescent probe H2-DCF-DA. Intracellular ROS accumulation was quantified spectrometrically at excitation 485 nm and emission 535 nm. Plates were read every 30 min for 2 h. Data are expressed as the mean ± SEM of three independent experiments (N = 3). Differences compared to the control were considered significant at *p < 0.01 and **p < 0.001 by one-way ANOVA
Next, we analyzed several factors which might be associated with PCAs lifespan extension activity. Many previous studies have noted that the dietary restriction is closely linked with lifespan and aging in many species including C. elegans (Walker et al. 2005; Bordone and Guarente 2005). Here in this study, we could estimate the food intake by counting pharyngeal pumping. As noted in Fig. 5a, a reduced pharyngeal pumping rate was detected from the PCA-treated worms. Interestingly, in spite of decreased food intake, PCA-treated worms (8th day of adulthood) exhibited significant increase in body movements compared to untreated worms (Fig. 5b), indicating PCA has not only lifespan extension properties, but also healthspan improving activity.

Effects of PCA on the pharynx pumping rate and body movement of wild-type N2 nematodes. On the 4th and 8th days of adulthood, the pharyngeal pumping rates (a) and body movement (b) were counted under a dissecting microscope for 1 min. Data are expressed as the mean ± SEM of three independent experiments (N = 3). Differences compared to the control were considered significant at **p < 0.001 by one-way ANOVA
In addition, recent studies have provided evidences that reproduction and longevity are directly coupled. Indeed, dramatic lifespan extension could be observed by targeted ablation of germline stem cells in C. elegans (Arantes-Oliveira et al. 2002). In this regards, we also examined whether PCA affects nematode’s reproduction. Interestingly, PCA shifted not only total offspring per nematode but the distribution of reproductive capacity. As shown in Fig. 6, the total number of progeny was significantly reduced by PCA. Moreover, PCA-treated worms displayed delayed egg-laying, demonstrating PCA might affect germline signaling results in altered germline developmental events such as spermatogenesis and oogenesis. Since germline signaling takes an important role in aging of soma in C. elegans, our results revealed that regulatory activity of PCA on the germline might be possibly related with extended lifespan.

Effects of PCA on the reproduction of wild-type N2 nematodes. Daily and total reproductive outputs were counted. The offspring of each worm were observed prior to L4 stage. Data are expressed as the mean ± SEM of three independent experiments (N = 3). Differences compared to the control were considered significant at *p < 0.01 and **p < 0.001 by one-way ANOVA
To address the possibility that PCA may shift worm’s lifespan directly, independent of its effects on the food intake and reproduction, the aged infertile worms (Adult day 8) were exposed to PCA and their lifespan was checked again. Interestingly, PCA-treated aged worms exhibited enhanced lifespan (Fig. 7a) without affecting pharyngeal pumping rate (Fig. 7b) in aged nematodes, though not as drastically as in the worms which were exposed to PCA after embryo isolation. These findings provide strong evidence that PCA can extends lifespan directly. However, further studies are required to elucidate the exact mechanism of PCAs lifespan extension and anti-aging activities.

Effects of PCA on the lifespan and pharynx pumping rate of aged nematodes. Age synchronized worms were grown in the NGM agar plate at 20 °C. On the 8th day of adulthood, worms were transferred to NGM agar plate in the absence or presence of PCA. a The mortality of each group was determined by daily counting of surviving and dead animals. b On the 12th day of adulthood, the pharyngeal pumping rates were counted under a dissecting microscope for 1 min. Statistical difference between the curves was analyzed by log-rank test. Error bars represent the standard error of mean (SEM). Differences compared to the control were considered by one-way ANOVA
References
Adachi, H., and N. Ishii. 2000. Effects of tocotrienols on life span and protein carbonylation in Caenorhabditis elegans. Journals of Gerontology A, Biological Sciences and Medical Sciences 55: B280–B285.
Aebi, H. 1984. Catalase in vitro. Methods in Enzymology 105: 121–126.
Ahn, D.R., S.I. Lee, and J.H. Yang. 2011. Superoxide radical scavengers from the whole plant of Veronica peregrina. Natural Product Sciences 17: 142–146.
Aires, D.J., G. Rockwell, T. Wang, J. Frontera, J. Wick, W. Wang, M. Tonkovic-Capin, J. Lu, E. Lezi, H. Zhu, and R.H. Swerdlow. 2012. Potentiation of dietary restriction-induced lifespan extension by polyphenols. Biochimica et Biophysica Acta 1822: 522–526.
An, L.J., S. Guan, G.F. Shi, Y.M. Bao, Y.L. Duan, and B. Jiang. 2006. Protocatechuic acid from Alpinia oxyphylla against MPP+-induced neurotoxicity in PC12 cells. Food and Chemical Toxicology 44: 436–443.
Antebi, A. 2007. Ageing: When less is more. Nature 447: 536–537.
Arantes-Oliveira, N., J. Apfeld, A. Dillin, and C. Kenyon. 2002. Regulation of life-span by germ-line stem cells in Caenorhabditis elegans. Science 295: 502–505.
Bokov, A., A. Chaudhuri, and A. Richardson. 2004. The role of oxidative damage and stress in aging. Mechanisms of Ageing and Development 125: 811–826.
Bordone, L., and L. Guarente. 2005. Calorie restriction, SIRT1 and metabolism: Understanding longevity. Nature Reviews Molecular Cell Biology 6: 298–305.
Brenner, S. 1974. The genetics of Caenorhabditis elegans. Genetics 77: 71–94.
Brown, M.K., J.L. Evans, and Y. Luo. 2006. Beneficial effects of natural antioxidants EGCG and alpha-lipoic acid on life span and age-dependent behavioral declines in Caenorhabditis elegans. Pharmacology, Biochemistry and Behavior 85: 620–628.
Canuelo, A., B. Gilbert-Lopez, P. Pacheco-Linan, E. Martinez-Lara, E. Siles, and A. Miranda-Vizuete. 2012. Tyrosol, a main phenol present in extra virgin olive oil, increases lifespan and stress resistance in Caenorhabditis elegans. Mechanisms of Ageing and Development 133: 563–574.
Finkel, T., and N.J. Holbrook. 2000. Oxidants, oxidative stress and the biology of ageing. Nature 408: 239–247.
Glenn, C.F., D.K. Chow, L. David, C.A. Cooke, M.S. Gami, W.B. Iser, K.B. Hanselman, I.G. Goldberg, and C.A. Wolkow. 2004. Behavioral deficits during early stages of aging in Caenorhabditis elegans result from locomotory deficits possibly linked to muscle frailty. Journals of Gerontology A, Biological Sciences and Medical Sciences 59: 1251–1260.
Golden, T.R., and S. Melov. 2007. Gene expression changes associated with aging in C. elegans. WormBook 12: 1–12.
Guarente, L., and C. Kenyon. 2000. Genetic pathways that regulate ageing in model organisms. Nature 408: 255–262.
Harini, R., and K.V. Pugalendi. 2010. Antihyperglycemic effect of protocatechuic acid on streptozotocin-diabetic rats. Journal of Basic and Clinical Physiology and Pharmacology 21: 79–91.
Harman, D. 1956. Aging: A theory based on free radical and radiation chemistry. Journals of Gerontology 11: 298–300.
Harrington, L.A., and C.B. Harley. 1988. Effect of vitamin E on lifespan and reproduction in Caenorhabditis elegans. Mechanisms of Ageing and Development 43: 71–78.
Herndon, L.A., P.J. Schmeissner, J.M. Dudaronek, P.A. Brown, K.M. Listner, Y. Sakano, M.C. Paupard, D.H. Hall, and M. Driscoll. 2002. Stochastic and genetic factors influence tissue-specific decline in ageing C. elegans. Nature 419: 808–814.
Horikawa, M., and K. Sakamoto. 2009. Fatty-acid metabolism is involved in stress-resistance mechanisms of Caenorhabditis elegans. Biochemical and Biophysical Research Communications 390: 1402–1407.
Ishii, N., N. Senoo-Matsuda, K. Miyake, K. Yasuda, T. Ishii, P.S. Hartman, and S. Furukawa. 2004. Coenzyme Q10 can prolong C. elegans lifespan by lowering oxidative stress. Mechanisms of Ageing and Development 125: 41–46.
Jeon, H. 2012. Anti-inflammatory activity of Veronica peregrina. Natural Product Sciences 18: 141–146.
Kim, K., O.N. Bae, K.M. Lim, J.Y. Noh, S. Kang, K.Y. Chung, and J.H. Chung. 2012. Novel antiplatelet activity of protocatechuic acid through the inhibition of high shear stress-induced platelet aggregation. Journal of Pharmacology and Experimental Therapeutics 343: 704–711.
Kwak, J.H., H.J. Kim, K.H. Lee, S.C. Kang, and O.P. Zee. 2009. Antioxidative iridoid glycosides and phenolic compounds from Veronica peregrina. Archives of Pharmacal Research 32: 207–213.
Lakowski, B., and S. Hekimi. 1998. The genetics of caloric restriction in Caenorhabditis elegans. Proceedings of the National Academy Science USA 95: 13091–13096.
Larsen, P.L. 1993. Aging and resistance to oxidative damage in Caenorhabditis elegans. Proceedings of the National Academy Science USA 90: 8905–8909.
Lee, E.Y., Y.H. Shim, D.J. Chitwood, S.B. Hwang, J. Lee, and Y.K. Paik. 2005. Cholesterol-producing transgenic Caenorhabditis elegans lives longer due to newly acquired enhanced stress resistance. Biochemical and Biophysical Research Communications 328: 929–936.
Lin, H.H., J.H. Chen, F.P. Chou, and C.J. Wang. 2011. Protocatechuic acid inhibits cancer cell metastasis involving the down-regulation of Ras/Akt/NF-kappaB pathway and MMP-2 production by targeting RhoB activation. British Journal of Pharmacology 162: 237–254.
Lithgow, G.J., T.M. White, S. Melov, and T.E. Johnson. 1995. Thermotolerance and extended life-span conferred by single-gene mutations and induced by thermal stress. Proceedings of the National Academy Science USA 92: 7540–7544.
Masella, R., C. Santangelo, M. D’Archivio, G. Li Volti, C. Giovannini, and F. Galvano. 2012. Protocatechuic acid and human disease prevention: Biological activities and molecular mechanisms. Current Medicinal Chemistry 19: 2901–2917.
Mekheimer, R.A., A.A. Sayed, and E.A. Ahmed. 2012. Novel 1,2,4-triazolo[1,5-a]pyridines and their fused ring systems attenuate oxidative stress and prolong lifespan of Caenorhabiditis elegans. Journal of Medicinal Chemistry 55: 4169–4177.
Munoz, M.J., and D.L. Riddle. 2003. Positive selection of Caenorhabditis elegans mutants with increased stress resistance and longevity. Genetics 163: 171–180.
Oliveira, B.F., J.A. Nogueira-Machado, and M.M. Chaves. 2010. The role of oxidative stress in the aging process. ScientificWorldJournal 10: 1121–1128.
Peng, C., H.Y. Chan, Y. Huang, H. Yu, and Z.Y. Chen. 2011. Apple polyphenols extend the mean lifespan of Drosophila melanogaster. Journal of Agriculture and Food Chemistry 59: 2097–2106.
Pietsch, K., N. Saul, S. Chakrabarti, S.R. Sturzenbaum, R. Menzel, and C.E. Steinberg. 2011. Hormetins, antioxidants and prooxidants: defining quercetin-, caffeic acid- and rosmarinic acid-mediated life extension in C. elegans. Biogerontology 12: 329–347.
Queen, B.L., and T.O. Tollefsbol. 2010. Polyphenols and aging. Current Aging Science 3: 34–42.
Sroka, Z., and W. Cisowski. 2003. Hydrogen peroxide scavenging, antioxidant and anti-radical activity of some phenolic acids. Food and Chemical Toxicology 41: 753–758.
Stojkovic, D.S., J. Zivkovic, M. Sokovic, J. Glamoclija, I.C. Ferreira, T. Jankovic, and Z. Maksimovic. 2013. Antibacterial activity of Veronica montana L. extract and of protocatechuic acid incorporated in a food system. Food and Chemical Toxicology 55: 209–213.
Tazearslan, C., S. Ayyadevara, P. Bharill, and R.J. Shmookler Reis. 2009. Positive feedback between transcriptional and kinase suppression in nematodes with extraordinary longevity and stress resistance. PLoS Genetics 5: e1000452.
Tsai, S.J., and M.C. Yin. 2012. Anti-glycative and anti-inflammatory effects of protocatechuic acid in brain of mice treated by d-galactose. Food and Chemical Toxicology 50: 3198–3205.
Walker, G., K. Houthoofd, J.R. Vanfleteren, and D. Gems. 2005. Dietary restriction in C. elegans: From rate-of-living effects to nutrient sensing pathways. Mechanisms of Ageing and Development 126: 929–937.
Wei, M., X. Chu, M. Guan, X. Yang, X. Xie, F. Liu, C. Chen, and X. Deng. 2013. Protocatechuic acid suppresses ovalbumin-induced airway inflammation in a mouse allergic asthma model. International Immunopharmacology 15: 780–788.
Wu, Z., J.V. Smith, V. Paramasivam, P. Butko, I. Khan, J.R. Cypser, and Y. Luo. 2002. Ginkgo biloba extract EGb 761 increases stress resistance and extends life span of Caenorhabditis elegans. Cellular and molecular biology (Noisy-le-grand) 48: 725–731.
Zhang, X., G.F. Shi, X.Z. Liu, L.J. An, and S. Guan. 2011. Anti-ageing effects of protocatechuic acid from Alpinia on spleen and liver antioxidative system of senescent mice. Cell Biochemistry and Function 29: 342–347.
Acknowledgments
Following are results of a study on the “Leades INdustry-university Cooperation of Woosuk University” Project, supported by the Ministry of Education, Science & Technology (MEST).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kim, Y.S., Seo, H.W., Lee, MH. et al. Protocatechuic acid extends lifespan and increases stress resistance in Caenorhabditis elegans . Arch. Pharm. Res. 37, 245–252 (2014). https://doi.org/10.1007/s12272-013-0183-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12272-013-0183-6