
Abstract
The present study examines the temporal variability of air–water CO2 fluxes (FCO2) and seawater carbonate chemistry in a Baja California coastal lagoon during an exceptionally warm anomaly that was developed in Northeast Pacific coasts during 2014. This oceanographic condition led to a summer-like season (weak upwelling condition) during the study period, which reached a maximum surface temperature anomaly of 2 °C in September 2014. San Quintín Bay acts as a source of CO2 to the atmosphere in 2014 (3.3 ± 4.8 mmol C m−2 day−1) with the higher positive fluxes mainly observed in summer months (9.0 ± 5.3 mmol C m−2 day−1). Net ecosystem production (NEP) switched seasonally between net heterotrophy and net autotrophy during the study period, with an annual average of 2.2 ± 7.1 mmol C m−2 day−1, which indicates that San Quintín Bay was a net autotrophic system during the atypical warm oceanographic condition in 2014. This pattern of seasonal variations in the carbon balance at San Quintín Bay appears to be linked to the life cycle of benthic communities, which play an important role in the whole-ecosystem metabolism. Under the limited input from external sources coupled with an increase in seawater temperatures, the recycled benthic carbon and nutrient fluxes play a major role to sustain water-column processes within the bay. Since the upwelling condition may influence the magnitude of the air–water CO2 fluxes, our results clearly indicated that San Quintín Bay is a net source of carbon to the atmosphere regardless of the adjacent oceanic conditions. Our study sheds light on the carbon dynamics and its metabolic implications in a shallow coastal ecosystem under a regional warm anomaly and contributes potentially relevant information in view of the likely future scenario of global climate change.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The global ocean is a key component of the Earth’s carbon reservoir, absorbing ∼1.8 Pg C year−1 of the CO2 emitted into the atmosphere (Doney et al. 2009). Generally, global climatology analyses of carbon fluxes do not consider the coastal ocean (∼7 % of total oceanic surface area) (e.g., Sabine et al. 2004; Takahashi et al. 2009). However, coastal waters are important biogeochemically active areas for the global carbon cycle and carbon sink, accounting for approximately 20 % of total ocean CO2 fluxes (Gypens et al. 2011; Bauer et al. 2013). Although coastal systems play a significant role in biogeochemical dynamics, large uncertainties remain on how these systems affect the ocean and global carbon budgets. Recent summaries of global air–water CO2 fluxes (FCO2) for continental shelves and estuaries estimated that the former act as a net global carbon sink (∼0.40 Pg C year−1), while inner estuaries, mangrove forests, and salt marshes are a carbon source (∼0.10 Pg C year−1) to the atmosphere (Borges 2005; Chen and Borges 2009; Chen et al. 2013). In many cases, FCO2 estimates are poorly represented due to the strong spatial and temporal heterogeneity of coastal ecosystems. The inorganic carbon exchange is influenced by a range of processes that include, for example, biological productivity, upwelling, calcium carbonate precipitation and dissolution, organic matter respiration, organic and inorganic carbon inputs from rivers, etc.; hence, the quantification of lateral and vertical CO2 fluxes is key for determining how these affect the oceanic carbon cycle (Bauer et al. 2013). Specific studies and a better understanding of carbon dynamics are needed in coastal marine systems worldwide, given the current lack of information on FCO2 for these systems. The Pacific coast of the Baja California peninsula, Mexico, is one of these areas of interest where the contribution of coastal carbon fluxes to the global synthesis of FCO2 is little known. Currently, reports related to the coastal carbonate system (dissolved inorganic carbon (DIC), total alkalinity (TA), pH, pCO2) and FCO2 are scarce (e.g. Hernández-Ayón et al. 2007a, b; Feely et al. 2008; Ribas-Ribas et al. 2011; Reimer et al. 2013; Muñoz-Anderson et al. 2015).
The western coastal ecosystems of the Baja California region are located at the southern limit of the California Current System (CCS), influenced by northerly wind-driven coastal upwelling events most of the year (Macías et al. 2012). Coastal upwelling carries dense, cold, CO2-, and nutrient-rich waters to the sea surface near the shore, supporting the high biological activity in this area (Lachkar and Gruber 2013). The peak of the upwelling season occurs during spring and summer, where subarctic waters and subtropical waters in the CCS off Baja California can variably contribute along the year (Durazo 2009). The seasonality of upwelling conditions and its anomalies in the Northwest Pacific coast are influenced primarily by coastal winds, large-scale winds, and offshore and remote ocean conditions (García-Reyes and Largier 2012). Moreover, variations in upwelling conditions at the seasonal-to-interannual timescale in the CCS are linked to the variability in biogeochemical dynamics and biological components in coastal ocean ecosystems (Checkley and Barth 2009).
This study was conducted during an anomalous warm oceanographic condition called “The Blob” that occurred since late 2013 until at least late 2015 across the North Pacific basin (Hartmann 2014; Bond et al. 2015). Areas of the North Pacific have been as much as 5 °C warmer than average, affecting regional weather and climate patterns (http://sccoos.org/projects/anomalies_workshop/). The impact of these regional climate variations and changing environmental conditions in terms of trophic status and metabolic consequences in coastal ecosystems is poorly understood (Testa et al. 2013). In order to better understand the effects of the 2014 oceanographic condition on the coastal carbon dynamics and ecosystem metabolism, we focused the present work on two main goals: first, to examine the spatial and temporal variability of seawater carbonate chemistry in San Quintín Bay, a Baja California Mediterranean-climate coastal lagoon and, second, to estimate, for the first time for this site, the seasonal changes of FCO2 and net ecosystem production (NEP), identifying the key underlying factors and processes. We discuss the role of primary producers and environmental conditions on the system’s trophic status and summarize the key implications of this anomalous condition on carbon dynamics and ecosystem metabolism in San Quintín Bay.
Material and Methods
Study Area
San Quintín Bay (30° 30′ N, 116° W) is a hypersaline coastal lagoon linked to the CCS, subject to frequent upwelling events, mainly in spring and summer, that supply the coastal lagoon with nutrients, phytoplankton, and DIC via tidal exchange (Ribas-Ribas et al. 2011). The lagoon is a well-mixed y-shaped system with only one connection to the ocean, covering an area of approximately 42 km2 with an average depth of 2 m. The seagrass Zostera marina is the main primary producer in San Quintín Bay, forming large meadows that cover approximately 40 % of the bay surface (Ward et al. 2004). Nevertheless, significant seasonal macroalgal blooms occurring within the bay can also be observed (Zertuche-González et al. 2009), as well as large phytoplankton biomass during the upwelling season. The latter constitutes the most important external source of organic matter into the system (Camacho-Ibar et al. 2003).
Field Sampling and Analytical Methods
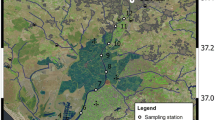
The role of San Quintín Bay as a carbon source or sink in 2014 was investigated by setting a network of ten fixed stations across the bay’s three biogeochemically distinct sections (Camacho-Ibar et al. 2003): the mouth (Mouth) (stations 1–3); the western arm, known as Bahía Falsa (BFa) (stations 4–7); and the eastern arm, known as Bahía San Quintín (bSQ) (stations 8–10) (Fig. 1). At each station, discrete water samples were collected monthly from November 2013 to November 2014 for carbonate system analyses. Seawater samples were stored in 500-mL borosilicate bottles and preserved with 100 μL saturated mercuric chloride (HgCl2). Bottles were sealed with Apiezon grease to prevent evaporation and contact with the atmosphere. DIC was measured following the coulometric method described by Johnson et al. (1987). Prior to the analysis, accuracy and precision were assessed using Certified Reference Material (CRM) for DIC provided by Dr. Andrew Dickson from the Scripps Institution of Oceanography (Dickson et al. 2003). The reference material gave an average difference of 4 μmol kg−1 relative to the certified maximum value, with a measurement error of 0.2 %. TA was measured with the potentiometric method described by Hernández-Ayón et al. (1999). Reference standard materials were used for the measurements, and the differences relative to the certified value were lower than 5 μmol kg−1, with a measurement error of 0.2 %.

San Quintín Bay with its different areas: mouth of the bay (Mouth), Bahía Falsa (BFa) and Bahía San Quintín (bSQ). Numbers indicate sampling stations in 2014, and the black star indicates the location of the oceanographic buoy
Given that San Quintín Bay is a hypersaline coastal lagoon where evaporation processes in the innermost parts of the system are often very important, DIC and TA values were normalized for salinity (normalized dissolved inorganic carbon (NDIC) and normalized total alkalinity (NTA), respectively) using the corresponding salinity for each sampling month from station 1 (oceanic signal). The average salinity during the study period at the outer part of the mouth was 33.64 ± 0.30. Monthly sampling was conducted only during high tide in order to eliminate variations due to tidal condition.
In situ pH (seawater scale) and pCO2 were calculated with the CO2Sys.xls program developed by Lewis and Wallace (1998), using the pair DIC-TA and the dissociation constants proposed by Mehrbach et al. (1973) as refitted by Dickson and Millero (1987). The precision of pH and pCO2 was ±0.01 pH units and ±20 μatm (±SE, n = 100), respectively. Surface seawater temperature (SST) and salinity were determined using a handheld multiparameter probe (YSI Pro Plus, USA) at the time of seawater sampling. The probe was calibrated prior to each field trip following the manufacturer’s recommendations.
Additionally, data from a time series were obtained from a buoy equipped with a multiparameter sonde (YSI 6920V2–2) anchored at the mouth of the bay (asterisk in Fig. 1). Observations were recorded at 15-min intervals for SST, pH (National Bureau of Standards (NBS) scale), and dissolved oxygen saturation (%DO) from February to December 2014 during the sampling period. These data were used to determine the following: (1) high-frequency physicochemical characteristics of ocean–bay seawater exchange and 2) the potential intrusion into the bay of water originated from coastal upwelling. The sonde was calibrated every 2 months during the sampling period following the manufacturer’s recommendations. The optical dissolved oxygen probe was calibrated using the “one-point saturated air” method with a measured precision of ±1 %, and the pH probe was calibrated against an NBS buffer with an error of ±0.01 pH units. The water residence time (τ) was calculated with a one-box model, as described by Camacho-Ibar et al. (2003) for San Quintín Bay.
In order to observe the local effects of upwelling conditions on eelgrass primary production, aboveground biomass of Z. marina (g dry weight (DW) m−2) was estimated each month throughout the sampling period in different eelgrass meadows corresponding to four study sites (stations 2, 4, 7, and 8) across San Quintín Bay. Plants were randomly collected by scuba diving using a 40-cm-diameter PVC corer (n = 3) and transported in separate plastic bags to the laboratory. DW of eelgrass aboveground biomass was evaluated after drying the samples at 60 °C to constant weight.
The monthly anomalies of the coastal upwelling index (UIA) during the sampling period for the nearest Pacific Ocean Station (30° N; 119° W) at San Quintín Bay were obtained from the National Oceanic and Atmospheric Administration (NOAA) (NOAA/NMFS Pacific Fisheries Environmental Laboratory), available online at http://upwell.pfeg.noaa.gov. The monthly offshore temperature-standardized anomalies during the study period were calculated from satellite images derived from MODIS-Aqua data (region: 116° W–115° W, 30° N–30° N) obtained from the National Aeronautics and Space Administration (NASA) and available online at http://gdata1.sci.gsfc.nasa.gov.
Biological and Temperature Control in pCO2 Calculations
Seawater pCO2 is affected by seasonal changes in temperature, physical processes (horizontal and vertical mixing), and biological processes (primary production, respiration, and calcium carbonate dissolution and precipitation) that influence total seawater CO2 concentrations in coastal systems (Takahashi et al. 2002; Barron et al. 2006). In order to remove the temperature effect from the observed pCO2 (pCO2 obs), the pCO2 obs values were normalized to 20.1 °C (pCO2 T mean), which was the mean annual area-averaged seawater temperature (T mean) measured in the San Quintín Bay system during the study period according to the method of Takahashi et al. (2002):
where pCO2 obs is the observed pCO2 at in situ temperature and T obs is the in situ mean monthly area-averaged seawater temperature measured in degrees Celsius. Changes in this parameter are indicative of variations in total CO2 due to the combined influence of horizontal mixing, vertical mixing, and biological processes, assuming that water temperature remains constant at 20.1 °C. Since San Quintín Bay is considered a well-mixed system, we assumed that the changes observed would result mostly from biological effects. The effect of biological factors on surface water pCO2 is represented in a given area by the seasonal amplitude of pCO2 values normalized to the mean annual temperature (pCO2 T mean), computed using the previous equation:
On the other hand, the effect of temperature on pCO2 was calculated as proposed by Takahashi et al. (2002):
where pCO2 T obs is the mean annual area-averaged observed pCO2 at in situ mean area-averaged seawater temperature for each month during the sampling period and pCO2mean is the mean annual area-averaged observed pCO2 (531 μatm).
The resulting values indicate the distribution of pCO2 that would be expected only from temperature changes if a parcel of water having the mean pCO2 of 538 μatm is subjected to seasonal temperature changes under isochemical conditions. The changes related to temperature were estimated as follows:
The importance of temperature (T) relative to biological parameters (B) can be expressed in terms of the following ratio (Takahashi et al. 2002):
Air–Water CO2 Flux Estimates
Air–water CO2 fluxes were calculated according to FCO2 = kα(ΔpCO2), where k is the CO2 transfer velocity, α is the CO2 solubility coefficient in seawater (Weiss 1974), and ΔpCO2 is the difference between the partial pressures of CO2 in water and air (ΔpCO2 = pCO2water − pCO2air). The pCO2air used to estimate ΔpCO2 was the monthly atmospheric data at the Pacific Ocean Station (POCN30) (30.0° N; 135.00° W), available from NOAA (http://www.esrl.noaa.gov/gmd/dv/site/site_table2.html), ranging from 397 to 399 μatm during the sampling period (November 2013 to November 2014). The parameter k was computed using the parameterization proposed by Ho et al. (2006) as a function of short-term steady wind velocities: k = 0.266u 2 10 (600/Sc)1/2, where “u 10” is the wind velocity at a height of 10 m, and “Sc” is the Schmidt number (Wanninkhof 1992). Monthly wind velocity (W) was obtained from a weather station located in San Quintín Bay.
Positive FCO2 values indicate gas fluxes from water to the atmosphere, while negative values indicate fluxes from the atmosphere to water. Since the calculation of pCO2water involved a standard error of ±20 μatm, the error associated with the FCO2 estimate is ±0.03 mmol C m−2 day−1 when wind velocity error was 8 %.
Whole-System Metabolism Calculation
The biogeochemical budget model according to Gordon et al. (1996) was used to estimate non-conservative DIC fluxes (ΔDIC). A one-box model was used for the San Quintín Bay system, following the procedure for nutrient budgets described by Camacho-Ibar et al. (2003). The whole-bay mean values (stations 2 to 10) and the outermost station (1) from the mouth of the bay value were used like the bay and the ocean end members, respectively. Station 1 was the only with merely oceanic signal, since the stations 2 to 10 were located near large eelgrass meadows and probably being more affected by biological processes. Net ecosystem production (NEP) (i.e., the difference between the gross primary production and the ecosystem respiration) was also computed from the non-conservative DIC fluxes to determine the system’s apparent net biogeochemical performance for carbon. Since ΔDIC values are attributed not only to the difference between production and respiration rates of organic matter (ΔDICo), but also to the net CO2 exchanges with the atmosphere (ΔDICg) and CaCO3 precipitation and dissolution (ΔDICc), NEP was estimated as follows (Gordon et al. 1996):
Using the expressions 6 and 7, NEP is defined as follows:
The contribution of CaCO3 precipitation and dissolution on NEP was assumed to be negligible (ΔDICc = 0) in the present study, considering the results and carbonate content in sediment samples observed by Ribas-Ribas et al. (2011) (ranging from 0 to 1.2 %) for San Quintín Bay. Otherwise to the best of our knowledge, no information on the magnitude of these processes is available for the bay.
Statistical Analyses
Temporal and spatial differences in hydrological and biogeochemical characteristics were analyzed using a non-parametric Kruskal–Wallis test, because the assumptions regarding normality and homoscedasticity criteria were not met for the data even when log transformed. The significance of correlations between environmental data and estimated parameters was tested using a Spearman’s correlation matrix. Statistical analyses were performed using the SIGMAPLOT 11 statistical package (Systat Software Inc., USA). The minimum significance level set for statistical difference was p < 0.05. Additionally, the combined uncertainties on the calculated non-conservative DIC fluxes were estimated by error propagation calculation.
Results
Seasonal Ocean–Bay Physicochemical Conditions
UIA in coastal waters off San Quintín Bay ranged between −70 and 5 m3 s−1·(100 m)−1 with an annual mean value of −23 ± 26 m3 s−1·(100 m)−1 (Table 1). Maximum negative UIA anomalies were observed from May to October 2014 (Fig. 2c), consistent with maximum temperature anomalies observed off San Quintín Bay in 2014 (Fig. 2d). Although the sampling period started under near-normal offshore SST (Fig. 2d), the SST anomaly exceeded 0.5 °C from May to November, peaking in September 2014. This month was anomalously 2 °C warmer versus the same month in the previous years observed in comparison with climatology data developed from July 2002 to May 2015. Temperature values at the mouth of the bay were higher than 20 °C during summer and fall 2014 (Fig. 2a).

Seasonal average values of a temperature (°C) and b salinity, for the three areas of San Quintín Bay; c monthly anomalies of the coastal upwelling index (UIA) (m3 s−1·(100 m)−1); and d monthly anomalies of the sea surface temperature (SST) during the study period
Seawater temperature in San Quintín Bay ranged between ∼15.0 and 26.0 °C during the study period, with minimum mean temperatures at the mouth (16.7 ± 1.7 °C) during spring months. Seawater temperatures were more than 4 °C lower at the mouth than at the head of both arms (Fig. 2a) and were higher toward the inner portion of bSQ in spring and summer. Salinity showed small seasonal differences, with an annual mean value of 34.33 ± 0.39 (Table 1). The minimum mean salinity was 33.35 ± 0.17 observed in the outer portion of the mouth in fall 2013, and the maximum mean salinity was 35.52 ± 0.98 observed in the inner part of bSQ in spring 2014 (Fig. 2b). Similar to temperature, salinity increased from the mouth of the bay toward the heads of bSQ and BFa, with increases of up to 5 units (Fig. 2b).
The timing of the water exchange varied seasonally for San Quintín Bay throughout the study period (Table 1). The annual mean residence time was 15 ± 6 days for 2014. The whole-system water exchange time estimated in this study was consistent with figures previously reported by Camacho-Ibar et al. (2003), with exchange rates of 12 ± 2 and 21 ± 9 days for summer and winter 2014, respectively. Wind velocity in San Quintín Bay revealed upwelling-favorable winds, with an annual mean of 3.1 ± 0.8 m s−1 during the study period and peak values observed from April to August (Table 1).
Figure 3 shows the time series of SST, pH, and %DO obtained from the oceanographic buoy located at the mouth of the bay. Seawater temperature ranged from 13 to 25 °C, fluctuating broadly from early spring to mid-fall and then decreasing toward the winter season. Temperatures were lowest in the spring, ranging from 13 to 19 °C and increasing steadily to reach a peak in September (Fig. 3a). The range of pH and %DO data during 2014 was 7.9 to 8.4 and 75 to 140 %, respectively, with means of 8.1 and 95 %, respectively and maxima between May and June 2014 (Fig. 3b, c). Pulses of cold water (<15 °C) were recorded in April, May, and early June, with occasional decreases in pH of up to ∼0.2 units and seawater %DO below 80 %, both of which suggest the input of water into the lagoon from an adjacent watershed with slightly different chemical properties.

Temporal variability in a temperature (°C), b pH, and c dissolved oxygen saturation (%) from February to December in 2014 as recorded by the oceanographic buoy located in the mouth of San Quintín Bay. Dashed lines show discrete sampling dates during the study period
Seasonal and Spatial Distribution of the Carbonate System and CO2 Fluxes
Figure 4a–d shows seasonal averages of DIC, TA, pH, and pCO2. DIC, TA, pH, and pCO2 recorded in San Quintín Bay were in the range of 1930 to 2330 μmol kg−1, 2210 to 2650 μmol kg−1, 7.75 to 8.18, and 382 to 919 μatm, respectively, with mean values of 2090 μmol kg−1, 2300 μmol kg−1, 7.95, and 538 μatm, respectively. Throughout the study period, DIC and TA concentrations were lower at the mouth of the bay than at the heads of BFa and bSQ, except in spring, where DIC values remained relatively similar along San Quintín Bay (Fig. 4a, b). Two contrasting environments were observed in the carbonate system parameters, mainly in spring (April–May) and summer (June–August) seasons, which showed significant differences (p < 0.05). In spring, low DIC values (<2000 μmol·kg−1) and low pCO2 values (<400 μatm) were consistent with high pH values (>8.1). In contrast, high DIC values (>2100 μmol·kg−1) and high pCO2 values (>700 μatm) were consistent with low pH values (<7.9) during the summer (Fig. 4a, c, d). On the other hand, seasonality of TA distribution showed a lower variability than seasonal changes of DIC, with relatively homogeneous concentrations between seasons.

Seasonal average CO2 system values for the three areas of San Quintín Bay during the study period: a dissolved inorganic carbon (μmol kg−1), b total alkalinity (μmol kg−1), c pHsw, and d partial pressure of CO2 (μatm)
As expected, DIC and TA normalized by salinity had an evaporation effect greatest in the innermost parts of San Quintín Bay. Both NDIC and NTA concentrations showed an increase from the mouth of the bay toward the arms, except in spring for NDIC, where a decrease was observed mainly at bSQ (Fig. 5a, b). These increasing concentrations were more notable in fall 2014 (>150 μmol kg−1), coinciding with the highest seawater temperatures recorded during the sampling period. The pCO2 air–water gradient (ΔpCO2) was positive for all seasons, except in spring at bSQ, where negative values were observed (Fig. 5c). Higher positive ΔpCO2 values (>400 μatm) were recorded in summer both in BFa and, to a lesser extent, in bSQ (Fig. 5c). During the anomalously warm 2014, FCO2 in San Quintín Bay ranged approximately between −4.9 and 22.6 mmol C m−2 day−1, with an annual mean of 3.3 ± 4.8 mmol C m−2 day−1 (Fig. 6a). Seasonal FCO2 averages were close to balance during the most of the year, except in summer months, when the CO2 exchange was higher (Fig. 6b). Overall, the whole system acted as a carbon source to the atmosphere, which peaked in July 2014 (15.1 ± 6.9 mmol C m−2 day−1) (Table 2).

Seasonal average values of a normalized dissolved inorganic carbon (μmol kg−1), b normalized total alkalinity (μmol kg−1), and c delta partial pressure of CO2 (μatm) for the three areas of San Quintín Bay during the study period

Spatial (a) and temporal (b) variability of air–water CO2 fluxes (mmol C m−2 day−1) during the sampling period in San Quintín Bay. Triangles and the dashed black line represent monthly wind velocity (m s−1). The zero line represents the equilibrium value of CO2 exchanges
Seasonal Variability of Eelgrass Biomass and Whole-System Metabolism
The eelgrass aboveground biomass varied seasonally between 70 and 250 g DW m−2 across the bay, with a peak in summer (238 ± 13 g DW m−2) and lower values in winter (81 ± 8 g DW m−2) (Fig. 7). The eelgrass biomass production rapidly increased early in the upwelling season, followed by a marked decrease in late summer toward November 2014, with average values below the 2013 fall–winter. To note, our eelgrass biomass estimate represents an underestimate of total biomass, as the biomass of roots and rhizomes was not considered.

Seasonal average values of aboveground biomass (g DW m−2) of seagrass Zostera marina in San Quintín Bay during the study period
In general, NEP values switched between net heterotrophy and net autotrophy during the study period, with values close to balance toward the end of 2014 (Fig. 8). NEP ranged approximately between −10 and 15 mmol C m−2 day−1, with negative values observed in December 2013 and July 2014 and positive values mainly during the upwelling season (April–June). Similar trend was observed for ΔDIC distribution, with a maximum peak in July (∼19 mmol C m−2 day−1) and minimum values estimated in spring and early summer. The annual mean NEP and ΔDIC estimates, with positive values (2.2 ± 7.1 mmol C m−2 day−1 and 1.1 ± 9.8 mmol C m−2 day−1, respectively), indicated that San Quintín Bay was a net autotrophic ecosystem during the study period (Table 2).

Non-conservative DIC fluxes (ΔDIC) using a one-box model and net ecosystem production (NEP) computed from DIC fluxes. ΔDIC and NEP estimates indicate that San Quintín Bay switched between net heterotrophy (organic respiration) and net autotrophy (inorganic uptake), approaching a balance from November 2013 to 2014
Discussion
Ocean–Bay Interactions and Biological Effects on Carbon Dynamics
The study period (November 2013 to November 2014) took place during the development of an unusual strongly positive temperature anomaly in the North Pacific Ocean, called The Blob, linked to a reduced cooling of the ocean during winter 2013–2014 (Hartmann 2014); this warm anomaly expanded and reached coastal waters in spring and summer 2014 (Bond et al. 2015). The anomalous oceanographic condition in 2014 markedly influenced the local dynamics of coastal waters off Baja California, where changes associated to the CCS wind patterns led to a drop in upwelling index values below average, as reflected by the monthly negative anomaly in the upwelling index (Fig. 2). These negative anomalous upwelling conditions occurred on shelf waters off San Quintín Bay, affecting the local ocean–bay interactions that largely influence and control biogeochemical processes within the system (Camacho-Ibar et al. 2003). The anomalous offshore SST condition was clearly observed in water adjacent to San Quintín Bay since May 2014, where the SST anomaly remained above 0.5 °C and reached a peak of 2 °C in mid-fall (Fig. 2d). The anomalous warm water was also observed in the surface temperature within the lagoon, both from discrete samples (Fig. 2a) and as recorded by the oceanographic buoy at the mouth of the bay (Fig. 3a). Considering that temperature is a suitable indicator of coastal upwelling waters flowing into San Quintín Bay (∼12 °C) (Farfán and Álvarez-Borrego 1983), no strong input of characteristic upwelling waters into the system was recorded during 2014.
In San Quintín Bay, where the upwelling index is closely associated to wind patterns (Ribas-Ribas et al. 2011), upwelling conditions determine the net fluxes of nutrients between the ocean and the lagoon, leading to strong changes in nutrient fluxes at the interannual, seasonal, and even single-upwelling-event timescales (Farfán and Álvarez-Borrego 1983; Camacho-Ibar et al. 2003; Hernández-Ayón et al. 2004). “Typical” nitrate concentrations under a “normal” upwelling condition are in the range between 3 and ∼13 μM (Hernández-Ayón et al. 2007a, b). Camacho-Ibar et al. (2003) reported nitrate concentrations of ∼0.5 to ∼0.9 μM in summer (weak upwelling), with a larger contribution of ammonium to the dissolved inorganic nitrogen (DIN) pool. On the other hand, Hernández-Ayón et al. (2004) described interannual fluctuations in nutrient dynamics in San Quintín Bay’s western arm during and after the 1997–1998 El Niño event. These authors observed that in Bahía Falsa (BFa), ammonium acted as a nitrogen source to the ocean during warm conditions, with significant benthic ammonium pulses (18–45 μM), while nitrate concentrations were low (0.8–2.0 μM). In our study, nitrate concentrations (data not shown) were approximately 0.6 ± 0.4 and 0.4 ± 0.4 μM on an annual basis and in spring–summer, respectively, similar to levels observed in San Quintín Bay in summer and warm El Niño conditions. These data suggest that the warm anomaly in 2014 maintained San Quintín Bay under a summer-like condition throughout the study period, characterized by weak upwelling events with higher seawater temperatures and depletion of incoming nutrients from adjacent shelf waters, relative to normal conditions. Under this scenario, nutrient recycling from sediments resulting from the decomposition of organic matter by in situ biota (via benthic fluxes) likely plays a key role as an autochthonous source of nitrogen and inorganic carbon for N-limited primary producer communities. The larger variations observed in pH and %DO from the buoy, particularly during the upwelling season (April–June), are consistent with this assumption, reflecting changes in biological production probably associated with a higher nutrient availability within the lagoon (Fig. 3b, c).
The distribution of carbon system parameters also reflects the importance of internal biogeochemical processes in San Quintín Bay in response to the anomalous oceanographic condition, as clearly seen in spring and summer (Fig. 4a–d). The internal supply of nutrients apparently linked to benthic fluxes during spring possibly stimulated a net autotrophy (i.e., primary production) that led to the decrease in DIC and pCO2, with the consequent rise in pH (Fig. 4a, c). A notably NDIC decrease at the inner arms of the bay was observed in spring months (Fig. 5a), which can be attributed to degasification and carbon uptake by macrophyte photosynthesis, previously reported by Ribas-Ribas et al. (2011). Higher DIC and pCO2 coupled with lower pH during summer jointly suggest the predominance of a net heterotrophy (aerobic respiration), probably enhanced by the rise in water temperature and higher autochthonous organic matter levels produced previously. These results are consistent with the seasonality of pCO2 at in situ temperatures, similar to the temperature-normalized pCO2 (Fig. 9), and clearly indicate the dominance of biological processes on pCO2 in the whole system (T/B = 0.3), which is typical of areas with marked seasonal changes in biological activity (Takahashi et al. 2002). Our findings suggest that despite the warm anomaly condition, temperature was not the driving force that controlled the pCO2 distribution in San Quintín Bay, with just a minor effect on the magnitude of air–water CO2 fluxes during the study period. However, temperature fluctuations may directly and indirectly affect net production and respiration rates in coastal ecosystems (e.g., decomposition rates, enzyme-catalyzed cell metabolism) (Testa et al. 2013). Therefore, the offshore anomalous SST condition observed in the lagoon could have directly influenced the magnitude of these biological activities.

Observed pCO2 (pCO2 obs, black circle line), temperature-normalized pCO2 at 20.1 °C (pCO2@T mean, white circle dashed line), and the mean area-averaged pCO2 at in situ temperature through the year (pCO2mean@T obs, black triangle dashed line)
The CO2 exchange estimated during 2014 resulted in CO2 sea–air outgassing mainly during the summer season, while the rest of the year was slight or close to balance (Table 2). Our findings are consistent with other studies in coastal ecosystems that showed overall annual air–water CO2 fluxes close to balance, with higher fluxes in summer (Chen and Borges 2009). The higher CO2 flux observed in our study during the upwelling season (∼5 mmol C m−2 day−1) was far lower than fluxes reported for San Quintín Bay during intense upwelling events (∼50 mmol C m−2 day−1) by Ribas-Ribas et al. (2011). The low DIC concentrations at the mouth of the bay (∼2000 μmol kg−1), similar to normal oceanic seawater values, in comparison with intense upwelling measurements (∼2150 μmol kg−1, Ribas-Ribas et al. 2011), suggest the absence of upwelling waters largely influenced by offshore conditions. Given the previously mentioned data, we propose that the spatial and temporal variability in net sea–air CO2 fluxes estimated in San Quintín Bay during 2014 probably reflected the biogeochemical carbon transformations driven primarily by benthic ecosystems.
Carbon Balance and NEP Changes during 2014
The tendency in net ecosystem metabolism (system status) either toward export (net autotrophic) or import (net heterotrophic), as well as in dissolved and particulate matter, represents the net effect of physical and biological processes that contribute to organic carbon balance in coastal aquatic ecosystems (Testa et al. 2013). On an annual basis, the metabolic balance of Mediterranean-climate coastal lagoons located within the CCS upwelling system are considered as net heterotrophic systems, largely associated with the tidal exchange of adjacent shelf waters and coastal systems (Smith and Hollibaugh 1997; Camacho-Ibar et al. 2003; Delgadillo-Hinojosa et al. 2008). In San Quintín Bay, while most studies have reported a net heterotrophic status explained by the import of labile phytoplanktonic carbon which fuels ecosystem respiration (e.g. Hernández-Ayón et al. 2004; Ibarra-Obando et al. 2007), net autotrophic conditions have been observed during intense upwelling events (Ribas-Ribas et al. 2011). The NEP and carbon balance (ΔDIC) estimates reported here revealed an evident seasonality in the system’s status, which shifts from net heterotrophy in winter and summer to net autotrophy mainly during the upwelling season (Fig. 8). This trend is consistent with previous studies in San Quintín Bay (e.g., Camacho-Ibar et al. 2003; Ribas-Ribas et al. 2011) and is in general agreement with observations in many temperate estuaries and shallow coastal lagoons, where NEP peaks in spring influenced by annual light and temperature cycles (McGlathery et al. 2001; Testa et al. 2013).
Overall, NEP ranges in 2014 were narrower than those reported in earlier studies for this site (Camacho-Ibar et al. 2003; Hernández-Ayón et al. 2004; Ribas-Ribas et al. 2011). Net heterotrophy values estimated during our study were slightly lower to net whole-system estimates previously reported under normal conditions in San Quintín Bay (∼−14 mmol C m−2 day−1) (Camacho-Ibar et al. 2003; Ibarra-Obando et al. 2004), while net autotrophy was between three and six times lower than values reported by Ribas-Ribas et al. (2011) during intense upwelling conditions. The higher net autotrophy reported by Ribas-Ribas et al. (2011) can be explained by the increased ecosystem productivity sensitive to external inorganic nutrient inputs and high CO2 concentrations from upwelled waters. In contrast, the temperature rise in 2014 probably favored respiration within the system (Hopkinson and Smith 2005), releasing inorganic carbon and nutrients that stimulated primary production. This production, in turn, yielded organic matter that fueled benthic respiration and internal supply of “recycled” nutrients for primary producer communities. Under this environment, the ecosystem status eventually approaches a balance condition (NEP = 0), where the periods of metabolic imbalance are influenced by the stimulated production or consumption of organic matter, as observed in Fig. 8. On an annual basis, contrary to what would be expected for San Quintín Bay, our positive NEP estimates suggest that primary production dominated the gross metabolism, apparently linked to the regional anomalous condition in 2014 (Table 2). This behavior could be explain due to the following: (1) limited external supply of organic material to account for an excess of respiration (net heterotrophy) over photosynthetic activity and (2) high benthic primary production rates (over respiration rates) fueled by carbon and nutrient release during remineralization within the system.
Benthic communities (i.e., seagrass beds, macroalgae, and benthic microalgae) are often highly productive components that dominate shallow marine ecosystems and play a key role in regulating the turnover rate of carbon and nutrients (McGlathery et al. 2007). San Quintín Bay is a coastal lagoon characterized by extensive eelgrass meadows that largely contribute to the metabolism of the benthic ecosystem, which is the key driver of the metabolic signal (70 %) (Ibarra-Obando et al. 2004). At low nutrient concentrations across the water column (summer-like condition), Z. marina is able to efficiently uptake sedimentary nutrient pools via underground biomass to support its production rates (Sandoval-Gil et al. 2015). Positively increased NEP values coincided with the onset of the eelgrass growth season (Fig. 7) and are associated with seasonal variations in both water temperature and light levels on eelgrass leaves (Cabello-Pasini et al. 2003; Sandoval-Gil et al. 2015). Aboveground eelgrass biomass dropped markedly from July to November 2014; this produced relatively large inputs of residual organic matter available in the water column and local sediments, which stimulated heterotrophy (Testa et al. 2013). This biomass drop resulted in average values below those of the previous fall–winter, probably because of the anomalously warm environment condition (Fig. 7). Similar to seagrasses, the nutrient storage capacity of macroalgae supports high growth rates with low-to-moderate nutrient availability, which contribute organic matter inputs to fuel respiration processes (McGlathery et al. 2013). As expected, a positive ΔDIC in mid-summer indicates that the system displayed high CO2 levels (Fig. 8), which led to a CO2 efflux from the decomposition of organic matter. Although both eelgrasses and macroalgae have been reported as important temporary carbon and DIN sinks at San Quintín Bay (Zertuche-González et al. 2009; Sandoval-Gil et al. 2015), organic matter bioavailability (i.e., labile biomass) drives the metabolic rates and nutrient budgets in coastal ecosystems (Testa et al. 2013). Considering that breakdown rates of macroalgal debris are generally higher (days-weeks) versus those of seagrass detritus (weeks–months) (McGlathery et al. 2013), macroalgae tissues would make a major potential source of DIC to sustain the FCO2 values observed in summer.
Based on the seasonal biomass records for the three most abundant macroalgae, i.e., Ulva sp., Gracilaria sp., and Chondracanthus squarrulosus in San Quintín Bay in 2014 (Table 3), the organic matter derived from macroalgae biomass from April to August represented approximately 12, 72, and 100 %, respectively (136 t of carbon) (Pacheco-Ruíz, personal communication). Assuming that most macroalgal detritus remained within the system, carbon fluxes associated with them would be sufficient to support respiration rates between 0.6 and 8.6 mmol C m−2 day−1—which could explain the peak FCO2 levels observed in mid-summer (Fig. 6b)—which contribute around to 45 % of the CO2 released into the water column (Fig. 8). The seasonal variation patterns in carbon balance in San Quintín Bay are seemingly linked, at least partially, to macroalgal communities that may temporarily stimulate the net heterotrophic metabolism of planktonic communities. However, other processes that occur both within sediments and at the water–sediment interface (e.g., microphytobenthos metabolism or anaerobic respiration) could be contributing factors on carbon dynamics in this coastal system as well (Ibarra-Obando et al. 2004).
The increased NTA coupled with higher NDIC concentrations, which were observed in the arms of the bay, mostly in winter and fall (Fig. 5), may reflect a major role of these sedimentary exchanges that affect directly or indirectly these quantities in shallow coastal areas (Thomas et al. 2009). Since carbon is added to seawater, as a consequence to sedimentary-improved regeneration of organic materials attributed to temperature and macrophyte production in San Quintín Bay, an increase in DIC could be expected for these seasons. On the other hand, accumulation of this organic matter supply has the potential to stimulate benthic, anaerobic degradation with a large enhancement of total alkalinity release from surface sediments (Thomas et al. 2009). In San Quintín Bay, benthic denitrification (remineralization of organic matter using nitrate as electron acceptor) represents a key sedimentary process coupled with benthic and water column nitrification (the aerobic oxidation of ammonium to nitrate) (Camacho-Ibar et al. 2003). This anaerobic process represents a loss of bioavailable water column nitrogen, reflecting low nitrate concentrations during summer and winter conditions (Camacho-Ibar et al. 2003). Moreover, since denitrification is an irreversible process (i.e., permanently remove reactive nitrogen from ecosystem), contributions of released ammonia by this anaerobic organic matter degradation constitute a net gain of the generated TA, which is of particular importance in shallow coastal areas (Thomas et al. 2009). This is evident, for example, in the observed values of TA in bSQ in fall 2014 (Fig. 5b), where the enhanced primary production from late winter until late summer stimulated anaerobic processes within the sediments, releasing large amounts of TA. Accordingly, the effect of DIC added by decaying biological activity is buffered by this increasing release of TA that prevented a strong pH decline and that maintained pCO2 values close to balance (Fig. 5c). As mentioned before, these results support the conclusion that benthic release of recycled carbon and nitrogen pools is probably fueled by biological activity in San Quintín Bay during the warm anomaly in 2014 and may explain the increased TA and DIC values observed at the water column toward the arms of the bay. Further processes that may enhance TA in coastal environments are the production of organic bases from microalgae or the dissolution of calcium carbonate. Early studies in San Quintín Bay have already reported the contribution of organic bases from microalgae to the titration alkalinity (Hernández-Ayón et al. 2007b; Ribas-Ribas et al. 2011). However, the calculated TA from pH (buoy data), salinity, and DIC measures suggests that TA values are not affected by organic alkalinity, with changes around ±7 μmol kg−1 from the mouth toward the arms of the bay. On the other hand, in benthic environments dominated by seagrass meadows, CaCO3 dynamics can also play an important role in carbon balance variations (Barron et al. 2006). Nevertheless, recent observations of the carbonate content in the sediments of San Quintín Bay by Ribas-Ribas et al. (2011) suggest that inorganic carbon precipitation or dissolution does not contribute significantly to internal DIC biogeochemical fluxes in this system. It can be concluded that benthic fluxes on surface sediments probably play a key regulatory role in the whole ecosystem through their storage and transformation capacity of organic matter and nutrients, which is favored by the anomalously warm environmental condition. So, a better understanding is needed about sedimentary processes and the role of benthic habitats to integrate the biogeochemical response of carbon dynamics in these environments to climate variations at a regional scale.
Summary
The atypical oceanographic condition developed in the North Pacific during 2014 led to a negative anomaly of the upwelling index in coastal waters off Baja California, which markedly influenced the local dynamics within this region. As a result, we observed a continuous summer-like season (weak upwelling condition) in San Quintín Bay during the study period (November 2013 to 2014). Despite the warm anomaly, temperature was not the key driver of the CO2 outgassing in this shallow system, largely influenced by the high seasonal variability in biological activity. San Quintín Bay acts as a source of CO2 to the atmosphere in 2014 (3.3 ± 4.8 mmol C m−2 day−1) with the higher positive fluxes mainly observed in summer months (9.0 ± 5.3 mmol C m−2 day−1). The spatial and temporal variability estimated in net air–water CO2 fluxes during 2014 appears to reflect a “baseline signal” of biogeochemical carbon transformations driven by an ecosystem dominated by macrophytes. On an annual basis, San Quintín Bay was a net autotrophic system during the anomalous warming, with an annual average of 2.2 ± 7.1 mmol C m−2 day−1. However, the net ecosystem status switched seasonally between net heterotrophy in winter and summer and net autotrophy during the upwelling season (April–June), apparently influenced by the life cycles of the dominant primary producers in response to annual light and temperature cycles.
The warm anomaly favored respiration rates that released inorganic carbon and dissolved nutrients, stimulating organic matter production that fueled benthic respiration and a local supply of recycled nutrients for N-limited primary producers. The seasonal biomass production of macroalgal communities was a potential source of CO2 during the study period and probably was the primary contributor to the net ecosystem metabolism in summer. Clearly, the data recorded in San Quintín Bay in 2014 suggest that macroalgal blooms are a potential local source of organic matter to fuel ecosystem respiration; hence, this factor should be taken into account in future investigations related to biogeochemical fluxes and ecosystem status in this shallow system. Eelgrass detritus contributed, to a lesser extent, to the summer net heterotrophy observed; however, in an annual cycle, the net production of Z. marina was a key contributor to the whole-ecosystem metabolism in San Quintín Bay. Since the upwelling condition may influence the magnitude of the air–water CO2 fluxes, our results clearly indicated that San Quintín Bay is a net source of carbon to the atmosphere regardless of the adjacent oceanic conditions. Our study sheds light on the relative importance of physical processes, biological communities, and environmental conditions in response to carbon dynamics and NEP in a shallow coastal ecosystem under a regional climate anomaly and provides potentially relevant information about global climate change scenarios in similar coastal systems.
References
Atkinson, M.J., and S.V. Smith. 1983. C:N:P ratios of benthic marine plants. Limnology and Oceanography 28: 568–574. doi:10.4319/lo.1983.28.3.0568.
Barrón, C., C.M. Duarte, M. Frankignoulle, and A.V. Borges. 2006. Organic carbon metabolism and carbonate dynamics in a Mediterranean seagrass (Posidonia oceanica) meadow. Estuaries and Coasts 29(3): 417–426.
Bauer, J.E., W.-J. Cai, P.A. Raymond, T.S. Bianchi, C.S. Hopkinson, and P.A.G. Regnier. 2013. The changing carbon cycle of the coastal ocean. Nature 504(7478): 61–70. doi:10.1038/nature12857.
Bond, N.A., M.F. Cronin, H. Freeland, and N. Mantua. 2015. Causes and impacts of the 2014 warm anomaly in the NE Pacific. Geophysical Research Letters. doi:10.1002/2015GL063306.
Borges, A.V. 2005. Do we have enough pieces of the jigsaw to integrate CO2 fluxes in the Coastal Ocean? Estuaries 28(1): 3–27.
Cabello-Pasini, A., R. Muñiz-Salazar, and D.H. Ward. 2003. Annual variations of biomass and photosynthesis in Zostera marina at its southern end of distribution in the North Pacific. Aquatic Botany 76(1): 31–47. doi:10.1016/S0304-3770(03)00012-3.
Camacho-Ibar, V.F., J.D. Carriquiry, and S.V. Smith. 2003. Non-conservative P and N fluxes and net ecosystem production in San Quintín Bay, Mexico. Estuaries 26(5): 1220–1237. doi:10.1007/BF02803626.
Checkley, D.M., and J.A. Barth. 2009. Patterns and processes in the California current system. Progress in Oceanography 83(1–4): 49–64. doi:10.1016/j.pocean.2009.07.028.
Chen, C.T.A., and A.V. Borges. 2009. Reconciling opposing views on carbon cycling in the coastal ocean: continental shelves as sinks and near-shore ecosystems as sources of atmospheric CO2. Deep Sea Research Part II: Topical Studies in Oceanography 56(8–10): 578–590.
Chen, C.T.A., T.H. Huang, Y.C. Chen, Y. Bai, X. He, and Y. Kang. 2013. Air-sea exchanges of coin the world’s coastal seas. Biogeosciences 10(10): 6509–6544. doi:10.5194/bg-10-6509-2013.
Delgadillo-Hinojosa, F., A. Zirino, O. Holm-Hansen, J.M. Hernández-Ayón, T.J. Boyd, B. Chadwick, and I. Rivera-Duarte. 2008. Dissolved nutrient balance and net ecosystem metabolism in a Mediterranean-climate coastal lagoon: San Diego Bay. Estuarine, Coastal and Shelf Sci 76(3): 594–607. doi:10.1016/j.ecss.2007.07.032.
Dickson, A.G., and F.J. Millero. 1987. A comparison of the equilibrium constants for the dissociation of carbonic acid in seawater media. Deep Sea Research Part A 34(10): 1733–1743.
Dickson, A.G., J.D. Afghan, and G.C. Anderson. 2003. Reference materials for oceanic CO2 analysis: a method for the certification of total alkalinity. Marine Chemistry 80(2–3): 185–197. doi:10.1016/S0304-4203(02)00133-0.
Doney, S.C., B. Tilbrook, S. Roy, N. Metzl, C. Le Quéré, M. Hood, and D. Bakker. 2009. Surface-ocean CO2 variability and vulnerability. Deep Sea Research Part II: Topical Studies in Oceanography 56(8–10): 504–511. doi:10.1016/j.dsr2.2008.12.016.
Durazo, R. 2009. Climate and upper ocean variability off Baja California, Mexico: 1997-2008. Progress in Oceanography 83(1–4): 361–368. doi:10.1016/j.pocean.2009.07.043.
Farfán, B.C., and S. Álvarez-Borrego. 1983. Variability and fluxes of nitrogen and organic carbon at the mouth of a coastal lagoon. Estuarine, Coastal and Shelf Sci. 17: 599–612.
Feely, R.A., C.L. Sabine, J.M. Hernandez-Ayon, D. Ianson, and B. Hales. 2008. Evidence for upwelling of corrosive “acidified” water onto the continental shelf. Science 320(5882): 1490–1492. doi:10.1126/science.1155676.
García-Reyes, M., and J.L. Largier. 2012. Seasonality of coastal upwelling off central and northern California: new insights, including temporal and spatial variability. Journal of Geophysical Research 117(C3): 1–17. doi:10.1029/2011JC007629.
Gordon, D.C. Jr., P.R. Boudreau, K.H. Mann, J.E. Ong, W.L. Silvert, S.V. Smith, G. Wattayakorn, F. Wulff, and T. Yanagi. 1996. LOICZ biogeochemical modeling guidelines. In LOICZ/R&S/95–5. The Netherlands: Land Ocean Interactions in the Coastal Zone, Texel.
Gypens, N., G. Lacroix, C. Lancelot, and A.V. Borges. 2011. Seasonal and inter-annual variability of air-sea CO2 fluxes and seawater carbonate chemistry in the Southern North Sea. Progress in Oceanography 88(1–4): 59–77. doi:10.1016/j.pocean.2010.11.004.
Hartmann, D.F. 2015. Pacific sea surface temperature and the winter of 2014. Geophysical Research Letters 42. doi:10.1002/2015GL063083.
Hernández-Ayón, J.M., S.L. Belli, and A. Zirino. 1999. pH, alkalinity and total CO2 in coastal seawater by potentiometric titration with a difference derivative readout. Anal. Chem. Acta 394(1): 101–108. doi:10.1016/S0003-2670(99)00207-X.
Hernández-Ayón, J.M., M.S. Galindo-Bect, V. Camacho-Ibar, Z. García-Esquivel, and F. Ley-Lou. 2004. Dinámica de los nutrientes en el brazo oeste de Bahía San Quintín, Baja California, México durante y después de El Niño 1997/1998. Ciencias Marinas 30(1 A): 119–132.
Hernández-Ayón, J. M., Camacho-Ibar, V. F., Mejía-Trejo, A., & Cabello-Pasini, A. 2007a. Variabilidad del CO2 total durante eventos de surgencia en Bahía San Quintín, Baja California, México. Book: Carbono en ecosistemas acuáticos de México, 187–200. doi:http://www.inecc.gob.mx/publicaciones/libros/531/cap12.pdf
Hernández-Ayón, J.M., A. Zirino, A.G. Dickson, T. Camiro-Vargas, and E. Valenzuela-Espinoza. 2007b. Estimating the contribution of organic bases from microalgae to the titration alkalinity in coastal seawaters. Limnology and Oceanography: Methods 5: 225e232.
Ho, D.T., C.S. Law, M.J. Smith, P. Schlosser, M. Harvey, and P. Hill. 2006. Measurements of air-sea gas exchange at high wind speeds in the Southern Ocean: implications for global parameterizations. Geophysical Research Letters 33: L16611. doi:10.1029/2006GL026817.
Hopkinson, C.S., and E.M. Smith. 2005. Estuarine respiration: an overview of benthic, pelagic, and whole system respiration. In Respiration in aquatic ecosystems, ed. P. del Giorgio, and PJledb Williams, 122–146. Oxford: Oxford University Press.
Ibarra-Obando, S.E., S.V. Smith, M. Poumian-Tapia, V. Camacho-Ibar, J.D. Carriquiry, and M. Montes-Hugo. 2004. Benthic metabolism in San Quintín Bay, Baja California Mexico. Mar. Ecol. Prog. Ser. 283: 99–112. doi:10.3354/meps283099.
Ibarra-Obando, S. E., Solana-Arellano, E., Poumian-Tapia, M., de la Torre, B. H., & Gaxiola-Castro, G. 2007. El Papel de Zostera marina en el Ciclo del Carbono en Bahía San Quintín, Baja California. Book: Carbono en ecosistemas acuáticos de México, 201–213.
Lachkar, Z., and N. Gruber. 2013. Response of biological production and air-sea CO2 fluxes to upwelling intensification in the California and Canary Current Systems. Journal of Marine Systems 109-110: 149–160. doi:10.1016/j.jmarsys.2012.04.003.
Lewis, E. & Wallace, D. W. R. 1998. Program developed for CO2 system calculations. Oak Ridge National Laboratory. ORNL/CDIAC-105.
Macías, D., P.J.S. Franks, M.D. Ohman, and M.R. Landry. 2012. Modeling the effects of coastal wind- and wind-stress curl-driven upwellings on plankton dynamics in the Southern California current system. Journal of Marine Systems 94: 107–119. doi:10.1016/j.jmarsys.2011.11.011.
McGlathery, K.J., I.C. Anderson, and A.C. Tyler. 2001. Magnitude and variability of benthic and pelagic metabolism in a temperate coastal lagoon. Marine Ecology Progress Series 216: 1–15.
McGlathery, K.J., K. Sundbäck, and I.C. Anderson. 2007. Eutrophication patterns in shallow coastal bays and lagoons. Marine Ecology Progress Series 348: 1–18.
McGlathery, K.J., K. Sundbäck, and P. Fong. 2013. Estuarine benthic algae. In Estuarine Ecology, Second edn, ed. J.W. Day, B.C. Crump, W.M. Kemp, and A. Yáñez-Arancibia. Hoboken, NJ: John Wiley & Sons, Inc.. doi:10.1002/9781118412787.ch8.
Mehrbach, C., C.H. Culberson, J.E. Hawley, and R.M. Pytkowicz. 1973. Measurement of the apparent dissociation constants of carbonic acid in seawater at atmospheric pressure. Limnology and Oceanography 18(6): 897e907.
Muñoz-Anderson, M.A., J.R. Lara-Lara, S. Alvarez-Borrego, C. Bazan-Guzman, and M. de la Cruz-Orozco. 2015. Water–air carbon fluxes in the coastal upwelling zone off northern Baja California. Ciencias Marinas 41(2): 157–168.
Reimer, J., R. Vargas, S.V. Smith, R. Lara-Lara, G. Gaxiola-Castro, J.M. Hernández-Ayón, A. Castro, M. Escoto-Rodríguez, and J. Martínez-Osuna. 2013. Air-sea CO2 fluxes in the near-shore and intertidal zones influenced by the California Current. Journal of Geophysical Research: Oceans Vol. 118: 1–16.
Ribas-Ribas, M., J.M. Hernández-Ayón, V.F. Camacho-Ibar, A. Cabello-Pasini, A. Mejia-Trejo, R. Durazo, et al. 2011. Effects of upwelling, tides and biological processes on the inorganic carbon system of a coastal lagoon in Baja California. Estuarine, Coastal and Shelf Sci 95(4): 367–376. doi:10.1016/j.ecss.2011.09.017.
Sabine, C.L., R.A. Feely, N. Gruber, R.M. Key, K. Lee, L. Bullister J., R. Wanninkhof, C.S. Wong, D.W.R. Wallace, B. Tilbrook, et al. 2004. The oceanic sink for anthropogenic CO2. Science 305(5682): 367–371.
Smith, S.V., and J.T. Hollibaugh. 1997. Annual cycle and interannual variability of ecosystem metabolism in a temperate climate embayment. Ecological Monographs 67(4): 509e533.
Takahashi, T., S.C. Sutherland, C. Sweeney, A. Poisson, N. Metzl, B. Tilbrook, and Y. Nojiri. 2002. Global sea-air CO2 flux based on climatological surface ocean pCO2, and seasonal biological and temperature effects. Deep Sea Research Part II: Topical Studies in Oceanography 49(9–10): 1601–1622. doi:10.1016/S0967-0645(02)00003-6.
Takahashi, T., S.C. Sutherland, R. Wanninkhof, C. Sweeney, R.A. Feely, D.W. Chipman, and H.J.W. de Baar. 2009. Climatological mean and decadal change in surface ocean pCO2, and net sea-air CO2 flux over the global oceans. Deep Sea Research Part II: Topical Studies in Oceanography 56(8–10): 554–577. doi:10.1016/j.dsr2.2008.12.009.
Testa, J.M., W.M. Kemp, C.S. Hopkinson, and S.V. Smith. 2013. Ecosystem metabolism. In Estuarine Ecology, Second edn, ed. J.W. Day, B.C. Crump, W.M. Kemp, and A. Yáñez-Arancibia. Hoboken, NJ: Wiley, Inc.. doi:10.1002/9781118412787.ch15.
Thomas, H., L.-S. Schiettecatte, K. Suykens, Y.J.M. Kone, E.H. Shad-wick, A.E.F. Prowe, Y. Bozec, H.J.W. de Baar, and A.V. Borges. 2009. Enhanced ocean carbon storage from anaerobic alkalinity generation in coastal sediments. Biogeosciences 6: 267–274. doi:10.5194/bg-6-267-2009.
Wanninkhof, R. 1992. Relationship between wind speed and gas exchange over the ocean. Journal of Geophysical Research 97(C5): 7373–7382.
Ward, D.H., T.L. Tibbitts, E. Carrera-González, and R. Kempka. 2004. Use of digital multispectral videography to assess seagrass distribution in San Quintín Bay, Baja California, Mexico. Ciencias Marinas 30(1A): 47–60.
Weiss, R.F. 1974. Carbon dioxide in water and seawater: the solubility of a non ideal gas. Marine Chemistry 2(3): 203–215.
Zertuche-González, J.A., V.F. Camacho-Ibar, I. Pacheco-Ruíz, A. Cabello-Pasini, L.A. Galindo-Bect, J.M. Guzmán-Calderón, and J. Espinoza-Avalos. 2009. The role of Ulva spp. as a temporary nutrient sink in a coastal lagoon with oyster cultivation and upwelling influence. Journal of Applied Phycology 21(6): 729–736. doi:10.1007/s10811-009-9408-y.
Acknowledgments
The authors wish to thank SEP-CONACYT for the support given for this work through the UABC project Estudio Integral del Ciclo del Nitrógeno en Bahía Falsa, Baja California (ref. no. 10412). We thank the cooperation of the Nautilus hatchery workers from San Quintín for their support in the field, and we appreciate the significant assistance of Dulce Magali López, Nevia Alfaro, Julieta Hernández, and Erika Santacruz during the field sampling and the laboratory analysis and the comments that improved the manuscript. This paper was pleasantly improved and strengthened by the constructive comments of various anonymous reviewers. María Elena Sánchez-Salazar contributed with the editing of the English language.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Alberto Vieira Borges
Rights and permissions
About this article

Cite this article
Ávila-López, M.C., Hernández-Ayón, J.M., Camacho-Ibar, V.F. et al. Air–Water CO2 Fluxes and Net Ecosystem Production Changes in a Baja California Coastal Lagoon During the Anomalous North Pacific Warm Condition. Estuaries and Coasts 40, 792–806 (2017). https://doi.org/10.1007/s12237-016-0178-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-016-0178-x