
Abstract
Contemporary deposition (artificial marker horizon, 3.5 years) and long-term accumulation rates (210Pb profiles, ~150 years) of sediment and associated carbon (C), nitrogen (N), and phosphorus (P) were measured in wetlands along the tidal Savannah and Waccamaw rivers in the southeastern USA. Four sites along each river spanned an upstream-to-downstream salinification gradient, from upriver tidal freshwater forested wetland (TFFW), through moderately and highly salt-impacted forested wetlands, to oligohaline marsh downriver. Contemporary deposition rates (sediment, C, N, and P) were greatest in oligohaline marsh and lowest in TFFW along both rivers. Greater rates of deposition in oligohaline and salt-stressed forested wetlands were associated with a shift to greater clay and metal content that is likely associated with a change from low availability of watershed-derived sediment to TFFW and to greater availability of a coastal sediment source to oligohaline wetlands. Long-term accumulation rates along the Waccamaw River had the opposite spatial pattern compared to contemporary deposition, with greater rates in TFFW that declined to oligohaline marsh. Long-term sediment and elemental mass accumulation rates also were 3–9× lower than contemporary deposition rates. In comparison to other studies, sediment and associated nutrient accumulation in TFFW are lower than downriver/estuarine freshwater, oligohaline, and salt marshes, suggesting a reduced capacity for surface sedimentation (short-term) as well as shallow soil processes (long-term sedimentation) to offset sea level rise in TFFW. Nonetheless, their potentially large spatial extent suggests that TFFW have a large impact on the transport and fate of sediment and nutrients in tidal rivers and estuaries.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Tidal freshwater forested wetlands (TFFW), also known as tidal swamps, occur at the interface of watersheds and estuaries. Non-tidal freshwater floodplains occur upriver from TFFW, and downriver are typically a progression of tidal herbaceous wetlands along gradients of increasing salinity towards the coastal zone (Odum 1988). TFFW floodplain ecosystems are extensive, likely occupying more land surface than tidal freshwater marshes in the USA (Field et al. 1991). Their hydrogeomorphic position makes them sensitive to both coastal processes, such as higher water and salinification due to sea level rise and human modifications to estuaries and tidal rivers, as well as watershed processes that influence freshwater discharge and sediment availability (Day et al. 2007). Yet TFFW have received relatively little study compared to other tidal and non-tidal wetlands (Conner et al. 2007). Greater understanding of the rates and controls on sediment trapping by TFFW along salinification gradients is essential for predicting the resilience of these ecosystems to sea level rise (Craft 2012; Ensign et al. 2014a, b) as well as their role in sediment and associated elemental budgets of estuaries (Noe et al. 2013).
The role of TFFW in coastal nutrient retention and transformations is relatively less known compared to that of other tidal wetlands (Anderson and Lockaby 2007). Nutrient inputs from sedimentation are critical to wetland ecosystem dynamics and biogeochemistry (Johnston 1991; Noe and Hupp 2009; Wolf et al. 2013). Sediment deposition can be the dominant source of N and P inputs compared to dissolved inorganic nutrient loading to the soil surface; sediment deposition is 90 and 66 % of total nitrogen (N) and phosphorus (P) inputs, respectively, to TFFW and oligohaline wetlands in the southeastern USA (Noe et al. 2013; this study). Nutrient trapping by sedimentation also can represent a large landscape-scale removal flux by TFFW, which is often spatially extensive where it occurs. Long-term N removal by sediment accumulation in freshwater, brackish, and salt marshes (downstream of TFFW) represents from 13 to 32 % of the watershed load of three estuaries (Loomis and Craft 2010). Furthermore, varying inputs of N and P from deposition could change the rates of biogeochemical cycling in TFFW (Hopkinson 1992; Wolf et al. 2013). Finally, salinification increases soil N and P mineralization in tidal freshwater wetlands, which could stimulate primary productivity or export nutrients to downriver estuaries (Weston et al. 2006; Jun et al. 2013; Ardón et al. 2013; Noe et al. 2013; Gao et al. 2014).
Sedimentation rates are generally lower in TFFW and tidal freshwater marshes compared to those of oligohaline marshes across the mid-Atlantic and Southeastern Atlantic coasts. Deposition rates in the upriver reaches of TFFW are generally greater than those of downriver reaches of TFFW as watershed sediment delivered to the tidal zone is trapped by upriver TFFW (Kroes et al. 2007; Ensign et al. 2014b; Hupp et al. 2015). Increased sediment deposition farther downriver of TFFW in oligohaline marsh coincides with increases in herbaceous biomass that may lead to a positive feedback mechanism between plant biomass and sediment trapping (Ensign et al. 2014a). Channel hydraulics also increase sediment concentrations in lower, oligohaline tidal rivers (Ensign et al. 2014b). Proximity to an estuarine turbidity maximum (ETM) in oligohaline reaches of estuaries also leads to enhanced wetland sediment deposition and accumulation rates (Darke and Megonigal 2003; Loomis and Craft 2010). Sources of sediment to tidal wetlands should change along longitudinal (upriver-downriver) gradients of tidal rivers based on magnitude and proximity of watershed vs. estuarine sediment (Windom et al. 1971). The rates and processes of sediment trapping by TFFW are essential for predicting the long-term fate of these ecosystems to sea level rise—TFFW vertical deposition and accretion rates are typically lower than local relative sea level rise (Craft 2012; Ensign et al. 2014a, b). Finally, sedimentation rates differ across temporal scales due to changes in sediment availability over time, sediment trapping rates, sea level rise, and post-depositional erosion, decomposition, and compaction of sediments (Craft 2012; Breithaupt et al. 2014; Hupp et al. 2015). Understanding the spatial and temporal variation in sedimentation rates in TFFW, and their controls, is a critical information gap.
The goals of this study were to (1) determine if nutrient sedimentation rates on intertidal floodplains differed along longitudinal gradients in tidal rivers; (2) evaluate if source(s) of deposited sediment change along longitudinal river gradients; (3) determine if there is a relationship between nutrient deposition and soil nutrient cycling, namely mineralization, in tidal river floodplains; and (4) compare short-term (deposition) vs. long-term (accumulation; following McKee et al. 1983) sediment and nutrient trapping rates. Addressing these goals will help identify the causes and consequences of TFFW ecosystem dynamics in response to climate and watershed change.
Methods
Study Sites
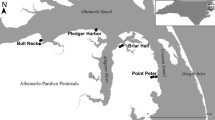
The longitudinal riverine transition from tidal freshwater forested to oligohaline marsh wetlands was studied along the floodplains of both the Waccamaw (SC, USA) and Savannah rivers (GA and SC, USA; Fig. 1). Four sites along each river ranged sequentially from continuously freshwater TFFW at the upstream end, moderately salt-impacted tidal forest (with moderate tree stress and mortality), highly salt-impacted tidal forest (with extensive tree mortality), and oligohaline marsh at the downstream end (see Krauss et al. 2009; Noe et al. 2013). The dominant tree species was Taxodium distichum [L.] L.C. Rich., with Nyssa aquatica L. codominant at continuously freshwater TFFW sites.

Map of sampling sites along the Savannah and Waccamaw River systems. The right panel shows the layout of contemporary (feldspar marker horizons) sediment deposition coring locations from this study (pink) and the earlier sampling reported in Ensign et al. (2014a) (white), and long-term sediment accumulation (210Pb core, yellow), using site W1 as an example
The Savannah River is a large alluvial (redwater) river that has a total watershed area of 27,390 km2 with a large portion in the Piedmont physiographic province. Discharge is affected by three dams upstream of the sites and constant dredging of the tidal portion of the river and estuary. Historically, the lower Savannah River was affected by a one-way tidal flap gate installed along one of three channel braids to the tidal portion of the Savannah River that caused wetland salinization (Pearlstine et al. 1993), but this gate caused only minor shifts in tidal swamp salinity (<1 ppt, Duberstein and Kitchens 2007). The Savannah River longitudinal transect includes a continuously freshwater tidal freshwater forest (soil porewater salinity of 0.1 ppt), a moderately salt-impacted forest (1.4 ppt), a highly salt-impacted forest (3.1 ppt), and an oligohaline marsh (3.5 ppt; Fig. 1).
The Waccamaw River, a predominantly blackwater river with a watershed area of 1981 km2 within the Coastal Plain physiographic province, is part of the Winyah Bay watershed (46,620 km2) that includes tributary inputs from the blackwater Sampit River (426 km2) and alluvial Pee Dee and Black rivers. The longitudinal river transect includes a continuously freshwater tidal forested wetland along the Waccamaw River (0.1 ppt), a moderately salt-impacted forest along the Waccamaw River (1.0 ppt), a highly salt-impacted forest along the lower Sampit River (1.9 ppt), and an oligohaline marsh along the lower Sampit River (3.2 ppt; Fig. 1). This longitudinal salinification gradient is hereafter referred to as the “Waccamaw River.”
Contemporary Deposition
Contemporary rates of sediment and nutrient deposition (g m−2 year−1) and vertical deposition (mm year−1) were measured by using artificial marker horizons. Marker horizons at each sites were placed on three transects oriented perpendicular to the river channel and separated by 50–100 m (Fig. 1). Transects ranged in length from 50 to 80 m, with five plots located equidistant along each transect, beginning 5 m from the channel. At each plot, a 2-cm-deep, 50-cm-wide square marker horizon was installed in September 2009 by using feldspar powder (Minspar 200, The Quartz Corp, Spruce Pine, NC). The clay becomes a firm, durable marker after it absorbs water, and allows accurate measurement of short-term net vertical deposition rates above the clay surface (Hupp and Bazemore 1993). Feldspar clay also accommodates fine root growth to facilitate sediment particle retention, as would occur naturally. Ensign et al. (2014a) reported rates of vertical deposition of sediment (mm year−1) from these marker horizons through February 2012.
In the current study, a subset of marker horizons in each site was sampled in March 2013, 3.5 years after installation. Five marker horizons at each of the moderately salt-impacted forest, highly salt-impacted forest, and oligohaline marsh sites were cored using a 7.4-cm-inner-diameter coring tube (three plots with associated soil mineralization measurement [see below; Noe et al. 2013] and two additional randomly chosen plots in each site). In the tidal freshwater forest sites, paired hummock and hollow marker horizons were sampled at three plots (six cores total per site). The core from each sampled marker horizon was extruded in the field, the depth of sediment above the feldspar was measured, and new sediment was separated from the feldspar marker horizon and returned to the laboratory (Noe and Hupp 2005). Sediment was dried at 60 °C until constant mass and weighed, identifiable roots and rhizomes >2 mm diameter were removed, and remaining (<2 mm) sediment was ground. Sediment was analyzed for loss-on-ignition (400 °C for 16 h, Nelson and Sommers 1996), particle size (LISST-100×, Wolf et al. 2011), total C and N (CHN elemental analyzer), and total P, Al, Ca, Fe, K, Mg, Na, and Ti (microwave-assisted acid digestion followed by ICP-OES, Noe et al. 2013). Sediment and nutrient deposition fluxes for each cored marker horizon were calculated by using the mass of new sediment, the diameter of the core tube, the duration of marker horizon deployment, and nutrient and organic/mineral content of sediment.
Long-Term Sedimentation
Deep soil cores for measuring long-term sediment and nutrient mass accumulation and vertical accumulation were collected at the Waccamaw River sites and analyzed for pollen, 14C, and radioisotope dating (Bernhardt, unpublished data), as well as nutrient analyses presented here. At the Waccamaw sites, a single 9.5-cm-diameter piston core was collected at the tidal freshwater forest, moderately salt-impacted tidal forest, and oligohaline marsh sites, whereas a single 7.0-cm-diameter vibracore was collected at the highly salt-impacted tidal forest site (due to greater density of deeper soil). Each core was collected to a depth of ~25 cm from a representative area at each site (within 50 m of a transect of feldspar marker horizons; Fig. 1). Cores were returned to the laboratory and sectioned into 1-cm (tidal forest sites) or 2-cm (oligohaline marsh) vertical increments. Soil increments were dried, weighed, ground, and then analyzed for 210Pb activity (Bernhardt, unpublished data) and total C, N, and P (same methods as for contemporary sedimentation, above) on separate subsamples. Cores also were collected at the Savannah River sites, but the coring technique that minimized compaction (Russian peat corer) did not provide sufficient material for radioisotope or nutrient elemental analysis following pollen analysis, and long-term sedimentation data from the Savannah River are not included here.
210Pb activity of each depth increment was determined directly by using alpha spectroscopy (after Flynn 1968) assuming 210Pb and its progeny are in secular equilibrium. Background 210Pb activity was first determined for each core by identifying the depth in the profile with constant 210Pb activity. Two cores had background 210Pb activity assigned to the lowest core increment, adding some uncertainty to 210Pb dating, but background 210Pb activity was uniform across all cores and less than 2 dpm g−1 (Supplemental Table 1). Values for missing increments were interpolated linearly from adjacent increments above and below the missing value. Missing 210Pb activity for the increment above the background layer in the core from the highly salt-impacted forest was interpolated from a natural log regression of unsupported 210Pb activity with depth. For missing 210Pb activity at the surface (tidal freshwater forest core), the average of unsupported 210Pb activity for all surficial increments down to 5 cm was used because of unvarying activity over this depth.
Unsupported 210Pb activity generally decreased exponentially with depth, but three of the four cores exhibited variable log-210Pb activity trends with depth indicating varying sedimentation rates over time. Therefore, the constant rate of supply (CRS) model (Appleby and Oldfield 1983) was used to calculate the age of sediment in each core increment as well as vertical accretion and sediment, C, N, and P mass accumulation rates in each increment (Corbett and Walsh 2015). Long-term mean rates of vertical accretion and sediment, C, N, and P accumulation in each core were calculated from the depth increments with unsupported 210Pb activity.
Soil N and P Mineralization
Surficial soil (0–5 cm) net N and P mineralization rates were measured in situ at these sites from 2009 to 2010 (Noe et al. 2013). Briefly, the mineralization method utilizes a modified resin core for measuring in situ rates of soil net inorganic N and P production over the duration of incubation (Noe 2011). Mineralization cores were measured quarterly over a year for 1–2 week-long incubations at three plots in each of the moderately salt-impacted forest, highly salt-impacted forest, and oligohaline marsh sites, with six plots in each of the tidal freshwater forest sites. Mineralization measurements were located about 1 m from an individual feldspar marker horizon that was cored during this study. Mean rates of soil N (NO3 − + NH4 +) and P (PO4 3−) mineralization in each plot are compared to measured deposition rates at the adjacent marker horizon from this study.
Statistical Analyses
Differences in sediment deposition rate between hummocks and hollows at the tidal freshwater forest sites were tested with one-way ANOVA. Differences in contemporary sedimentation rates and characteristics between rivers and longitudinally along rivers were tested using factorial two-way ANOVAs followed by Fisher’s LSD post hoc tests for significant model terms. Spatial variation in the elemental concentrations of contemporary deposited sediment was evaluated with principal components analysis (PCA). Correlation between contemporary deposition rates and soil N and P mineralization rates was tested with Spearman rank correlations.
Results
Contemporary Deposition (Marker Horizons)
Organic matter, C, and N concentrations in sediment deposited over 3.5 years on intertidal floodplains decreased downriver along both the Savannah and Waccamaw rivers, with a stronger trend on the Savannah River (Tables 1, 2). In other words, deposited sediment was more mineral with increasing salinity and conversion to oligohaline marsh. Along the Waccamaw River, deposition of moderately organic material at the highly salt-impacted forest, located at Turkey Creek along the Sampit River, disrupted the downriver trend. Downriver trends in N concentration were more similar along both rivers. Phosphorus concentration increased downriver along the Savannah River, but was relatively constant along the Waccamaw River. Mean organic matter content of recently deposited sediment ranged from 49 to 15 % among sites, indicating the predominance of mineral sediment deposition on tidal floodplains along these rivers. Deposited sediment was predominantly (>55 %) in the silt size fraction. The clay content of newly deposited sediment increased downriver with a corresponding decrease in silt content for both rivers.
The two rivers also differed in the composition of recently deposited sediment (Table 2), with more clay and Mg and less fine sand and N along the Savannah River compared to Waccamaw River (Table 1). Most of the other sediment composition measurements differed among the rivers depending on the site along the river (river by site interaction term). For example, organic, C, and P content differed between the Savannah and Waccamaw rivers depending on the longitudinal position (site) along each river. However, silt, K, and Na content did not differ among the rivers irrespective of longitudinal site (no interaction).
The elemental composition of deposited sediment revealed spatial trends in sediment characteristics between and especially along the two rivers. PCA component 1 represents a strong longitudinal spatial gradient in metal content along the rivers, explaining 59 % of variation (Fig. 2). The concentrations of Ti, Fe, Mg, P, and Al were positively loaded, and C, N, Ca, Na, and K were negatively loaded on component 1. Sites sorted along component 1, with a gradient from tidal freshwater forest (negative loading) to salt-impacted tidal forests and oligohaline marsh (positive loading, Fig. 3). Within each longitudinal riverine position, sites along the Savannah River always loaded more positively to component 1 than the corresponding site along the Waccamaw River. Component 2 (20 % of variation) comprised a spatial gradient in organic content with both C and N negatively loaded and K, Na, and Al concentrations positively loaded (Fig. 2). Variation in component 2 scores was high among rivers and sites and represents within-site variation in organic content among plots.

Principal components analysis of elemental concentrations in contemporary deposited sediment

Contemporary sediment deposition and vertical deposition rates (site average ± 1 s.e.) vs. site score along the first principal component (PCA1) of sediment elemental concentrations. Savannah River = open symbols, Waccamaw River = gray symbols
Rates of sediment deposition (g m−2 year−1) did not differ between hummocks and hollows at the tidal freshwater forest sites (P = 0.449, ANOVA, n = 6); thus, deposition rates were pooled for comparing the tidal freshwater forest sites to other sites. Sediment deposition rates differed greatly across the study sites (Fig. 4), with significant trends longitudinally along rivers and between rivers, and their interaction term, for total, mineral, P, and N deposition rates (Table 2). Sediment deposition rates of organic material and C did not differ along or between rivers, but their interaction term was significant, indicating site-specific variation. Sediment deposition rates were greater on the alluvial Savannah River compared to the more blackwater Waccamaw River (Fig. 4). All sediment deposition rates generally increased downriver, with the lowest rates in the tidal freshwater forests and increasing rates along the salinity gradient into oligohaline marsh. However, the highly salt-impacted forests along both rivers disrupted the downriver trend, with the highest (Savannah) and lowest (Waccamaw) mean rates measured anywhere in this study. On average, deposited sediment was 79 % mineral and 21 % organic.

Contemporary sediment and nutrient deposition rates (site average ± 1 s.e.) along the longitudinal (upriver-downriver) profile of tidal rivers
The average contemporary vertical deposition rate of all plots in this study (2009–2013, 9.0 mm year−1) was similar to rates measured earlier at the same plots (2009–2012, 9.2 mm year−1; subset of plots in Ensign et al. 2014a). However, the comparison of rates in individual plots measured in 2012 and remeasured in 2013 reveals fine-scale variability in rates over time (Fig. 5). In particular, average vertical deposition rates increased through time at the Savannah River plots (10.5 mm year−1 in 2012 and 11.8 mm year−1 in 2013) and decreased on average at the Waccamaw River plots (7.8 mm year−1 in 2012 and 6.1 mm year−1 in 2013). Watershed discharge and tidal river stage variations over time for the two rivers show no differences that could account for the differing changes in deposition rates between the two time periods (Savannah River near Clyo, USGS gage 02198840; Pee Dee River near Bucksport, USGS gage 02135200; Waccamaw River at Conway Marina, USGS gage 02110704; Savannah River at I-95, USGS gage 02198840; Waccamaw River near Pawleys Island, USGS gag 021108125). Most (78 %) of remeasured plots had increased depth (mm) of deposited sediment in 2013 compared to 2012, indicating net deposition, whereas 22 % experienced some erosion.

Vertical deposition rates over individual marker horizons installed in 2009 and measured in 2012 (subset of plots in Ensign et al. 2014a) and 2013 (this study). Savannah River = open symbols, Waccamaw River = gray symbols
The elemental composition of deposited sediment related to the mass and vertical deposition rates (Fig. 3). Site mean sediment deposition rate increased with site loading along PCA component 1, irrespective of the river. Downriver oligohaline marsh sites, nearest the estuary, had the highest sediment deposition rates and most positive loading along component 1 (Fig. 3) due to their elevated concentrations of Ti, Fe, Mg, P, and Al (Table 1, Fig. 2). The same pattern exists for vertical deposition rates (Fig. 3) due to relatively small differences in sediment bulk density among sites (Table 1).
Long-Term Accumulation (210Pb Cores)
Mean long-term mass accumulation (g m−2 year−1) and vertical accretion (mm year−1) rates, measured by using 210Pb CRS dating of soil core increments, generally decreased from tidal freshwater forest downriver to oligohaline marsh along the Waccamaw River (Table 3). The Waccamaw downriver decrease in long-term mean accumulation was most clear for sediment and P accumulation and vertical accretion, with the greatest rates at the tidal freshwater forest, intermediate rates at the two salt-impacted forests, and generally the lowest rates at oligohaline marsh (Table 3). In contrast, long-term C and N accumulation rates peaked at the highly salt-impacted tidal forest, were intermediate at tidal freshwater and moderately salt-stressed forest, and were lowest at oligohaline marsh along the Waccamaw River. Thus, mineral sediment accumulation decreased nearly monotonically from freshwater to oligohaline wetlands, whereas organic accumulation was more consistent among the freshwater and moderately and highly salt-impacted tidal forests and lowest in oligohaline marsh of the Waccamaw River. This downriver pattern of less long-term sediment accumulation is the opposite of the downriver pattern of greater contemporary sediment deposition measured with artificial marker horizons (Table 3).
Accumulation rates decreased in deeper, older increments of the cores, although not monotonically (Fig. 6). Maximum datable ages of the cores ranged from 90 to 169 years (Supplemental Table 1). The oldest ages for given depths were found in the oligohaline marsh core (Fig. 6), corresponding to the lowest vertical accretion rates (Supplemental Table 1). More modern increments (<20 years) had much greater accumulation rates than older increments. This trend is strongest for sediment and P accumulation and vertical accretion in the core from tidal freshwater forest (W1, Fig. 6).

Downcore profiles of 210Pb-derived sediment ages (CRS model; top left panel) and calculated rates of sediment and nutrient accumulation over time along the Waccamaw River. W1 = tidal freshwater forest, W2 = moderately salt-impacted tidal forest, W3 = highly salt-impacted forest, W4 = oligohaline marsh
Long-term sediment and nutrient accumulation rates (average of all datable increments) were much lower than contemporary sediment deposition rates measured by using marker horizons in the same site (Table 3). Mean sediment deposition rate among the Waccamaw River sites was 1657 g m−2 year−1 over the 3.5 years of marker horizon measurements compared to 224 g m−2 year−1 of accumulation over the 100- to 150-year duration of 210Pb dating (7× lower). Carbon, N, and P accumulation and vertical accretion rates on average were 4×, 3×, 9×, and 2× lower, respectively, using 210Pb dating compared to deposition determined from marker horizons. As previously mentioned, the rank order of rates among the four sites was nearly the opposite for contemporary and long-term sedimentation. Even when considering only the more recently accumulated soil increments near the top of the 210Pb dated cores (Fig. 6), contemporary rates of deposition exceeded long-term accumulation of sediment.
Relationship Between Short-Term Sediment Inputs and Soil Nutrient Mineralization
Soil net N and P mineralization rates, measured in situ in 2009–2010 (Noe et al. 2013), were compared to contemporary deposition inputs (2009–2013) measured at the same plots. For an individual element, the only significant correlation between soil mineralization flux and depositional flux was for P (r s = 0.529, P = 0.010, Spearman rank correlation, n = 23). Nitrogen mineralization, ammonification, and nitrification were unrelated to N or C deposition (r s < 0.316, P > 0.142) or any other depositional flux. In addition to P deposition rate, soil P mineralization also was correlated with vertical deposition rate, sediment deposition rate, and mineral sediment deposition rate (r s > 0.431, P < 0.040), but not N (r s = 0.404, P = 0.056) or C deposition rates (r s = 0.230, P = 0.291).
Discussion
The large extent and watershed position of tidal wetlands suggest their potential to have large influence on nutrient and sediment delivery to estuaries. For example, N sedimentation in tidal marshes was up to 32 % of watershed N load (Loomis and Craft 2010). Nutrient trapping by sedimentation in TFFW might also provide a large landscape nutrient removal mechanism from tidal waters along these rivers because the TFFW area is often extensive. Although TFFW have long-term soil N and P accumulation rates that are lower than those of downstream tidal marshes (Craft 2012; this study, see below), these fluxes could extrapolate to be quantitatively important to estuarine loading. TFFW also have high rates of nitrate removal by denitrification (Ensign et al. 2008, 2013), further increasing their role in regulating coastal N availability. However, the upstream limit of tidal freshwater forested wetlands is unclear; delineation of tidal inundation is difficult using remote sensing imagery. Nonetheless, TFFW are likely more extensive than tidal freshwater marsh in the estuaries of the USA (Field et al. 1991). In addition to the large influence of TFFW and other intertidal wetlands on influencing N and P loading to estuaries and the coastal zone, non-tidal floodplains of the Coastal Plain also have large trapping rates that can be a large proportion of watershed sediment, N, and P loads (Noe and Hupp 2009; Hupp et al. 2015). Thus, lowland floodplain wetlands cumulatively remove large quantities of pollutants from rivers that would otherwise impact estuarine ecosystems.
Sedimentation studies in tidal wetlands have shown that contemporary deposition rates are greater than long-term accumulation rates in the same sites (Neubauer et al. 2002; Craft 2007; Breithaupt et al. 2014; Hupp et al. 2015). The sites along the Waccamaw River showed 7× greater contemporary sedimentation rates (3.5-year duration marker horizons) compared to rates over the past ca. 150 years (210Pb dating). Compaction and decomposition of deposited material and temporal variation in sedimentation and erosion all contribute to this common discrepancy (Cahoon 2014). Short-term patterns of sedimentation are also more variable over time (Delgado et al. 2013) and space (Breithaupt et al. 2014) than long-term patterns due to rare, large sedimentation events (Sadler 1981). Average contemporary (2009–2013) vertical accretion rates of all plots in this study were similar to average rates measured earlier at the same plots (2009–2012; subset of plots in Ensign et al. 2014a). However, comparison of changes in rates of individual plots measured in 2012 and 2013 reveals considerable plot-scale spatial variability, as well as changes in rates between the two rivers. The increase in deposition rate on the Savannah River compared to the decrease on the Waccamaw River over the period from 2012 to 2013 could not be explained by differences in watershed discharge or tidal river stage excursions. In summary, temporal variability in vertical accretion in our study was dependent on spatial scale, from plot to river to region, without obvious explanation. In addition to this scale-dependent variation in spatial and temporal patterns of sedimentation under typical conditions, tropical cyclones can result in very large amounts of episodic deposition in intertidal forested wetlands (Castañeda-Moya et al. 2010; Ensign et al. 2014b).
The large discrepancy in sediment mass trapping rate over time is likely reflective of changes in sediment availability over time. Mineral sediment comprised 79 % of total sedimentation in this study, but is unlikely to have meaningful mass loss over time due to mineral dissolution. Similarly, compaction would not change the mass of sediment in storage. Further, widespread, large rates erosion of floodplain sediment over long time periods is not likely in these low-energy environments, with only 22 % of marker horizons experiencing erosion over a 1-year period but none eroding over 3.5 years. Instead, we speculate that the reversal of upriver-downriver trends in the contemporary vs. long-term sedimentation rates is due to long-term changes in the position of the ETM in the Waccamaw River complex. Historically, sedimentation rates were greatest in the TFFW. This may reflect the much larger watershed loads of sediment in the past associated with the legacy of soil erosion associated with historic land-use change (Meade 1969; Hupp et al. 2009, 2015). In the modern era, sedimentation rates were greatest in the oligohaline marsh. This may reflect an upriver transgression of saline and relatively sediment-rich waters into formerly TFFW that had converted into oligohaline marsh, resulting in an increase in sediment availability and trapping. Alternatively, dredging for shipping downstream in the Waccamaw estuary may have increased contemporary sediment availability to wetlands nearest the estuary.
Long-term accumulation rates of sediment, N, P, and C in TFFW were much lower in this study compared to those of other rivers in the southeastern USA (Craft 2012). Collectively, these sediment and associated nutrient accumulation rates in TFFW are lower than rates in tidal freshwater marshes (Neubauer et al. 2005; Megonigal and Neubauer 2009; Loomis and Craft 2012; Drexler et al. 2013). Tidal freshwater marshes can have similar long-term N sediment accumulation rates as brackish marshes, but greater than salt marshes, and lower P accumulation rates than brackish marshes (Craft 2007; Loomis and Craft 2010). In contrast, oligohaline marsh in this study had much lower long-term N accumulation and somewhat lower P accumulation than TFFW. In summary, long-term material trapping by wetlands varies along tidal gradients from freshwater to seawater, with TFFW having among the lowest rates.
As found in this study, others have shown that contemporary sediment and associated nutrient deposition is greater in brackish than tidal freshwater wetlands, due to proximity of the ETM, tidal channel hydraulics, or vegetative roughness (Darke and Megonigal 2003; Morse et al. 2004; Ensign et al. 2014a, b). No difference in sediment deposition was observed between tidal freshwater marsh and salt marsh, however (Temmerman et al. 2003), suggesting greatest contemporary deposition rates in estuaries likely occur in oligohaline marsh. Within the TFFW zone, sediment accretion has been shown to decrease from the upper reaches of tidal freshwater rivers (near the limit of tide) to the downriver limit of TFFW (near oligohaline wetlands) (Baldwin 2007; Ensign et al. 2014b). These longitudinal gradients in contemporary deposition suggest greater sedimentation in the upper reaches of TFFW due to greater availability of watershed-derived fluvial sediment, minimization of sedimentation in the lower reaches of TFFW after exhaustion of watershed sediment load by sediment removal in the upper reaches, and an increase in sedimentation in oligohaline marshes due to the ETM increasing the availability of estuarine or marine sediment.
The characteristics of deposited sediment suggest a change in sediment sources along tidal rivers. Sediment deposited on the tidal floodplains of the Waccamaw and Savannah rivers was enriched in clay at sites closer to the estuary, and also were enriched in Ti, Fe, Mg, P, and Al but depleted in K, Na, Ca, C, and N. Clay concentration and mineralogy change along the lengths of estuarine rivers due to differing watershed (Piedmont vs. Coastal Plain) and marine sources (Windom et al. 1971). Clay (coastal sediment) and quartz sand (Coastal Plain watershed sediment) are likely end-members of sediment sources in estuaries with strongly differing metal content (Hanson et al. 1993). In addition to differing mineralogical end-members from watershed and estuarine sources, metal concentrations in suspended sediment also vary along estuarine salinity gradients. Fe concentrations in suspended particles increase with salinity in estuaries (Windom 1976), which can also increase the concentrations of other metals (Bianchi 2007). Similarly, wetland soil Mg concentrations increased along a freshwater to oligohaline to mesohaline gradient in a Florida estuary (Williams et al. 2014).
These trends in the elemental geochemistry of deposited sediment, and inferred change in sediment source, covaried with sediment deposition rate along the longitudinal tidal freshwater to oligohaline wetland gradient of the tidal rivers in this study. The greater deposition rates in salt-impacted tidal forests and oligohaline marshes were associated with a shift in sediment geochemistry that suggests a change in sediment source from trapping of watershed-derived sediment in TFFW to coastal-derived sediment in salt-stressed forests and oligohaline marshes. The observed changes in the rate of deposition and the chemistry of deposited sediment suggest greater sediment trapping in oligohaline marshes is due to an increase in sediment availability from a coastal sediment source. This enhanced availability of sediment to tidal freshwater and oligohaline marshes along lower tidal rivers is often assumed to be associated with the ETM (Darke and Megonigal 2003; Loomis and Craft 2010). Flood-tide dominant transport also can move upstream fine sediment from the coast, estuarine sediment mobilized by dredging, and entrained channel bed sediment in tidal rivers (Meade 1969), leading to enhanced deposition in oligohaline marshes (Ensign et al. 2014b). Similarly, oceanic and coastal sediment can be transported to and deposited in estuaries (Newell et al. 2004). The exception to this longitudinal trend in the two rivers was the Waccamaw highly salt-impacted tidal forest that had the lowest sediment deposition rate along the Waccamaw River system. This exception is likely due to the landscape position of this site along a small tributary of the Sampit River with very small catchment area and presumably low watershed sediment loading as well as little ETM sediment availability—the chemistry of the small amount of sediment deposited at this location suggests a watershed source. Collectively, recent research is identifying a minimum of sediment trapping in lower TFFW, associated with less availability of both continental and coastal sediment, that fits more generic models of sediment dynamics in estuaries (e.g., Biggs and Howell 1984).
Relationship Between Sediment Inputs and Soil Nutrient Mineralization
Phosphorus inputs from sediment deposition increased soil P mineralization in these TFFW and oligohaline marshes. Noe et al. (2013) identified a significant correlation between spatiotemporal variation in soil P mineralization turnover rate (day−1) and soil conductivity at the same sites. However, spatial pattern in contemporary P deposition rate is more strongly correlated with P mineralization rate (g-P m−2 year−1, r s = 0.529) and P turnover rate (r s = 0.248, P = 0.254) than is soil conductivity with P mineralization rate (r s = −0.044, P = 0.840) or P turnover rate (r s = 0.243, P = 0.265) at the subset of plots where both deposition and mineralization were measured. In summary, greater inputs of P from sediment deposition, and not greater salinity, is the best explanation for enhanced soil P mineralization rates in oligohaline marshes along the Savannah and Waccamaw rivers.
Enhanced input of nutrients to wetlands has been shown to stimulate internal wetland biogeochemical cycling (Hopkinson 1992; Wolf et al. 2013). The relationship between sediment deposition and soil mineralization of P, but lack of a relationship for N, suggests that P mineralization is limited by the rate of P inputs to the TFFW and oligohaline marsh ecosystems. The linkage of P deposition and mineralization suggests that the deposited particles have labile forms of P. The lability of deposited sediment P could be due to either direct microbial mineralization of organic P, desorption of P associated with reducing conditions, or addition of salt to soils (Noe et al. 2013). In contrast, N mineralization is decoupled from N inputs. Instead, N mineralization along tidal rivers is influenced by the production of large quantities of labile litter from dying trees as salinification converts TFFW into oligohaline marsh (Noe et al. 2013). The linkage between P inputs from coastally derived sediments and internal ecosystem biogeochemical processes in TFFW and oligohaline marsh is similar to some mangrove ecosystems where inputs of P from marine sediment regulate ecosystem productivity (Castañeda-Moya et al. 2010).
Conclusions
The changing chemical signature of deposited sediment firmly links longitudinal changes in sediment trapping by wetlands along tidal rivers and estuaries to a changing source from watershed to coastal sediment. This change in sediment source is associated with sediment availability and explains the sedimentation minimum that is often observed in TFFW compared to oligohaline marshes. The minimal sediment trapping and accretion in TFFW (Craft 2012; this study) suggest that they are most sensitive to current and future sea level rise impacts, whereas the resiliency of oligohaline marshes would be greater due to enhanced sediment availability from coastal sources (Kirwan et al. 2010). However, longitudinal patterns of sedimentation can change depending on the time frame of measurement (Hupp et al. 2015), with opposing longitudinal sedimentation patterns from contemporary deposition vs. long-term accumulation measurements. These findings have important implications for the linkages among watershed sediment loading, wetland resilience to sea level rise, and estuarine nutrient budgets.
References
Anderson, C.J., and B.G. Lockaby. 2007. Soils and biogeochemistry of tidal freshwater forested wetlands. In Ecology of tidal freshwater forested wetlands of the southeastern United States, ed. W.H. Conner, T.W. Doyle, and K.W. Krauss, 65–88. Dordrecht: Springer.
Appleby, P.G., and F. Oldfield. 1983. The assessment of 210Pb data from sites with varying sediment accumulation rates. Hydrobiologia 103: 29–35.
Ardón, M., J.L. Morse, B.P. Colman, and E.S. Bernhardt. 2013. Drought-induced saltwater incursion leads to increased wetland nitrogen export. Global Change Biology 19: 2976–2985.
Baldwin, A.H. 2007. Vegetation and seed bank studies of salt-pulsed swamps of the Nanticoke River, Chesapeake Bay. In Ecology of tidal freshwater forested wetlands of the southeastern United States, ed. W.H. Conner, T.W. Doyle, and K.W. Krauss, 139–160. Dorcrecht: Springer.
Bianchi, T.S. 2007. Biogeochemistry of estuaries. New York: Oxford University Press.
Biggs, R.B., and B.A. Howell. 1984. The estuary as a sediment trap; alternate approaches to estimating its filtering efficiency. In The estuary as a filter, ed. V.S. Kennedy, 107–129. New York: Academic.
Breithaupt, J.L., J.M. Smoak, T.J. Smith, and C.J. Sanders. 2014. Temporal variability of carbon and nutrient burial, sediment accretion, and mass accumulation over the past century in a carbonate platform mangrove forest of the Florida Everglades. Journal of Geophysical Research, Biogeosciences 119: 2032–2048.
Cahoon, D. 2014. Estimating relative sea-level rise and submergence potential at a coastal wetland. Estuaries and Coasts. doi:10.1007/s12237-014-9872-8.
Castañeda-Moya, E., R. Twilley, V. Rivera-Monroy, K. Zhang, S. Davis III, and M. Ross. 2010. Sediment and nutrient deposition associated with Hurricane Wilma in mangroves of the Florida Coastal Everglades. Estuaries and Coasts 33: 45–58.
Conner, W.H., T.W. Doyle, and K.W. Krauss (eds.). 2007. Ecology of tidal freshwater forested wetlands of the southeastern United States. Dorcrecht: Springer.
Corbett, D.R., and J.P. Walsh. 2015. 210lead and 137cesium. In Handbook of sea-level research, ed. I. Shennan, A.J. Long, and B.P. Horton, 361–372. Chichester: Wiley.
Craft, C. 2007. Freshwater input structures soil properties, vertical accretion, and nutrient accumulation of Georgia and U.S. tidal marshes. Limnology and Oceanography 2007: 1220–1230.
Craft, C. 2012. Tidal freshwater forest accretion does not keep pace with sea level rise. Global Change Biology 18: 3615–3623.
Darke, A.K., and J.P. Megonigal. 2003. Control of sediment deposition rates in two mid-Atlantic coast tidal freshwater wetlands. Estuarine, Coastal and Shelf Science 57: 255–268.
Day, R.H., T.M. Williams, and C.M. Swarzenski. 2007. Hydrology of tidal freshwater forested wetlands of the southeastern United States. In Ecology of tidal freshwater forested wetlands of the southeastern United States, ed. W.H. Conner, T.W. Doyle, and K.W. Krauss, 29–63. Dorcrecht: Springer.
Delgado, P., P.F. Hensel, C.W. Swarth, M. Ceroni, and R. Boumans. 2013. Sustainability of a tidal freshwater marsh exposed to a long-term hydrologic barrier and sea level rise. Estuaries and Coasts 36: 585–594.
Drexler, J.Z., K.W. Krauss, M.C. Sasser, C.C. Fuller, C.M. Swarzenski, A. Powell, K.M. Swanson, and J. Orlando. 2013. A long-term comparison of carbon sequestration rates in impounded and naturally tidal freshwater marshes along the lower Waccamaw River, South Carolina. Wetlands 33: 965–974.
Duberstein, J., and W. Kitchens. 2007. Community composition of select areas of tidal freshwater forest along the Savannah River. In Ecology of tidal freshwater forested wetlands of the southeastern United States, ed. W.H. Conner, T.W. Doyle, and K.W. Krauss, 321–348. Dorcrecht: Springer.
Ensign, S., M. Piehler, and M. Doyle. 2008. Riparian zone denitrification affects nitrogen flux through a tidal freshwater river. Biogeochemistry 91: 133–150.
Ensign, S., K. Siporin, M. Piehler, M. Doyle, and L. Leonard. 2013. Hydrologic versus biogeochemical controls of denitrification in tidal freshwater wetlands. Estuaries and Coasts 36: 519–532.
Ensign, S.H., C.R. Hupp, G.B. Noe, K.W. Krauss, and C.L. Stagg. 2014a. Sediment accretion in tidal freshwater forests and oligohaline marshes of the Waccamaw and Savannah rivers, USA. Estuaries and Coasts 37: 1107–1119.
Ensign, S.H., G.B. Noe, and C.R. Hupp. 2014b. Linking channel hydrology with riparian wetland accretion in tidal rivers. Journal of Geophysical Research, Earth Surface 119: 28–44.
Field, D.W., A.J. Reyer, P.V. Genovese, and B.D. Shearer. 1991. Coastal wetlands of the United States, an accounting of a valuable national resource. Rockville: National Oceanic and Atmospheric Administration.
Flynn, W.W. 1968. The determination of low levels of polonium-210 in environmental materials. Analytica Chimica Acta 43: 221–227.
Gao, H., J. Bai, X. He, Q. Zhao, Q. Lu, and J. Wang. 2014. High temperature and salinity enhance soil nitrogen mineralization in a tidal freshwater marsh. PLoS ONE 9, e95011.
Hanson, P.J., D.W. Evans, D.R. Colby, and V.S. Zdanowicz. 1993. Assessment of elemental contamination in estuarine and coastal environments based on geochemical and statistical modeling of sediments. Marine Environmental Research 36: 237–266.
Hopkinson, C.S. 1992. A comparison of ecosystem dynamics in freshwater wetlands. Estuaries and Coasts 15: 549–562.
Hupp, C.R., and D.E. Bazemore. 1993. Temporal and spatial patterns of wetland sedimentation, West Tennessee. Journal of Hydrology 141: 179–196.
Hupp, C.R., A.R. Pierce, and G.B. Noe. 2009. Floodplain geomorphic processes and environmental impacts of human alteration along coastal plain rivers, USA. Wetlands 29: 413–429.
Hupp, C.R., E.R. Schenk, D.E. Kroes, D.A. Willard, P.A. Townsend, and R.K. Peet. 2015. Patterns of floodplain sediment deposition along the regulated lower Roanoke River, North Carolina: annual, decadal, centennial scales. Geomorphology 228: 666–680.
Johnston, C.A. 1991. Sediment and nutrient retention by freshwater wetlands: effects on surface water quality. Critical Reviews in Environmental Control 21: 491–565.
Jun, M., A.E. Altor, and C.B. Craft. 2013. Effects of increased salinity and inundation on inorganic nitrogen exchange and phosphorus sorption by tidal freshwater floodplain forest soils, Georgia (USA). Estuaries and Coasts 36: 508–518.
Kirwan, M.L., G.R. Guntenspergen, A. D’Alpaos, J.T. Morris, S.M. Mudd, and S. Temmerman. 2010. Limits on the adaptability of coastal marshes to rising sea level. Geophysical Research Letters 37, L23401.
Krauss, K., J. Duberstein, T. Doyle, W. Conner, R. Day, L. Inabinette, and J. Whitbeck. 2009. Site condition, structure, and growth of bald cypress along tidal/non-tidal salinity gradients. Wetlands 29: 505–519.
Kroes, D., C. Hupp, and G. Noe. 2007. Sediment, nutrient, and vegetation trends along the tidal, forested Pocomoke River, Maryland. In Ecology of tidal freshwater forested wetlands of the southeastern United States, ed. W.H. Conner, T.W. Doyle, and K.W. Krauss, 113–137. Dorcrecht: Springer.
Loomis, M.J., and C.B. Craft. 2010. Carbon sequestration and nutrient (nitrogen, phosphorus) accumulation in river-dominated tidal marshes, Georgia, USA. Soil Science Society of America Journal 74: 1028–1036.
McKee, B.A., C.A. Nittrouer, and D.J. DeMaster. 1983. Concepts of sediment deposition and accumulation applied to the continental shelf near the mouth of the Yangtze River. Geology 11: 631–633.
Meade, R.H. 1969. Landward transport of bottom sediments in estuaries of the Atlantic coastal plain. Journal of Sedimentary Research 39: 222–234.
Megonigal, J.P., and S.C. Neubauer. 2009. Biogeochemistry of tidal freshwater wetlands. In Coastal wetlands: an integrated ecosystem approach, ed. G.M.E. Perillo, E. Wolanski, D.R. Cahoon, and M.M. Brinson, 535–562. Amsterdam: Elsevier.
Morse, J.L., J.P. Megonigal, and M.R. Walbridge. 2004. Sediment nutrient accumulation and nutrient availability in two tidal freshwater marshes along the Mattaponi River, Virginia, USA. Biogeochemistry 69: 175–206.
Nelson, D.W., and L.E. Sommers. 1996. Total carbon, organic carbon, and organic matter. In Methods of soil analysis, part 2, microbiological and biochemical properties, ed. A.L. Page, 961–1010. Madison: ASA and SSSA.
Neubauer, S.C., I.C. Anderson, J.A. Constantine, and S.A. Kuehl. 2002. Sediment deposition and accretion in a mid-Atlantic (USA) tidal freshwater marsh. Estuarine, Coastal and Shelf Science 54: 713–727.
Neubauer, S., I. Anderson, and B. Neikirk. 2005. Nitrogen cycling and ecosystem exchanges in a Virginia tidal freshwater marsh. Estuaries and Coasts 28: 909–922.
Newell, W.L., I. Clark, and O. Bricker. 2004. Distribution of Holocene sediment in Chesapeake Bay as interpreted from submarine geomorphology of the submerged landforms, selected core holes, bridge borings and seismic profiles. Open File Report 2004-1235. Reston: U.S. Geological Survey.
Noe, G.B. 2011. Measurement of net nitrogen and phosphorus mineralization in wetland soils using a modification of the resin-core technique. Soil Science Society of America Journal 75: 760–770.
Noe, G.B., and C.R. Hupp. 2005. Carbon, nitrogen, and phosphorus accumulation in floodplains of Atlantic Coastal Plain rivers, USA. Ecological Applications 15: 1178–1190.
Noe, G.B., and C.R. Hupp. 2009. Retention of riverine sediment and nutrient loads by coastal plain floodplains. Ecosystems 12: 728–746.
Noe, G.B., K.W. Krauss, B.G. Lockaby, W.H. Conner, and C.R. Hupp. 2013. The effect of increasing salinity and forest mortality on soil nitrogen and phosphorus mineralization in tidal freshwater forested wetlands. Biogeochemistry 114: 225–244.
Odum, W.E. 1988. Comparative ecology of tidal freshwater and salt marshes. Annual Review of Ecology and Systematics 19: 147–176.
Pearlstine, L.G., W.M. Kitchens, P.J. Latham, and R.D. Bartleson. 1993. Tide gate influences on a tidal marsh. Journal of the American Water Resources Association 29: 1009–1019.
Sadler, P.M. 1981. Sediment accumulation rates and the completeness of stratigraphic sections. The Journal of Geology 89: 569–584.
Temmerman, S., G. Govers, S. Wartel, and P. Meire. 2003. Spatial and temporal factors controlling short-term sedimentation in a salt and freshwater tidal marsh, Scheldt estuary, Belgium, SW Netherlands. Earth Surface Processes and Landforms 28: 739–755.
Weston, N.B., R.E. Dixon, and S.B. Joye. 2006. Ramifications of increased salinity in tidal freshwater sediments: geochemistry and microbial pathways of organic matter mineralization. Journal of Geophysical Research, Biogeosciences 111, G01009.
Williams, A.A., N.T. Lauer, and C.T. Hackney. 2014. Soil phosphorus dynamics and saltwater intrusion in a Florida estuary. Wetlands 34: 535–544.
Windom, H.L. 1976. Geochemical interactions of heavy metals in southeastern salt marsh environments. U.S. EPA-600/3-76-023. Corvallis: Environmental Protection Agency.
Windom, H.L., W.J. Neal, and K.C. Beck. 1971. Mineralogy of sediments in three Georgia estuaries. Journal of Sedimentary Research 41: 497–504.
Wolf, K., C. Ahn, and G. Noe. 2011. Development of soil properties and nitrogen cycling in created wetlands. Wetlands 31: 699–712.
Wolf, K.L., G.B. Noe, and C. Ahn. 2013. Hydrologic connectivity to streams increases nitrogen and phosphorus inputs and cycling in soils of created and natural floodplain wetlands. Journal of Environmental Quality 42: 1245–1255.
Acknowledgments
We thank Jackie Batson, Ed Schenk, Nick Ostroski, Andrew Kunz, Scott Ensign, Kristin Wolf, Sara Ulrich, and Marci Marot for field and laboratory assistance. Support was provided by the U.S. Geological Survey Climate and Land Use Change Research and Development Program and the U.S. Geological Survey National Research Program. Comments from reviewers were particularly helpful for improving the manuscript. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US Government.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by David Reide Corbett
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 56 kb)
Rights and permissions
About this article

Cite this article
Noe, G.B., Hupp, C.R., Bernhardt, C.E. et al. Contemporary Deposition and Long-Term Accumulation of Sediment and Nutrients by Tidal Freshwater Forested Wetlands Impacted by Sea Level Rise. Estuaries and Coasts 39, 1006–1019 (2016). https://doi.org/10.1007/s12237-016-0066-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12237-016-0066-4