
Abstract
Exosomes, nano-sized extracellular vesicles secreted by most cell types, are found in all kinds of biological fluids and tissues, including the central nervous system (CNS). The proposed functions of these vesicles include roles in cell–cell signaling, removal of cellular debris, and transfer of pathogens between cells. Many studies have revealed that exosomes derived from the CNS occur in the cerebrospinal fluid and peripheral body fluids, and their contents are altered during disease, making them an appealing target for biomarker development in Parkinson’s disease (PD). Exosomes have been shown to spread toxic α-synuclein (αsyn) between cells and induce apoptosis, which suggests a key mechanism underlying the spread of αsyn aggregates in the brain and the acceleration of pathology in PD. However, potential neuroprotective roles of exosomes in PD have also been reported. On the treatment side, as drug delivery vehicles, exosomes have been used to deliver small interfering RNAs and catalase to the brain, and have shown clear therapeutic effects in a mouse model of PD. These features of exosomes in PD make them extremely interesting from the point of view of developing novel diagnostic and therapeutic approaches.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Exosomes are small membranous vesicles that originate from endosomes. They are secreted by virtually all cell types, including neurons, and can be isolated from conditioned cell media or bodily fluids such as cerebrospinal fluid (CSF), plasma, and urine. The exosome contents change during disease, which makes them particularly attractive targets for novel diagnostic approaches, and the important functions of exosomes such as cellular communication allow them to act as notable contributors to both health and disease. In this review, we discuss the biogenesis of exosomes and their roles in the central nervous system (CNS), and reveal their multiple roles in Parkinson’s disease (PD), the second most common neurodegenerative disease after Alzheimer’s disease (AD).
The Biogenesis of Exosomes and Their Roles in the CNS
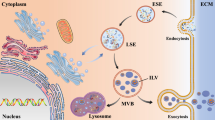
Exosomes are generated within multivesicular bodies (MVBs) that contain intraluminal vesicles in the endosomal system. The endosomal system consists of primary endocytic vesicles, early endosomes (EEs), and MVBs. EEs are located near the cell membrane and act as the first port of call for primary endocytosed vesicles. After leaving the EE, the cargo is recycled to the plasma membrane or targeted to MVBs. MVBs then fuse with plasma and endosomal membranes to release exosomes into the extracellular space [1, 2].
Exosomes typically have a diameter of 40–150 nm. They present a cup-shaped morphology under the transmission electron microscope, but cryo-electron microscopy of unfixed exosomes has revealed a spherical morphology [3]. Exosomes are secreted in vitro and in vivo by various types of cells, including those of the CNS, under physiological and pathological conditions. They have been isolated from blood, CSF, urine, saliva, and milk [4,5,6,7]. Due to their different cellular ancestries, exosomes carry cell-type-specific proteins and lipids. For instance, exosomes derived from oligodendrocytes carry myelin proteins [8]. Several proteins are specifically found on the surface of all exosomes and thereby serve as exosomal markers. They include the multivesicular endosome proteins Alix and tsg101, integrins, the tetraspanins CD63, CD89, CD81, CD9, and CD82, and the heat-shock proteins hsp70 and hsp90, as well as the endosomal and endosome maturation-related proteins flotillin and annexin [9]. On the other hand, proteins such as transferrin receptor and GM130 derived from the nucleus, mitochondria, Golgi apparatus, or endoplasmic reticulum are not detectable in exosomes and thereby serve as negative markers. Apart from various types of proteins, nucleic acid constituents including messenger RNA (mRNA), small non-coding microRNAs (miRNAs), and double-stranded DNA have also been detected in exosomes and may function in cardiovascular protection and repair [10, 11].
Recently, many types of cells in the CNS, including oligodendrocytes [8, 12], neurons [13, 14], and astrocytes [15] have been confirmed to secrete exosomes. It is believed that exosomes as potential carriers can disseminate disease pathology in neurodegenerative disorders. Exosomes secreted from PrP (prion protein)-expressing neurons are reported to contain PrPsc (scrapie isoform) and PrPc (cellular isoform) when parental cells are infected with PrPsc [16, 17]. Moreover, there is evidence that exosomes facilitate the unique transmissible nature of prions. For instance, the exosomes from prion-infected cells can cause prion propagation in uninfected recipient cells and produce prion disease-like clinical features when inoculated into mice [18]. While prion disease has generally been thought to be the only neurodegenerative disease that is transmissible, misfolded forms of proteins involved in other neurodegenerative disorders such as AD, PD, and amyotrophic lateral sclerosis may also spread to different brain regions similar to prions [19, 20]. The exosomes from either neurons or brain tissues contain amyloid-β precursor protein (APP) and APP-processing products, C-terminal fragments and amyloid-β [21, 22]. Furthermore, neuronal exosomes accelerate the propensity of soluble amyloid peptides to form fibrils [23]. Misfolded Cu/Zn superoxide dismutase (SOD1) propagation between cells can be mediated through exosomes. This may help to explain why SOD1 aggregates propagate in a prion-like manner in neurons and how sporadic amyotrophic lateral sclerosis spreads systemically from region to region in a progressive manner [24, 25].
Several properties of exosomes make them excellent candidates for carrying biomarkers. Their cargos of cell-type-specific proteins and nucleic acids are likely to reflect the core of pathogenic intracellular processes. Moreover, exosomes can cross the blood-brain barrier (BBB) so that CNS-derived exosomes can reach the peripheral blood and provide protein biomarkers of CNS disease. For instance, altered levels of autolysosomal proteins such as cathepsin D, lysosome-associated membrane protein 1, and ubiquitinylated proteins, as well as AD-associated proteins including P-S396-tau, P-T181-tau, and Aβ1-42, are found in CNS-derived blood exosomes in patients with AD and appear to reflect the pathology up to 10 years before clinical onset [26, 27].
Exosomes in Parkinson’s Disease
PD, a progressive movement disorder, is the second most common neurodegenerative disease after AD and affects ~2% of the population aged over 65 years [28]. The common clinical manifestations are the characteristic motor symptoms of resting tremor, bradykinesia, muscular rigidity, and postural instability. In PD, degeneration of dopaminergic neurons in the substantia nigra leads to an imbalance of excitatory (acetylcholine) and inhibitory (dopamine, DA) neurotransmitters, which causes the motor symptoms [29].
One of the major pathological characteristics of PD is the presence of Lewy bodies [30]. The misfolding and fibrillar aggregation of α-synuclein (αsyn) constitutes the main component of Lewy bodies and Lewy neurites in both the genetic and sporadic forms of PD [30]. Other mechanisms include reactive oxygen species, neuroinflammation, excitotoxicity, apoptosis, and loss of trophic factors. However, none of these is the only contributor to the generation of PD; rather, molecular pathways act together to induce the degeneration of dopaminergic neurons.
At present, there is no cure for PD, and the goals of treatment are to alleviate the symptoms for the comfort of patients and to minimize the dyskinesia. Levodopa is the primary line of DA replacement therapy, which restores deficits resulting from the loss of dopaminergic neurons. This provides the greatest symptomatic benefit and alleviates the motor symptoms [31]. However, as the effectiveness diminishes with time, clinicians have to delay prescription of levodopa as much as possible. Reports have shown that only ~25% of patients treated with levodopa for 5 years continue to have a good response [32, 33]. In addition, adverse effects of levodopa (dyskinesia, psychosis, hallucinations, and hypotension) have been noted [34,35,36]. Furthermore, although there are other therapeutic options such as Sinemet (carbidopa-levodopa), Rasagiline, antioxidants, and factors that inhibit apoptosis, as well as operative treatments such as deep brain stimulation, their side-effects are also of serious concern [37,38,39,40,41].
There is a presymptomatic phase during PD progression, in which clinical signs do not appear until 70%–80% of the dopaminergic terminals in the striatum and >50% of the dopaminergic neurons in the substantia nigra have been lost [42]. This highlights the importance of early detection and intervention, which may prevent the degeneration of dopaminergic neurons. Moreover, numerous studies have shown that the development of disease-modifying treatments for PD is hampered by the lack of sensitive and specific biomarkers.
Exosomes as Biomarkers in PD
A biomarker has been defined as “a role that is objectively measured and evaluated as an indicator of physiological processes, pathogenic processes or pharmacologic response to a therapeutic intervention” [43]. At present, the diagnosis of PD is typically based on observation of the motor symptoms but the misdiagnosis rate is appreciably high, particularly in the early stages and in the presymptomatic phase [44]. Biomarkers of PD have gained increasing attention, and the discovery of disease-related proteins in exosomes isolated from plasma/CSF samples from PD patients has inspired research into the use of exosomes as biomarkers.
αsyn in the CSF has been consistently reported to be significantly lower in PD patients than in controls [45,46,47]. However, as peripheral cells, especially red blood cells and platelets, can produce abundant αsyn [48, 49], the use of blood αsyn as a biomarker has been found to be readily accessible but inconsistent [50,51,52]. By using the anti-L1CAM (neural cell adhesion molecule L1) antibody which specifically identifies exosomes derived from the CNS, Shi et al. discovered that CNS-derived exosomes can efflux into blood and the level of αsyn from CNS-derived exosomes in plasma is substantially higher in PD patients and is associated with the severity of the disease [53]. Therefore, they concluded that plasma CNS-derived exosomal αsyn can serve as a PD biomarker with high sensitivity and specificity [53]. In CSF samples, Stuendl and co-workers found decreased exosomal αsyn levels in PD patients, consistent with the total αsyn levels in CSF [54]. It is likely that tau is transported from brain to blood by exosomes, and CNS-derived exosomal tau in plasma is significantly higher in PD patients than in controls and is correlated with CSF total tau and phosphorylated tau [55].
Mutations in the leucine-rich repeat kinase 2 (LRRK2) gene cause late-onset PD in both familial and sporadic late-onset populations [56, 57]. In 2013, Fraser et al. reported that LRRK2 is secreted by exosomes from various kinds of cells where LRRK2 is natively expressed, including duct epithelial cells in the kidney, neurons, and macrophages. Thus, LRRK2 can be detected through the purification of exosomes from clinical samples of urine or CSF, which provides a foundation for using exosomal LRRK2 as a biomarker in clinical trials [58], and this has been confirmed by Ho and coworkers in Korean PD patients [59]. In addition, Ho et al. found that the protein levels of LRRK2 and DJ-1 (PD causative gene products) in urinary exosomes show clear gender-dependent differences. In male patients, the DJ-1 levels are significantly higher in PD than in controls and increase in an age-dependent manner in PD [59]. Another research team also reported gender differences of Ser(P)-1292 LRRK2 levels in urinary exosomes [60]. They showed that the exosomal Ser(P)-1292 LRRK2 levels are higher in idiopathic PD, especially in females, than in controls, and are correlated with the severity of cognitive impairment and the loss of activities of daily living, independent of age. Although the accuracy is not high, the Ser(P)-1292 LRRK2 levels in urinary exosomes appear to predict several aspects of PD severity [60]. In the same year, these authors focused on familial PD, and found higher ratios of phosphorylated Ser-1292 LRRK2 to total LRRK2 in urinary exosomes from LRRK2 mutation carriers than in those of non-carriers (with or without PD). Among the carriers, those with PD have a higher ratio of Ser(P)-1292 LRRK2 to total LRRK2 in urinary exosomes than those without PD. Moreover, an elevated ratio predicts the LRRK2 mutation status and PD risk among LRRK2 mutation carriers [61].
In addition to proteins, it has long been known that exosomes contain various RNA species. Gui and co-workers explored the exosomal miRNAs isolated from the CSF in PD and AD patients. They found that 16 exosomal miRNAs are upregulated while 11 miRNAs are down-regulated in PD patients compared with controls. Among these, miR-1 and miR-19b-3p have been validated and are significantly reduced in PD CSF exosomes. On the contrary, miR-153, miR-409-3p, miR-10a-5p, and let-7g-3p are increased. On the other hand, different levels of mRNA transcripts such as APP, αsyn, and long non-coding RNAs (RP11-462G22.1 and PCA3) occur in CSF exosomes from PD compared to AD patients [62]. This evidence shows the potential value of CSF exosomal RNA for the diagnosis and assessment of PD.
The above studies support the idea that exosomes have potential value in PD diagnosis (summarized in Table 1). However, substantiation in large population studies is still needed, and the sensitivity and specificity of these biomarkers need to be further confirmed by clinical studies. At the same time, there is a pressing need to develop exosome isolation methods that exclude lipoprotein aggregates and contaminating proteins. They are of particular importance for improving the diagnostic value of exosomes.
Exosomes as Both a Curse and a Blessing in the Progress of PD
Lewy bodies often initially occur in the periphery, gradually affect the brain stem, and eventually appear in the cortex [63]. In addition, Lewy bodies have been observed in mesencephalic stem-cell transplants in recipients with PD, suggesting host-to-graft transfer of αsyn pathology [64], and seeded propagation of αsyn is a leading hypothesis to explain the interneuronal transmission of pathology [65,66,67]. Exosomes are biologically active vesicles that participate in intercellular communication [68, 69]. They have the exceptional property of interacting with target cell membranes and thereby act as nanocarriers of biological information. Ghidoni et al. have proposed the “Trojan horse” hypothesis of exosomes in neurodegeneration; a mechanism leading to the death of cells by shipping toxic agents in exosomes from cell to cell [70]. In fact, many researchers believe that exosomes act as potential intercellular carriers of pathogenic proteins and cause impaired neuronal function [9, 19, 20, 71].
It is widely accepted that αsyn oligomers cause neuronal death [72]. αsyn in exosomes may not only serve as a biomarker for PD, but also as a potentially pivotal player in the propagation of the neurotoxic form of αsyn and its spread in the brain. αsyn has been detected in exosomes separated from the conditioned media of SH-SY5Y cells with wild-type αsyn and inducible β-galactosidase, and the exosome-associated αsyn impacts the viability of neighboring neurons [73]. In 2011, Alvarez et al. published a study in which they demonstrated that exosomes from SH-SY5Y cells overexpressing αsyn efficiently transfer αsyn to normal SH-SY5Y cells. Moreover, when the release of αsyn from exosomes increases, its transmission to recipient cells increases correspondingly [74]. Soon after that, Danzer et al. performed a similar study and found that exosome-associated αsyn oligomers in vitro are preferentially internalized by recipient cells and are more toxic than free αsyn oligomers [71]. Moreover, the aggregation of exogenous αsyn can be accelerated by exosomes, caused by the exosome lipid [75]. αsyn can induce an increase of exosomal secretion by microglial BV-2 cells, and these activated exosomes cause increased apoptosis of cortical neurons. The authors concluded that exosomes from activated microglia may be important mediators of αsyn-induced neurodegeneration in PD [76]. Some years later, Kunadt et al. provided evidence for exosomal αsyn in the CNS in vivo [77]. The next year, these authors published a study showing that CSF exosomes derived from PD patients contain a pathogenic species of αsyn, and these exosomes can initiate the oligomerization of soluble α-synuclein in target cells in a dose-dependent manner and confer disease pathology [54]. These studies support the notion that exosomes contain αsyn “seeds” or “strains” that spread αsyn aggregates in the brain and aid pathology.
Though the intercellular transfer of αsyn by exosomes may cause the spread of pathology, neurons can benefit from the reduction of its intracellular level by αsyn externalization via exosomes. The surviving DA neurons in the substantia nigra of PD patients express higher levels of ATP13A2 mRNA and protein than controls [78, 79]. Gitler et al. further showed inhibitory effects of ATP13A2 on the αsyn toxicity in primary DA neurons [80]. Then, Kong and co-workers reported that elevated ATP13A2 expression increases the externalization of exosome-associated αsyn and thereby reduces the intracellular αsyn levels, while reduced ATP13A2 expression correspondingly leads to decreased levels of exosomal αsyn [81]. Further functional evidence for the neuroprotective efficacy of exosomes came from a study by Tomlinson et al. showing that PD-derived microvesicles are protective in models of neuronal stress [82]. These results indicate a potential neuroprotective role of exosomes in PD.
The evidence shows contradictory roles of exosomes in PD, which may be due to the properties of exosome complexes that are not completely clear. But different exosomes from different cellular ancestries in the brain have unique characteristics, so their roles in PD pathology may be distinctly different. In future, studies of exosomes may need to take into consideration their sources.
Exosomes as Drug-Delivery Vehicles for PD Therapy
It has been reported that ~98% of all potent drugs that may be therapeutic for many diseases in the CNS failed in clinic trials because of their inability to cross the BBB [83]. As naturally-occurring nano-sized vesicles, exosomes can cross the BBB and have attracted considerable attention as drug-delivery vehicles [84, 85]. The incorporation of therapeutic agents into exosomes preserves the therapeutic activity, increases the circulation time, and improves delivery to the brain. For instance, exosomes have been harnessed for the systemic delivery of therapeutic agents such as curcumin and exogenous siRNA across the BBB [84, 86, 87]. In PD, when macrophages are transfected with plasmid DNA encoding catalase, the exosomes secreted from these macrophages are packed with catalase genetic material including plasmid DNA and mRNA, active catalase, and nuclear factor κ-B, a transcription factor involved in encoded gene expression. What is more, these contents can be efficiently transferred by exosomes to neurons and thereby result in de novo protein synthesis in target cells. Ultimately, this approach results in reduced inflammation and neuroprotection in PD mice [88]. Peripheral injection of modified exosomes expressing rabies virus glycoprotein loaded with siRNA to αsyn leads to decreased αsyn mRNA and protein levels in S129D αsyn transgenic mice [89]. Exosomes secreted by monocytes and macrophages loaded with catalase efficiently accumulate in neurons and microglial cells in the brain, decrease brain inflammation, and significantly increase neuronal survival in PD mice [90].
Accumulating evidence suggests that exosomes can be engineered to target neurons or even specific neuronal populations and be effectively used for the treatment of various neurodegenerative disorders. However, several problems must be solved before they are used in the clinical setting. First, exosomes are complex systems containing various molecular constituents that raise multiple safety issues. Thus, future studies will focus on the engineering of exosome-mimetic delivery systems containing only the desired therapeutic molecules. On the other hand, more experiments are needed to evaluate possible adverse effects related to the therapeutic administration of exosomes. It is known that exosomes vary considerably among different cellular sources, so it is important to identify the best sources. The role of exosomes as potential novel therapeutic tools against neurodegenerative diseases has recently (2016) been discussed in detail in a review article by Jarmalaviciute and Pivoriunas [91].
Conclusions
The field of exosome studies has recently been attracting increasing interest. In recent years, exosomes have risen from being considered mere cellular “trash-cans” to possible biomarkers and multipotent therapeutic targets of various diseases, among which PD is one of the most important. Though exosomes have been shown to possess the ability to propagate PD pathology as well as to hinder it, we believe that eventually exosomes can be harnessed for the benefit of PD patients.
References
Heijnen HF, Schiel AE, Fijnheer R, Geuze HJ, Sixma JJ. Activated platelets release two types of membrane vesicles: microvesicles by surface shedding and exosomes derived from exocytosis of multivesicular bodies and alpha-granules. Blood 1999, 94: 3791–3799.
Le Blanc I, Luyet PP, Pons V, Ferguson C, Emans N, Petiot A, et al. Endosome-to-cytosol transport of viral nucleocapsids. Nat Cell Biol 2005, 7: 653–664.
Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol 2013, 200: 373–383.
Caby MP, Lankar D, Vincendeau-Scherrer C, Raposo G, Bonnerot C. Exosomal-like vesicles are present in human blood plasma. Int Immunol 2005, 17: 879–887.
Pisitkun T, Shen RF, Knepper MA. Identification and proteomic profiling of exosomes in human urine. Proc Natl Acad Sci U S A 2004, 101: 13368–13373.
Ogawa Y, Kanai-Azuma M, Akimoto Y, Kawakami H, Yanoshita R. Exosome-like vesicles with dipeptidyl peptidase IV in human saliva. Biol Pharm Bull 2008, 31: 1059–1062.
Vella LJ, Greenwood DL, Cappai R, Scheerlinck JP, Hill AF. Enrichment of prion protein in exosomes derived from ovine cerebral spinal fluid. Vet Immunol Immunopathol 2008, 124: 385–393.
Kramer-Albers EM, Bretz N, Tenzer S, Winterstein C, Mobius W, Berger H, et al. Oligodendrocytes secrete exosomes containing major myelin and stress-protective proteins: Trophic support for axons? Proteomics Clin Appl 2007, 1: 1446–1461.
Simons M, Raposo G. Exosomes–vesicular carriers for intercellular communication. Curr Opin Cell Biol 2009, 21: 575–581.
Emanueli C, Shearn AI, Angelini GD, Sahoo S. Exosomes and exosomal miRNAs in cardiovascular protection and repair. Vascul Pharmacol 2015, 71: 24–30.
Thakur BK, Zhang H, Becker A, Matei I, Huang Y, Costa-Silva B, et al. Double-stranded DNA in exosomes: a novel biomarker in cancer detection. Cell Res 2014, 24: 766–769.
Bakhti M, Winter C, Simons M. Inhibition of myelin membrane sheath formation by oligodendrocyte-derived exosome-like vesicles. J Biol Chem 2011, 286: 787–796.
Faure J, Lachenal G, Court M, Hirrlinger J, Chatellard-Causse C, Blot B, et al. Exosomes are released by cultured cortical neurones. Mol Cell Neurosci 2006, 31: 642–648.
Lachenal G, Pernet-Gallay K, Chivet M, Hemming FJ, Belly A, Bodon G, et al. Release of exosomes from differentiated neurons and its regulation by synaptic glutamatergic activity. Mol Cell Neurosci 2011, 46: 409–418.
Taylor AR, Robinson MB, Gifondorwa DJ, Tytell M, Milligan CE. Regulation of heat shock protein 70 release in astrocytes: role of signaling kinases. Dev Neurobiol 2007, 67: 1815–1829.
Veith NM, Plattner H, Stuermer CA, Schulz-Schaeffer WJ, Burkle A. Immunolocalisation of PrPSc in scrapie-infected N2a mouse neuroblastoma cells by light and electron microscopy. Eur J Cell Biol 2009, 88: 45–63.
Vella LJ, Sharples RA, Lawson VA, Masters CL, Cappai R, Hill AF. Packaging of prions into exosomes is associated with a novel pathway of PrP processing. J Pathol 2007, 211: 582–590.
Guo BB, Bellingham SA, Hill AF. Stimulating the release of exosomes increases the intercellular transfer of prions. J Biol Chem 2016, 291: 5128–5137.
Bellingham SA, Guo BB, Coleman BM, Hill AF. Exosomes: vehicles for the transfer of toxic proteins associated with neurodegenerative diseases? Front Physiol 2012, 3: 124.
Coleman BM, Hill AF. Extracellular vesicles–Their role in the packaging and spread of misfolded proteins associated with neurodegenerative diseases. Semin Cell Dev Biol 2015, 40: 89–96.
Vingtdeux V, Hamdane M, Loyens A, Gele P, Drobeck H, Begard S, et al. Alkalizing drugs induce accumulation of amyloid precursor protein by-products in luminal vesicles of multivesicular bodies. J Biol Chem 2007, 282: 18197–18205.
Perez-Gonzalez R, Gauthier SA, Kumar A, Levy E. The exosome secretory pathway transports amyloid precursor protein carboxyl-terminal fragments from the cell into the brain extracellular space. J Biol Chem 2012, 287: 43108–43115.
Yuyama K, Sun H, Mitsutake S, Igarashi Y. Sphingolipid-modulated exosome secretion promotes clearance of amyloid-beta by microglia. J Biol Chem 2012, 287: 10977–10989.
Grad LI, Yerbury JJ, Turner BJ, Guest WC, Pokrishevsky E, O’Neill MA, et al. Intercellular propagated misfolding of wild-type Cu/Zn superoxide dismutase occurs via exosome-dependent and -independent mechanisms. Proc Natl Acad Sci U S A 2014, 111: 3620–3625.
Munch C, O’Brien J, Bertolotti A. Prion-like propagation of mutant superoxide dismutase-1 misfolding in neuronal cells. Proc Natl Acad Sci U S A 2011, 108: 3548–3553.
Goetzl EJ, Boxer A, Schwartz JB, Abner EL, Petersen RC, Miller BL, et al. Altered lysosomal proteins in neural-derived plasma exosomes in preclinical Alzheimer disease. Neurology 2015, 85: 40–47.
Fiandaca MS, Kapogiannis D, Mapstone M, Boxer A, Eitan E, Schwartz JB, et al. Identification of preclinical Alzheimer’s disease by a profile of pathogenic proteins in neurally derived blood exosomes: A case-control study. Alzheimers Dement 2015, 11: 600–607 e601.
Lang AE, Lozano AM. Parkinson’s disease. Second of two parts. N Engl J Med 1998, 339: 1130–1143.
Olanow CW, Stern MB, Sethi K. The scientific and clinical basis for the treatment of Parkinson disease. Neurology 2009, 72: S1–136.
Goedert M. Alpha-synuclein and neurodegenerative diseases. Nat Rev Neurosci 2001, 2: 492–501.
Olanow CW, Tatton WG. Etiology and pathogenesis of Parkinson’s disease. Annu Rev Neurosci 1999, 22: 123–144.
Agid Y, Olanow CW, Mizuno Y. Levodopa: why the controversy? Lancet 2002, 360: 575.
Fahn S, Oakes D, Shoulson I, Kieburtz K, Rudolph A, Lang A, et al. Levodopa and the progression of Parkinson’s disease. N Engl J Med 2004, 351: 2498–2508.
Lieberman AN, Pedersen B. Levodopa and adventitious movements. Lancet 1970, 2: 985.
Celesia GG, Barr AN. Psychosis and other psychiatric manifestations of levodopa therapy. Arch Neurol 1970, 23: 193–200.
McDowell FH, Lee JE. Levodopa, Parkinson’s disease, and hypotension. Ann Intern Med 1970, 72: 751–752.
Guay DR. Rasagiline (TVP-1012): a new selective monoamine oxidase inhibitor for Parkinson’s disease. Am J Geriatr Pharmacother 2006, 4: 330–346.
Olanow CW, Schapira AH, LeWitt PA, Kieburtz K, Sauer D, Olivieri G, et al. TCH346 as a neuroprotective drug in Parkinson’s disease: a double-blind, randomised, controlled trial. Lancet Neurol 2006, 5: 1013–1020.
Kordower JH, Emborg ME, Bloch J, Ma SY, Chu Y, Leventhal L, et al. Neurodegeneration prevented by lentiviral vector delivery of GDNF in primate models of Parkinson’s disease. Science 2000, 290: 767–773.
Lang AE, Gill S, Patel NK, Lozano A, Nutt JG, Penn R, et al. Randomized controlled trial of intraputamenal glial cell line-derived neurotrophic factor infusion in Parkinson disease. Ann Neurol 2006, 59: 459–466.
Smeding HM, Speelman JD, Huizenga HM, Schuurman PR, Schmand B. Predictors of cognitive and psychosocial outcome after STN DBS in Parkinson’s Disease. J Neurol Neurosurg Psychiatry 2011, 82: 754–760.
Fearnley JM, Lees AJ. Ageing and Parkinson’s disease: substantia nigra regional selectivity. Brain 1991, 114 (Pt 5): 2283–2301.
Biomarkers Definitions Working G. Biomarkers and surrogate endpoints: preferred definitions and conceptual framework. Clin Pharmacol Ther 2001, 69: 89–95.
Tolosa E, Wenning G, Poewe W. The diagnosis of Parkinson’s disease. Lancet Neurol 2006, 5: 75–86.
Hong Z, Shi M, Chung KA, Quinn JF, Peskind ER, Galasko D, et al. DJ-1 and alpha-synuclein in human cerebrospinal fluid as biomarkers of Parkinson’s disease. Brain 2010, 133: 713–726.
Mollenhauer B, Locascio JJ, Schulz-Schaeffer W, Sixel-Doring F, Trenkwalder C, Schlossmacher MG. alpha-Synuclein and tau concentrations in cerebrospinal fluid of patients presenting with parkinsonism: a cohort study. Lancet Neurol 2011, 10: 230–240.
Shi M, Bradner J, Hancock AM, Chung KA, Quinn JF, Peskind ER, et al. Cerebrospinal fluid biomarkers for Parkinson disease diagnosis and progression. Ann Neurol 2011, 69: 570–580.
Barbour R, Kling K, Anderson JP, Banducci K, Cole T, Diep L, et al. Red blood cells are the major source of alpha-synuclein in blood. Neurodegener Dis 2008, 5: 55–59.
Marques O, Outeiro TF. Alpha-synuclein: from secretion to dysfunction and death. Cell Death Dis 2012, 3: e350.
Lee PH, Lee G, Park HJ, Bang OY, Joo IS, Huh K. The plasma alpha-synuclein levels in patients with Parkinson’s disease and multiple system atrophy. J Neural Transm (Vienna) 2006, 113: 1435–1439.
Shi M, Zabetian CP, Hancock AM, Ginghina C, Hong Z, Yearout D, et al. Significance and confounders of peripheral DJ-1 and alpha-synuclein in Parkinson’s disease. Neurosci Lett 2010, 480: 78–82.
Li QX, Mok SS, Laughton KM, McLean CA, Cappai R, Masters CL, et al. Plasma alpha-synuclein is decreased in subjects with Parkinson’s disease. Exp Neurol 2007, 204: 583–588.
Shi M, Liu C, Cook TJ, Bullock KM, Zhao Y, Ginghina C, et al. Plasma exosomal alpha-synuclein is likely CNS-derived and increased in Parkinson’s disease. Acta Neuropathol 2014, 128: 639–650.
Stuendl A, Kunadt M, Kruse N, Bartels C, Moebius W, Danzer KM, et al. Induction of alpha-synuclein aggregate formation by CSF exosomes from patients with Parkinson’s disease and dementia with Lewy bodies. Brain 2016, 139: 481–494.
Shi M, Kovac A, Korff A, Cook TJ, Ginghina C, Bullock KM, et al. CNS tau efflux via exosomes is likely increased in Parkinson’s disease but not in Alzheimer’s disease. Alzheimers Dement 2016, 12: 1125–1131.
Zimprich A, Biskup S, Leitner P, Lichtner P, Farrer M, Lincoln S, et al. Mutations in LRRK2 cause autosomal-dominant parkinsonism with pleomorphic pathology. Neuron 2004, 44: 601–607.
Gilks WP, Abou-Sleiman PM, Gandhi S, Jain S, Singleton A, Lees AJ, et al. A common LRRK2 mutation in idiopathic Parkinson’s disease. Lancet 2005, 365: 415–416.
Fraser KB, Moehle MS, Daher JP, Webber PJ, Williams JY, Stewart CA, et al. LRRK2 secretion in exosomes is regulated by 14-3-3. Hum Mol Genet 2013, 22: 4988–5000.
Ho DH, Yi S, Seo H, Son I, Seol W. Increased DJ-1 in urine exosome of Korean males with Parkinson’s disease. Biomed Res Int 2014, 2014: 704678.
Fraser KB, Rawlins AB, Clark RG, Alcalay RN, Standaert DG, Liu N, et al. Ser(P)-1292 LRRK2 in urinary exosomes is elevated in idiopathic Parkinson’s disease. Mov Disord 2016, 31: 1543–1550
Fraser KB, Moehle MS, Alcalay RN, West AB, Consortium LC. Urinary LRRK2 phosphorylation predicts parkinsonian phenotypes in G2019S LRRK2 carriers. Neurology 2016, 86: 994–999.
Gui Y, Liu H, Zhang L, Lv W, Hu X. Altered microRNA profiles in cerebrospinal fluid exosome in Parkinson disease and Alzheimer disease. Oncotarget 2015, 6: 37043–37053.
Braak H, Del Tredici K, Rub U, de Vos RA, Jansen Steur EN, Braak E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol Aging 2003, 24: 197–211.
Kordower JH, Chu Y, Hauser RA, Freeman TB, Olanow CW. Lewy body-like pathology in long-term embryonic nigral transplants in Parkinson’s disease. Nat Med 2008, 14: 504–506.
Desplats P, Lee HJ, Bae EJ, Patrick C, Rockenstein E, Crews L, et al. Inclusion formation and neuronal cell death through neuron-to-neuron transmission of alpha-synuclein. Proc Natl Acad Sci U S A 2009, 106: 13010–13015.
Lee HJ, Patel S, Lee SJ. Intravesicular localization and exocytosis of alpha-synuclein and its aggregates. J Neurosci 2005, 25: 6016–6024.
Lee HJ, Suk JE, Bae EJ, Lee SJ. Clearance and deposition of extracellular alpha-synuclein aggregates in microglia. Biochem Biophys Res Commun 2008, 372: 423–428.
Fevrier B, Raposo G. Exosomes: endosomal-derived vesicles shipping extracellular messages. Curr Opin Cell Biol 2004, 16: 415–421.
Ratajczak J, Wysoczynski M, Hayek F, Janowska-Wieczorek A, Ratajczak MZ. Membrane-derived microvesicles: important and underappreciated mediators of cell-to-cell communication. Leukemia 2006, 20: 1487–1495.
Ghidoni R, Benussi L, Binetti G. Exosomes: the Trojan horses of neurodegeneration. Med Hypotheses 2008, 70: 1226–1227.
Danzer KM, Kranich LR, Ruf WP, Cagsal-Getkin O, Winslow AR, Zhu L, et al. Exosomal cell-to-cell transmission of alpha synuclein oligomers. Mol Neurodegener 2012, 7: 42.
Sung JY, Kim J, Paik SR, Park JH, Ahn YS, Chung KC. Induction of neuronal cell death by Rab5A-dependent endocytosis of alpha-synuclein. J Biol Chem 2001, 276: 27441–27448.
Emmanouilidou E, Melachroinou K, Roumeliotis T, Garbis SD, Ntzouni M, Margaritis LH, et al. Cell-produced alpha-synuclein is secreted in a calcium-dependent manner by exosomes and impacts neuronal survival. J Neurosci 2010, 30: 6838–6851.
Alvarez-Erviti L, Seow Y, Schapira AH, Gardiner C, Sargent IL, Wood MJ, et al. Lysosomal dysfunction increases exosome-mediated alpha-synuclein release and transmission. Neurobiol Dis 2011, 42: 360–367.
Grey M, Dunning CJ, Gaspar R, Grey C, Brundin P, Sparr E, et al. Acceleration of alpha-synuclein aggregation by exosomes. J Biol Chem 2015, 290: 2969–2982.
Chang C, Lang H, Geng N, Wang J, Li N, Wang X. Exosomes of BV-2 cells induced by alpha-synuclein: important mediator of neurodegeneration in PD. Neurosci Lett 2013, 548: 190–195.
Kunadt M, Eckermann K, Stuendl A, Gong J, Russo B, Strauss K, et al. Extracellular vesicle sorting of alpha-Synuclein is regulated by sumoylation. Acta Neuropathol 2015, 129: 695–713.
Ramirez A, Heimbach A, Grundemann J, Stiller B, Hampshire D, Cid LP, et al. Hereditary parkinsonism with dementia is caused by mutations in ATP13A2, encoding a lysosomal type 5 P-type ATPase. Nat Genet 2006, 38: 1184–1191.
Ramonet D, Podhajska A, Stafa K, Sonnay S, Trancikova A, Tsika E, et al. PARK9-associated ATP13A2 localizes to intracellular acidic vesicles and regulates cation homeostasis and neuronal integrity. Hum Mol Genet 2012, 21: 1725–1743.
Gitler AD, Chesi A, Geddie ML, Strathearn KE, Hamamichi S, Hill KJ, et al. Alpha-synuclein is part of a diverse and highly conserved interaction network that includes PARK9 and manganese toxicity. Nat Genet 2009, 41: 308–315.
Kong SM, Chan BK, Park JS, Hill KJ, Aitken JB, Cottle L, et al. Parkinson’s disease-linked human PARK9/ATP13A2 maintains zinc homeostasis and promotes alpha-Synuclein externalization via exosomes. Hum Mol Genet 2014, 23: 2816–2833.
Tomlinson PR, Zheng Y, Fischer R, Heidasch R, Gardiner C, Evetts S, et al. Identification of distinct circulating exosomes in Parkinson’s disease. Ann Clin Transl Neurol 2015, 2: 353–361.
Pardridge WM. Drug transport across the blood-brain barrier. J Cereb Blood Flow Metab 2012, 32: 1959–1972.
Zhuang X, Xiang X, Grizzle W, Sun D, Zhang S, Axtell RC, et al. Treatment of brain inflammatory diseases by delivering exosome encapsulated anti-inflammatory drugs from the nasal region to the brain. Mol Ther 2011, 19: 1769–1779.
Alvarez-Erviti L, Seow Y, Yin H, Betts C, Lakhal S, Wood MJ. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol 2011, 29: 341–345.
Lai CP, Breakefield XO. Role of exosomes/microvesicles in the nervous system and use in emerging therapies. Front Physiol 2012, 3: 228.
El Andaloussi S, Lakhal S, Mager I, Wood MJ. Exosomes for targeted siRNA delivery across biological barriers. Adv Drug Deliv Rev 2013, 65: 391–397.
Haney MJ, Zhao Y, Harrison EB, Mahajan V, Ahmed S, He Z, et al. Specific transfection of inflamed brain by macrophages: a new therapeutic strategy for neurodegenerative diseases. PLoS One 2013, 8: e61852.
Cooper JM, Wiklander PB, Nordin JZ, Al-Shawi R, Wood MJ, Vithlani M, et al. Systemic exosomal siRNA delivery reduced alpha-synuclein aggregates in brains of transgenic mice. Mov Disord 2014, 29: 1476–1485.
Haney MJ, Klyachko NL, Zhao Y, Gupta R, Plotnikova EG, He Z, et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J Control Release 2015, 207: 18–30.
Jarmalaviciute A, Pivoriunas A. Exosomes as a potential novel therapeutic tools against neurodegenerative diseases. Pharmacol Res 2016, 113: 816–822.
Acknowledgements
This review was supported by the grants of International Cooperative Key Project of National Natural Science Foundation of China (81520108010) and the Natural Science Foundation of Shaoxing Municipality, Zhejiang Province, China (2016QN020).
Author information
Authors and Affiliations
Corresponding author
Additional information
Xiaoqing Wu and Tingting Zheng have contributed equally to this review.
Rights and permissions
About this article

Cite this article
Wu, X., Zheng, T. & Zhang, B. Exosomes in Parkinson’s Disease. Neurosci. Bull. 33, 331–338 (2017). https://doi.org/10.1007/s12264-016-0092-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12264-016-0092-z