
Abstract
Cardiovascular diseases represent one of the major causes for increasing rates of human morbidity and mortality across the world. This reinforces the necessity for the development of novel diagnostics and therapies for the early identification and cure of heart diseases. MicroRNAs are evolutionarily conserved small regulatory non-coding RNAs that regulate the expression of large number of genes. They are involved in several cellular pathophysiological pathways and have been shown to play a significant role in the pathogenesis of many disease states. Recent studies have correlated dysregulated miRNA expressions to diseased hearts and also shown the relevance of miRNA in growth, development, function, and stress responsiveness of the heart. The possibility of exploiting miRNAs to develop diagnostic markers or manipulating them to obtain therapeutic effects is very attractive since they have very specific targets in a particular cellular pathway. In this review we will summarize the role played by miRNAs in the heart and discuss the scope of utilizing miRNA-based strategies in the clinics for the benefit of mankind.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Diseases of the heart are the number one cause of death globally in the industrialized world. These are projected to remain the single leading cause of death around the world due to aging, sedentary lifestyle, obesity, and diabetes. Life in its entirety is dependent on the second-to-second function of the heart. Even subtle perturbations during cardiac development in the embryo or during transmission of electric impulses in the adult heart can be catastrophic. Several studies have highlighted the need for precise regulation of key molecular pathways during embryonic development especially in the cardiovascular system. Despite a detailed understanding of the molecular mechanisms underlying cardiac development, cardiac malformations are the most prevalent birth defect and the number one cause of noninfectious infant mortality [41]. Thus there is an innate need for innovative new therapies and diagnostic tools for cardiovascular diseases. Recent studies have unveiled a novel class of gene regulators called microRNAs (miRNAs) that have been demonstrated to play unexpected and powerful roles in diverse aspects of cardiac development and function. In addition, miRNAs have also been implied in the pathogenesis of diverse disease states of the heart thereby opening up new avenues for the development of miRNA-based diagnostics and therapeutics for cardiovascular diseases. In this review, we will briefly discuss miRNA biology and the role of miRNAs in the heart. We will also briefly analyze the opportunities and challenges that we face by translating this knowledge into the clinical setting.
miRNA Biogenesis
Mature miRNAs are an evolutionarily conserved class of small non-coding RNAs typically 22 nucleotide long that have gained status as important regulators of cardiac development and pathological processes [22, 64]. In general, they are transcribed by RNA polymerase II as primary transcripts (pri-miRNAs) [67], though some miRNAs that are interspersed among repetitive DNA elements like Alu repeats can also be transcribed by RNA polymerase III [10]. A significant number of pri-miRNAs are expressed as polycistronic RNAs [3]. The pri-miRNAs are subsequently processed by Drosha and DGCR8 (Pasha) complex resulting in an ∼70 nucleotide long-stem loop structure called precursor-miRNA (pre-miRNA). From the nucleus, the pre-miRNAs are then exported to cytoplasm by exportin-5, where they are processed by Dicer, an RNAse III enzyme, and bound to miRNA-induced silencing complex (RISC) to form the mature miRNAs of 18–23 nucleotide length. The key proteins in the RISC complex are Argonaute 2 and transactivation-responsive RNA-binding protein. In the RISC complex, the active strand is selectively retained and the passenger or star (*) strand is degraded. The mature miRNA along with the RISC binds to complementary sites in the mRNA transcripts and negatively regulate gene expression [6, 33, 66]. miRNAs that bind to mRNA targets with imperfect complementarity, regulate the target gene at the level of protein translation, while miRNAs that bind to their mRNA targets with perfect complementarity induce target mRNA cleavage [33, 133]. miRNA biogenesis is shown in Fig. 1.

Biogenesis of microRNAs: (A) transcription of primary microRNAs (pri-miRNAs) by RNA polymerase II in the nucleus. (B) Pri-miRNA is processed by Drosha and DGCR8 to form a precursor-miRNA (pre-miRNA) of about 70 nucleotides in length. (C) Pre-miRNA is exported to cytoplasm by exportin 5. (D) Final processing of pre-miRNA to mature duplex miRNA by RNase enzyme Dicer. (E) Incorporation of mature duplex into RNA-induced silencing complex where miRNA* strand is selectively degraded. (F) Binding of complex to the target mRNA guided by mature miRNA. (G) Negative regulation of protein translation or degradation of the mRNA transcript based on the complementarity of the miRNA to the target sequence. Some figure templates were adopted from Ambion website
miRNA-Mediated Target Gene Regulation
miRNAs regulate gene expression at the post-transcriptional level by mRNA degradation, translational repression, or miRNA-mediated mRNA decay. About 700 miRNAs have been identified so far and about 1,000 are predicted in humans [39]. Each miRNA is able to regulate expression of multiple target genes often involved in the same cellular pathway [37]. Therefore, it can be safely presumed that about 50% of human genes are regulated by miRNAs. Conversely, each gene can be target of multiple miRNAs thereby providing wide regulatory potential [37, 68]. In general, miRNA interacts with the 3′ untranslated region (UTR) of their target gene. However, recent reports demonstrate that miRNA targeting can occur in 5′UTR, open reading frames as well as in promoter regions [54, 65, 77, 90]. In some instances miRNAs can also upregulate gene expression and positively impact protein translation [125].
In mammals, miRNA-mediated gene regulation is generally accomplished by imperfect base pairing of miRNAs to their target genes. Several mechanisms that occur such as protein translation pre-initiation [81, 89] and post-initiation [88] translational repression of target genes have been put forward. Cytoplasmic structures such as P-bodies or processing bodies have been implicated in miRNA regulation [69]. These P-bodies store the repressed mRNA and restrict the access of protein translational machineries to mRNA [33]. Understanding miRNA–mRNA association is a complex task. A single miRNA can regulate or fine tune multiple genes. On the other hand, a mRNA transcript can be regulated by one or more miRNAs, depending on the number of miRNA complementary sites in the 3′UTR of the target gene [51]. Details on miRNA target prediction and regulation is reviewed by Bartel [6].
Cardiac Development and Regeneration
miRNAs in Normal Heart
The expression of miRNAs is dynamic, and is largely dependent on tissue or cell types, metabolic status and disease state. As in any other tissue, cardiac tissue also displays a ‘signature expression’ pattern for miRNAs. The most abundant miRNAs in heart are miR-1, let-7, miR-133, miR-126-3p, miR-143, miR-30c, and miR-22 [60]. Among these, miR-1 accounts for 45% of all mouse miRNAs found in heart, suggesting a role for these miRNAs in tissue specification or cell lineage decisions [60]. Recently, Rao and coworkers performed deep sequencing of miRNAs from heart tissue in male and female mice [94] and observed striking abundance of miR-1 (that accounted for nearly 40%) compared to other known miRNAs in heart. In addition, they also observed other miRNAs, including miR-29a, miR-26a, let-7 family members, to be more abundant than miR-133a, although miR-1 and miR-133 are coregulated. This suggests a role for mechanisms other than transcription (e.g., processing or stability) to dramatically alter steady state levels of mature miRNAs. Furthermore, they performed cardiomyocyte-specific deletion of dgcr8, a gene required for miRNA biogenesis. This revealed a fully penetrant phenotype that begins with left ventricular malfunction, progressing to a dilated cardiomyopathy and premature lethality [94]. The reported phenotype is similar to that described for the knockout of Dicer in the adult myocardium [25]. Taken together, this suggests that the dramatic cardiac phenotype observed in the absence of dicer or dgcr8, maybe contributed by just a handful of miRNAs [94]. However, one cannot rule out the contribution of low abundance or temporally regulated miRNAs in the heart. This further implies that to develop therapies against cardiovascular diseases, we need to decipher the molecular function of these specific miRNAs associated with heart.
The heart is derived from multiple cell lineages and must differentiate into unique regions, each possessing different physiological, electric and anatomic properties. Distinct patterns of gene expression, resulting from tight temporal and spatial regulation, guide the differentiation of each region of the heart. One approach to study the importance of miRNAs in the normal development of vertebrates has been to create loss-of-function mutations in Dicer. These studies in zebrafish and mice revealed the importance of miRNAs in development including cardiogenesis [8, 128]. Both zebrafish and mice lacking Dicer died at gastrulation indicating the essentiality of miRNAs for normal development. Further studies aimed at generating cardiac-specific deletion of Dicer in mice, using Cre recombinase under the control of the endogenous Nkx2.5 regulatory elements [137]. This resulted in embryonic lethality by E12.5 due to poorly developed ventricular myocardium. It will be interesting to determine whether miRNAs play a role in cardiac lineage specification by deleting Dicer during earlier steps of cardiogenesis. It is also understood that Dicer activity is required for normal function of mature heart, as adult mice lacking Dicer in the myocardium have high incidence of sudden death, cardiac hypertrophy and reactivation of fetal cardiac gene program [25]. Further studies aiming at targeted deletion of Dicer in specific regions of the heart will unravel the role of miRNAs in distinct aspects of cardiac development.
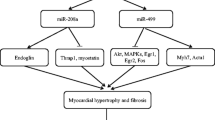
Of the 700 mammalian miRNAs identified to date, miR-1, miR-133 and miR-208 are considered muscle specific, being primarily expressed in cardiac and skeletal muscles [15, 59, 93, 105, 121, 137, 138]. miR-1 and miR-133 are derived from a common precursor transcript (bicistronic) and multiple highly conserved loci encode mature miR-1 (miR-1-1 and miR1-2) and miR-133 (miR-133a-1, miR-133a-2, and miR133b) transcripts, which appear to be genetically redundant. miR-208 is encoded by an intron in the α-MHC gene [121]. Cardiac muscle-specific expression of miR-1 and miR-133 are controlled by serum response factor/myocardin while miR-208 expression is associated with the expression of α–MHC gene. Recently, three myosin genes (Myh6, Myh7, and Myh7b) were shown to encode related intronic miRNAs (miR-208a, miR-208b, and miR-499) that control muscle performance [119].
In contrast, the skeletal muscle-specific expression of miR-1 and miR-133 are controlled by MyoD/MEF2 [15, 70]. These data suggest that transcription factors play a predominant role in the tissue-specific expression of miRNAs. In addition, these miRNAs also show differential cis-regulation, which in turn modulates their regulation of target mRNAs in the muscle differentiation pathway downstream of MEF2. For instance, the upstream miR-1/133 enhancer directs expression within the ventricular chambers [138], while the intragenic miR-1/133 enhancer is active in the atrial and ventricular chambers [70].
Cardiac Regeneration
Myocardial regeneration provides one of the most attractive therapeutic options to restore myocardial function. Several studies have shown the existence of multiple sources of stem cells for myocardial regeneration [7, 52]. Cardiac injury also promotes homing of circulating hematopoietic progenitor cells to sites of ischemia [55, 109]. The role of miRNAs in determining cardiac differentiation of mouse embryonic stem cells was addressed by Ivey and coworkers [47]. They observed miR-1 and miR-133 to be highly expressed in mouse cardiomyocytes derived from mouse embryonic stem cells. These miRNAs were potent repressors of nonmuscle gene expression and cell fate while enhancing mesodermal gene expression during mouse and human ES cell differentiation. Further they attributed the effects of miR-1 in part to be mediated by translational repression of the Notch ligand Delta-like 1 (Dll-1). miR-126 is another miRNA expressed in hematopoietic stem cells which can potentially contribute to myocardial regeneration [61]. Recently, miR-145 and miR-143, which are direct transcriptional targets of serum response factor, myocardin and Nkx2-5, were shown to regulate smooth muscle cell fate [21]. While miR-145 was shown to be sufficient to induce differentiation of multipotent neural crest stem cells into vascular smooth muscle, miR-145 and miR-143 function to regulate the quiescent versus proliferative phenotype of smooth muscle cells. In an earlier study, miR-145 was shown to be a novel vascular smooth muscle cell phenotypic marker and modulator that is capable of controlling vascular neointimal lesion formation [18].
Aberrant miRNA Function in Diseased Heart
miRNA expression profiles in a given tissue is also dependent on the disease state. A particular pathological process is often associated with a particular gene expression pattern as depicted in various disease conditions [11, 73, 106]. These signature patterns can aid in the diagnosis and prognosis of human diseases, as evidenced by recent studies in cancer and cardiovascular diseases. The implications of miRNA in cardiovascular diseases have become a rapidly evolving field and have been extensively reviewed recently [5, 13, 22, 64]. Hence in this review we will briefly explain the recent findings with special focus on the clinical significance of these studies. A summary of the different miRNAs implicated in cardiovascular diseases are listed in Table 1 with their potential targets and functions in heart.
Cardiac Hypertrophy
The adult heart is susceptible to many biomechanical stresses and pathologic stimuli and even slight perturbations during cardiogenesis can lead to catastrophe. In order to adapt to the workload and impaired cardiac function, the heart may respond by undergoing extensive cardiac remodeling known as cardiac hypertrophy [43]. This process involves redeployment of the molecular events controlling cardiac gene expression during development. Though this is an adaptive mechanism that is beneficial in the short-term, prolonged hypertrophy can lead to heart failure and sudden death [36].
To date, there had been several studies on the role of miRNAs in cardiac hypertrophy [14, 17, 97, 112, 115, 120, 121]. The common finding of these studies is that an array of miRNAs is significantly altered in their expression in either direction, and that single miRNAs can critically determine the progression of this disease [63]. Furthermore, using both gain- and loss-of-function approaches in mice, different groups have established a correlation between miRNAs and hypertrophy by demonstrating that stress-regulated miRNAs can trigger both positive and negative influence on the hypertrophic growth response [14, 120, 121]. For instance, cardiac-specific overexpression of miR-195, which is upregulated in both human and mouse hypertrophic hearts, can result in dilated cardiomyopathy and heart failure in mice within 2 weeks of birth [120]. However, the mechanisms underlying this regulation are still not clear.
On the other hand, deletion of miR-208, which is essential for expression of genes involved in cardiac hypertrophy, but not cardiogenesis, resulted in aberrant response to cardiac stress [121]. This miRNA targeted expression of a thyroid hormone signaling pathway component, Thrap1, and provides an example for miRNA function linked to classical hormone-regulated muscle physiology. Recently, miR-208a was found to be sufficient to induce cardiac arrhythmias and was identified as a key regulatory molecule necessary for proper cardiac conduction [12].
Another miRNA, miR-21, implicated in tumor-related cell growth and apoptosis [4, 83], is consistently induced by stress signals in vitro and in vivo [17, 97, 112, 120]. However, there is some controversy regarding its exact role in cardiac hypertrophy. Recently, Thum and coworkers [116] validated miR-21 as a disease target for heart failure, which has therapeutic implications. miR-21 regulates the ERK–MAP kinase signaling pathway through inhibition of sprouty homologue 1 (Spry1), in cardiac fibroblasts, which has impact on global cardiac structure and function. They further demonstrated that in vivo silencing of miR-21 by a chemically modified antagomir in a mouse pressure-overload-induced disease model reduced cardiac ERK–MAP kinase activity and inhibited interstitial fibrosis and attenuated cardiac dysfunction.
Heart Failure
Heart failure is a frequent and fatal outcome of hypertrophy developed under pathological circumstances. Most of our current knowledge on miRNAs in heart failure comes from miRNA expression analyses performed using microarrays in either experimental models that lead to pathologic remodeling of the heart [14, 97, 120] or in cultured cardiomyocytes that have been stimulated with peptides that mimic neurohormonal activation and/or have been shown to provoke myocyte hypertrophy in vitro [14, 17, 112, 120]. Studies on miRNA expression profiling in heart failure in humans is largely limited by the lack of standardized protocols, the small number of patients and the high degree of variability in expression levels between patients. A comparison of the altered expression profiles of miRNAs in various animal models of heart disease and in human heart failure [46, 115] revealed good concordance between these two sources of heart failure, especially for miRNAs that show increased levels of expression [26]. Twenty-five miRNAs were upregulated in one or more myocardial samples from failing human hearts and experimental models. This indicates that changes in miRNA expression profiles in experimental models may provide further insights into our understanding of heart failure. On the other hand, there was far less concordance in the expression of downregulated miRNAs, which may be due to the different microarray platforms used in the above studies.
Ikeda and coworkers [46] were the first group to do a global analysis of miRNA expression in human heart disease and to show the commonalities in expression between distinct disease etiologies. They compared miRNA expression in three different types of failing hearts (ischemic or dilated cardiomyopathy and aortic stenosis) with nonfailing heart. Among the 87 miRNAs detected in the heart, ∼50% were differentially expressed in at least one disease group, while seven miRNAs showed similar regulation in all three disease states. By classifying samples by etiological diagnosis, this study provides proof-of-concept that miRNA expression profiles may be useful in other more challenging and clinically relevant etiology prediction problems and supports further studies on miRNAs as potential biomarkers for determining prognosis and response to therapy.
A hallmark of heart failure is the reactivation of fetal gene program. A study by Thum and coworkers [115] compared miRNA expression profiles from failing, nonfailing, and fetal human hearts and concluded that miRNA levels may contribute to this reactivation. They observed that a large fraction (>80%) of miRNAs were either up- or downregulated in the same direction in fetal and failing heart in comparison with normal heart. In yet another recent study involving miRNA profiling in 70 patients [86], two novel miRNAs, miR-7, and miR-378 were observed to be downregulated in end-stage heart failure (dilated cardiomyopathy). Furthermore, they performed network analysis that suggested that a major process affected by miRNA regulation associated with heart failure progression is probably through NFκB target genes. Using transfected cells and mice, they show that NFκB, which is not a predicted target for miR-378, is altered following changes in miR-378. Thus, they show for the first time that perturbation in miRNAs leads to modulation in the network by a feed-in mechanism thus indicating a pivotal role for miRNAs in altering global signaling networks during progression of cardiac pathology. This study lays a foundation for the concept that therapeutic interventions with miRNAs would have global effects on signaling networks.
Cardiac Fibrosis
After myocardial infarction (MI), cardiac fibroblasts undergo negative remodeling by excessive secretion of extracellular matrix. Myocardial fibrosis leads to mechanical stiffness leading to contractile dysfunction and prevents myocardial repair. It also predisposes to cardiac arrhythmias through abnormalities in cardiac conductance. Van Rooij and coworkers [122], in a seminal work, elucidated the role of miRNAs in cardiac fibrosis following MI by profiling expression in the border zone of the infarct and remote myocardium at two time points (3 and 14 days). They observed dramatic downregulation of miR-29 family in the border zone and implicated miR-29 in the induction of collagens and other extracellular matrix genes. miR-29 is also downregulated in cardiac hypertrophy and failure [120] suggesting that therapeutic upregulation of miR-29 in response to ischemic event or cardiac stress may prevent onset of cardiac fibrosis. Earlier, we and others have shown significant downregulation of miR-29 family miRNAs in rhabdomyosarcoma (skeletal muscle tumors), implicating miR-29 in muscle cell differentiation [106, 127]. miR-21 is another miRNA that was found to be enriched in cardiac fibroblasts compared to cardiomyocytes [114]. miR-21 was shown to augment ERK/MAP kinase signaling thereby controlling interstitial fibrosis and inhibition of miR-21 using antagomirs reversed the effect [116]. Recently, miR-133 and miR-30 were shown to directly downregulate connective tissue growth factor (CTGF), a key molecule in the process of fibrosis and a potential therapeutic target [28]. By overexpression and knockdown in cultured cardiomyocytes and fibroblasts, the authors establish an important role for these miRNAs in the control of structural changes in the extracellular matrix of the myocardium by targeting CTGF.
Arrhythmias
Arrhythmias arise from a perturbation in the electrophysiological properties of heart muscle. Though arrhythmias arise as a result of cardiac disease or mutation in ion channel genes, miRNAs, miR-1 and miR-133 have recently been implicated to function in cardiac conduction system [75, 76, 132, 137]. Targeted deletion of miR-1-2 in mice resulted in 50% lethality attributed to ventricular septal defects [137]. Mutant mice that survived until birth experienced sudden death following electrophysiological defects. This effect was attributed to the upregulation of Irx5 transcription factor that regulates the cardiac ventricular repolarization gradient. The involvement of miR-1 in cardiac conductance was further confirmed by over expression studies in normal and infarcted rat hearts by Yang and coworkers, who also reported upregulation of miR-1 in individuals with coronary heart disease [132]. Arrythmias are common after heart attack and decreasing miR-1 levels using an antisense oligonucleotide in rat infarct model reverses predisposition to arrythmias by upregulation of ion channel genes, KCNJ2 and GJA1 [132]. Recent studies have focused on using miR-1 as therapeutic target for the prevention of ischemic arrhythmias [74, 101]. These studies analyzed whether the beneficial effects of beta-blockers and tanshinone IIA in reducing ischemic arrhythmias and cardiac mortality are related to downregulation of miR-1. miR-208a has also been shown to be sufficient to induce arrhythmias in mice [12].
Coronary Angioplasty
Percutaneous coronary intervention (PCI) is a key therapeutic tool for the treatment of various forms of ischemic heart diseases. Though a successful technique, PCI yields the risk of restenosis, i.e., the recurrence of obstruction in the treated segment and efforts are ongoing to overcome this phenomenon. Ji and coworkers [49] demonstrated for the first time aberrant expression of miRNAs in the vascular walls after carotid artery balloon angioplasty in rats. They showed that miR-21 knockdown increased cell apoptosis and induced significant negative effect on neointimal lesion formation and identified PTEN and Bcl-2, two pro-apoptotic proteins as targets for miR-21. Although some of these observed effects of miR-21 may depend on the culture conditions of vascular smooth muscle cells [104], these data suggest that miRNAs could be potential targets for anti-restenotic therapies. More recently, the same group has indicated miR-221 and miR-222 as additional regulators for vascular smooth muscle cell proliferation and neointimal hyperplasia [71]. Using both gain-of-function and loss-of-function approaches, they identified p27 (Kip1) and p57 (Kip2) as potential targets for these miRNAs in these cells. Further by knocking down miR-221 and miR-222 in rat carotid arteries, they demonstrated therapeutic potential for these miRNAs in suppressing cell proliferation in vivo and neointimal lesion formation after angioplasty.
Ischemia/Reperfusion
Yet another risk associated with PCI is reperfusion injury. The rapid restoration of blood flow following coronary intervention often results in significant reperfusion injury to the myocardium and microvasculature. Following reperfusion, there is an immediate rise in oxygen tension leading to the production of free radicals from the electron transport chain and other sources that in many cases cause irreversible damage to cell membranes. In addition, the initiation of apoptosis contributes to progressive loss of myocytes and adverse left ventricular remodeling. The molecular mechanisms that regulate gene expression during myocardial ischemia/reperfusion (I/R) are not clearly understood.
Muscle-specific miR-1 was found to be upregulated in response to I/R injury in rat heart [111] and in a rat model of MI [102, 132]. miR-1 has been shown to elicit pro-apoptotic responses by post-transcriptional repression of HSP60 and HSP70 in cardiomyocytes induced by oxidative stress [131]. miR-1 is involved in apoptotic cell death induced by cardiac ischemia through the post-transcriptional repression of the anti-apoptotic proteins Bcl2 and IGF-1 [102, 111]. Recently miR-320 was shown to be involved in the regulation of I/R-induced cardiac injury and dysfunction via targeting heat shock protein 20 [95]. In this study, they performed miRNA arrays to detect miRNA expression in murine hearts subjected to I/R in vivo and ex vivo and found miR-320 to be the only miRNA that is significantly downregulated. Gain-of-function and loss-of-function studies confirmed the role of miR-320 in reducing infarct size by preventing apoptosis and thus constitute a new therapeutic target for ischemic heart diseases [95].
It has been shown that preconditioning hearts to stresses such as sub-lethal ischemia or mild heat shock improves myocardial survival after subsequent prolonged I/R injury [24, 45, 79]. Recently, efforts have been made to understand the mechanisms underlying preconditioning at transcriptional level. Roy and coworkers [96] provided the first evidence for the global changes in miRNA expression following I/R in murine heart. They characterized miRNA expression profiles following 2 and 7 days of I/R. Their analyses revealed 13 miRs to be upregulated on day 2 post-I/R, nine miRs to be upregulated on day 7, and six miRs to be uniquely downregulated on day 7 post-I/R. Furthermore, using tissues from infarct site, captured by laser micro dissection and pressure catapulting, they showed that miR-21, which was rapidly induced in cardiac fibroblasts following I/R, regulates MMP2 expression via PTEN pathway. The significance of miR-21 in influencing myocardial infarction outcomes was also pointed out in two independent studies [27, 135]. The first study was using mice subjected to cytoprotective heat shock [135]. Here the authors showed reduction in infarct size following injection of chemically synthesized exogenous miR-21 whereas co-treatment with a miR-21 inhibitor abolished the effect. They observed downregulation of pro-apoptotic proteins suggesting a potential role of miRNAs in reducing MI through repression of apoptosis. The same group, in yet another study showed that ischemic preconditioning induced miRNAs that trigger cardioprotection (viz., miR-1, miR-21, and miR-24), possibly through upregulating eNOS, HSP70, and heat shock transcription factor, HSF-1, that are implicated in the delayed phase of ischemic preconditioning in the heart [134].
The second group examined miRNA expression signature in the early phase of acute myocardial infarction (AMI) [27] in rat hearts 6 h after AMI. They observed 38 miRNAs to be differentially expressed in infarcted areas and 33 miRNAs to be aberrantly expressed in the border areas as compared to the expression signature in the noninfarcted areas. Remarkably, miR-21 expression was found to be significantly downregulated in infarcted areas, but upregulated in border areas. The downregulation of miR-21 in the infarcted areas was inhibited by ischemic preconditioning. Furthermore, overexpression of miR-21, in rat models of AMI using adenovirus expressing miR-21, decreased myocardial infarct size by 29% at 24 h after AMI. Using both gain-of-function and loss-of-function approaches in cultured cardiac myocytes, they identified programmed cell death 4 and activator protein 1 pathway to be targeted by miR-21 to provide the protective effect on ischemia-induced cell apoptosis.
Ischemic preconditioning was also attempted with two cycles of 30-min ischemia/re-oxygenation in bone-marrow-derived mesenchymal stem cells (MSCs) [53]. In this study, the authors observed that preconditioning significantly reduced apoptosis in MSCs through nuclear translocation of HIF-1α and concomitant induction of miR-210. Notably, multiple I/R cycles regulated miR-210 more effectively and hence promoted MSCs survival as compared with single cycle hypoxia of an equal duration. Using a rat model of acute myocardial infarction, they further showed that preconditioning induced miR-210 plays a key role in bone-marrow-derived MSC survival by regulating FLICE-associated huge protein/caspase-8-associated protein-2 (Casp8ap2) which has a regulatory role in Fas-mediated apoptosis.
Angiogenesis
One of the endogenous mechanisms for efficient repair following MI is through the formation of new blood vessels that provide collateral circulation. Myocardial vascularization requires signaling by angiogenic growth factors like vascular endothelial growth factor (VEGF) and fibroblast growth factor (FGF) [99, 108]. miRNAs have been shown to participate in angiogenesis and vascular development (reviewed in [57, 117]). The role of miRNAs in angiogenesis was demonstrated by inducing silencing of two key enzymes Dicer and Drosha [58] in murine endothelial cells. Genetic silencing of Dicer and Drosha significantly reduced capillary sprouting of endothelial cells and tube forming activity in vitro. However, silencing of Dicer but not of Drosha significantly decreased migration of endothelial cells and reduced angiogenesis in vivo. This study further showed that silencing of these enzymes caused reduction of endothelial let-7f and miR-27b which promote angiogenesis by targeting antiangiogenic genes like thrombospondin-1. miR-17-92 cluster is another family that is highly expressed in human endothelial cells [9]. miR-92a, a component of this cluster, has been shown to control angiogenesis. Overexpression of miR-92a in endothelial cells blocked angiogenesis in vitro and in vivo while systemic administration of antagomir designed to inhibit miR-92a in mouse models of limb ischemia and MI, led to enhanced blood vessel growth and functional recovery of damaged tissue. Several proangiogenic proteins, including the integrin subunit alpha5, are potentially targeted by miR-92a. These studies reflect miR-92a as another valuable therapeutic target in the setting of ischemic disease.
miR-126 is another positive regulator of angiogenic signaling in endothelial cells and has an essential role in neoangiogenesis following MI and in maintenance of vascular integrity in vivo [35, 123, 126]. Following coronary ligation, a model for MI, miR-126 mutant mice showed reduced vascularization of infarcted myocardium, more extensive fibrosis and loss of functional myocardium and were more prone to cardiac rupture. miR-126 enhances the proangiogenic actions of VEGF and FGF and promotes blood vessel formation by repressing the expression of Spred-1 [35, 126] and PIK3R2 [35]. Both Spred-1 and PIK3R2 are intracellular inhibitors of angiogenic signaling by acting as negative regulators of MAP kinase and phosphatidylinositol 3-kinase pathways, respectively. These findings suggest that strategies to elevate endothelial specific miR-126 expression to promote new blood vessel formation in ischemic myocardium can enhance cardiac repair.
The role of miRNAs in regulating endothelial cells in response to hypoxia was studied by Fasanaro and coworkers [34]. They observed miR-210 upregulation in human endothelial cells in response to hypoxia affecting cell survival, migration, and differentiation. miR-210 overexpression in normoxic endothelial cells stimulated angiogenesis while the converse was observed with miR-210 blockade. Ephrin A3, a known regulator of angiogenesis and VEGF signaling, was identified as a target for miR-210. In another study, a Dicer knockdown approach was used to test the significance of miRNA in regulating the redox state and angiogenic response of human microvascular endothelial cells [103]. Lowering of miRNA content induced vascular endothelial growth factor expression but diminished the angiogenic response of endothelial cells. The reduced miRNA levels induced expression of the transcription factor HBP1 that negatively regulates expression of p47phox of the NADPH oxidase complex. This study provides the first evidence that redox signaling in cells is subjected to regulation by miRNA. Recently, miR-217 was identified in human atherosclerotic lesions and was shown to induce premature senescence-like phenotype, leading to impairment in angiogenesis [84]. These actions were mediated by inhibition of silent information regulator SIRT1 and modulation of FOXO1 (forkhead box O1) and endothelial nitric oxide synthase acetylation. These studies show miR-217 as potentially amenable to therapeutic manipulation for prevention of endothelial dysfunction in metabolic disorders.
miRNA-Based Strategies in the Clinics
The multitude of research in the field of cardiovascular biology clearly emphasizes the profound role played by miRNAs in the etiology and progression of cardiovascular diseases. These in turn highlight the potential of miRNAs as targets for development of diagnostic and prognostic biomarkers and/or for therapeutic purposes. Through their ability to regulate mRNA levels post-transcriptionally, miRNAs bring about finer regulation of the protein factors that regulate cardiogenesis. This is especially significant in congenital heart defects that results from haploinsufficiency or heterozygous point mutations [78, 92]. By fine tuning target protein levels, miRNA can modulate critical pathways and allow restoration of protein levels to a point that crosses the disease threshold. In addition, with miRNAs implicated in cardiac biology, mutations in miRNA genes or their target sequences can correlate to genetic disorders as exemplified by a study where a SNP was identified in the 3′UTR of myostatin transcript in Texel sheep that created an aberrant miR-1 target site [20]. With the advent of high-throughput sequencing of whole genomes, it will be possible to decode allelic mutations in miRNA genes and/or their target sites associated with congenital defects.
As a Diagnostic Tool
As discussed above, many diseases are characterized by an abnormal miRNA expression pattern. Identification of miRNA signature for specific diseases will be useful in early and differential diagnosis of diseases characterized by abnormal miRNA levels in tissues or in plasma. For instance, we have showed earlier that unique miRNA expression patterns in sarcoma can help identify misdiagnosed cases [106]. There has been increasing evidence suggesting the potential utilization of circulating miRNAs in various body fluids as biomarkers for disease diagnosis [16, 19, 23, 38, 44, 62, 85, 87, 124]. For instance, measurement of serum levels of miR-92 and miR-141 has been suggested for screening for colorectal and prostate cancer, respectively [85, 87]. In addition, miR-141 has also been reported as a pregnancy associated marker [19]. The potential of miRNAs as biomarkers for cardiovascular diseases is only starting to be explored and will be an active field of research for the next few years. Because of their abundance and relative stability in plasma, miRNAs can serve as easily accessible markers to monitor tissue injury. Due to their small size and tissue specificity, miRNAs can leak into the circulating plasma from injured cells and thereby aid in identifying the injured cell type. Recently, miR-208 was shown to be a useful indicator of myocardial injury [50]. The authors used an isoproterenol-induced myocardial injury model to confirm whether cardiac-specific miRNAs leak out from injured cardiomyocytes in response to myocardial injury. The plasma concentration of miR-208 in myocardial injury model correlated to the concentration of cardiac troponin 1 (cTn1). Circulating cTn1 is a validated marker of isoproterenol-induced myocardial injury [136]. However, cTn1 concentrations are commonly increased in end-stage renal disease, even without acute coronary syndrome [1]. Hence in this study [50], the authors assessed miR-208 levels in a renal-infarction model and observed no detectable levels of this miRNA in this model. Thus, miRNA-based markers maybe superior to the currently available markers.
miR-1 has been earlier shown to exhibit aberrant upregulation in ischemic myocardium [132]. By using blood samples from 159 patients, and using real-time RT-PCR technique for quantifying miRNAs in plasma, the same group has now identified circulating miR-1 as a potential independent biomarker for acute myocardial infarction [2]. They found significantly higher miR-1 levels in plasma from AMI patients compared to non-AMI subjects. These elevated levels of miR-1 dropped to normal by the time of patient discharge following medication. Thus, miR-1 can be a useful novel marker for the diagnosis and progression of AMI and its response to treatment. These studies shed light on the potential of miRNAs in assessing risk for developing a disease and also for early diagnosis and to evaluate the efficacy of treatment.
Efforts are also underway to understand dynamic regulation of miRNA during heart failure and treatment [82, 100]. Matkovich and coworkers analyzed miRNA and mRNA profiles in patients with or without biomechanical support from left ventricular assist devices [82]. They observed all the 28 miRNAs that were deregulated in failing heart to get normalized in patients undergoing treatment. However, of the 444 altered mRNAs, only 29 were normalized by as much as 25% in these failing hearts, which were on support device. Based on their studies, the authors suggest miRNAs to be more sensitive than mRNAs to the actual functional status of end-stage heart failure [82]. With the current knowledge on miRNA expression in heart, future studies can be aimed at developing miRNAs as diagnostic and prognostic markers.
As a Therapeutic Tool
miRNAs are also useful as targets for therapeutic applications mainly due to a few salient properties. For example, miRNAs exhibit signature expression patterns in various diseases. This will aid in identifying specific miRNAs as targets that can be efficiently manipulated to fine tune gene regulation. Another factor that contributes to the therapeutic property of miRNA is their small size which makes their in vivo delivery feasible [32, 56]. One other interesting and most significant advantage of targeting miRNAs is due to the fact that miRNAs are predicted to have multiple mRNA targets [6], some of which seem to work in concert to control a common pathway or biological function. However, this can also be a major disadvantage in that it can bring about “off-target” side effects. For instance, miR-133 is significantly downregulated in cardiac hypertrophy and thus increasing miR-133 levels by introducing synthetic miR-133 can potentially control pathological hypertrophy [14]. However, an increased level of miR-133 induces cardiac arrhythmias [130]. Thus, it is important to understand miRNA biology and function pertaining to the heart in detail before considering it for clinical applications.
miRNA levels are either upregulated or downregulated in different diseases. Hence miRNAs need to be finely modulated so as to either knockdown pathogenic or aberrantly expressed miRNAs or induce expression of beneficial miRNAs to threshold levels. There are several approaches to achieve these in vivo and in vitro as has been exemplified by the several gain-of-function and loss-of-function studies outlined in this review.
miRNA Mimics
One of the methods to upregulate cellular levels of specific miRNAs is by using short double-stranded oligonucleotides which serve as mimics. In this, one strand serves as the guide and is the mature miRNA sequence while the complementary strand is the passenger (Fig. 1) and is complexed with the mature miRNA sequence [80]. The double-stranded structure enables efficient recognition and loading into the RISC. In order to ensure incorporation of only the guide strand and to reduce off-target silencing, position-specific chemical modifications has been suggested [48]. These constructs are analogous to siRNA molecules and are successfully utilized in vitro, though, to date, there are no reports of its efficacy in vivo.
AntagomiRs
In scenarios where a miRNA is causal to disease state, efficient reduction of the specific miRNA would be therapeutically advantageous. This can be achieved by using chemically modified single-stranded reverse complement oligonucleotides called antagomirs. They function either as competitive inhibitors by binding to mature miRNA within RISC or they can bind to pre-miRNA and prevent its processing and entry into RISC or further upstream by interfering with the processing and export of pre- or pri-miRNA from the nucleus. The first successful mammalian in vivo study using antagomiRs was reported by Krutzfeldt and co workers [56], where they inhibited a liver-specific miR-122. Here they used 2′-O-methyl modified cholesterol conjugated oligos to facilitate cellular uptake. Further studies attempted modifying cellular uptake by using high-density lipoproteins [129], MOE (2′-O-methoxyethyl phosphorothioate) [32] and also using locked-nucleic-acid-modified oligonucleotides [31], which has also been tested in non-human primates [30]. With respect to heart, in vivo inhibition in mice of miR-133 and miR-29 with antagomiRs were performed to implicate their roles in cardiac hypertrophy and cardiac fibrosis, respectively [14, 122].
Sponges and Erasers
In addition to modulating specific miRNAs, several approaches to interfere miRNA-target mRNA interactions have been developed. One way of modifying miRNA function is by scavenging out specific miRNA and thereby preventing it from interacting with its target mRNA. An expression vector based approach was reported for miRNA sponging wherein a series of perfectly and imperfectly paired tandem binding sites of a miRNA of interest are included in the 3′UTR of a reporter gene construct [29]. The multiplexed binding sites serve as competitive inhibitors and help in shutting off miRNA families, since a single sponge can block all miRNAs that recognize same sequence. In vivo, miRNA sponges has been used to inhibit breast cancer metastasis by specifically inhibiting miR-31 [118]. miRNA erasers are comparable to sponges; however, erasers use only two copies of the perfectly complementary antisense sequence of the miRNA [98]. For instance, Sprouty 2 was demonstrated to be the target for miR-21 using miR-21 eraser approach [98]. Though these approaches have been proposed, their in vivo efficacy needs to be tested. However, recently this approach was tested in vivo in Drosophila to understand the function of miRNAs at spatiotemporal resolution [72].
Future Directions
A major challenge in the advancement of miRNA-based therapeutics is concerning its systemic delivery into targeted sites, which can be specific organs or specific cell types within an organ, as in the heart. Not much is known regarding the pharmacokinetics and tissue distribution of these synthetic small RNA molecules. Native nucleic acids are rapidly degraded in blood by endogenous nucleases and phosphodiesterases. This has been overcome by several chemical modifications, but these are mostly demonstrated only for liver. Moreover, there is always the risk of over-saturating endogenous miRNA pathways thereby causing deregulation of key miRNAs leading to undesired effects [40]. Hence further work needs to be done to improve efficacy of the available delivery systems. Cues can be drawn from RNAi-therapeutics which has striking similarities with miRNA-based approach. Recently, there has been a report on targeted RNAi for the treatment of heart failure [107]. The authors report a successful treatment strategy for heart failure in a rat model of transaortic banding by RNAi targeting a key regulator of cardiac Ca (2+) homeostasis, phospholamban. One encouraging observation was that there was no evidence for miRNA deregulation or hepatotoxicity during these long-term RNAi therapies in the heart.
To conclude, there is a definite need for more mechanistic studies on miRNA-based therapeutics for a proper transition of knowledge from bench to bedside. With the advances in the understanding of the biology and function of miRNA and the horizontal flow of information from cancer and other infectious diseases to cardiovascular field, the transition to miRNA-based therapies is in the foreseeable future.
References
Abbas, N. A., John, R. I., Webb, M. C., Kempson, M. E., Potter, A. N., Price, C. P., et al. (2005). Cardiac troponins and renal function in nondialysis patients with chronic kidney disease. Clinical Chemistry, 51, 2059–2066.
Ai, J., Zhang, R., Li, Y., Pu, J., Lu, Y., Jiao, J., et al. (2010). Circulating microRNA-1 as a potential novel biomarker for acute myocardial infarction. Biochemical and Biophysical Research Communications, 391, 73–77.
Altuvia, Y., Landgraf, P., Lithwick, G., Elefant, N., Pfeffer, S., Aravin, A., et al. (2005). Clustering and conservation patterns of human microRNAs. Nucleic Acids Research, 33, 2697–2706.
Asangani, I. A., Rasheed, S. A., Nikolova, D. A., Leupold, J. H., Colburn, N. H., Post, S., et al. (2008). MicroRNA-21 (miR-21) post-transcriptionally downregulates tumor suppressor Pdcd4 and stimulates invasion, intravasation and metastasis in colorectal cancer. Oncogene, 27, 2128–2136.
Barringhaus, K. G., & Zamore, P. D. (2009). MicroRNAs: Regulating a change of heart. Circulation, 119, 2217–2224.
Bartel, D. P. (2009). MicroRNAs: Target recognition and regulatory functions. Cell, 136, 215–233.
Beltrami, A. P., Urbanek, K., Kajstura, J., Yan, S. M., Finato, N., Bussani, R., et al. (2001). Evidence that human cardiac myocytes divide after myocardial infarction. New England Journal of Medicine, 344, 1750–1757.
Bernstein, E., Kim, S. Y., Carmell, M. A., Murchison, E. P., Alcorn, H., Li, M. Z., et al. (2003). Dicer is essential for mouse development. Nature Genetics, 35, 215–217.
Bonauer, A., Carmona, G., Iwasaki, M., Mione, M., Koyanagi, M., Fischer, A., et al. (2009). MicroRNA-92a controls angiogenesis and functional recovery of ischemic tissues in mice. Science, 324, 1710–1713.
Borchert, G. M., Lanier, W., & Davidson, B. L. (2006). RNA polymerase III transcribes human microRNAs. Nature Structural & Molecular Biology, 13, 1097–1101.
Calin, G. A., Ferracin, M., Cimmino, A., Di Leva, G., Shimizu, M., Wojcik, S. E., et al. (2005). A microRNA signature associated with prognosis and progression in chronic lymphocytic leukemia. New England Journal of Medicine, 353, 1793–1801.
Callis, T. E., Pandya, K., Seok, H. Y., Tang, R. H., Tatsuguchi, M., Huang, Z. P., et al. (2009). MicroRNA-208a is a regulator of cardiac hypertrophy and conduction in mice. Journal of Clinical Investigation, 119, 2772–2786.
Callis, T. E., & Wang, D. Z. (2008). Taking microRNAs to heart. Trends in Molecular Medicine, 14, 254–260.
Care, A., Catalucci, D., Felicetti, F., Bonci, D., Addario, A., Gallo, P., et al. (2007). MicroRNA-133 controls cardiac hypertrophy. Nature Medicine, 13, 613–618.
Chen, J. F., Mandel, E. M., Thomson, J. M., Wu, Q., Callis, T. E., Hammond, S. M., et al. (2006). The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nature Genetics, 38, 228–233.
Chen, X., Ba, Y., Ma, L., Cai, X., Yin, Y., Wang, K., et al. (2008). Characterization of microRNAs in serum: A novel class of biomarkers for diagnosis of cancer and other diseases. Cell Res, 18, 997–1006.
Cheng, Y., Ji, R., Yue, J., Yang, J., Liu, X., Chen, H., et al. (2007). MicroRNAs are aberrantly expressed in hypertrophic heart: Do they play a role in cardiac hypertrophy? American Journal of Pathology, 170, 1831–1840.
Cheng, Y., Liu, X., Yang, J., Lin, Y., Xu, D. Z., Lu, Q., et al. (2009). MicroRNA-145, a novel smooth muscle cell phenotypic marker and modulator, controls vascular neointimal lesion formation. Circulation Research, 105, 158–166.
Chim, S. S., Shing, T. K., Hung, E. C., Leung, T. Y., Lau, T. K., Chiu, R. W., et al. (2008). Detection and characterization of placental microRNAs in maternal plasma. Clinical Chemistry, 54, 482–490.
Clop, A., Marcq, F., Takeda, H., Pirottin, D., Tordoir, X., Bibe, B., et al. (2006). A mutation creating a potential illegitimate microRNA target site in the myostatin gene affects muscularity in sheep. Nature Genetics, 38, 813–818.
Cordes, K. R., Sheehy, N. T., White, M. P., Berry, E. C., Morton, S. U., Muth, A. N., et al. (2009). miR-145 and miR-143 regulate smooth muscle cell fate and plasticity. Nature, 460, 705–710.
Cordes, K. R., & Srivastava, D. (2009). MicroRNA regulation of cardiovascular development. Circulation Research, 104, 724–732.
Cortez, M. A., & Calin, G. A. (2009). MicroRNA identification in plasma and serum: A new tool to diagnose and monitor diseases. Expert Opinion on Biol Ther, 9, 703–711.
Currie, R. W., Tanguay, R. M., & Kingma, J. G., Jr. (1993). Heat-shock response and limitation of tissue necrosis during occlusion/reperfusion in rabbit hearts. Circulation, 87, 963–971.
da Costa Martins, P. A., Bourajjaj, M., Gladka, M., Kortland, M., van Oort, R. J., Pinto, Y. M., et al. (2008). Conditional dicer gene deletion in the postnatal myocardium provokes spontaneous cardiac remodeling. Circulation, 118, 1567–1576.
Divakaran, V., & Mann, D. L. (2008). The emerging role of microRNAs in cardiac remodeling and heart failure. Circulation Research, 103, 1072–1083.
Dong, S., Cheng, Y., Yang, J., Li, J., Liu, X., Wang, X., et al. (2009). MicroRNA expression signature and the role of microRNA-21 in the early phase of acute myocardial infarction. Journal of Biological Chemistry, 284, 29514–29525.
Duisters, R. F., Tijsen, A. J., Schroen, B., Leenders, J. J., Lentink, V., van der Made, I., et al. (2009). miR-133 and miR-30 regulate connective tissue growth factor: Implications for a role of microRNAs in myocardial matrix remodeling. Circ Res, 104, 170–178. 176p following 178.
Ebert, M. S., Neilson, J. R., & Sharp, P. A. (2007). MicroRNA sponges: Competitive inhibitors of small RNAs in mammalian cells. Nat Methods, 4, 721–726.
Elmen, J., Lindow, M., Schutz, S., Lawrence, M., Petri, A., Obad, S., et al. (2008). LNA-mediated microRNA silencing in non-human primates. Nature, 452, 896–899.
Elmen, J., Lindow, M., Silahtaroglu, A., Bak, M., Christensen, M., Lind-Thomsen, A., et al. (2008). Antagonism of microRNA-122 in mice by systemically administered LNA-antimiR leads to up-regulation of a large set of predicted target mRNAs in the liver. Nucleic Acids Research, 36, 1153–1162.
Esau, C., Davis, S., Murray, S. F., Yu, X. X., Pandey, S. K., Pear, M., et al. (2006). miR-122 regulation of lipid metabolism revealed by in vivo antisense targeting. Cell Metab, 3, 87–98.
Eulalio, A., Huntzinger, E., & Izaurralde, E. (2008). Getting to the root of miRNA-mediated gene silencing. Cell, 132, 9–14.
Fasanaro, P., D'Alessandra, Y., Di Stefano, V., Melchionna, R., Romani, S., Pompilio, G., et al. (2008). MicroRNA-210 modulates endothelial cell response to hypoxia and inhibits the receptor tyrosine kinase ligand Ephrin-A3. Journal of Biological Chemistry, 283, 15878–15883.
Fish, J. E., Santoro, M. M., Morton, S. U., Yu, S., Yeh, R. F., Wythe, J. D., et al. (2008). miR-126 regulates angiogenic signaling and vascular integrity. Developments in Cell, 15, 272–284.
Frey, N., & Olson, E. N. (2003). Cardiac hypertrophy: The good, the bad, and the ugly. Annual Review of Physiology, 65, 45–79.
Friedman, R. C., Farh, K. K., Burge, C. B., & Bartel, D. P. (2009). Most mammalian mRNAs are conserved targets of microRNAs. Genome Research, 19, 92–105.
Gilad, S., Meiri, E., Yogev, Y., Benjamin, S., Lebanony, D., Yerushalmi, N., et al. (2008). Serum microRNAs are promising novel biomarkers. PLoS ONE, 3, e3148.
Griffiths-Jones, S., Saini, H. K., van Dongen, S., & Enright, A. J. (2008). miRBase: Tools for microRNA genomics. Nucleic Acids Research, 36, D154–D158.
Grimm, D., Streetz, K. L., Jopling, C. L., Storm, T. A., Pandey, K., Davis, C. R., et al. (2006). Fatality in mice due to oversaturation of cellular microRNA/short hairpin RNA pathways. Nature, 441, 537–541.
Hoffman, J. I. (1995). Incidence of congenital heart disease: II. Prenatal incidence. Pediatric Cardiology, 16, 155–165.
Horie, T., Ono, K., Nishi, H., Iwanaga, Y., Nagao, K., Kinoshita, M., et al. (2009). MicroRNA-133 regulates the expression of GLUT4 by targeting KLF15 and is involved in metabolic control in cardiac myocytes. Biochemical and Biophysical Research Communications, 389, 315–320.
Hunter, J. J., & Chien, K. R. (1999). Signaling pathways for cardiac hypertrophy and failure. New England Journal of Medicine, 341, 1276–1283.
Hunter, M. P., Ismail, N., Zhang, X., Aguda, B. D., Lee, E. J., Yu, L., et al. (2008). Detection of microRNA expression in human peripheral blood microvesicles. PLoS ONE, 3, e3694.
Hutter, M. M., Sievers, R. E., Barbosa, V., & Wolfe, C. L. (1994). Heat-shock protein induction in rat hearts. A direct correlation between the amount of heat-shock protein induced and the degree of myocardial protection. Circulation, 89, 355–360.
Ikeda, S., Kong, S. W., Lu, J., Bisping, E., Zhang, H., Allen, P. D., et al. (2007). Altered microRNA expression in human heart disease. Physiological Genomics, 31, 367–373.
Ivey, K. N., Muth, A., Arnold, J., King, F. W., Yeh, R. F., Fish, J. E., et al. (2008). MicroRNA regulation of cell lineages in mouse and human embryonic stem cells. Cell Stem Cell, 2, 219–229.
Jackson, A. L., Burchard, J., Leake, D., Reynolds, A., Schelter, J., Guo, J., et al. (2006). Position-specific chemical modification of siRNAs reduces “off-target” transcript silencing. RNA, 12, 1197–1205.
Ji, R., Cheng, Y., Yue, J., Yang, J., Liu, X., Chen, H., et al. (2007). MicroRNA expression signature and antisense-mediated depletion reveal an essential role of microRNA in vascular neointimal lesion formation. Circulation Research, 100, 1579–1588.
Ji, X., Takahashi, R., Hiura, Y., Hirokawa, G., Fukushima, Y., & Iwai, N. (2009). Plasma miR-208 as a biomarker of myocardial injury. Clinical Chemistry, 55, 1944–1949.
Johnson, S. M., Grosshans, H., Shingara, J., Byrom, M., Jarvis, R., Cheng, A., et al. (2005). RAS is regulated by the let-7 microRNA family. Cell, 120, 635–647.
Kajstura, J., Urbanek, K., Rota, M., Bearzi, C., Hosoda, T., Bolli, R., et al. (2008). Cardiac stem cells and myocardial disease. Journal of Molecular and Cellular Cardiology, 45, 505–513.
Kim, H. W., Haider, H. K., Jiang, S., & Ashraf, M. (2009). Ischemic preconditioning augments survival of stem cells via miR-210 expression by targeting caspase-8 associated protein 2. Journal of Biological Chemistry, 284, 33161–33168.
Kloosterman, W. P., Wienholds, E., Ketting, R. F., & Plasterk, R. H. (2004). Substrate requirements for let-7 function in the developing zebrafish embryo. Nucleic Acids Research, 32, 6284–6291.
Kocher, A. A., Schuster, M. D., Szabolcs, M. J., Takuma, S., Burkhoff, D., Wang, J., et al. (2001). Neovascularization of ischemic myocardium by human bone-marrow-derived angioblasts prevents cardiomyocyte apoptosis, reduces remodeling and improves cardiac function. Nature Medicine, 7, 430–436.
Krutzfeldt, J., Rajewsky, N., Braich, R., Rajeev, K. G., Tuschl, T., Manoharan, M., et al. (2005). Silencing of microRNAs in vivo with ‘antagomirs’. Nature, 438, 685–689.
Kuehbacher, A., Urbich, C., & Dimmeler, S. (2008). Targeting microRNA expression to regulate angiogenesis. Trends in Pharmacological Sciences, 29, 12–15.
Kuehbacher, A., Urbich, C., Zeiher, A. M., & Dimmeler, S. (2007). Role of Dicer and Drosha for endothelial microRNA expression and angiogenesis. Circulation Research, 101, 59–68.
Kwon, C., Han, Z., Olson, E. N., & Srivastava, D. (2005). MicroRNA1 influences cardiac differentiation in Drosophila and regulates Notch signaling. Proceedings of the National Academy of Sciences of the United States of America, 102, 18986–18991.
Lagos-Quintana, M., Rauhut, R., Yalcin, A., Meyer, J., Lendeckel, W., & Tuschl, T. (2002). Identification of tissue-specific microRNAs from mouse. Current Biology, 12, 735–739.
Landgraf, P., Rusu, M., Sheridan, R., Sewer, A., Iovino, N., Aravin, A., et al. (2007). A mammalian microRNA expression atlas based on small RNA library sequencing. Cell, 129, 1401–1414.
Laterza, O. F., Lim, L., Garrett-Engele, P. W., Vlasakova, K., Muniappa, N., Tanaka, W. K., et al. (2009). Plasma MicroRNAs as sensitive and specific biomarkers of tissue injury. Clinical Chemistry, 55, 1977–1983.
Latronico, M. V., Catalucci, D., & Condorelli, G. (2007). Emerging role of microRNAs in cardiovascular biology. Circulation Research, 101, 1225–1236.
Latronico, M. V., & Condorelli, G. (2009). MicroRNAs and cardiac pathology. Nature Reviews Cardiol, 6, 419–429.
Lee, I., Ajay, S. S., Yook, J. I., Kim, H. S., Hong, S. H., Kim, N. H., et al. (2009). New class of microRNA targets containing simultaneous 5′-UTR and 3′-UTR interaction sites. Genome Res, 19, 1175–1183.
Lee, R. C., Feinbaum, R. L., & Ambros, V. (1993). The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell, 75, 843–854.
Lee, Y., Kim, M., Han, J., Yeom, K. H., Lee, S., Baek, S. H., et al. (2004). MicroRNA genes are transcribed by RNA polymerase II. EMBO Journal, 23, 4051–4060.
Lewis, B. P., Burge, C. B., & Bartel, D. P. (2005). Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell, 120, 15–20.
Liu, J., Valencia-Sanchez, M. A., Hannon, G. J., & Parker, R. (2005). MicroRNA-dependent localization of targeted mRNAs to mammalian P-bodies. Nature Cell Biology, 7, 719–723.
Liu, N., Williams, A. H., Kim, Y., McAnally, J., Bezprozvannaya, S., Sutherland, L. B., et al. (2007). An intragenic MEF2-dependent enhancer directs muscle-specific expression of microRNAs 1 and 133. Proceedings of the National Academy of Sciences of the United States of America, 104, 20844–20849.
Liu, X., Cheng, Y., Zhang, S., Lin, Y., Yang, J., & Zhang, C. (2009). A necessary role of miR-221 and miR-222 in vascular smooth muscle cell proliferation and neointimal hyperplasia. Circulation Research, 104, 476–487.
Loya, C. M., Lu, C. S., Van Vactor, D., & Fulga, T. A. (2009). Transgenic microRNA inhibition with spatiotemporal specificity in intact organisms. Nat Methods, 6, 897–903.
Lu, J., Getz, G., Miska, E. A., Alvarez-Saavedra, E., Lamb, J., Peck, D., et al. (2005). MicroRNA expression profiles classify human cancers. Nature, 435, 834–838.
Lu, Y., Zhang, Y., Shan, H., Pan, Z., Li, X., Li, B., et al. (2009). MicroRNA-1 downregulation by propranolol in a rat model of myocardial infarction: A new mechanism for ischaemic cardioprotection. Cardiovascular Research, 84, 434–441.
Luo, X., Lin, H., Pan, Z., Xiao, J., Zhang, Y., Lu, Y., et al. (2008). Down-regulation of miR-1/miR-133 contributes to re-expression of pacemaker channel genes HCN2 and HCN4 in hypertrophic heart. Journal of Biological Chemistry, 283, 20045–20052.
Luo, X., Xiao, J., Lin, H., Li, B., Lu, Y., Yang, B., et al. (2007). Transcriptional activation by stimulating protein 1 and post-transcriptional repression by muscle-specific microRNAs of IKs-encoding genes and potential implications in regional heterogeneity of their expressions. Journal of Cellular Physiology, 212, 358–367.
Lytle, J. R., Yario, T. A., & Steitz, J. A. (2007). Target mRNAs are repressed as efficiently by microRNA-binding sites in the 5′ UTR as in the 3′ UTR. Proceedings of the National Academy of Sciences of the United States of America, 104, 9667–9672.
Mandel, E. M., Callis, T. E., Wang, D. Z., & Conlon, F. L. (2005). Transcriptional mechanisms of congenital heart disease. Drug Discovery Today, 2, 33–38.
Marber, M. S., Latchman, D. S., Walker, J. M., & Yellon, D. M. (1993). Cardiac stress protein elevation 24 hours after brief ischemia or heat stress is associated with resistance to myocardial infarction. Circulation, 88, 1264–1272.
Martinez, J., Patkaniowska, A., Urlaub, H., Luhrmann, R., & Tuschl, T. (2002). Single-stranded antisense siRNAs guide target RNA cleavage in RNAi. Cell, 110, 563–574.
Mathonnet, G., Fabian, M. R., Svitkin, Y. V., Parsyan, A., Huck, L., Murata, T., et al. (2007). MicroRNA inhibition of translation initiation in vitro by targeting the cap-binding complex eIF4F. Science, 317, 1764–1767.
Matkovich, S. J., Van Booven, D. J., Youker, K. A., Torre-Amione, G., Diwan, A., Eschenbacher, W. H., et al. (2009). Reciprocal regulation of myocardial microRNAs and messenger RNA in human cardiomyopathy and reversal of the microRNA signature by biomechanical support. Circulation, 119, 1263–1271.
Meng, F., Henson, R., Wehbe-Janek, H., Ghoshal, K., Jacob, S. T., & Patel, T. (2007). MicroRNA-21 regulates expression of the PTEN tumor suppressor gene in human hepatocellular cancer. Gastroenterology, 133, 647–658.
Menghini, R., Casagrande, V., Cardellini, M., Martelli, E., Terrinoni, A., Amati, F., et al. (2009). MicroRNA-217 modulates endothelial cell senescence via silent information regulator 1. Circulation, 120, 1524–1532.
Mitchell, P. S., Parkin, R. K., Kroh, E. M., Fritz, B. R., Wyman, S. K., Pogosova-Agadjanyan, E. L., et al. (2008). Circulating microRNAs as stable blood-based markers for cancer detection. Proceedings of the National Academy of Sciences of the United States of America, 105, 10513–10518.
Naga Prasad, S. V., Duan, Z. H., Gupta, M. K., Surampudi, V. S., Volinia, S., Calin, G. A., et al. (2009). Unique microRNA profile in end-stage heart failure indicates alterations in specific cardiovascular signaling networks. Journal of Biological Chemistry, 284, 27487–27499.
Ng, E. K., Chong, W. W., Jin, H., Lam, E. K., Shin, V. Y., Yu, J., et al. (2009). Differential expression of microRNAs in plasma of patients with colorectal cancer: A potential marker for colorectal cancer screening. Gut, 58, 1375–1381.
Petersen, C. P., Bordeleau, M. E., Pelletier, J., & Sharp, P. A. (2006). Short RNAs repress translation after initiation in mammalian cells. Molecular Cell, 21, 533–542.
Pillai, R. S., Bhattacharyya, S. N., Artus, C. G., Zoller, T., Cougot, N., Basyuk, E., et al. (2005). Inhibition of translational initiation by Let-7 MicroRNA in human cells. Science, 309, 1573–1576.
Place, R. F., Li, L. C., Pookot, D., Noonan, E. J., & Dahiya, R. (2008). MicroRNA-373 induces expression of genes with complementary promoter sequences. Proceedings of the National Academy of Sciences of the United States of America, 105, 1608–1613.
Rane, S., He, M., Sayed, D., Vashistha, H., Malhotra, A., Sadoshima, J., et al. (2009). Downregulation of miR-199a derepresses hypoxia-inducible factor-1alpha and Sirtuin 1 and recapitulates hypoxia preconditioning in cardiac myocytes. Circulation Research, 104, 879–886.
Ransom, J., & Srivastava, D. (2007). The genetics of cardiac birth defects. Seminars in Cell & Developmental Biology, 18, 132–139.
Rao, P. K., Kumar, R. M., Farkhondeh, M., Baskerville, S., & Lodish, H. F. (2006). Myogenic factors that regulate expression of muscle-specific microRNAs. Proceedings of the National Academy of Sciences of the United States of America, 103, 8721–8726.
Rao, P. K., Toyama, Y., Chiang, H. R., Gupta, S., Bauer, M., Medvid, R., et al. (2009). Loss of cardiac microRNA-mediated regulation leads to dilated cardiomyopathy and heart failure. Circulation Research, 105, 585–594.
Ren, X. P., Wu, J., Wang, X., Sartor, M. A., Qian, J., Jones, K., et al. (2009). MicroRNA-320 is involved in the regulation of cardiac ischemia/reperfusion injury by targeting heat-shock protein 20. Circulation, 119, 2357–2366.
Roy, S., Khanna, S., Hussain, S. R., Biswas, S., Azad, A., Rink, C., et al. (2009). MicroRNA expression in response to murine myocardial infarction: miR-21 regulates fibroblast metalloprotease-2 via phosphatase and tensin homologue. Cardiovascular Research, 82, 21–29.
Sayed, D., Hong, C., Chen, I. Y., Lypowy, J., & Abdellatif, M. (2007). MicroRNAs play an essential role in the development of cardiac hypertrophy. Circulation Research, 100, 416–424.
Sayed, D., Rane, S., Lypowy, J., He, M., Chen, I. Y., Vashistha, H., et al. (2008). MicroRNA-21 targets Sprouty2 and promotes cellular outgrowths. Molecular Biology of the Cell, 19, 3272–3282.
Scheinowitz, M., Abramov, D., & Eldar, M. (1997). The role of insulin-like and basic fibroblast growth factors on ischemic and infarcted myocardium: A mini review. International Journal of Cardiology, 59, 1–5.
Schipper, M. E., van Kuik, J., de Jonge, N., Dullens, H. F., & de Weger, R. A. (2008). Changes in regulatory microRNA expression in myocardium of heart failure patients on left ventricular assist device support. Journal of Heart and Lung Transplantation, 27, 1282–1285.
Shan, H., Li, X., Pan, Z., Zhang, L., Cai, B., Zhang, Y., et al. (2009). Tanshinone IIA protects against sudden cardiac death induced by lethal arrhythmias via repression of microRNA-1. British Journal of Pharmacology, 158, 1227–1235.
Shan, Z. X., Lin, Q. X., Fu, Y. H., Deng, C. Y., Zhou, Z. L., Zhu, J. N., et al. (2009). Upregulated expression of miR-1/miR-206 in a rat model of myocardial infarction. Biochemical and Biophysical Research Communications, 381, 597–601.
Shilo, S., Roy, S., Khanna, S., & Sen, C. K. (2008). Evidence for the involvement of miRNA in redox regulated angiogenic response of human microvascular endothelial cells. Arteriosclerosis, Thrombosis, and Vascular Biology, 28, 471–477.
Silvestri, P., Rigattieri, S., & Loschiavo, P. (2008). Does the effect of microRNAs in vascular neointimal formation depend on cell cycle phase? Circulation Research, 102, e101. author reply e102.
Sokol, N. S., & Ambros, V. (2005). Mesodermally expressed Drosophila microRNA-1 is regulated by Twist and is required in muscles during larval growth. Genes and Development, 19, 2343–2354.
Subramanian, S., Lui, W. O., Lee, C. H., Espinosa, I., Nielsen, T. O., Heinrich, M. C., et al. (2008). MicroRNA expression signature of human sarcomas. Oncogene, 27, 2015–2026.
Suckau, L., Fechner, H., Chemaly, E., Krohn, S., Hadri, L., Kockskamper, J., et al. (2009). Long-term cardiac-targeted RNA interference for the treatment of heart failure restores cardiac function and reduces pathological hypertrophy. Circulation, 119, 1241–1252.
Syed, I. S., Sanborn, T. A., & Rosengart, T. K. (2004). Therapeutic angiogenesis: A biologic bypass. Cardiology, 101, 131–143.
Takahashi, T., Kalka, C., Masuda, H., Chen, D., Silver, M., Kearney, M., et al. (1999). Ischemia- and cytokine-induced mobilization of bone marrow-derived endothelial progenitor cells for neovascularization. Nature Medicine, 5, 434–438.
Takaya, T., Ono, K., Kawamura, T., Takanabe, R., Kaichi, S., Morimoto, T., et al. (2009). MicroRNA-1 and MicroRNA-133 in spontaneous myocardial differentiation of mouse embryonic stem cells. Circolo J, 73, 1492–1497.
Tang, Y., Zheng, J., Sun, Y., Wu, Z., Liu, Z., & Huang, G. (2009). MicroRNA-1 regulates cardiomyocyte apoptosis by targeting Bcl-2. Int Heart J, 50, 377–387.
Tatsuguchi, M., Seok, H. Y., Callis, T. E., Thomson, J. M., Chen, J. F., Newman, M., et al. (2007). Expression of microRNAs is dynamically regulated during cardiomyocyte hypertrophy. Journal of Molecular and Cellular Cardiology, 42, 1137–1141.
Taulli, R., Bersani, F., Foglizzo, V., Linari, A., Vigna, E., Ladanyi, M., et al. (2009). The muscle-specific microRNA miR-206 blocks human rhabdomyosarcoma growth in xenotransplanted mice by promoting myogenic differentiation. Journal of Clinical Investigation, 119, 2366–2378.
Thum, T., Catalucci, D., & Bauersachs, J. (2008). MicroRNAs: Novel regulators in cardiac development and disease. Cardiovascular Research, 79, 562–570.
Thum, T., Galuppo, P., Wolf, C., Fiedler, J., Kneitz, S., van Laake, L. W., et al. (2007). MicroRNAs in the human heart: A clue to fetal gene reprogramming in heart failure. Circulation, 116, 258–267.
Thum, T., Gross, C., Fiedler, J., Fischer, T., Kissler, S., Bussen, M., et al. (2008). MicroRNA-21 contributes to myocardial disease by stimulating MAP kinase signalling in fibroblasts. Nature, 456, 980–984.
Urbich, C., Kuehbacher, A., & Dimmeler, S. (2008). Role of microRNAs in vascular diseases, inflammation, and angiogenesis. Cardiovascular Research, 79, 581–588.
Valastyan, S., Reinhardt, F., Benaich, N., Calogrias, D., Szasz, A. M., Wang, Z. C., et al. (2009). A pleiotropically acting microRNA, miR-31, inhibits breast cancer metastasis. Cell, 137, 1032–1046.
van Rooij, E., Quiat, D., Johnson, B. A., Sutherland, L. B., Qi, X., Richardson, J. A., et al. (2009). A family of microRNAs encoded by myosin genes governs myosin expression and muscle performance. Developments in Cell, 17, 662–673.
van Rooij, E., Sutherland, L. B., Liu, N., Williams, A. H., McAnally, J., Gerard, R. D., et al. (2006). A signature pattern of stress-responsive microRNAs that can evoke cardiac hypertrophy and heart failure. Proceedings of the National Academy of Sciences of the United States of America, 103, 18255–18260.
van Rooij, E., Sutherland, L. B., Qi, X., Richardson, J. A., Hill, J., & Olson, E. N. (2007). Control of stress-dependent cardiac growth and gene expression by a microRNA. Science, 316, 575–579.
van Rooij, E., Sutherland, L. B., Thatcher, J. E., DiMaio, J. M., Naseem, R. H., Marshall, W. S., et al. (2008). Dysregulation of microRNAs after myocardial infarction reveals a role of miR-29 in cardiac fibrosis. Proceedings of the National Academy of Sciences of the United States of America, 105, 13027–13032.
van Solingen, C., Seghers, L., Bijkerk, R., Duijs, J. M., Roeten, M. K., van Oeveren-Rietdijk, A. M., et al. (2009). Antagomir-mediated silencing of endothelial cell specific microRNA-126 impairs ischemia-induced angiogenesis. Journal of Cellular and Molecular Medicine, 13, 1577–1585.
Vasilescu, C., Rossi, S., Shimizu, M., Tudor, S., Veronese, A., Ferracin, M., et al. (2009). MicroRNA fingerprints identify miR-150 as a plasma prognostic marker in patients with sepsis. PLoS ONE, 4, e7405.
Vasudevan, S., Tong, Y., & Steitz, J. A. (2007). Switching from repression to activation: MicroRNAs can up-regulate translation. Science, 318, 1931–1934.
Wang, H., Garzon, R., Sun, H., Ladner, K. J., Singh, R., Dahlman, J., et al. (2008). NF-kappaB-YY1-miR-29 regulatory circuitry in skeletal myogenesis and rhabdomyosarcoma. Cancer Cell, 14, 369–381.
Wang, S., Aurora, A. B., Johnson, B. A., Qi, X., McAnally, J., Hill, J. A., et al. (2008). The endothelial-specific microRNA miR-126 governs vascular integrity and angiogenesis. Developments in Cell, 15, 261–271.
Wienholds, E., Koudijs, M. J., van Eeden, F. J., Cuppen, E., & Plasterk, R. H. (2003). The microRNA-producing enzyme Dicer1 is essential for zebrafish development. Nature Genetics, 35, 217–218.
Wolfrum, C., Shi, S., Jayaprakash, K. N., Jayaraman, M., Wang, G., Pandey, R. K., et al. (2007). Mechanisms and optimization of in vivo delivery of lipophilic siRNAs. Nature Biotechnology, 25, 1149–1157.
Xiao, J., Luo, X., Lin, H., Zhang, Y., Lu, Y., Wang, N., et al. (2007). MicroRNA miR-133 represses HERG K+ channel expression contributing to QT prolongation in diabetic hearts. Journal of Biological Chemistry, 282, 12363–12367.
Xu, C., Lu, Y., Pan, Z., Chu, W., Luo, X., Lin, H., et al. (2007). The muscle-specific microRNAs miR-1 and miR-133 produce opposing effects on apoptosis by targeting HSP60, HSP70 and caspase-9 in cardiomyocytes. Journal of Cell Science, 120, 3045–3052.
Yang, B., Lin, H., Xiao, J., Lu, Y., Luo, X., Li, B., et al. (2007). The muscle-specific microRNA miR-1 regulates cardiac arrhythmogenic potential by targeting GJA1 and KCNJ2. Nature Medicine, 13, 486–491.
Yekta, S., Shih, I. H., & Bartel, D. P. (2004). MicroRNA-directed cleavage of HOXB8 mRNA. Science, 304, 594–596.
Yin, C., Salloum, F. N., & Kukreja, R. C. (2009). A novel role of microRNA in late preconditioning: Upregulation of endothelial nitric oxide synthase and heat shock protein 70. Circulation Research, 104, 572–575.
Yin, C., Wang, X., & Kukreja, R. C. (2008). Endogenous microRNAs induced by heat-shock reduce myocardial infarction following ischemia-reperfusion in mice. FEBS Letters, 582, 4137–4142.
York, M., Scudamore, C., Brady, S., Chen, C., Wilson, S., Curtis, M., et al. (2007). Characterization of troponin responses in isoproterenol-induced cardiac injury in the Hanover Wistar rat. Toxicologic Pathology, 35, 606–617.
Zhao, Y., Ransom, J. F., Li, A., Vedantham, V., von Drehle, M., Muth, A. N., et al. (2007). Dysregulation of cardiogenesis, cardiac conduction, and cell cycle in mice lacking miRNA-1-2. Cell, 129, 303–317.
Zhao, Y., Samal, E., & Srivastava, D. (2005). Serum response factor regulates a muscle-specific microRNA that targets Hand2 during cardiogenesis. Nature, 436, 214–220.
Author information
Authors and Affiliations
Corresponding author
Additional information
Note
Due to the space restrictions we could not cite many other significant contributions made in this rapidly progressing field.
Rights and permissions
About this article
Cite this article
Kartha, R.V., Subramanian, S. MicroRNAs in Cardiovascular Diseases: Biology and Potential Clinical Applications. J. of Cardiovasc. Trans. Res. 3, 256–270 (2010). https://doi.org/10.1007/s12265-010-9172-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12265-010-9172-z